Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank


Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
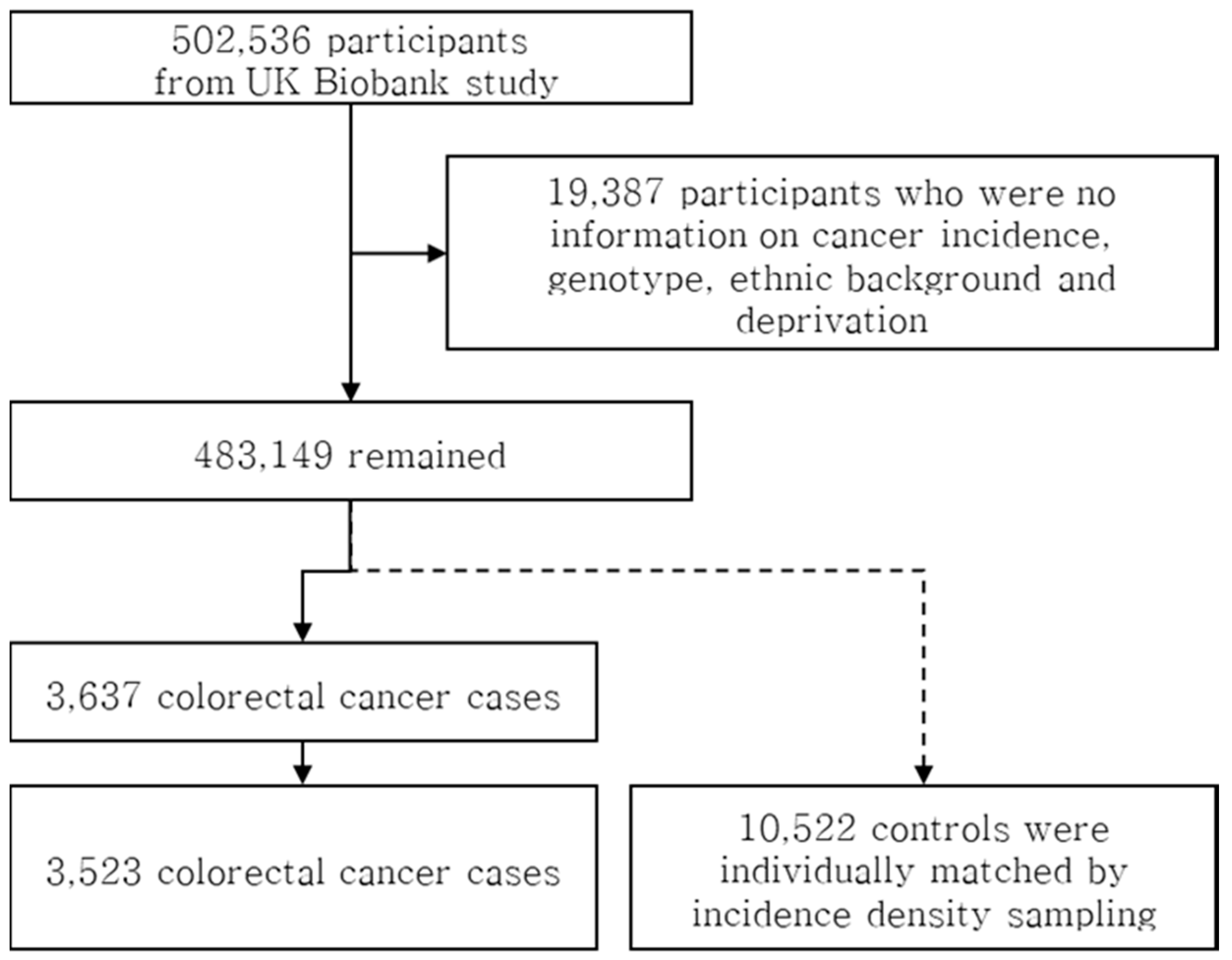
2.1. Participant Characteristics
2.2. Association of SNPs in Genes of the Citric Acid Cycle with the Risk of Colorectal Cancer
2.3. Interaction between SNPs in Genes of the Citric Acid Cycle and Contributors to Energy Balance on the Risk of Colorectal Cancer
2.4. Pairwise SNP-SNP Interactions of SNPs within the Citric Acid Cycle on the Risk of Colorectal Cancer
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Data Collection and Measurements
4.3. Outcome Ascertainment
4.4. Genotyping
4.5. Marker Selection
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Won, Y.-J.; Park, Y.R.; Jung, K.-W.; Kong, H.-J.; Lee, E.S. Cancer Statistics in Korea: Incidence, Mortality, Survival, and Prevalence in 2017. J. Korean Cancer Assoc. 2020, 52, 335–350. [Google Scholar] [CrossRef] [PubMed]
- World Cancer Research Fund, American Institute for Cancer Research Continuous Update Project Expert Report 2018: Diet, Nutrition, Physical Activity and Colorectal Cancer. 10 November 2018. Available online: https://www.wcrf.org/sites/default/files/Colorectal-cancer-report.pdf (accessed on 11 October 2020).
- Westerterp, K.R. Physical activity and physical activity induced energy expenditure in humans: Measurement, determinants, and effects. Front. Physiol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Ravussin, E.; Bogardus, C. Relationship of genetics, age, and physical fitness to daily energy expenditure and fuel utilization. Am. J. Clin. Nutr. 1989, 49, 968–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, P.S.; Bergouignan, A.; Cornier, M.-A.; Jackman, M.R. Biology’s response to dieting: The impetus for weight regain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R581–R600. [Google Scholar] [CrossRef] [Green Version]
- Koivunen, P.; Hirsilä, M.; Remes, A.M.; Hassinen, I.E.; Kivirikko, K.I.; Myllyharju, J. Inhibition of Hypoxia-inducible Factor (HIF) Hydroxylases by Citric Acid Cycle Intermediates: Possible links between cell metabolism and stabilization of hif. J. Biol. Chem. 2007, 282, 4524–4532. [Google Scholar] [CrossRef] [Green Version]
- Pezzuto, A.; Carico, E. Role of HIF-1 in Cancer Progression: Novel Insights. A Review. Curr. Mol. Med. 2018, 18, 343–351. [Google Scholar] [CrossRef]
- Pezzuto, A.; Perrone, G.; Orlando, N.; Citarella, F.; Ciccozzi, M.; Scarlata, S.; Tonini, G. A close relationship between HIF-1α; expression and bone metastases in advanced NSCLC, a retrospective analysis. Oncotarget 2019, 10, 7071–7079. [Google Scholar] [CrossRef] [Green Version]
- Cummins, T.D.; Holden, C.R.; Sansbury, B.E.; Gibb, A.A.; Shah, J.; Zafar, N.; Tang, Y.; Hellmann, J.; Rai, S.N.; Spite, M.; et al. Metabolic remodeling of white adipose tissue in obesity. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E262–E277. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Selak, M.A.; Armour, S.M.; MacKenzie, E.D.; Boulahbel, H.; Watson, D.G.; Mansfield, K.D.; Pan, Y.; Simon, M.C.; Thompson, C.B.; Gottlieb, E. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell. 2005, 7, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.; Qiu, Y.; Zou, X.; Chen, T.; Xie, G.; Cheng, Y.; Dong, T.; Zhao, L.; Feng, B.; Hu, X.; et al. Metabonomics Identifies Serum Metabolite Markers of Colorectal Cancer. J. Proteome Res. 2013, 12, 3000–3009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Xie, G.; Chen, T.; Qiu, Y.; Zou, X.; Zheng, M.; Tan, B.; Feng, B.; Dong, T.; He, P.; et al. Distinct Urinary Metabolic Profile of Human Colorectal Cancer. J. Proteome Res. 2012, 11, 1354–1363. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Posener, K.; Negelein, E. On Metabolism of Tumors; Constable: London, UK, 1930. [Google Scholar]
- Gogvadze, V.; Zhivotovsky, B.; Orrenius, S. The Warburg effect and mitochondrial stability in cancer cells. Mol. Asp. Med. 2010, 31, 60–74. [Google Scholar] [CrossRef]
- Samudio, I.; Fiegl, M.; Andreeff, M. Mitochondrial Uncoupling and the Warburg Effect: Molecular Basis for the Reprogramming of Cancer Cell Metabolism. Cancer Res. 2009, 69, 2163–2166. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.M.; Mucka, P.; Kern, J.G.; Feng, H. The emerging role and targetability of the TCA cycle in cancer metabolism. Protein Cell 2018, 9, 216–237. [Google Scholar] [CrossRef]
- Janeway, K.A.; Kim, S.Y.; Lodish, M.; Nosé, V.; Rustin, P.; Gaal, J.; Dahia, P.L.M.; Liegl, B.; Ball, E.R.; Raygada, M.; et al. Defects in succinate dehydrogenase in gastrointestinal stromal tumors lacking KIT and PDGFRA mutations. Proc. Natl. Acad. Sci. USA 2011, 108, 314–318. [Google Scholar] [CrossRef] [Green Version]
- Sajnani, K.; Islam, F.; Smith, R.A.; Gopalan, V.; Lam, A.K. Genetic alterations in Krebs cycle and its impact on cancer pathogenesis. Biochimie 2017, 135, 164–172. [Google Scholar] [CrossRef]
- Killian, J.K.; Miettinen, M.; Walker, R.L.; Wang, Y.; Zhu, Y.J.; Waterfall, J.J.; Noyes, N.; Retnakumar, P.; Yang, Z.; Smith, W.I.; et al. Recurrent epimutation of SDHC in gastrointestinal stromal tumors. Sci. Transl. Med. 2014, 6, 268ra177. [Google Scholar] [CrossRef]
- Bardella, C.; Pollard, P.J.; Tomlinson, I. SDH mutations in cancer. Biochim. Biophys. Acta-Bioenerg. 2011, 1807, 1432–1443. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Selak, M.A.; Gottlieb, E. Succinate dehydrogenase and fumarate hydratase: Linking mitochondrial dysfunction and cancer. Oncogene 2006, 25, 4675–4682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Soga, T.; Pollard, P.J. Oncometabolites: Linking altered metabolism with cancer. J. Clin. Invest. 2013, 123, 3652–3658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurenti, G.; Tennant, D.A. Isocitrate dehydrogenase (IDH), succinate dehydrogenase (SDH), fumarate hydratase (FH): Three players for one phenotype in cancer? Biochem. Soc. Trans. 2016, 44, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; He, X.; Chen, Y.; Cao, H.; Wang, J.; Liu, X.; Wang, S.; Wan, S.; Xing, J. Genetic variations in genes of metabolic enzymes predict postoperational prognosis of patients with colorectal cancer. Mol. Cancer 2015, 14, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Zhou, F.; Wang, J.; Cao, H.; Chen, Y.; Liu, X.; Zhang, Z.; Dai, J.; He, X. Functional polymorphisms of ATP citrate lyase gene predicts clinical outcome of patients with advanced colorectal cancer. World J. Surg. Oncol. 2015, 13, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limsui, D.; Vierkant, R.A.; Tillmans, L.S.; Wang, A.H.; Weisenberger, D.J.; Laird, P.W.; Lynch, C.F.; Anderson, K.E.; French, A.J.; Haile, R.W.; et al. Cigarette Smoking and Colorectal Cancer Risk by Molecularly Defined Subtypes. JNCI J. Natl. Cancer Inst. 2010, 102, 1012–1022. [Google Scholar] [CrossRef]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The UK Biobank resource with deep phenotyping and genomic data. Nature 2018, 562, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M.; et al. UK Biobank: An Open Access Resource for Identifying the Causes of a Wide Range of Complex Diseases of Middle and Old Age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef] [Green Version]
- Clayton, D.; Hills, M. Statistical Models in Epidemiology; Oxford University Press: New York, NY, USA, 1995; Volume 14, pp. 104–105. [Google Scholar]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic: Report of a WHO Consultation; WHO Technical Report Series; World Health Organization: Geneva, Switzerland, 2000; pp. 1–252. [Google Scholar]
- Townsend, P.; Phillimore, P.; Beattie, A. Health and Deprivation: Inequality and the North, 1st ed.; Routledge: London, UK, 1988. [Google Scholar]
- Møller, H.; Richards, S.; Hanchett, N.; Riaz, S.P.; Lüchtenborg, M.; Holmberg, L.; Robinson, D. Completeness of case ascertainment and survival time error in English cancer registries: Impact on 1-year survival estimates. Br. J. Cancer. 2011, 105, 170–176. [Google Scholar] [CrossRef]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar]
- Cotter, D.; Guda, P.; Fahy, E.; Subramaniam, S. MitoProteome: Mitochondrial protein sequence database and annotation system. Nucleic Acids Res. 2004, 32, D463–D467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Smigielski, E.M.; Sirotkin, K.; Ward, M.; Sherry, S.T. dbSNP: A database of single nucleotide polymorphisms. Nucleic Acids Res. 2000, 28, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Zou, G.Y. On the Estimation of Additive Interaction by Use of the Four-by-two Table and Beyond. Am. J. Epidemiol. 2008, 168, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Knol, M.J.; VanderWeele, T.J.; Groenwold, R.H.H.; Klungel, O.H.; Rovers, M.M.; Grobbee, D.E. Estimating measures of interaction on an additive scale for preventive exposures. Eur. J. Epidemiol. 2011, 26, 433–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2004, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Gene | SNP | Chr: Position | Allele (a < A) | MAF | p for HWE | Call Rate (%) | |
|---|---|---|---|---|---|---|---|
| Control | CRC Case | ||||||
| SDHC | rs16832884 | 1: 161368670 | G < A | 0.061 | 0.063 | 0.883 | 99.7 |
| SDHC | rs17395595 | 1: 161374656 | G < A | 0.148 | 0.147 | 0.788 | 99.9 |
| MDH1 | rs2278718 | 2: 63588667 | C < A | 0.249 | 0.244 | 0.365 | 99.8 |
| IDH1 | rs34218846 | 2: 208243593 | T < C | 0.056 | 0.054 | 1.000 | 99.7 |
| SUCLG2 | rs902320 | 3: 67360679 | T < C | 0.270 | 0.261 | 0.451 | 99.9 |
| SUCLG2 | rs902321 | 3: 67360742 | G < A | 0.395 | 0.389 | 0.296 | 99.8 |
| SUCLG2 | rs35494829 | 3: 67375857 | C < T | 0.113 | 0.101 | 0.829 | 99.9 |
| SUCLG2 | rs2363712 | 3: 67376176 | T < C | 0.327 | 0.317 | 0.289 | 99.9 |
| SDHA | rs6962 | 5: 256394 | A < G | 0.129 | 0.129 | 0.099 | 99.9 |
| SDHA | rs34511054 | 5: 264041 | C < A | 0.059 | 0.061 | 0.651 | 99.8 |
| ACO1 | rs7042042 | 9: 32451146 | A < G | 0.356 | 0.356 | 0.740 | 99.9 |
| ACO1 | rs10970986 | 9: 32453280 | C < T | 0.291 | 0.294 | 0.919 | 99.9 |
| OGDHL | rs11101224 | 10: 49742930 | A < G | 0.179 | 0.179 | 0.096 | 99.7 |
| OGDHL | rs751595 | 10: 49756610 | A < G | 0.191 | 0.188 | 0.395 | 99.6 |
| DLAT | rs10891314 | 11: 112045923 | A < G | 0.368 | 0.349 | 0.570 | 99.9 |
| PCK2 | rs55733026 | 14: 24095963 | G < A | 0.074 | 0.068 | 1.000 | 99.2 |
| PCK2 | rs1951634 | 14: 24100525 | T < G | 0.254 | 0.252 | 0.738 | 99.9 |
| PCK2 | rs35618680 | 14: 24103603 | A < G | 0.090 | 0.088 | 0.796 | 99.1 |
| IDH3A | rs11555541 | 15: 78149427 | C < T | 0.495 | 0.495 | 0.418 | 99.9 |
| IDH3A | rs17674205 | 15: 78169115 | G < A | 0.089 | 0.084 | 0.833 | 100.0 |
| ACLY | rs8065502 | 17: 41892360 | A < G | 0.085 | 0.085 | 0.355 | 99.6 |
| ACLY | rs2304497 | 17: 41909521 | G < T | 0.125 | 0.126 | 0.232 | 99.9 |
| Characteristics and Categories | Control, n (%) | Case, n (%) |
|---|---|---|
| N | 10,522 | 3523 |
| Age at enrollment | ||
| 36–40 | 24 (0.2) | 8 (0.2) |
| 41–45 | 272 (2.6) | 93 (2.6) |
| 46–50 | 575 (5.5) | 193 (5.5) |
| 51–55 | 1237 (11.8) | 412 (11.7) |
| 56–60 | 2007 (19.1) | 673 (19.1) |
| 61–65 | 3361 (31.9) | 1123 (31.9) |
| 66–70 | 3046 (28.9) | 1021 (29.0) |
| Sex | ||
| Men | 6052 (57.5) | 2024 (57.5) |
| Women | 4470 (42.5) | 1499 (42.5) |
| Ethnic background | ||
| White | 10,284 (97.7) | 3423 (97.2) |
| Mixed | 35 (0.3) | 18 (0.5) |
| Asian or Asian British | 83 (0.8) | 31 (0.9) |
| Black or Black British | 76 (0.7) | 28 (0.8) |
| Chinese | 11 (0.1) | 6 (0.2) |
| Other | 33 (0.3) | 17 (0.5) |
| Assessment center at which participant consented | ||
| Barts | 230 (2.2) | 76 (2.2) |
| Birmingham | 399 (3.8) | 135 (3.8) |
| Bristol | 887 (8.4) | 296 (8.4) |
| Bury | 699 (6.6) | 236 (6.7) |
| Cardiff | 453 (4.3) | 152 (4.3) |
| Croydon | 408 (3.9) | 136 (3.9) |
| Edinburgh | 445 (4.2) | 148 (4.2) |
| Glasgow | 474 (4.5) | 158 (4.5) |
| Hounslow | 472 (4.5) | 157 (4.5) |
| Leeds | 928 (8.8) | 309 (8.8) |
| Liverpool | 673 (6.4) | 226 (6.4) |
| Manchester | 339 (3.2) | 114 (3.2) |
| Middlesbrough | 353 (3.4) | 118 (3.3) |
| Newcastle | 903 (8.6) | 300 (8.5) |
| Nottingham | 704 (6.7) | 236 (6.7) |
| Oxford | 378 (3.6) | 128 (3.6) |
| Reading | 682 (6.5) | 229 (6.5) |
| Sheffield | 572 (5.4) | 193 (5.5) |
| Stockport | 12 (0.1) | 5 (0.1) |
| Stoke | 460 (4.4) | 154 (4.4) |
| Swansea | 45 (0.4) | 15 (0.4) |
| Wrexham | 6 (0.1) | 2 (0.1) |
| Townsend deprivation index at recruitment | ||
| [−6.26, 3.65] | 2788 (26.5) | 933 (26.5) |
| (−3.65, 2.15] | 2733 (26.0) | 916 (26.0) |
| (−2.15, 0.515] | 2450 (23.3) | 820 (23.3) |
| (0.515, 11] | 2551 (24.2) | 854 (24.2) |
| Gene-SNP | Colon Cancer | Rectal Cancer | ||||
|---|---|---|---|---|---|---|
| Controls, n (%) | Cases, n (%) | OR (95% CIs) | Controls, n (%) | Cases, n (%) | OR (95% CIs) | |
| SDHC-rs16832884 | ||||||
| CC | 6183 (88.0) | 2061 (87.6) | 1.00 (reference) | 3190 (88.7) | 1059 (88.0) | 1.00 (reference) |
| CT | 816 (11.6) | 285 (12.1) | 1.04 (0.90–1.20) | 393 (10.9) | 140 (11.6) | 1.08 (0.87–1.32) |
| TT | 27 (0.4) | 6 (0.3) | 0.64 (0.26–1.55) | 14 (0.4) | 4 (0.3) | 0.85 (0.28–2.59) |
| CT + TT | 1.03 (0.89–1.19) | 1.07 (0.87–1.31) | ||||
| Per T allele | 1.01 (0.88–1.16) | 1.06 (0.87–1.29) | ||||
| SDHC-rs17395595 | ||||||
| AA | 5062 (72.4) | 1680 (71.6) | 1.00 (reference) | 2616 (73.0) | 895 (74.5) | 1.00 (reference) |
| AG | 1785 (25.5) | 613 (26.1) | 1.04 (0.94–1.16) | 883 (24.6) | 280 (23.3) | 0.93 (0.80–1.08) |
| GG | 142 (2.0) | 52 (2.2) | 1.11 (0.81–1.54) | 84 (2.3) | 26 (2.2) | 0.89 (0.57–1.39) |
| AG + GG | 1.05 (0.94–1.16) | 0.93 (0.80–1.07) | ||||
| Per G allele | 1.05 (0.95–1.15) | 0.93 (0.82–1.07) | ||||
| MDH1-rs2278718 | ||||||
| GG | 3991 (56.9) | 1350 (57.4) | 1.00 (reference) | 2003 (55.7) | 688 (57.3) | 1.00 (reference) |
| GA | 2580 (36.8) | 857 (36.5) | 0.98 (0.89–1.09) | 1365 (38.0) | 435 (36.2) | 0.93 (0.81–1.07) |
| AA | 444 (6.3) | 143 (6.1) | 0.95 (0.78–1.16) | 228 (6.3) | 78 (6.5) | 1.00 (0.77–1.32) |
| GA + AA | 0.98 (0.89–1.08) | 0.94 (0.83–1.07) | ||||
| Per A allele | 0.98 (0.91–1.06) | 0.97 (0.87–1.07) | ||||
| IDH1-rs34218846 | ||||||
| GG | 6252 (89.0) | 2100 (89.3) | 1.00 (reference) | 3212 (89.4) | 1080 (89.9) | 1.00 (reference) |
| GA | 750 (10.7) | 245 (10.4) | 0.98 (0.84–1.14) | 368 (10.2) | 117 (9.7) | 0.95 (0.77–1.18) |
| AA | 19 (0.3) | 6 (0.3) | 0.93 (0.37–2.33) | 13 (0.4) | 5 (0.4) | 1.07 (0.38–3.03) |
| GA + AA | 0.97 (0.84–1.13) | 0.96 (0.77–1.18) | ||||
| Per A allele | 0.97 (0.84–1.13) | 0.96 (0.79–1.18) | ||||
| SUCLG2-rs902320 | ||||||
| GG | 3753 (53.4) | 1280 (54.5) | 1.00 (reference) | 1921 (53.5) | 650 (54.0) | 1.00 (reference) |
| GA | 2730 (38.9) | 907 (38.6) | 0.97 (0.88–1.08) | 1415 (39.4) | 476 (39.6) | 0.99 (0.87–1.14) |
| AA | 541 (7.7) | 162 (6.9) | 0.87 (0.72–1.05) | 256 (7.1) | 77 (6.4) | 0.89 (0.69–1.17) |
| GA + AA | 0.96 (0.87–1.05) | 0.98 (0.86–1.11) | ||||
| Per A allele | 0.95 (0.88–1.03) | 0.97 (0.87–1.07) | ||||
| SUCLG2-rs902321 | ||||||
| TT | 2605 (37.2) | 870 (37.1) | 1.00 (reference) | 1315 (36.6) | 439 (36.7) | 1.00 (reference) |
| TG | 3282 (46.9) | 1118 (47.7) | 1.02 (0.92–1.13) | 1701 (47.4) | 586 (49.0) | 1.03 (0.89–1.19) |
| GG | 1114 (15.9) | 358 (15.3) | 0.96 (0.84–1.11) | 572 (15.9) | 172 (14.4) | 0.90 (0.74–1.10) |
| TG + GG | 1.00 (0.91–1.10) | 1.00 (0.87–1.14) | ||||
| Per G allele | 0.99 (0.92–1.06) | 0.97 (0.88–1.06) | ||||
| SUCLG2-rs35494829 | ||||||
| CC | 5516 (78.7) | 1919 (81.9) | 1.00 (reference) | 2812 (78.7) | 945 (78.8) | 1.00 (reference) |
| CT | 1402 (20.0) | 404 (17.3) | 0.83 (0.73–0.94) | 715 (20.0) | 241 (20.1) | 1.00 (0.85–1.18) |
| TT | 87 (1.2) | 19 (0.8) | 0.64 (0.39–1.05) | 47 (1.3) | 14 (1.2) | 0.89 (0.49–1.63) |
| CT + TT | 0.82 (0.72–0.92) | 1.00 (0.85–1.17) | ||||
| Per T allele | 0.82 (0.74–0.92) | 0.99 (0.86–1.15) | ||||
| SUCLG2-rs2363712 | ||||||
| TT | 3184 (45.4) | 1087 (46.3) | 1.00 (reference) | 1643 (45.8) | 560 (46.6) | 1.00 (reference) |
| TC | 3048 (43.5) | 1018 (43.4) | 0.98 (0.89–1.08) | 1561 (43.5) | 531 (44.1) | 1.00 (0.87–1.14) |
| CC | 777 (11.1) | 242 (10.3) | 0.91 (0.77–1.07) | 386 (10.8) | 112 (9.3) | 0.86 (0.68–1.08) |
| TC + CC | 0.97 (0.88–1.06) | 0.97 (0.85–1.10) | ||||
| Per C allele | 0.96 (0.90–1.03) | 0.95 (0.86–1.05) | ||||
| SDHA-rs6962 | ||||||
| GG | 5297 (75.5) | 1787 (76.0) | 1.00 (reference) | 2726 (75.9) | 896 (74.5) | 1.00 (reference) |
| GT | 1605 (22.9) | 529 (22.5) | 0.98 (0.87–1.09) | 808 (22.5) | 294 (24.4) | 1.10 (0.94–1.28) |
| TT | 110 (1.6) | 34 (1.4) | 0.92 (0.62–1.35) | 57 (1.6) | 13 (1.1) | 0.68 (0.37–1.26) |
| GT + TT | 0.97 (0.87–1.09) | 1.08 (0.92–1.25) | ||||
| Per T allele | 0.97 (0.88–1.07) | 1.04 (0.90–1.19) | ||||
| SDHA-rs34511054 | ||||||
| GG | 6202 (88.5) | 2063 (88.0) | 1.00 (reference) | 3176 (88.6) | 1065 (88.7) | 1.00 (reference) |
| GA | 776 (11.1) | 275 (11.7) | 1.07 (0.92–1.24) | 399 (11.1) | 130 (10.8) | 0.97 (0.79–1.20) |
| AA | 31 (0.4) | 7 (0.3) | 0.67 (0.30–1.53) | 9 (0.3) | 6 (0.5) | 1.99 (0.71–5.60) |
| GA + AA | 1.05 (0.91–1.22) | 1.00 (0.81–1.22) | ||||
| Per A allele | 1.03 (0.90–1.19) | 1.02 (0.84–1.24) | ||||
| ACO1-rs7042042 | ||||||
| GG | 2898 (41.3) | 946 (40.2) | 1.00 (reference) | 1501 (41.8) | 515 (42.8) | 1.00 (reference) |
| GA | 3241 (46.2) | 1110 (47.2) | 1.05 (0.95–1.16) | 1619 (45.1) | 544 (45.2) | 0.98 (0.85–1.13) |
| AA | 880 (12.5) | 295 (12.5) | 1.02 (0.88–1.19) | 467 (13.0) | 144 (12.0) | 0.90 (0.73–1.11) |
| GA + AA | 1.04 (0.95–1.15) | 0.96 (0.84–1.10) | ||||
| Per A allele | 1.02 (0.95–1.10) | 0.96 (0.87–1.05) | ||||
| ACO1-rs10970986 | ||||||
| AA | 3561 (50.8) | 1174 (50.0) | 1.00 (reference) | 1776 (49.5) | 586 (48.7) | 1.00 (reference) |
| AG | 2850 (40.6) | 992 (42.2) | 1.06 (0.96–1.17) | 1510 (42.0) | 501 (41.6) | 1.01 (0.88–1.16) |
| GG | 603 (8.6) | 183 (7.8) | 0.93 (0.78–1.11) | 305 (8.5) | 116 (9.6) | 1.15 (0.91–1.45) |
| AG + GG | 1.04 (0.94–1.14) | 1.03 (0.91–1.18) | ||||
| Per G allele | 1.00 (0.93–1.08) | 1.05 (0.95–1.16) | ||||
| OGDHL-rs11101224 | ||||||
| AA | 4690 (66.8) | 1557 (66.2) | 1.00 (reference) | 2438 (67.9) | 825 (68.5) | 1.00 (reference) |
| AG | 2121 (30.2) | 717 (30.5) | 1.02 (0.92–1.13) | 1046 (29.1) | 351 (29.2) | 0.99 (0.86–1.15) |
| GG | 209 (3.0) | 77 (3.3) | 1.12 (0.85–1.46) | 109 (3.0) | 28 (2.3) | 0.76 (0.49–1.16) |
| AG + GG | 1.03 (0.93–1.13) | 0.97 (0.84–1.12) | ||||
| Per G allele | 1.03 (0.94–1.12) | 0.95 (0.84–1.08) | ||||
| OGDHL-rs751595 | ||||||
| TT | 4589 (65.5) | 1529 (65.2) | 1.00 (reference) | 2353 (65.5) | 798 (66.3) | 1.00 (reference) |
| TC | 2174 (31.0) | 734 (31.3) | 1.01 (0.91–1.12) | 1101 (30.7) | 378 (31.4) | 1.01 (0.88–1.17) |
| CC | 243 (3.5) | 83 (3.5) | 1.01 (0.78–1.31) | 136 (3.8) | 28 (2.3) | 0.60 (0.40–0.92) |
| TC + CC | 1.01 (0.92–1.12) | 0.97 (0.84–1.11) | ||||
| Per C allele | 1.01 (0.93–1.10) | 0.93 (0.82–1.05) | ||||
| DLAT-rs10891314 | ||||||
| GG | 2759 (39.6) | 996 (42.7) | 1.00 (reference) | 1442 (40.5) | 486 (40.8) | 1.00 (reference) |
| GA | 3231 (46.4) | 1073 (46.0) | 0.92 (0.83–1.02) | 1672 (46.9) | 557 (46.8) | 0.99 (0.86–1.14) |
| AA | 975 (14.0) | 266 (11.4) | 0.75 (0.65–0.88) | 450 (12.6) | 148 (12.4) | 0.98 (0.79–1.22) |
| GA + AA | 0.88 (0.80–0.97) | 0.99 (0.86–1.13) | ||||
| Per A allele | 0.88 (0.82–0.95) | 0.99 (0.90–1.09) | ||||
| PCK2-rs55733026 | ||||||
| AA | 6050 (86.2) | 2048 (87.1) | 1.00 (reference) | 3060 (85.1) | 1042 (86.8) | 1.00 (reference) |
| AG | 937 (13.3) | 296 (12.6) | 0.93 (0.81–1.07) | 516 (14.4) | 147 (12.2) | 0.83 (0.68–1.01) |
| GG | 35 (0.5) | 8 (0.3) | 0.68 (0.32–1.47) | 18 (0.5) | 12 (1.0) | 2.03 (0.95–4.33) |
| AG + GG | 0.92 (0.80–1.06) | 0.87 (0.72–1.05) | ||||
| Per G allele | 0.92 (0.80–1.05) | 0.92 (0.77–1.10) | ||||
| PCK2-rs1951634 | ||||||
| CC | 3918 (55.9) | 1314 (56.0) | 1.00 (reference) | 1993 (55.6) | 669 (55.7) | 1.00 (reference) |
| CT | 2652 (37.8) | 881 (37.5) | 0.99 (0.89–1.09) | 1351 (37.7) | 460 (38.3) | 1.02 (0.89–1.17) |
| TT | 444 (6.3) | 152 (6.5) | 1.01 (0.84–1.23) | 243 (6.8) | 72 (6.0) | 0.88 (0.67–1.17) |
| CT + TT | 0.99 (0.90–1.09) | 1.00 (0.87–1.14) | ||||
| Per T allele | 1.00 (0.92–1.08) | 0.98 (0.88–1.09) | ||||
| PCK2-rs35618680 | ||||||
| GG | 5798 (82.7) | 1953 (83.3) | 1.00 (reference) | 2976 (83.0) | 998 (83.2) | 1.00 (reference) |
| GA | 1156 (16.5) | 370 (15.8) | 0.95 (0.84–1.08) | 580 (16.2) | 194 (16.2) | 0.99 (0.83–1.19) |
| AA | 56 (0.8) | 21 (0.9) | 1.10 (0.67–1.83) | 31 (0.9) | 8 (0.7) | 0.76 (0.35–1.66) |
| GA + AA | 0.96 (0.85–1.09) | 0.98 (0.82–1.17) | ||||
| Per A allele | 0.97 (0.86–1.09) | 0.97 (0.83–1.14) | ||||
| IDH3A-rs11555541 | ||||||
| AA | 1823 (26.0) | 600 (25.5) | 1.00 (reference) | 903 (25.2) | 309 (25.7) | 1.00 (reference) |
| AC | 3447 (49.1) | 1178 (50.1) | 1.04 (0.93–1.16) | 1814 (50.5) | 595 (49.4) | 0.96 (0.82–1.12) |
| CC | 1749 (24.9) | 575 (24.4) | 1.00 (0.87–1.14) | 872 (24.3) | 300 (24.9) | 1.00 (0.84–1.21) |
| AC + CC | 1.03 (0.92–1.14) | 0.97 (0.84–1.13) | ||||
| Per C allele | 1.00 (0.94–1.07) | 1.00 (0.91–1.10) | ||||
| IDH3A-rs17674205 | ||||||
| TT | 5788 (83.1) | 1957 (84.0) | 1.00 (reference) | 2949 (82.7) | 1008 (84.1) | 1.00 (reference) |
| TC | 1127 (16.2) | 357 (15.3) | 0.94 (0.83–1.07) | 588 (16.5) | 180 (15.0) | 0.90 (0.75–1.08) |
| CC | 51 (0.7) | 16 (0.7) | 0.96 (0.54–1.70) | 28 (0.8) | 10 (0.8) | 1.09 (0.53–2.26) |
| TC + CC | 0.94 (0.83–1.07) | 0.91 (0.76–1.08) | ||||
| Per C allele | 0.95 (0.84–1.07) | 0.92 (0.78–1.09) | ||||
| ACLY-rs8065502 | ||||||
| AA | 5890 (83.9) | 1959 (83.3) | 1.00 (reference) | 3009 (83.8) | 1017 (84.5) | 1.00 (reference) |
| AG | 1060 (15.1) | 377 (16.0) | 1.07 (0.94–1.21) | 561 (15.6) | 180 (15.0) | 0.95 (0.79–1.14) |
| GG | 67 (1.0) | 16 (0.7) | 0.72 (0.42–1.25) | 21 (0.6) | 7 (0.6) | 0.99 (0.42–2.32) |
| AG + GG | 1.04 (0.92–1.18) | 0.95 (0.79–1.14) | ||||
| Per G allele | 1.02 (0.91–1.15) | 0.95 (0.80–1.13) | ||||
| ACLY-rs2304497 | ||||||
| GG | 5414 (77.2) | 1790 (76.2) | 1.00 (reference) | 2721 (75.7) | 923 (76.7) | 1.00 (reference) |
| GA | 1482 (21.1) | 524 (22.3) | 1.07 (0.95–1.20) | 813 (22.6) | 258 (21.4) | 0.94 (0.80–1.10) |
| AA | 121 (1.7) | 35 (1.5) | 0.88 (0.60–1.28) | 60 (1.7) | 22 (1.8) | 1.09 (0.66–1.78) |
| GA + AA | 1.05 (0.94–1.18) | 0.95 (0.82–1.11) | ||||
| Per A allele | 1.03 (0.93–1.14) | 0.97 (0.84–1.11) | ||||
| Environmental Variable | Noncarriers | Carriers | Pinteraction | ||||
|---|---|---|---|---|---|---|---|
| Controls, n (%) | Cases, n (%) | OR (95% CIs) | Controls, n (%) | Cases, n (%) | OR (95% CIs) | ||
| SDHC-rs17395595, G < A | |||||||
| Obesity, BMI | 0.0023 | ||||||
| <30 kg/m2 | 3833 (75.9) | 1163 (69.8) | 1.00 (reference) | 1416 (73.7) | 495 (74.5) | 1.00 (reference) | |
| ≥30 kg/m2 | 1216 (24.1) | 504 (30.2) | 1.42 (1.24–1.63) | 506 (26.3) | 169 (25.5) | 0.92 (0.69–1.23) | |
| MDH1-rs2278718, C < A | |||||||
| Severe obesity, BMI | 0.0229 | ||||||
| <40 kg/m2 | 3906 (98.1) | 1317 (98.4) | 1.00 (reference) | 2973 (98.6) | 973 (97.5) | 1.00 (reference) | |
| ≥40 kg/m2 | 76 (1.9) | 21 (1.6) | 1.18 (0.67–2.07) | 42 (1.4) | 25 (2.5) | 1.47 (0.76–2.84) | |
| SUCLG2-rs902320, T < C | |||||||
| Severe obesity, BMI | 0.0437 | ||||||
| <40 kg/m2 | 3672 (98.0) | 1250 (98.3) | 1.00 (reference) | 3216 (98.7) | 1039 (97.7) | 1.00 (reference) | |
| ≥40 kg/m2 | 76 (2.0) | 22 (1.7) | 0.89 (0.50–1.59) | 42 (1.3) | 24 (2.3) | 1.95 (1.00–3.81) | |
| SUCLG2-rs902321, G < A | |||||||
| Severe obesity, BMI | 0.0071 | ||||||
| <40 kg/m2 | 2544 (97.8) | 851 (98.6) | 1.00 (reference) | 4321 (98.6) | 1435 (97.7) | 1.00 (reference) | |
| ≥40 kg/m2 | 58 (2.2) | 12 (1.4) | 0.82 (0.39–1.69) | 60 (1.4) | 34 (2.3) | 1.74 (1.07–2.82) | |
| PCK2-rs55733026, G < A | |||||||
| Daily energy intake, kcal | 0.0376 | ||||||
| Men, ≤ 0.9; women, ≤ 0.85 | 340 (47.0) | 99 (43.4) | 1.00 (reference) | 61 (50.8) | 21 (52.5) | 1.00 (reference) | |
| Men, > 0.9; women, > 0.85 | 383 (53.0) | 129 (56.6) | 1.22 (0.79–1.90) | 59 (49.2) | 19 (47.5) | 0.86 (0.45–1.45) | |
| ACLY-rs2304497, G < T | |||||||
| Vigorous physical activity | 0.045 | ||||||
| Not sufficient | 1533 (54.0) | 428 (49.7) | 1.00 (reference) | 422 (50.0) | 134 (51.3) | 1.00 (reference) | |
| Sufficient | 1304 (46.0) | 434 (50.3) | 1.19 (0.98–1.46) | 422 (50.0) | 127 (48.7) | 1.44 (0.75–2.78) | |
| Environmental Variable | Noncarriers | Carriers | Pinteraction | ||||
|---|---|---|---|---|---|---|---|
| Controls, n (%) | Cases, n (%) | OR (95% CIs) | Controls, n (%) | Cases, n (%) | OR (95% CIs) | ||
| MDH1-rs2278718, C < A | |||||||
| Obesity, BMI | 0.045 | ||||||
| <30 kg/m2 | 1503 (75.3) | 528 (76.7) | 1.00 (reference) | 1227 (77.1) | 370 (72.8) | 1.00 (reference) | |
| ≥30 kg/m2 | 493 (24.7) | 160 (23.3) | 0.91 (0.72–1.15) | 364 (22.9) | 138 (27.2) | 1.39 (1.04–1.87) | |
| SUCLG2-rs902321, G < A | |||||||
| Severe obesity, BMI | 0.0468 | ||||||
| <40 kg/m2 | 1283 (97.7) | 432 (98.6) | 1.00 (reference) | 2240 (98.9) | 739 (98.0) | 1.00 (reference) | |
| ≥40 kg/m2 | 30 (2.3) | 6 (1.4) | 0.40 (0.13–1.24) | 26 (1.1) | 15 (2.0) | 1.38 (0.67–2.84) | |
| SUCLG2-rs35494829, C < T | |||||||
| Severe obesity, BMI | 0.0457 | ||||||
| <40 kg/m2 | 2766 (98.6) | 921 (98.0) | 1.00 (reference) | 742 (97.8) | 253 (99.2) | 1.00 (reference) | |
| ≥40 kg/m2 | 40 (1.4) | 19 (2.0) | 1.51 (0.83–2.75) | 17 (2.2) | 2 (0.8) | 0.39 (0.04–3.78) | |
| Abdominal obesity, WHR | 0.0159 | ||||||
| Men, ≤0.9; women, ≤0.85 | 1152 (41.0) | 333 (35.4) | 1.00 (reference) | 290 (38.1) | 107 (42.0) | 1.00 (reference) | |
| Men, >0.9; women, >0.85 | 1658 (59.0) | 609 (64.6) | 1.35 (1.12–1.63) | 471 (61.9) | 148 (58.0) | 1.11 (0.64–1.94) | |
| OGDHL-rs11101224, A < G | |||||||
| Abdominal obesity, WHR | 0.0193 | ||||||
| Men, ≤0.9; women, ≤0.85 | 1003 (41.2) | 288 (35.0) | 1.00 (reference) | 448 (38.8) | 154 (40.7) | 1.00 (reference) | |
| Men, >0.9; women, >0.85 | 1432 (58.8) | 535 (65.0) | 1.37 (1.12–1.68) | 707 (61.2) | 224 (59.3) | 1.08 (0.75–1.55) | |
| OGDHL-rs751595, G < A | |||||||
| Abdominal obesity, WHR | 0.0056 | ||||||
| Men, ≤0.9; women, ≤0.85 | 977 (41.6) | 276 (34.7) | 1.00 (reference) | 474 (38.3) | 166 (41.0) | 1.00 (reference) | |
| Men, >0.9; women, >0.85 | 1374 (58.4) | 520 (65.3) | 1.40 (1.14–1.72) | 762 (61.7) | 239 (59.0) | 0.96 (0.68–1.35) | |
| Gene-SNP | Gene-SNP | Controls, n (%) | Cases, n (%) | ORs (95% CIs) | AP (95% CIs) |
|---|---|---|---|---|---|
| SDHC-rs17395595 | IDH3A-rs11555541 | −0.348 (−0.628–0.068) | |||
| AA | AA | 1357 (19.4) | 414 (17.7) | 1.00 (reference) | |
| AG + GG | AA | 455 (6.5) | 184 (7.8) | 1.33 (1.09–1.63) | |
| AA | AC + CC | 3698 (53.0) | 1266 (54.0) | 1.12 (0.99–1.27) | |
| AG + GG | AC + CC | 1471 (21.1) | 481 (20.5) | 1.08 (0.93–1.25) | |
| MDH1-rs2278718 | SUCLG2-rs902321 | −0.301 (−0.525–0.077) | |||
| GG | TT | 1484 (21.2) | 463 (19.8) | 1.00 (reference) | |
| GA + AA | TT | 1117 (16.0) | 407 (17.4) | 1.17 (1.00–1.37) | |
| GG | TG + GG | 2492 (35.7) | 881 (37.6) | 1.13 (1.00–1.29) | |
| GA + AA | TG + GG | 1896 (27.1) | 592 (25.3) | 1.00 (0.87–1.15) | |
| IDH1-rs34218846 | IDH3A-rs11555541 | −0.507 (−0.978–0.036) | |||
| GG | AA | 1638 (23.4) | 522 (22.2) | 1.00 (reference) | |
| GA + AA | AA | 183 (2.6) | 78 (3.3) | 1.33 (1.00–1.76) | |
| GG | AC + CC | 4606 (65.7) | 1578 (67.1) | 1.07 (0.96–1.20) | |
| GA + AA | AC + CC | 586 (8.4) | 173 (7.4) | 0.93 (0.76–1.13) | |
| SUCLG2-rs902320 | IDH3A-rs17674205 | −0.570 (−0.966–0.174) | |||
| GG | TT | 3140 (45.1) | 1044 (44.9) | 1.00 (reference) | |
| GA + AA | TT | 2645 (38.0) | 909 (39.1) | 1.03 (0.93–1.15) | |
| GG | TC + CC | 579 (8.3) | 222 (9.5) | 1.17 (0.98–1.39) | |
| GA + AA | TC + CC | 599 (8.6) | 151 (6.5) | 0.76 (0.63–0.93) | |
| SUCLG2-rs902321 | OGDHL-rs751595 | −0.259 (−0.503–0.015) | |||
| TG + GG | TT | 2920 (41.8) | 948 (40.5) | 1.00 (reference) | |
| TT | TT | 1656 (23.7) | 578 (24.7) | 1.08 (0.96–1.22) | |
| TG + GG | TC + CC | 1462 (20.9) | 525 (22.4) | 1.10 (0.97–1.25) | |
| TT | TC + CC | 942 (13.5) | 288 (12.3) | 0.94 (0.81–1.09) | |
| SUCLG2-rs902321 | IDH3A-rs11555541 | −0.258 (−0.500–0.016) | |||
| TG + GG | AA | 1169 (16.7) | 359 (15.3) | 1.00 (reference) | |
| TT | AA | 648 (9.3) | 240 (10.2) | 1.20 (0.99–1.44) | |
| TG + GG | AC + CC | 3222 (46.1) | 1117 (47.6) | 1.12 (0.98–1.29) | |
| TT | AC + CC | 1954 (27.9) | 630 (26.9) | 1.05 (0.91–1.22) | |
| SUCLG2-rs902321 | IDH3A-rs17674205 | −0.491 (−0.862–0.121) | |||
| TT | TT | 2174 (31.3) | 703 (30.3) | 1.00 (reference) | |
| TG + GG | TT | 3590 (51.7) | 1250 (53.8) | 1.07 (0.96–1.19) | |
| TT | TC + CC | 407 (5.9) | 158 (6.8) | 1.21 (0.98–1.48) | |
| TG + GG | TC + CC | 769 (11.1) | 212 (9.1) | 0.86 (0.72–1.02) | |
| SUCLG2-rs35494829 | IDH3A-rs17674205 | −0.358 (−0.716–0.001) | |||
| CT + TT | TT | 1254 (18.1) | 342 (14.7) | 1.00 (reference) | |
| CC | TT | 4515 (65.0) | 1606 (69.2) | 1.31 (1.14–1.49) | |
| CT + TT | TC + CC | 223 (3.2) | 76 (3.3) | 1.26 (0.94–1.68) | |
| CC | TC + CC | 952 (13.7) | 296 (12.8) | 1.15 (0.97–1.38) | |
| SUCLG2-rs2363712 | IDH3A-rs11555541 | −0.282 (−0.508–0.055) | |||
| TC + CC | AA | 1038 (14.8) | 305 (13.0) | 1.00 (reference) | |
| TT | AA | 780 (11.1) | 295 (12.6) | 1.27 (1.06–1.53) | |
| TC + CC | AC + CC | 2784 (39.8) | 955 (40.7) | 1.16 (1.00–1.35) | |
| TT | AC + CC | 2399 (34.3) | 792 (33.7) | 1.12 (0.96–1.30) | |
| SUCLG2-rs2363712 | IDH3A-rs17674205 | −0.496 (−0.866–0.126) | |||
| TT | TT | 2651 (38.1) | 878 (37.8) | 1.00 (reference) | |
| TC + CC | TT | 3126 (45.0) | 1074 (46.2) | 1.04 (0.94–1.15) | |
| TT | TC + CC | 507 (7.3) | 196 (8.4) | 1.17 (0.98–1.41) | |
| TC + CC | TC + CC | 666 (9.6) | 176 (7.6) | 0.81 (0.68–0.97) | |
| OGDHL-rs11101224 | OGDHL-rs751595 | −0.440 (−0.857–0.022) | |||
| AG + GG | TT | 294 (4.2) | 82 (3.5) | 1.00 (reference) | |
| AA | TT | 4292 (61.3) | 1446 (61.7) | 1.20 (0.93–1.55) | |
| AG + GG | TC + CC | 2031 (29.0) | 709 (30.2) | 1.24 (0.96–1.61) | |
| AA | TC + CC | 382 (5.5) | 107 (4.6) | 1.00 (0.72–1.39) | |
| DLAT-rs10891314 | IDH3A-rs11555541 | −0.288 (−0.530–0.046) | |||
| GG | AA | 703 (10.1) | 223 (9.6) | 1.00 (reference) | |
| GA + AA | AA | 1104 (15.9) | 374 (16.0) | 1.07 (0.88–1.30) | |
| GG | AC + CC | 2051 (29.5) | 773 (33.1) | 1.19 (1.01–1.42) | |
| GA + AA | AC + CC | 3099 (44.5) | 965 (41.3) | 0.98 (0.83–1.16) |
| Gene-SNP | Gene-SNP | Controls, n (%) | Cases, n (%) | ORs (95% CIs) | AP (95% CIs) |
|---|---|---|---|---|---|
| SDHC-rs17395595 | IDH3A-rs11555541 | −0.341 (−0.672–0.010) | |||
| AG + GG | AA | 258 (7.2) | 68 (5.7) | 1.00 (reference) | |
| AA | AA | 644 (18.0) | 239 (19.9) | 1.42 (1.04–1.93) | |
| AG + GG | AC + CC | 707 (19.8) | 238 (19.8) | 1.29 (0.95–1.75) | |
| AA | AC + CC | 1966 (55.0) | 656 (54.6) | 1.28 (0.96–1.69) | |
| SUCLG2-rs902320 | SDHA-rs6962 | −0.390 (−0.774–0.006) | |||
| GA + AA | GG | 1273 (35.5) | 395 (32.9) | 1.00 (reference) | |
| GG | GG | 1449 (40.4) | 500 (41.6) | 1.12 (0.96–1.30) | |
| GA + AA | GT + TT | 392 (10.9) | 158 (13.1) | 1.30 (1.04–1.62) | |
| GG | GT + TT | 472 (13.2) | 149 (12.4) | 1.02 (0.82–1.26) | |
| SUCLG2-rs902321 | ACO1-rs7042042 | −0.431 (−0.784–0.078) | |||
| TG + GG | GG | 957 (26.7) | 301 (25.2) | 1.00 (reference) | |
| TT | GG | 541 (15.1) | 211 (17.6) | 1.25 (1.01–1.53) | |
| TG + GG | GA + AA | 1312 (36.7) | 456 (38.1) | 1.11 (0.94–1.32) | |
| TT | GA + AA | 768 (21.5) | 228 (19.1) | 0.95 (0.78–1.16) | |
| SUCLG2-rs2363712 | ACO1-rs7042042 | −0.368 (−0.681–0.054) | |||
| TC + CC | GG | 836 (23.4) | 258 (21.5) | 1.00 (reference) | |
| TT | GG | 663 (18.5) | 256 (21.3) | 1.25 (1.02–1.53) | |
| TC + CC | GA + AA | 1106 (30.9) | 385 (32.0) | 1.13 (0.94–1.35) | |
| TT | GA + AA | 975 (27.2) | 303 (25.2) | 1.01 (0.83–1.22) | |
| SDHA-rs34511054 | ACLY-rs2304497 | −0.704 (−1.362–0.047) | |||
| GA + AA | GG | 314 (8.8) | 93 (7.8) | 1.00 (reference) | |
| GG | GG | 2398 (66.9) | 830 (69.2) | 1.17 (0.92–1.49) | |
| GA + AA | GA + AA | 93 (2.6) | 43 (3.6) | 1.56 (1.02–2.40) | |
| GG | GA + AA | 777 (21.7) | 234 (19.5) | 1.01 (0.77–1.33) | |
| SDHC-rs17395595 | IDH3A-rs11555541 | −0.348 (−0.628–0.068) | |||
| AA | AA | 1357 (19.4) | 414 (17.7) | 1.00 (reference) | |
| AG + GG | AA | 455 (6.5) | 184 (7.8) | 1.33 (1.09–1.63) | |
| AA | AC + CC | 3698 (53.0) | 1266 (54.0) | 1.12 (0.99–1.27) | |
| AG + GG | AC + CC | 1471 (21.1) | 481 (20.5) | 1.08 (0.93–1.25) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.; Song, N.; Choi, J.-Y.; Shin, A. Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank. Cancers 2020, 12, 2939. https://doi.org/10.3390/cancers12102939
Cho S, Song N, Choi J-Y, Shin A. Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank. Cancers. 2020; 12(10):2939. https://doi.org/10.3390/cancers12102939
Chicago/Turabian StyleCho, Sooyoung, Nan Song, Ji-Yeob Choi, and Aesun Shin. 2020. "Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank" Cancers 12, no. 10: 2939. https://doi.org/10.3390/cancers12102939
APA StyleCho, S., Song, N., Choi, J. -Y., & Shin, A. (2020). Effect of Citric Acid Cycle Genetic Variants and Their Interactions with Obesity, Physical Activity and Energy Intake on the Risk of Colorectal Cancer: Results from a Nested Case-Control Study in the UK Biobank. Cancers, 12(10), 2939. https://doi.org/10.3390/cancers12102939