The Role of E3, E4 Ubiquitin Ligase (UBE4B) in Human Pathologies


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Ubiquitination and Ubiquitin Proteasome System
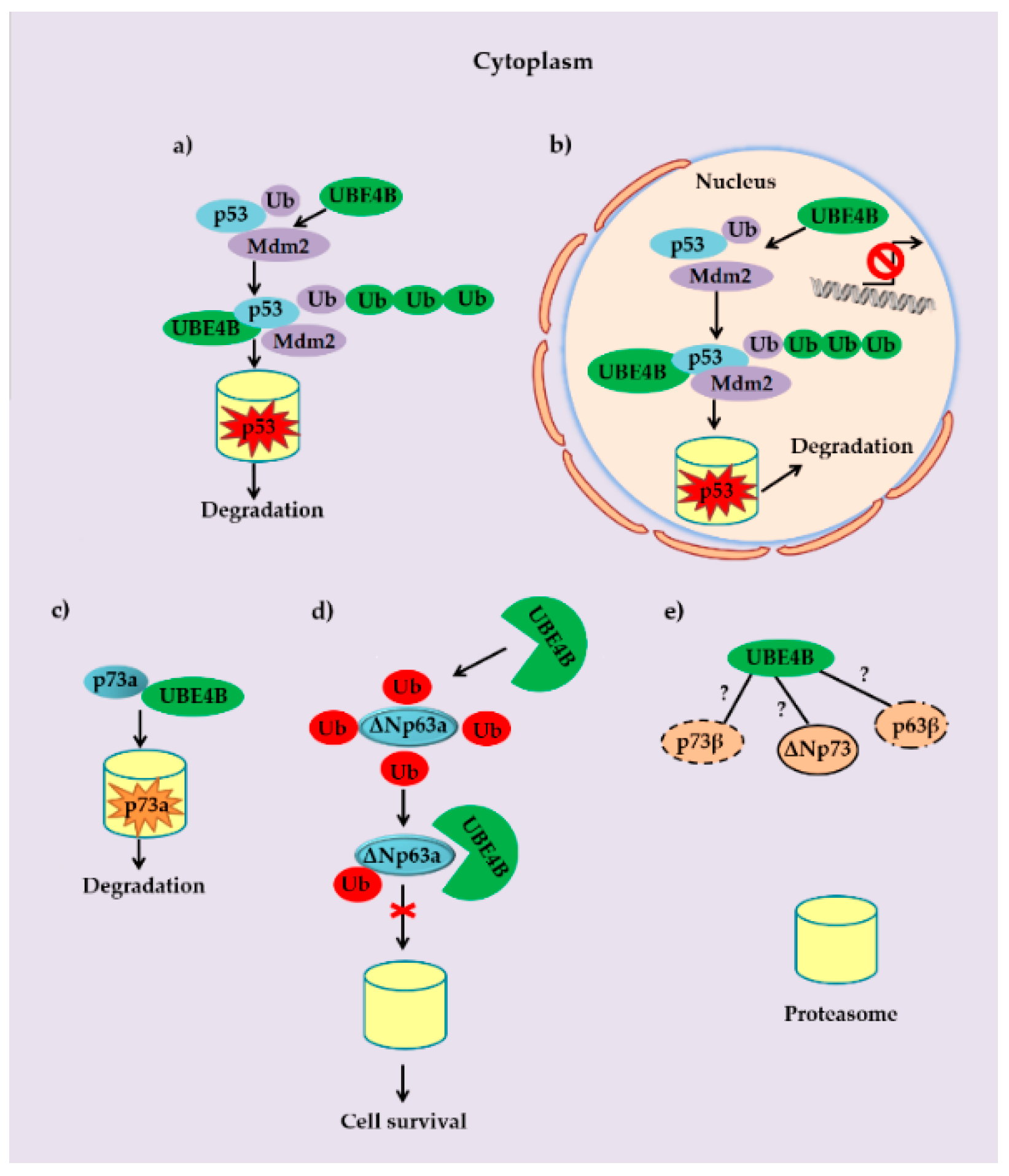
3. UBE4B and p53 Family Members
4. UBE4B Ubiquitin Ligase in Human Pathology
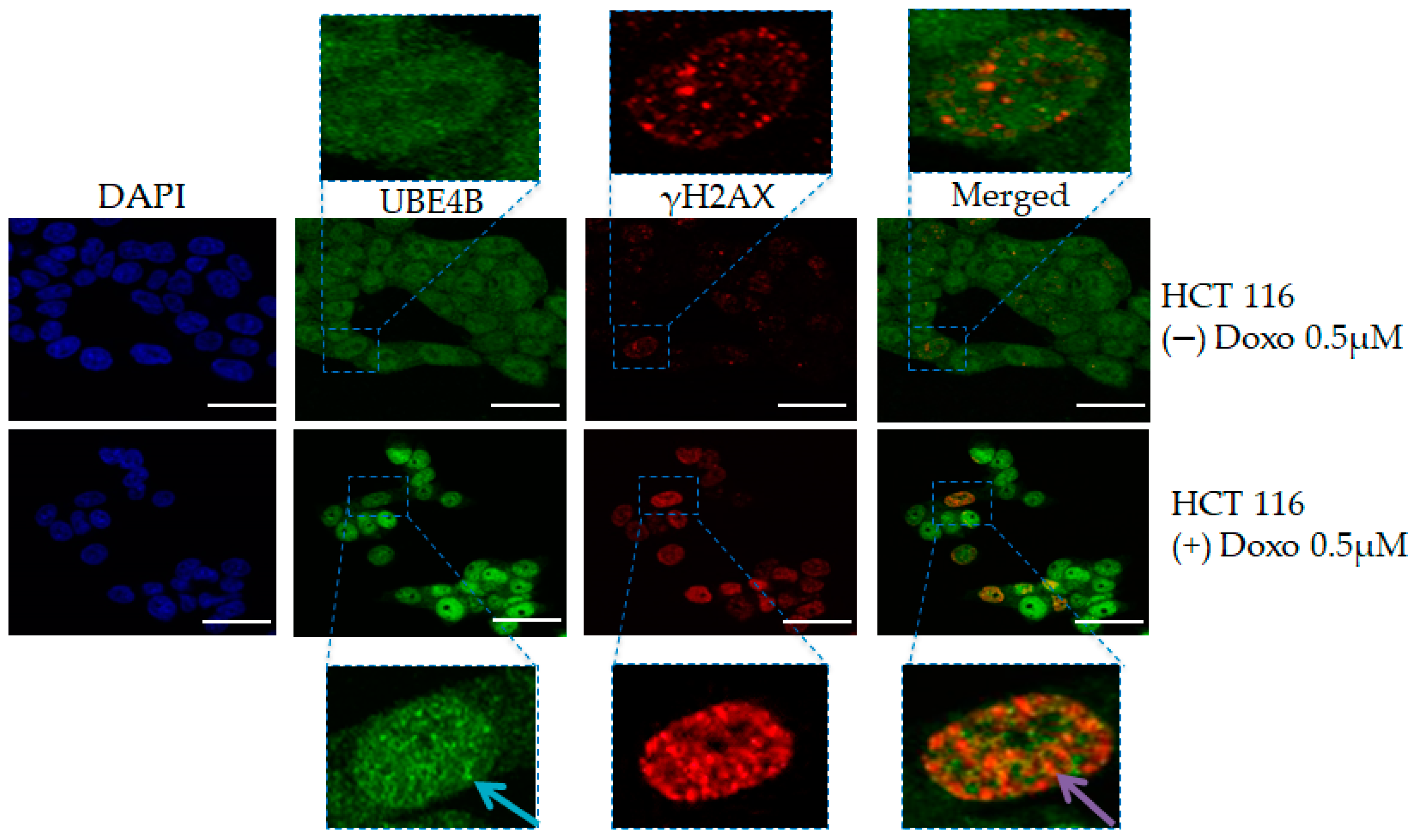
4.1. UBE4B and Neurodegenerative Diseases
4.2. UBE4B and Cancer
5. Druggability Analysis
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weissman, A.M. Themes and variations on ubiquitylation. Nat. Rev. Mol. Cell Biol. 2001, 2, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chen, Z.J. The novel functions of ubiquitination in signaling. Curr. Opin. Cell Biol. 2004, 16, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Ciechanover, A. The ubiquitin system. Ann. Rev. Biochem. 1998, 67, 425–479. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.L.; Wickliffe, K.E.; Dong, K.C.; Yu, C.; Bosanac, I.; Bustos, D.; Phu, L.; Kirkpatrick, D.S.; Hymowitz, S.G.; Rape, M.; et al. K11-linked polyubiquitination in cell cycle control revealed by a K11 linkage-specific antibody. Mol. Cell 2010, 39, 477–484. [Google Scholar] [CrossRef]
- Chondrogianni, N.; Gonos, E. Structure and Function of the Ubiquitin–Proteasome System: Modulation of Components. Prog. Mol. Biol. Transl. Sci. 2012, 109, 41–74. [Google Scholar] [CrossRef]
- Streich, F.; Lima, C. Structural and Functional Insights to Ubiquitin-Like Protein Conjugation. Ann. Rev. Biophys. 2014, 43, 357–379. [Google Scholar] [CrossRef] [Green Version]
- Karve, T.M.; Cheema, A.K. Small Changes Huge Impact: The Role of Protein Posttranslational Modifications in Cellular Homeostasis and Disease. J. Amino Acids 2011. [Google Scholar] [CrossRef] [Green Version]
- Contino, G.; Amati, F.; Pucci, S.; Pontieri, E.; Pichiorri, F.; Novelli, A.; Botta, A.; Mango, R.; Nardone, A.M.; Sangiuolo, F.C.; et al. Expression analysis of the gene encoding for the U-box-type ubiquitin ligase UBE4A in human tissues. Gene 2004, 328, 69–74. [Google Scholar] [CrossRef]
- Hosoda, M.; Ozaki, T.; Miyazaki, K.; Hayashi, S.; Furuya, K.; Watanabe, K.; Nakagawa, T.; Hanamoto, T.; Todo, S.; Nakagawara, A. UFD2a mediates the proteasomal turnover of p73 without promoting p73 ubiquitination. Oncogene 2005, 24, 7156–7169. [Google Scholar] [CrossRef] [Green Version]
- Caren, H.; Holmstrand, A.; Sjöberg, R.M.; Martinsson, T. The two human homologues of yeast UFD2 ubiquitination factor, UBE4A and UBE4B, are located in common neuroblastoma deletion regions and are subject to mutations in tumours. Eur. J. Cancer 2006, 42, 381–387. [Google Scholar] [CrossRef]
- Jana, N.R. Protein homeostasis and aging: Role of ubiquitin protein ligases. Neurochem. Int. 2012, 60, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. Intracellular protein degradation: From a vague idea thru the lysosome and the ubiquitin–proteasome system and onto human diseases and drug targeting. Cell Death Differ. 2005, 12, 1178–1190. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulos, K.; Kriegenburg, F.; Tatham, M.H.; Rosner, H.I.; Medina, B.; Larsen, I.B.; Brandstrup, R.; Hardwick, K.G.; Hay, R.T.; Kragelund, B.B.; et al. Dss1 Is a 26S Proteasome Ubiquitin Receptor. Mol. Cell 2014, 56, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.E.; Sarkar, S.; Rubinsztein, D.C. The ubiquitin proteasome system in Huntington’s disease and the spinocerebellar ataxias. BMC Biochem. 2007, 8 (Suppl. 1), S2. [Google Scholar] [CrossRef] [Green Version]
- Finley, D. Recognition and processing of ubiquitin-protein conjugates by the proteasome. Ann. Rev. Biochem. 2009, 78, 477–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochstrasser, M. Protein degradation or regulation: Ub the judge. Cell 1996, 84, 813–815. [Google Scholar] [CrossRef] [Green Version]
- Kerscher, O.; Felberbaum, R.; Hochstrasser, M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Ann. Rev. Cell Dev. Biol. 2006, 22, 159–180. [Google Scholar] [CrossRef] [Green Version]
- Varshavsky, A. The early history of the ubiquitin field. Protein Sci. 2006, 15, 647–654. [Google Scholar] [CrossRef] [Green Version]
- Pickart, C.M. Mechanisms underlying ubiquitination. Ann. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef]
- Schrader, E.K.; Harstad, K.G.; Matouschek, A. Targeting proteins for degradation. Nat. Chem. Biol. 2009, 5, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, S.K. Seven-transmembrane receptors and ubiquitination. Circ. Res. 2007, 100, 1142–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndsen, C.; Wolberger, C. New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 2014, 21, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Hoeller, D.; Dikic, I. Targeting the ubiquitin system in cancer therapy. Nature 2009, 458, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Vucic, D.; Dixit, V.M.; Wertz, I.E. Ubiquitylation in apoptosis: A post-translational modification at the edge of life and death. Nat. Rev. Mol. Cell Biol. 2011, 12, 439–452. [Google Scholar] [CrossRef]
- Buetow, L.; Huang, D.T. Structural insights into the catalysis and regulation of E3 ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 2016, 17, 626–642. [Google Scholar] [CrossRef] [Green Version]
- Morreale, F.; Walden, H. Types of Ubiquitin Ligases. Cell 2016, 165, 248. [Google Scholar] [CrossRef]
- Metzger, M.B.; Pruneda, J.N.; Klevit, R.E.; Weissman, A.M. RING-type E3 ligases: Master manipulators of E2 ubiquitin-conjugating enzymes and ubiquitination. Biochim. Biophys. Acta 2014, 1843, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Ronai, Z.A. Dysregulation of ubiquitin ligases in cancer. Drug Resist. Updat. 2015, 23, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lu, L.Y.; Yu, X. The role of BRCA1 in DNA damage response. Protein Cell 2010, 1, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wang, C.; Huang, K.; Xia, F.; Parvin, J.D. Mondal N3Regulation of 53BP1 Protein Stability by RNF8 and RNF168 Is Important for Efficient DNA Double-Strand Break Repair. PLoS ONE 2014, 9, e110522. [Google Scholar] [CrossRef]
- Chroma, K.; Mistrik, M.; Moudry, P.; Gursky, J.; Liptay, M.; Strauss, R.; Skrott, Z.; Vrtel, R.; Bartkova, J.; Kramara, J.; et al. Tumors overexpressing RNF168 show altered DNA repair and responses to genotoxic treatments, genomic instability and resistance to proteotoxic stress. Oncogene 2016, 36, 2405–2422. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, T. Multiubiquitylation by E4 enzymes: ‘one size’ doesn’t fit all. Trends Biochem. Sci. 2005, 30, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Pop, M.S.; Kulikov, R.; Love, I.M.; Kung, A.L.; Grossman, S.R. CBP and p300 are cytoplasmic E4 polyubiquitin ligases for p53. Proc. Natl. Acad. Sci. USA 2009, 106, 16275–16280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatakeyama, S.; Yada, M.; Matsumoto, M.; Ishida, N.; Nakayama, K.I. U box proteins as a new family of ubiquitin-protein ligases. J. Biol. Chem. 2001, 276, 33111–33120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranes-Bachar, K.; Levy-Barda, A.; Oehler, J.; Ramadan, K.; Ziv, Y.; Shiloh, Y. The Ubiquitin E3/E4 Ligase UBE4A Adjusts Protein Ubiquitylation and Accumulation at Sites of DNA Damage, Facilitating Double-Strand Break Repair. Mol. Cell 2018, 69, 866–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.S.; Ma, P.C.; Ota, I.M.; Varshavsky, A. A proteolytic pathway that recognizes ubiquitin as a degradation signal. J. Biol. Chem. 1995, 270, 17442–17456. [Google Scholar] [CrossRef] [Green Version]
- Koegl, M.; Hoppe, T.; Schlenker, S.; Ulrich, H.D.; Mayer, T.U.; Jentsch, S. A novel ubiquitination factor, E4, is involved in multiubiquitin chain assembly. Cell 1999, 96, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lv, Y.; Zhang, Y.; Gao, H. Regulation of p53 Level by UBE4B in Breast Cancer. PLoS ONE 2014, 9, e90154. [Google Scholar] [CrossRef]
- Ackermann, L.; Schell, M.; Pokrzywa, W.; Kevei, É.; Gartner, A.; Schumacher, B.; Hoppe, T. E4 ligase—Specific ubiquitination hubs coordinate DNA double-strand-break repair and apoptosis. Nat. Struct. Mol. Biol. 2016, 23, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Kon, N.; Li, M.; Zhang, W.; Qin, J.; Gu, W. ARF-BP1/Mule is a critical mediator of the ARF tumor suppressor. Cell 2005, 121, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Brooks, C.L.; Gu, W. p53 ubiquitination: Mdm2 and beyond. Mol. Cell 2006, 21, 307–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sane, S.; Rezvani, K. Essential Roles of E3 Ubiquitin Ligases in p53 Regulation. Int. J. Mol. Sci. 2017, 18, 442. [Google Scholar] [CrossRef] [PubMed]
- Bond, G.L.; Hirshfield, K.M.; Kirchhoff, T.; Alexe, G.; Bond, E.E.; Robins, H.; Bartel, F.; Taubert, H.; Wuerl, P.; Hait, W.; et al. MDM2 SNP309 Accelerates Tumor Formation in a Gender-Specific and Hormone-Dependent Manner. Cancer Res. 2006, 66, 5104–5410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urso, L.; Calabrese, F.; Favaretto, A.; Conte, P.; Pasello, G. Critical review about MDM2 in cancer: Possible role in malignant mesothelioma and implications for treatment. Crit. Rev. Oncol. Hematol. 2016, 97, 220–230. [Google Scholar] [CrossRef]
- Medina-Medina, I.; Martínez-Sánchez, M.; Hernández-Monge, J.; Fahraeus, R.; Muller, P.; Olivares-Illana, V. p53 promotes its own polyubiquitination by enhancing the HDM2 and HDMX interaction. Protein Sci. 2018, 27, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Pomeroy, S.L.; Ferreira, M.; Teider, N.; Mariani, J.; Nakayama, K.I.; Hatakeyama, S.; Tron, V.A.; Saltibus, L.F.; Spyracopoulos, L.; et al. UBE4B promotes Hdm2-mediated degradation of the tumor suppressor p53. Nat. Med. 2011, 17, 347–355. [Google Scholar] [CrossRef]
- Zeinab, R.A.; Wu, H.; Sergi, C.; Leng, R. UBE4B: A Promising Regulatory Molecule in Neuronal Death and Survival. Int. J. Mol. Sci. 2012, 13, 16865–16879. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Jensen, J.P.; Ludwig, R.L.; Vousden, K.H.; Weissman, A.M. Mdm2 is a RING finger-dependent ubiquitin protein ligase for itself and p53. J. Biol. Chem. 2000, 275, 8945–8951. [Google Scholar] [CrossRef] [Green Version]
- Mammen, A.L.; Mahoney, J.A.; St Germain, A.; Badders, N.; Taylor, J.P.; Rosen, A.; Spinette, S. A Novel Conserved Isoform of the Ubiquitin Ligase UFD2a/UBE4B Is Expressed Exclusively in Mature Striated Muscle Cells. PLoS ONE 2011, 6, e28861. [Google Scholar] [CrossRef]
- Riley, M.F.; Lozano, G. The Many Faces of MDM2 Binding Partners. Genes Cancer 2012, 3, 226–239. [Google Scholar] [CrossRef] [Green Version]
- Iwakuma, T.; Lozano, G. MDM2, An Introduction. Mol. Cancer Res. 2003, 1, 933–1000. [Google Scholar] [CrossRef] [Green Version]
- Haupt, S.; Vijayakumaran, R.; Jeffreena Miranda, P.; Burgess, A.; Lim, E.; Haupt, Y. The role of MDM2 and MDM4 in breast cancer development and prevention. J. Mol. Cell Biol. 2017, 9, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nag, S.; Qin, J.; Srivenugopal, K.S.; Wang, M.; Zhang, R. The MDM2-p53 pathway revisited. J. Biomed. Res. 2013, 27, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Gu, W. Dual Roles of MDM2 in the Regulation of p53: Ubiquitination Dependent and Ubiquitination Independent Mechanisms of MDM2 Repression of p53 Activity. Genes Cancer 2012, 3, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Pant, V.; Lozano, G. Limiting the power of p53 through the ubiquitin proteasome pathway. Genes Dev. 2018, 28, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.F.; Pan, Q.Z.; Pan, K.; Weng, D.S.; Wang, Q.J.; Zhao, J.J.; He, J.; Liu, Q.; Wang, D.D.; Jiang, S.S.; et al. Expression and Prognostic Role of Ubiquitination Factor E4B in Primary Hepatocellular Carcinoma. Mol. Carcinog. 2016, 55, 64–76. [Google Scholar] [CrossRef]
- Du, C.; Wu, H.; Leng, R.P. UBE4B targets phosphorylated p53 at serines 15 and 392 for degradation. Oncotarget 2016, 7, 2823–2836. [Google Scholar] [CrossRef]
- Slack, R.S.; Belliveau, D.J.; Rosenberg, M.; Atwal, J.; Lochmuller, H.; Aloyz, R.; Haghighi, A.; Lach, B.; Seth, P.; Cooper, E.; et al. Adenovirus-mediated gene transfer of the tumor suppressor, p53, induces apoptosis in postmitotic neurons. J. Cell Biol. 1996, 135, 1085–1096. [Google Scholar] [CrossRef]
- Yuan, J.; Yankner, B.A. Apoptosis in the nervous system. Nature 2000, 407, 802–809. [Google Scholar] [CrossRef]
- Matsumoto, M.; Yada, M.; Hatakeyama, S.; Ishimoto, H.; Tanimura, T.; Tsuji, S.; Kakizuka, A.; Kitagawa, M.; Nakayama, K.I. Molecular clearance of ataxin-3 is regulated by a mammalian E4. EMBO J. 2004, 23, 659–669. [Google Scholar] [CrossRef]
- Ward, J.M.; La Spada, A.R. Ataxin-3, DNA Damage Repair, and SCA3 Cerebellar Degeneration: On the Path to Parsimony? PLoS Genet. 2015, 11, e1004937. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, S.R.; Wu, H.; Wang, B.; Abuetabh, Y.; Sergi, C.; Leng, R.P. The Regulation of Tumor Suppressor p63 by the Ubiquitin-Proteasome System. Int. J. Mol. Sci. 2016, 17, 2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, W.B.; Govoni, G.; Ho, D.; Atwal, J.K.; Barnabe-Heider, F.; Keyes, W.M.; Mills, A.A.; Miller, F.D.; Kaplan, D.R. p63 is an essential proapoptotic protein during neural development. Neuron 2005, 48, 743–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awais, R.; Spiller, D.G.; White, M.R.H.; Paraoan, L. p63 is required beside p53 for PERP-mediated apoptosis in uveal melanoma. Br. J. Cancer 2016, 115, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Upadhyay, S.; Chang, X.; Nagpal, J.K.; Trink, B.; Sidransky, D. U-box-type ubiquitin E4 ligase, UFD2a attenuates cisplatin mediated degradation of ΔNp63α. Cell Cycle 2008, 7, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Nakagawara, A. p73, a sophisticated p53 family member in the cancer world. Cancer Sci. 2005, 96, 729–737. [Google Scholar] [CrossRef]
- Melino, G.; de Laurenzi, V.; Vousden, K.H. p73: Friend or foe in tumorigenesis. Nat. Rev. Cancer 2002, 2, 605–615. [Google Scholar] [CrossRef]
- Jost, C.A.; Marin, M.C.; Kaelin, W.G., Jr. p73 is a simian (correction of human) p53-related protein that can induce apoptosis. Nature 1997, 389, 191–194. [Google Scholar] [CrossRef]
- Goldschneider, D.; Horvilleur, E.; Plassa, L.F.; Guillaud-Bataille, M.; Million, K.; Wittmer-Dupret, E.; Danglot, G.; de Thé, H.; Bénard, J.; May, E.; et al. Expression of C-terminal deleted p53 isoforms in neuroblastoma. Nucleic Acids Res. 2006, 34, 5603–5612. [Google Scholar] [CrossRef] [Green Version]
- Flores, E.R.; Tsai, K.Y.; Crowley, D.; Sengupta, S.; Yang, A.; McKeon, F.; Jacks, T. p63 and p73 are required for p53-dependent apoptosis in response to DNA damage. Nature 2002, 416, 560–564. [Google Scholar] [CrossRef]
- Rossi, M.; De Laurenzi, V.; Munarriz, E.; Green, D.R.; Liu, Y.C.; Vousden, K.H.; Cesareni, G.; Melino, G. The ubiquitinprotein ligase Itch regulates p73 stability. EMBO J. 2005, 24, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasini, R.; Mak, T.W.; Melino, G. The impact of p53 and p73 on aneuploidy and cancer. Trends Cell Biol. 2008, 18, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Chen, L.; Jost, C.A.; Maya, R.; Keller, D.; Wang, X.; Kaelin, W.G., Jr.; Oren, M.; Chen, J.; Lu, H. MDM2 suppresses p73 function without promoting p73 degradation. Mol. Cell. Biol. 1999, 19, 3257–3266. [Google Scholar] [CrossRef] [Green Version]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An oncogene induced DNA damage model for cancer development. Science 2008, 319, 1352–1355. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wu, H.; Chai, C.; Lewis, J.; Pichiorri, F.; Eisenstat, D.D.; Pomeroy, S.L.; Leng, R.P. MicroRNA-1301 suppresses tumor cell migration and invasion by targeting the p53/UBE4B pathway in multiple human cancer cells. Cancer Lett. 2017, 401, 20–32. [Google Scholar] [CrossRef]
- Gorgoulis, V.G.; Pefani, D.-E.; Pateras, I.S.; Trougakos, I.P. Integrating the DNA damage and protein stress responses during cancer development and treatment. J. Pathol. 2018, 246, 12–40. [Google Scholar] [CrossRef] [Green Version]
- Park, C.W.; Ryu, K.Y. Cellular ubiquitin pool dynamics and homeostasis. BMB Rep. 2014, 47, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Kaneko-Oshikawa, C.; Nakagawa, T.; Yamada, M.; Yoshikawa, H.; Matsumoto, M.; Yada, M.; Hatakeyama, S.; Nakayama, K.; Nakayama, K.I. Mammalian E4 is required for cardiac development and maintenance of the nervous system. Mol. Cell Biol. 2005, 25, 10953–10964. [Google Scholar] [CrossRef] [Green Version]
- Spinette, S.; Lengauer, C.; Mahoney, J.A.; Jallepalli, P.V.; Wang, Z.; Casciola-Rosen, L.; Rosen, A. Ufd2, a Novel Autoantigen in Scleroderma, Regulates Sister Chromatid Separation. Cell Cycle 2004, 3, 1638–1644. [Google Scholar] [CrossRef]
- Fernández-Sáiz, V.; Buchberger, A. Imbalances in p97 co-factor interactions in human proteinopathy. EMBO Rep. 2010, 11, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Krona, C.; Ejeskär, K.; Abel, F.; Kogner, P.; Bjelke, J.; Björk, E.; Sjöberg, R.M.; Martinsson, T. Screening for gene mutations in a 500 kb neuroblastoma tumor suppressor candidate region in chromosome 1p; mutation and stage-specific expression in UBE4B/UFD2. Oncogene 2003, 22, 2343–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefeuvre, M.; Gunduz, M.; Nagatsuka, H.; Gunduz, E.; Al Sheikh Ali, M.; Beder, L.; Fukushima, K.; Yamanaka, N.; Shimizu, K.; Nagai, N. Fine deletion analysis of 1p36 chromosomal region in oral squamous cell carcinomas. J. Oral. Pathol. Med. 2009, 38, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.P.; Freeman, M.R. Wallerian Degeneration, WldS, and Nmnat. Ann. Rev. Neurosci. 2010, 33, 245–267. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release 2018-1: SiteMap, Version 4.6. Available online: https://www.schrodinger.com/citations (accessed on 8 October 2019).
- Yuan, Y.; Pei, J.; Lai, L. Binding Site Detection and Druggability Prediction of Protein Targets for Structure-Based Drug Design. Curr. Pharm. Des. 2013, 19, 2326–2333. [Google Scholar] [CrossRef]
- Le Guilloux, V.; Schmidtke, P.; Tuffery, P. Fpocket: An open source platform for ligand pocket detection. BMC Bioinform. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release 2018-1: Prime, Version 5.1. Available online: https://www.schrodinger.com/prime (accessed on 8 October 2019).
- Baek, G.H.; Kim, I.; Rao, H. The Cdc48 ATPase modulates the interaction between two proteolytic factors Ufd2 and Rad23. Proc. Natl. Acad. Sci. USA 2011, 108, 13558–13563. [Google Scholar] [CrossRef] [Green Version]
- Tu, D.; Li, W.; Ye, Y.; Brunger, A.T. Structure and function of the yeast U-box-containing ubiquitin ligase Ufd2p. Proc. Natl. Acad. Sci. USA 2007, 104, 15599–15606. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Yoon, S.K.; Yoon, J.B. TRIP12 functions as an E3 ubiquitin ligase of APP-BP1. Biochem. Biophys. Res. Commun. 2008, 374, 294–298. [Google Scholar] [CrossRef]
- Starita, L.M.; Pruneda, J.N.; Lo, R.S.; Fowler, D.M.; Kim, H.J.; Hiatt, J.B.; Shendure, J.; Brzovic, P.S.; Fields, S.; Klevit, R.E. Activity enhancing mutations in an E3 ubiquitin ligase identified by high-throughput mutagenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 1263–1272. [Google Scholar] [CrossRef] [Green Version]
- Periz, G.; Lu, J.; Zhang, T.; Kankel, M.W.; Jablonski, A.M.; Kalb, R.; McCampbell, A.; Wang, J. Regulation of Protein Quality Control by UBE4B and LSD1 through p53-Mediated Transcription. PLoS Biol. 2015, 13, e1002114. [Google Scholar] [CrossRef] [PubMed]
- Ortolan, T.G.; Tongaonkar, P.; Lambertson, D.; Chen, L.; Schauber, C.; Madura, K. The DNA repair protein Rad23 is a negative regulator of multi-ubiquitin chain assembly. Nat. Cell Biol. 2000, 2, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, K.; Schell, M.; Hoppe, T.; Kashkar, H. Regulation of the DNA damage response by ubiquitin conjugation. Front. Genet. 2015, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citterio, E. Fine-tuning the ubiquitin code at DNA double-strand breaks: Deubiquitinating enzymes at work. Front. Genet. 2015, 6, 282. [Google Scholar] [CrossRef] [Green Version]
- Zage, P.E.; Sirisaengtaksin, N.; Liu, Y.; Gireud, M.; Brown, B.S.; Palla, S.; Richards, K.N.; Hughes, D.P.; Bean, A.J. UBE4B levels are correlated with clinical outcomes in neuroblastoma patients and with altered neuroblastoma cell proliferation and sensitivity to epidermal growth factor receptor inhibitors. Cancer 2013, 119, 915–923. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data from Cellular, Animal Models and Tissues | ||||||
|---|---|---|---|---|---|---|
| Pathological Condition | Cause | Role of UBE4B | Possible Alteration (s) of UBE4B | Type of Organism | Type of Tissue | Reference |
| Machado-Joseph disease/Spinocerebellar ataxia type 3 (SCA3) | increased CAG repeat tract in ATXN3 | Ectopically overexpressed UBE4B poly-ubiquitinates ataxin-3 resulting in the degradation of the latter | − | Human cancer cell lines and SCA3 Drosophila model | − | [60] |
| Scleroderma_Autoimmune disease | − | Autoantigen in 10% of patients, regulation of mitotic progression | Phosphorylated during mitosis as a result its conformation changes dramatically and the protein doesn’t work properly leading to mitotic abnormalities | Human cancer cell lines | − | [79] |
| Inclusion body myopathy with Paget disease of bone and frontotemporal dementia (IBMPFD) | Mutant VCP | Interaction of mutated p97 with UBE4B weakens while its interaction with ataxin-3 becomes intensified | − | Human cancer cell lines | Muscular | [80] |
| Neuroblastoma (NBL) | MYCN amplification and chromosome 1p deletions | − | Deletion of the UBE4B gene/one base substitution in exon 9 observed in stage 3 tumor (dysfunctional protein)/low expression protein in high stage tumors with poor prognosis | NBL cell line | NBL frozen tissues, peripheral blood | [10,81] |
| Oral Squamous Cell Carcinoma (OSCC) | Multiple genetic events involving inactivation of tumor suppressor genes | − | Deletion of the UBE4B gene | − | Tissues from OSCC patients | [82] |
| Ependymoma, Medulloblastoma | Multiple genetic events | Negative regulation of p53 by UBE4B | Protein is frequently upregulated due to the mRNA overexpression, attributed to gene amplification | Human brain cancer cell lines and mouse model | Human tissues derived from brain tumors | [46] |
| Breast Cancer | Multiple genetic events | Negative regulation of p53 by UBE4B | Protein overexpression | Human breast cancer cell lines and mouse model | Human tissues derived from breast cancer patients | [38] |
| Hepatocellular Carcinoma (HCC) | Multiple genetic events | Negative regulation of p53 by UBE4B | Overexpression of protein and mRNA levels | Human HCC cell lines | Human tissues derived from HCC patients | [56] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoniou, N.; Lagopati, N.; Balourdas, D.I.; Nikolaou, M.; Papalampros, A.; Vasileiou, P.V.S.; Myrianthopoulos, V.; Kotsinas, A.; Shiloh, Y.; Liontos, M.; et al. The Role of E3, E4 Ubiquitin Ligase (UBE4B) in Human Pathologies. Cancers 2020, 12, 62. https://doi.org/10.3390/cancers12010062
Antoniou N, Lagopati N, Balourdas DI, Nikolaou M, Papalampros A, Vasileiou PVS, Myrianthopoulos V, Kotsinas A, Shiloh Y, Liontos M, et al. The Role of E3, E4 Ubiquitin Ligase (UBE4B) in Human Pathologies. Cancers. 2020; 12(1):62. https://doi.org/10.3390/cancers12010062
Chicago/Turabian StyleAntoniou, Nikolaos, Nefeli Lagopati, Dimitrios Ilias Balourdas, Michail Nikolaou, Alexandros Papalampros, Panagiotis V. S. Vasileiou, Vassilios Myrianthopoulos, Athanassios Kotsinas, Yosef Shiloh, Michalis Liontos, and et al. 2020. "The Role of E3, E4 Ubiquitin Ligase (UBE4B) in Human Pathologies" Cancers 12, no. 1: 62. https://doi.org/10.3390/cancers12010062
APA StyleAntoniou, N., Lagopati, N., Balourdas, D. I., Nikolaou, M., Papalampros, A., Vasileiou, P. V. S., Myrianthopoulos, V., Kotsinas, A., Shiloh, Y., Liontos, M., & Gorgoulis, V. G. (2020). The Role of E3, E4 Ubiquitin Ligase (UBE4B) in Human Pathologies. Cancers, 12(1), 62. https://doi.org/10.3390/cancers12010062