CREPT Promotes Melanoma Progression Through Accelerated Proliferation and Enhanced Migration by RhoA-Mediated Actin Filaments and Focal Adhesion Formation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
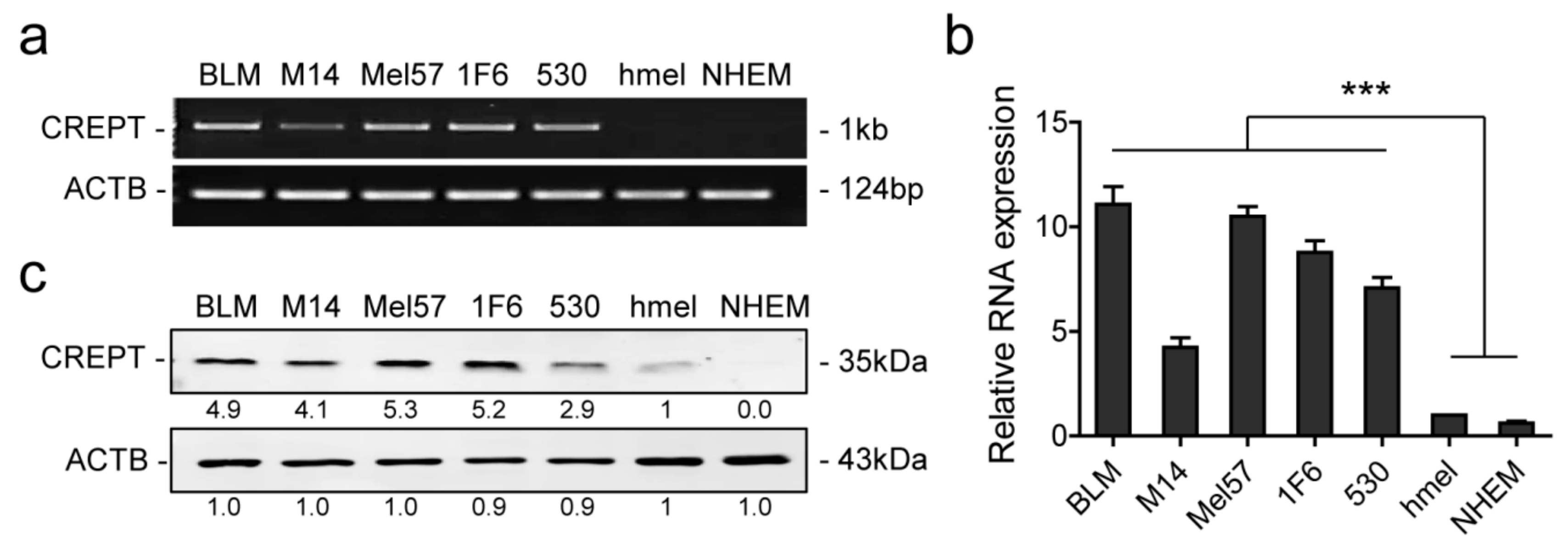
2.1. Differential Expression of CREPT in Melanoma Cells and Melanocytes
2.2. Depleted or Enforced Expression of CREPT Influences Melanoma Cells’ Proliferative Capacity
2.3. CREPT Promotes Melanoma Cell Migration and Invasion In Vitro
2.4. Global Gene Expression Profiling Based on CREPT Expression Modification
2.5. CREPT Regulates Actin Filament Polymerization and Focal Adhesion Formation
2.6. CREPT Influences Focal Adhesion Signaling-Related Molecules
2.7. The Role of RhoA on Actin Filaments and Cell Migration
3. Discussion
4. Materials and Methods
4.1. Antibodies and Plasmids
4.2. Cell Lines and Culture Conditions
4.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Quantitative PCR
- CREPT forward: 5’-TAT AGG TAC CAT GTC CTC CTT CTC TGA G-3’,
- CREPT reverse: 5’-TAT ACT CGA GCT AGT CAG TTG AAA ACA GGT C-3’;
- ACTB forward: 5’- GTC ATT CCA AAT ATG AGA TGC GT-3’,
- ACTB reverse: 5’- AAT GCT ATC ACC TCC CCT GT-3’.
4.4. Western Blotting
4.5. Cell Transfection
4.6. Cell Proliferation Assay
4.7. Colony Formation Assay
4.8. Ring-Barrier Migration Assay
4.9. 3D Invasion Assay
4.10. Gene Expression Profiling
4.11. Immunofluorescence Staining
4.12. Small GTPase Activation Assay
4.13. Inactivation of Rho
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weide, B.; Elsasser, M.; Buttner, P.; Pflugfelder, A.; Leiter, U.; Eigentler, T.K.; Bauer, J.; Witte, M.; Meier, F.; Garbe, C. Serum markers lactate dehydrogenase and s100b predict independently disease outcome in melanoma patients with distant metastasis. Br. J. Cancer 2012, 107, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Robert, C.; Grob, J.J.; Stroyakovskiy, D.; Karaszewska, B.; Hauschild, A.; Levchenko, E.; Sileni, V.C.; Schachter, J.; Garbe, C.; Bondarenko, I.; et al. Five-year outcomes with dabrafenib plus trametinib in metastatic melanoma. N. Engl. J. Med. 2019, 381, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-year survival with combined nivolumab and ipilimumab in advanced melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C.; Ribas, A.; Schachter, J.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.M.; Lotem, M.; et al. Pembrolizumab versus ipilimumab in advanced melanoma (keynote-006): Post-hoc 5-year results from an open-label, multicentre, randomised, controlled, phase 3 study. Lancet Oncol. 2019, 20, 1239–1251. [Google Scholar] [CrossRef]
- Chaffer, C.L.; Weinberg, R.A. A perspective on cancer cell metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef]
- Lauffenburger, D.A.; Horwitz, A.F. Cell migration: A physically integrated molecular process. Cell 1996, 84, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.C. Cell-matrix contact structures. Cell. Mol. Life Sci. 2001, 58, 371–392. [Google Scholar] [CrossRef]
- Burridge, K.; Chrzanowska-Wodnicka, M. Focal adhesions, contractility, and signaling. Annu. Rev. Cell Dev. Biol. 1996, 12, 463–518. [Google Scholar] [CrossRef] [Green Version]
- Cramer, L.P. Organization and polarity of actin filament networks in cells: Implications for the mechanism of myosin-based cell motility. Biochem. Soc. Symp. 1999, 65, 173–205. [Google Scholar] [PubMed]
- Siret, C.; Terciolo, C.; Dobric, A.; Habib, M.C.; Germain, S.; Bonnier, R.; Lombardo, D.; Rigot, V.; Andre, F. Interplay between cadherins and alpha 2 beta 1 integrin differentially regulates melanoma cell invasion. Br. J. Cancer 2015, 113, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, S.J.; Zhang, W.; Ni, N.N.; Wu, Q.; Wang, X.P.; Lin, Y.K.; Sun, J.F. Expression of rho gtpases family in melanoma cells and its influence on cytoskeleton and migration. Oncotarget 2017, 8, 30112–30122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, R.M.; Aplin, A.E. Rnd3 regulation of the actin cytoskeleton promotes melanoma migration and invasive outgrowth in three dimensions. Cancer Res. 2009, 69, 2224–2233. [Google Scholar] [CrossRef] [Green Version]
- Haass, N.K.; Smalley, K.S.; Li, L.; Herlyn, M. Adhesion, migration and communication in melanocytes and melanoma. Pigment Cell Res. 2005, 18, 150–159. [Google Scholar] [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef]
- Das, A.M.; Pescatori, M.; Vermeulen, C.E.; Rens, J.A.; Seynhaeve, A.L.; Koning, G.A.; Eggermont, A.M.; Ten Hagen, T.L. Melanomas prevent endothelial cell death under restrictive culture conditions by signaling through akt and p38 mapk/ erk-1/2 cascades. Oncoimmunology 2016, 5, e1219826. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Olsen, J.B.; Guo, X.; Zhong, G.; Ruan, E.D.; Marcon, E.; Young, P.; Guo, H.; Li, J.; Moffat, J.; et al. Control of the RNA polymerase II phosphorylation state in promoter regions by CTD interaction domain-containing proteins RPRD1A and RPRD1B. Transcription 2011, 2, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Xu, C.; Guo, X.; Hunter, G.O.; Kuznetsova, O.V.; Tempel, W.; Marcon, E.; Zhong, G.; Guo, H.; Kuo, W.W.; et al. RPRD1A and RPRD1B are human RNA polymerase II C-terminal domain scaffolds for Ser5 dephosphorylation. Nat. Struct. Mol. Biol. 2014, 21, 686–695. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.D.; Wu, Y.Y.; Wang, Y.Y.; Ren, F.L.; Wang, D.J.; Su, F.Q.; Zhang, Y.Q.; Yang, X.; Jin, G.H.; Hao, X.B.; et al. Crept accelerates tumorigenesis by regulating the transcription of cell-cycle-related genes. Cancer Cell 2012, 21, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qiu, H.; Hu, W.; Li, S.; Yu, J. RPRD1B promotes tumor growth by accelerating the cell cycle in endometrial cancer. Oncol. Rep. 2014, 31, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yang, L.; He, Y.; Zhu, B.; Ren, F.; Fan, X.; Wang, Y.; Li, M.; Li, J.; Kuang, Y.; et al. Crept/RPRD1B associates with Aurora B to regulate Cyclin B1 expression for accelerating the G2/M transition in gastric cancer. Cell Death Dis. 2018, 9, 1172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, C.; Duan, X.; Ren, F.; Li, S.; Jin, Z.; Wang, Y.; Feng, Y.; Liu, Z.; Chang, Z. Crept/RPRD1B, a recently identified novel protein highly expressed in tumors, enhances the β-catenin·TCF4 transcriptional activity in response to Wnt signaling. J. Biol. Chem. 2014, 289, 22589–22599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, W.M.; Wang, W.P.; Sun, Y.; Ni, Y.F.; Xing, H.; Xia, J.H.; Wang, X.J.; Zhang, Z.P.; Li, X.F. Inhibiting CREPT reduces the proliferation and migration of non-small cell lung cancer cells by down-regulating cell cycle related protein. Am. J. Transl. Res. 2016, 8, 2097–2113. [Google Scholar]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers--assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Li, W.M.; Zheng, G.X.; Xia, J.H.; Yang, G.; Sun, J.Y.; Wang, X.J.; Wen, M.M.; Sun, Y.; Zhang, Z.P.; Jin, F.G. Cell cycle-related and expression-elevated protein in tumor overexpression is associated with proliferation behaviors and poor prognosis in non-small-cell lung cancer. Cancer Sci. 2018, 109, 1012–1023. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.X.; Li, W.M.; Zuo, B.L.; Guo, Z.Y.; Xi, W.J.; Wei, M.; Chen, P.; Wen, W.H.; Yang, A.G. High expression of CREPT promotes tumor growth and is correlated with poor prognosis in colorectal cancer. Biochem. Biophys. Res. Commun. 2016, 480, 436–442. [Google Scholar] [CrossRef]
- She, Y.; Liang, J.; Chen, L.; Qiu, Y.; Liu, N.; Zhao, X.; Huang, X.; Wang, Y.; Ren, F.; Chang, Z.; et al. CREPT expression correlates with poor prognosis in patients with retroperitoneal leiomyosarcoma. Int. J. Clin. Exp. Pathol. 2014, 7, 6596–6605. [Google Scholar]
- Zhang, Y.Q.; Wang, S.Y.; Kang, W.; Liu, C.X.; Dong, Y.J.; Ren, F.L.; Wang, Y.Y.; Zhang, J.L.; Wang, G.P.; To, K.F.; et al. CREPT facilitates colorectal cancer growth through inducing Wnt/beta-catenin pathway by enhancing p300-mediated beta-catenin acetylation. Oncogene 2018, 37, 3485–3500. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Si, G.; Sun, H.S.; Si, F.C. Inhibition of CREPT restrains gastric cancer growth by regulation of cycle arrest, migration and apoptosis via ROS-regulated p53 pathway. Biochem. Biophys. Res. Commun. 2018, 496, 1183–1190. [Google Scholar] [CrossRef]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Ciobanasu, C.; Faivre, B.; Le Clainche, C. Actin dynamics associated with focal adhesions. Int. J. Cell Biol. 2012, 2012, 941292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezratty, E.J.; Partridge, M.A.; Gundersen, G.G. Microtubule-induced focal adhesion disassembly is mediated by dynamin and focal adhesion kinase. Nat. Cell Biol. 2005, 7, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Hoock, S.C.; Ritter, A.; Steinhauser, K.; Roth, S.; Behrends, C.; Oswald, F.; Solbach, C.; Louwen, F.; Kreis, N.N.; Yuan, J. Rita modulates cell migration and invasion by affecting focal adhesion dynamics. Mol. Oncol. 2019, 13, 2121–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Wirtz, D. Focal adhesion size uniquely predicts cell migration. FASEB J. 2013, 27, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, M.; Zigmond, S.; Boone, C. Formins: Signaling effectors for assembly and polarization of actin filaments. J. Cell Sci. 2003, 116, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Rose, R.; Weyand, M.; Lammers, M.; Ishizaki, T.; Ahmadian, M.R.; Wittinghofer, A. Structural and mechanistic insights into the interaction between rho and mammalian dia. Nature 2005, 435, 513–518. [Google Scholar] [CrossRef]
- Ren, F.; Wang, R.; Zhang, Y.; Liu, C.; Wang, Y.; Hu, J.; Zhang, L.; Chang, Z. Characterization of a monoclonal antibody against CREPT, a novel protein highly expressed in tumors. Monoclon. Antib. Immunodiagn. Immunother. 2014, 33, 401–408. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine b colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Das, A.M.; Eggermont, A.M.; ten Hagen, T.L. A ring barrier-based migration assay to assess cell migration in vitro. Nat. Protoc. 2015, 10, 904–915. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using david bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horzum, U.; Ozdil, B.; Pesen-Okvur, D. Step-by-step quantitative analysis of focal adhesions. MethodsX 2014, 1, 56–59. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Seynhaeve, A.L.B.; Brouwer, R.W.W.; van IJcken, W.F.J.; Yang, L.; Wang, Y.; Chang, Z.; ten Hagen, T.L.M. CREPT Promotes Melanoma Progression Through Accelerated Proliferation and Enhanced Migration by RhoA-Mediated Actin Filaments and Focal Adhesion Formation. Cancers 2020, 12, 33. https://doi.org/10.3390/cancers12010033
Liu H, Seynhaeve ALB, Brouwer RWW, van IJcken WFJ, Yang L, Wang Y, Chang Z, ten Hagen TLM. CREPT Promotes Melanoma Progression Through Accelerated Proliferation and Enhanced Migration by RhoA-Mediated Actin Filaments and Focal Adhesion Formation. Cancers. 2020; 12(1):33. https://doi.org/10.3390/cancers12010033
Chicago/Turabian StyleLiu, Hui, Ann L. B. Seynhaeve, Rutger W. W. Brouwer, Wilfred F. J. van IJcken, Liu Yang, Yinyin Wang, Zhijie Chang, and Timo L. M. ten Hagen. 2020. "CREPT Promotes Melanoma Progression Through Accelerated Proliferation and Enhanced Migration by RhoA-Mediated Actin Filaments and Focal Adhesion Formation" Cancers 12, no. 1: 33. https://doi.org/10.3390/cancers12010033
APA StyleLiu, H., Seynhaeve, A. L. B., Brouwer, R. W. W., van IJcken, W. F. J., Yang, L., Wang, Y., Chang, Z., & ten Hagen, T. L. M. (2020). CREPT Promotes Melanoma Progression Through Accelerated Proliferation and Enhanced Migration by RhoA-Mediated Actin Filaments and Focal Adhesion Formation. Cancers, 12(1), 33. https://doi.org/10.3390/cancers12010033