Pulmonary Carcinoid Surface Receptor Modulation Using Histone Deacetylase Inhibitors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
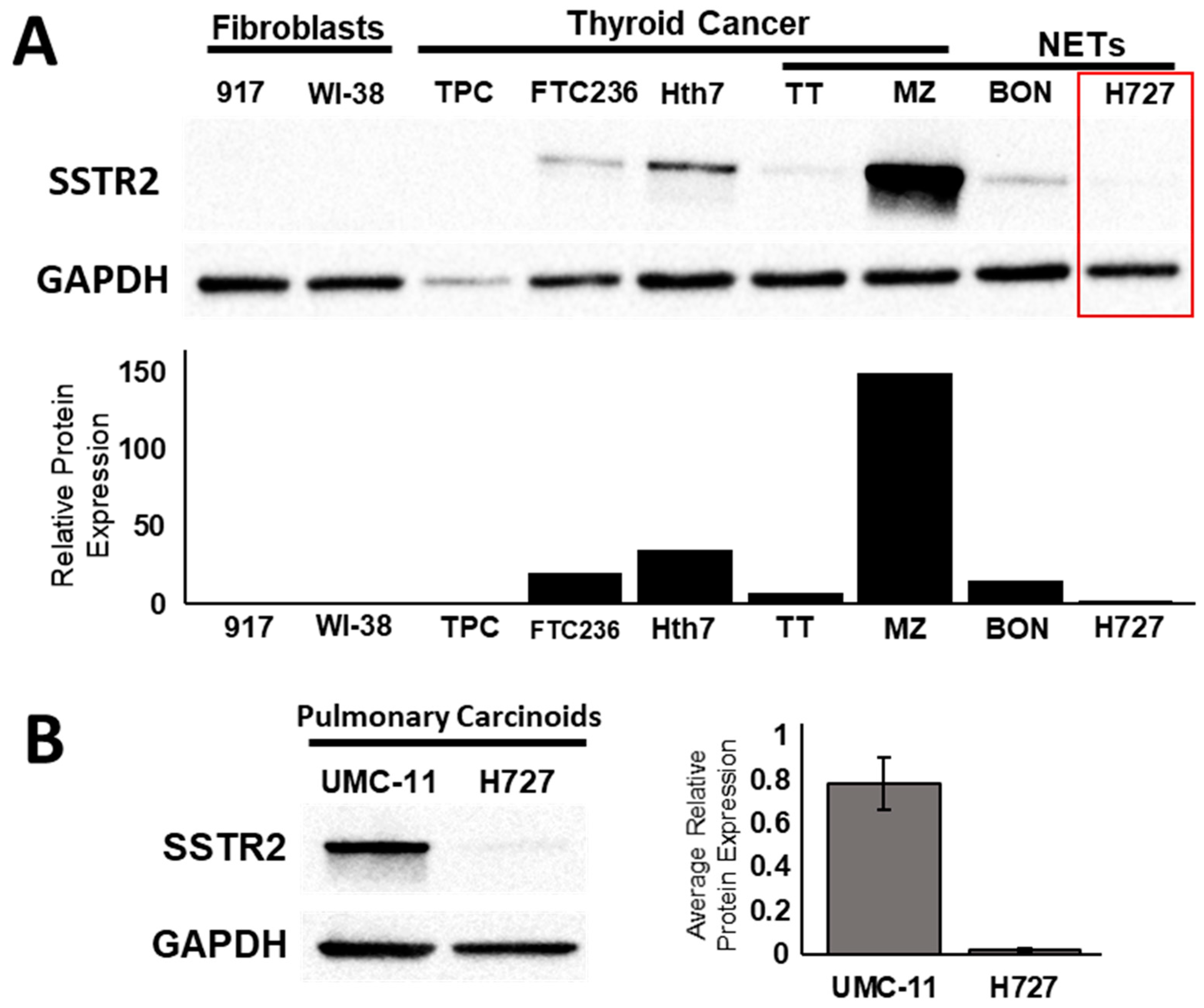
2.1. Transcriptional and Translational Induction of SSTR2
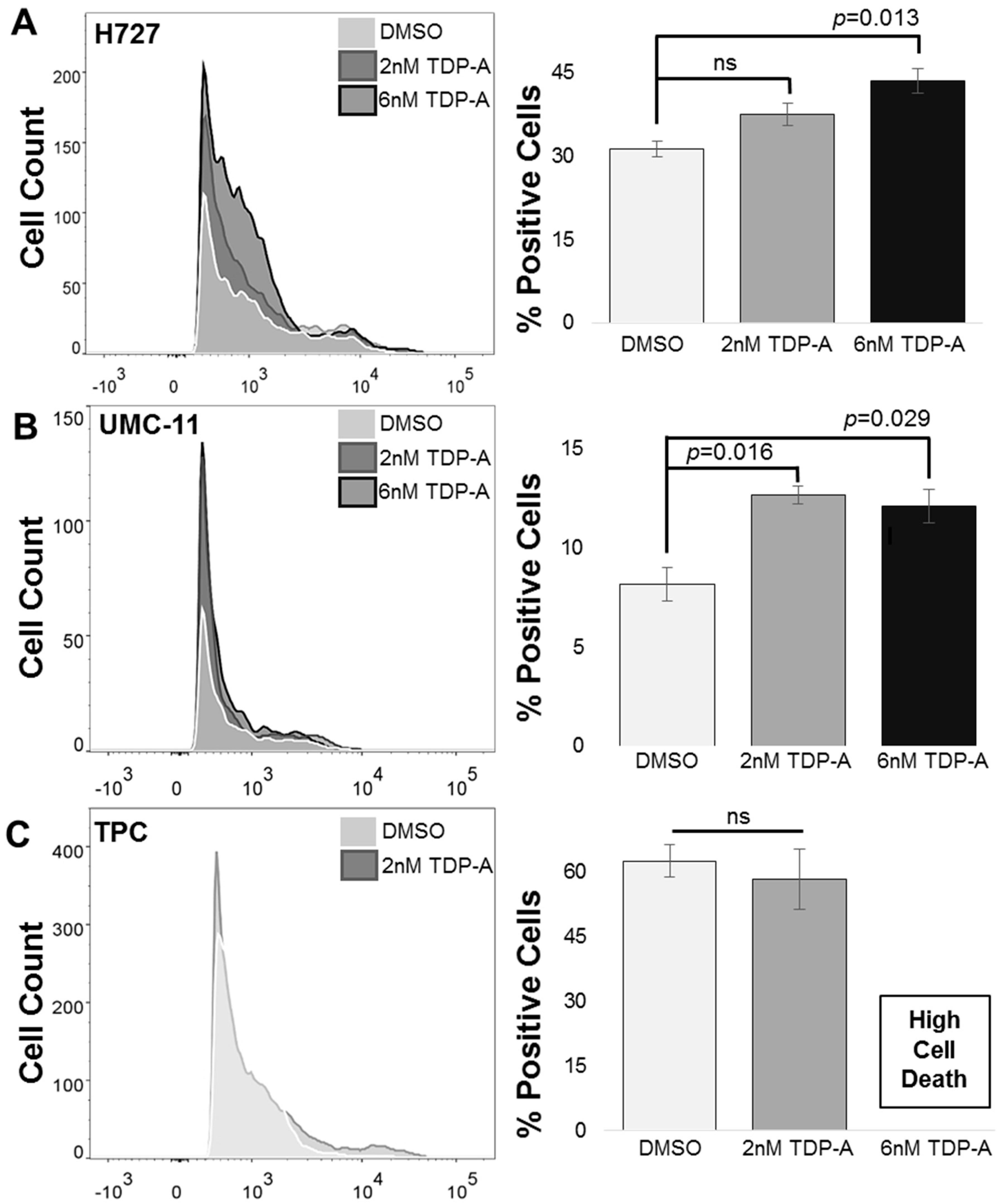
2.2. Detection of Functional SSTR2 Expression in Pulmonary Carcinoids
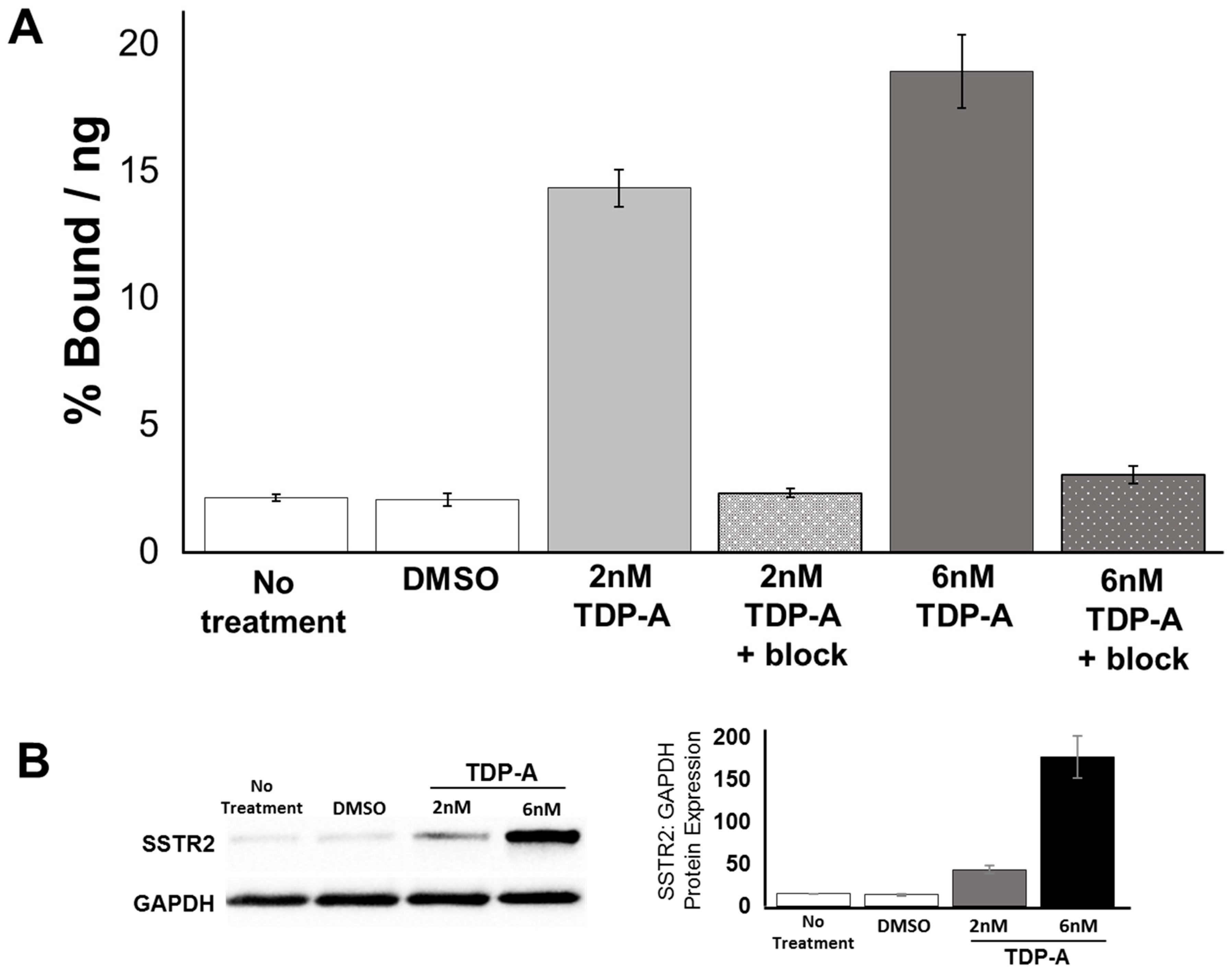
2.3. Cell Uptake of [68Ga]DOTATATE PET/CT In Vitro
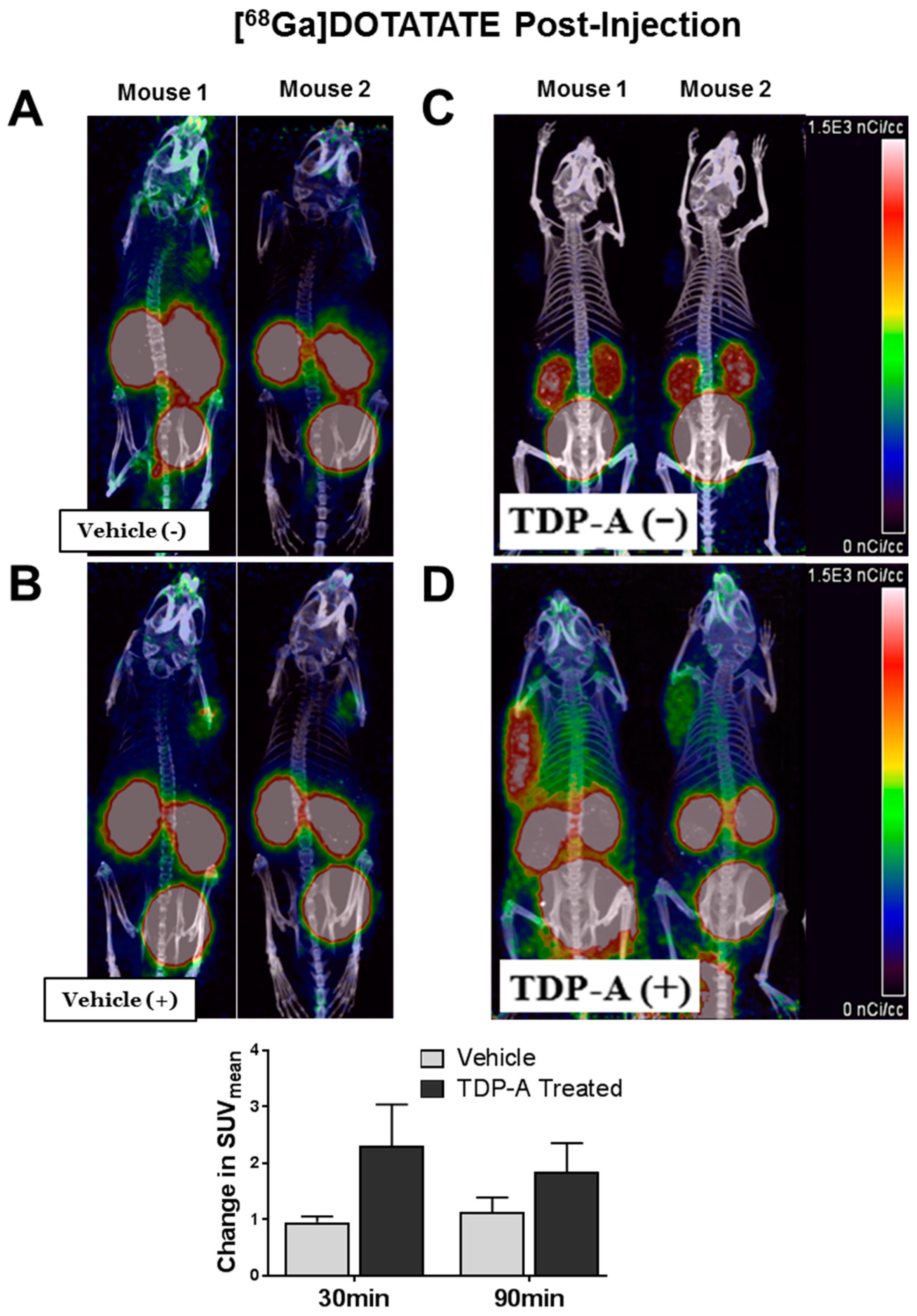
2.4. [68Ga]DOTATATE PET/CT Small Animal Imaging
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Real Time Quantitative PCR
4.3. Western Blot Analysis
4.4. Flow Cytometry
4.5. In Vitro [68Ga]DOTATATE PET/CT Uptake
4.6. Small Animal [68Ga]-DOTATATE PET/CT Imaging
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Travis, W.D. Lung tumours with neuroendocrine differentiation. Eur. J. Cancer 2009, 45, 251–266. [Google Scholar] [CrossRef]
- Chen, L.C.; Travis, W.D.; Krug, L.M. Pulmonary neuroendocrine tumors: What (little) do we know? J. Natl. Compr. Cancer Netw. 2006, 4, 623–630. [Google Scholar] [CrossRef]
- Lim, E. Surigical management of bronchopulmonary and thymic neuroendocrine tumours. Gastroenteropancreatic and thoracic neuroendocrine tumors. BioScienifica 2011, 185–189, in print. [Google Scholar]
- Caplin, M.E.; Pavel, M.; Ruszniewski, P. Lanreotide in metastatic enteropancreatic neuroendocrine tumors. N. Engl. J. Med. 2014, 371, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Otte, A.; Herrmann, R.; Heppeler, A.; Behe, M.; Jermann, E.; Powell, P.; Maecke, H.R.; Muller, J.; Powell, P. Yttrium-90 DOTATOC: First clinical results. Eur. J. Nucl. Med. 1999, 26, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Marincek, N.; Radojewski, P.; Dumont, R.A.; Brunner, P.; Müller-Brand, J.; Maecke, H.R.; Briel, M.; Walter, M.A. Somatostatin receptor-targeted radiopeptide therapy with 90Y-DOTATOC and 177Lu-DOTATOC in progressive meningioma: Long-term results of a phase II clinical trial. J. Nucl. Med. 2015, 56, 171–176. [Google Scholar] [CrossRef]
- Oberg, K. Molecular imaging radiotherapy: Theranostics for personalized patient management of neuroendocrine tumors (NETs). Theranostics 2012, 2, 448–458. [Google Scholar] [CrossRef]
- Karaca, B.; Degirmenci, M.; Ozveren, A.; Atmaca, H.; Bozkurt, E.; Karabulut, B.; Sanli, U.A.; Uslu, R. Docetaxel in combination with octreotide shows synergistic apoptotic effect by increasing SSTR2 and SSTR5 expression levels in prostate and breast cancer cell lines. Cancer Chemother. Pharmacol. 2015, 75, 1273–1280. [Google Scholar] [CrossRef]
- He, Y.; Yuan, X.; Lei, P. The antiproliferative effects of somatostatin receptor subtype 2 in breast cancer cells. Acta Pharamacol. Sin. 2009, 30, 1053–1059. [Google Scholar] [CrossRef] [Green Version]
- Pauwels, E.; Cleeren, F.; Bormans, G.; Deroose, C.M. Somatostatin receptor PET ligands—The next generation for clinical practice. Am. J. Nucl. Med. Mol. Imaging 2018, 8, 311–331. [Google Scholar]
- Reubi, J.C.; Schar, J.C.; Waser, B.; Wenger, S.; Heppeler, A.; Schmitt, J.S.; Macke, H.R. Affinity profiles for human somatostatin receptor subtypes SST1-SST5 of somatostatin radiotracers selected for scintigraphic and radiotherapeutic use. Eur. J. Nucl. Med. 2000, 27, 273–282. [Google Scholar] [CrossRef]
- Wild, D.; Macke, H.R.; Waser, B.; Reubi, J.C.; Ginj, M.; Rasch, H.; Muller-Brand, J.; Hofmann, M. 68Ga-DOTANOC: A first compound for PET imaging with high affinity for somatostatin receptor subtypes 2 and 5. Eur. J. Nucl. Med. Mol. Imaging 2005, 32, 724. [Google Scholar] [CrossRef]
- Righi, L.; Volante, M.; Tavaglione, V.; Billè, A.; Daniele, L.; Angusti, T.; Inzani, F.; Pelosi, G.; Rindi, G.; Papotti, M. Somatostatin receptor tissue distribution in lung neuroendocrine tumours: A clinicopathologic and immunohistochemical study of 218 ‘clinically aggressive’ cases. Ann. Oncol. 2010, 21, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Taelman, V.F.; Radojewski, P.; Marincek, N.; Ben-Shlomo, A.; Grotzky, A.; Olariu, C.; Perren, A.; Stettler, C.; Krause, T.; Meier, L.P.; et al. Upregulation of Key Molecules for Targeted Imaging and Therapy. J. Nucl. Med. 2016, 57, 1805–1810. [Google Scholar] [CrossRef] [Green Version]
- Jaskula-Sztul, R.; Eide, J.; Tesfazghi, S.; Dammalapati, A.; Harrison, A.D.; Yu, M.N.; Scheinebeck, C.; Winston-McPherson, G.; Kupcho, K.R.; Robers, M.B.; et al. Tumor suppressor role of Notch3 in Medullary Thyroid Carcinoma revealed by genetic and pharmacological induction. Mol. Cancer Ther. 2015, 14, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.E.; Lowe, V.J.; Aubry, M.C.; Allen, M.S.; Jett, J.R. The Utility of Fluorodeoxyglucose Positron Emission Tomography in the Evaluation of Carcinoid Tumors Presenting as Pulmonary Nodules. Chest 2006, 131, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Henkes, L.M.; Doughty, L.B.; He, M.; Wang, D.; Meyer-Almes, F.J.; Cheng, Y.Q. Thailandepsins: Bacterial products with potent histone deacetylase inhibitory activities and broad-spectrum antiproliferative activities. J. Nat. Prod. 2011, 74, 2031–2038. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cheng, Y.Q. Thailandepsin A. Acta Crystallogr. Sect. E Struct. Rep. Online 2011, 67, o2948–o2949. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Li, Y.P.; Wang, C.; Ahmad, S.; Vu, M.; Kuma, K.; Cheng, Y.Q.; Lam, K.S. Disulfide cross-linked micelles of novel HDAC inhibitor thailandepsin A for the treatment of breast cancer. Biomaterials 2015, 67, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Weinlander, E.; Somnay, Y.; Harrison, A.D.; Wang, C.; Cheng, Y.Q.; Jaskula-Sztul, R.; Yu, X.M.; Chen, H. The novel histone deacetylase inhibitor thailandepsin A inhibits anaplastic thyroid cancer growth. J. Surg. Res. 2014, 190, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.J.; Cheng, Y.Q.; Khabele, D. Thailandepsins are new small molecule class I HDAC inhibitors with potent cytotoxic activity in ovarian cancer cells: A preclinical study of epigenetic ovarian cancer therapy. J. Ovarian Res. 2012, 5, 12. [Google Scholar] [CrossRef]
- Jang, S.; Janssen, A.; Aburjania, Z.; Robers, M.B.; Harrison, A.; Dammalapati, A.; Cheng, Y.Q.; Chen, H.; Jaskula-Sztul, R. Histone deacetylase inhibitor thailandepsin-A activates Notch signaling and suppresses neuroendocrine cancer cell growth in vivo. Oncotarget 2017, 8, 70828–70840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanakis, G.; Grimelius, L.; Spathis, A.; Tringidou, R.; Rassidakis, G.Z.; Öberg, K.; Kaltsas, G.; Tsolakis, A.V. Expression of Somatostatin Receptors 1-5 and Dopamine Receptor 2 in Lung Carcinoids: Implications for a Therapeutic Role. Neuroendocrinology 2015, 101, 211–222. [Google Scholar] [CrossRef]
- Reubi, J.C.; Waser, B. Triple-Peptide Receptor Targeting In Vitro Allows Detection of All Tested Gut and Bronchial NETs. J. Nucl. Med. 2015, 56, 613–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, F.; Ejeskar, K.; Kogner, P.; Martinsson, T. Gain of chromosome arm 17q is associated with unfavourable prognosis in neuroblastoma, but does not involve mutations in the somatostatin receptor 2 (SSTR2) gene at 17q24. Br. J. Cancer 1999, 81, 1402–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daskalakis, K.; Kaltsas, G.; Öberg, K.; Tsolakis, A.V. Lung Carcinoids: Long-Term Surgical Results and the Lack of Prognostic Value of Somatostatin Receptors and Other Novel Immunohistochemical Markers. Neuroendocrinology 2018, 107, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Sestini, R.; Orlando, C.; Peri, A.; Tricarico, C.; Pazzagli, M.; Serio, M.; Pagani, A.; Bussolati, G.; Granchi, S.; Maggi, M. Quantitation of somatostatin receptor type 2 gene expression in neuroblastoma cell lines and primary tumors using competitive reverse transcription-polymerase chain reaction. Clin. Cancer Res. 1996, 2, 1757–1765. [Google Scholar] [PubMed]
- Jaskula-Sztul, R.; Xu, W.; Chen, G.; Harrison, A.; Dammalapati, A.; Nair, R.; Cheng, Y.; Gong, S.; Chen, H. Thailandepsin A-loaded and octreotide-functionalized unimolecular micelles for targeted neuroendocrine cancer therapy. Biomaterials 2016, 91, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tirosh, A.; Kebebew, E. The utility of 68Ga-DOTATATE positron-emission tomography/computed tomography in the diagnosis, management, follow-up and prognosis of neuroendocrine tumors. Future Oncol. 2018, 14, 111–122. [Google Scholar] [CrossRef]
- Yoon, S.; Eom, G.H. HDAC and HDAC inhibitor: From cancer to cardiovascular diseases. Chonnam. Med. J. 2016, 52, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Somnay, Y.R.; Dull, B.Z.; Eide, J.; Jaskula-Sztul, R.; Chen, H. Chrysin suppresses the achaete-scute complex-like1 and alters the neuroendocrine phenotype of carcinoids. Cancer Gene. Ther. 2015, 22, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Somnay, Y.R.; Yu, X.M.; Lloyd, R.; Leverson, G.; Aburjania, Z.; Jang, S.; Jaskula-Sztul, R.; Chen, H. Notch3 Expression Correlates with Thyroid Cancer Differentiation, Induces Apoptosis, and Predicts Disease Prognosis. Cancer 2017, 123, 769–782. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guenter, R.E.; Aweda, T.; Carmona Matos, D.M.; Whitt, J.; Chang, A.W.; Cheng, E.Y.; Liu, X.M.; Chen, H.; Lapi, S.E.; Jaskula-Sztul, R. Pulmonary Carcinoid Surface Receptor Modulation Using Histone Deacetylase Inhibitors. Cancers 2019, 11, 767. https://doi.org/10.3390/cancers11060767
Guenter RE, Aweda T, Carmona Matos DM, Whitt J, Chang AW, Cheng EY, Liu XM, Chen H, Lapi SE, Jaskula-Sztul R. Pulmonary Carcinoid Surface Receptor Modulation Using Histone Deacetylase Inhibitors. Cancers. 2019; 11(6):767. https://doi.org/10.3390/cancers11060767
Chicago/Turabian StyleGuenter, Rachael E., Tolulope Aweda, Danilea M. Carmona Matos, Jason Whitt, Alexander W. Chang, Eric Y. Cheng, X. Margaret Liu, Herbert Chen, Suzanne E. Lapi, and Renata Jaskula-Sztul. 2019. "Pulmonary Carcinoid Surface Receptor Modulation Using Histone Deacetylase Inhibitors" Cancers 11, no. 6: 767. https://doi.org/10.3390/cancers11060767
APA StyleGuenter, R. E., Aweda, T., Carmona Matos, D. M., Whitt, J., Chang, A. W., Cheng, E. Y., Liu, X. M., Chen, H., Lapi, S. E., & Jaskula-Sztul, R. (2019). Pulmonary Carcinoid Surface Receptor Modulation Using Histone Deacetylase Inhibitors. Cancers, 11(6), 767. https://doi.org/10.3390/cancers11060767