Dopamine D1 Receptor in Cancer


Abstract
:Simple Summary
Abstract
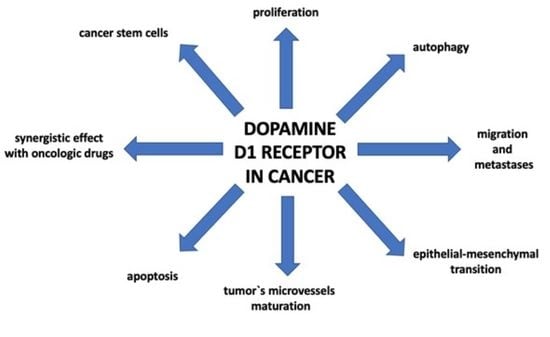
1. Introduction
2. D1 Receptor and Its Signaling Pathways
3. Effects of D1 Receptor in Various Tumor Types
3.1. Breast Cancer
3.1.1. In Vitro Studies
3.1.2. In Vivo Studies
3.1.3. Cancer Stem Cells
3.2. Nervous System Neoplasms
3.2.1. In Vitro Studies
3.2.2. In Vivo Studies
3.3. Lymphoproliferative Disorders
3.4. Lung Cancer
3.4.1. In Vitro Studies
3.4.2. In Vivo Studies
3.5. Gastrointestinal System Cancers
3.5.1. In Vitro Studies
3.5.2. In Vivo Studies
3.6. Other Tumors
4. D1 Receptor and Angiogenesis
5. Perspectives and Future Direction
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Amenta, F.; Ricci, A.; Tayebati, S.K.; Zaccheo, D. The peripheral dopaminergic system: Morphological analysis, functional and clinical applications. Ital. J. Anat. Embryol. 2002, 107, 145–167. [Google Scholar] [PubMed]
- Eisenhofer, G.; Aneman, A.; Friberg, P.; Hooper, D.; Fandriks, L.; Lonroth, H.; Hunyady, B.l.; Mezey, E. Substantial Production of Dopamine in the Human Gastrointestinal Tract. J. Clin. Endocrinol. Metab. 1997, 82, 3864–3871. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, S.; Choe, H.K.; Chung, S.; Son, G.H.; Kim, K. Implications of Circadian Rhythm in Dopamine and Mood Regulation. Mol. Cells 2017, 40, 450–456. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wise, R.A.; Baler, R. The dopamine motive system: Implications for drug and food addiction. Nat. Rev. Neurosci. 2017, 18, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, J. Dopamine: Opening the door of movement. Mov. Disord. 2018, 33, 1269. [Google Scholar] [CrossRef]
- Westbrook, A.; Braver, T.S. Dopamine Does Double Duty in Motivating Cognitive Effort. Neuron 2016, 89, 695–710. [Google Scholar] [CrossRef] [Green Version]
- Jose, P.A.; Eisner, G.M.; Felder, R.A. Regulation of blood pressure by dopamine receptors. Nephron. Physiol. 2003, 95, p19–p27. [Google Scholar] [CrossRef]
- Pinoli, M.; Marino, F.; Cosentino, M. Dopaminergic Regulation of Innate Immunity: A Review. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2017, 12, 602–623. [Google Scholar] [CrossRef]
- Peters, M.A.; Walenkamp, A.M.; Kema, I.P.; Meijer, C.; de Vries, E.G.; Oosting, S.F. Dopamine and serotonin regulate tumor behavior by affecting angiogenesis. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer Chemother. 2014, 17, 96–104. [Google Scholar] [CrossRef]
- Contreras, F.; Fouillioux, C.; Bolívar, A.; Simonovis, N.; Hernández-Hernández, R.; Armas-Hernandez, M.J.; Velasco, M. Dopamine, hypertension and obesity. J. Hum. Hypertens 2002, 16 (Suppl. 1), S13–S17. [Google Scholar] [CrossRef] [Green Version]
- Anselmi, L.; Toti, L.; Bove, C.; Hampton, J.; Travagli, R.A. A Nigro-Vagal Pathway Controls Gastric Motility and is Affected in a Rat Model of Parkinsonism. Gastroenterology 2017, 153, 1581–1593. [Google Scholar] [CrossRef]
- Kondej, M.; Stępnicki, P.; Kaczor, A.A. Multi-Target Approach for Drug Discovery against Schizophrenia. Int. J. Mol. Sci. 2018, 19, 3105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, P.M.; Pacheco, R. Targeting the Dopaminergic System in Autoimmunity. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2019. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Yu, X.; Zheng, H.; Sun, X.; Lu, Y.; Zhang, Y.; Li, C.; Bi, X. The incidence rate of cancer in patients with schizophrenia: A meta-analysis of cohort studies. Schizophr. Res. 2018, 195, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Chou, F.H.-C.; Tsai, K.-Y.; Su, C.-Y.; Lee, C.-C. The incidence and relative risk factors for developing cancer among patients with schizophrenia: A nine-year follow-up study. Schizophr. Res. 2011, 129, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Driver, J.A.; Logroscino, G.; Buring, J.E.; Gaziano, J.M.; Kurth, T. A prospective cohort study of cancer incidence following the diagnosis of Parkinson’s disease. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1260–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fois, A.F.; Wotton, C.J.; Yeates, D.; Turner, M.R.; Goldacre, M.J. Cancer in patients with motor neuron disease, multiple sclerosis and Parkinson’s disease: Record linkage studies. J. Neurol. Neurosurg. Psychiatry 2010, 81, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Rugbjerg, K.; Friis, S.; Lassen, C.F.; Ritz, B.; Olsen, J.H. Malignant melanoma, breast cancer and other cancers in patients with Parkinson’s disease. Int. J. Cancer 2012, 131, 1904–1911. [Google Scholar] [CrossRef]
- Robles, A.I.; Yang, P.; Jen, J.; McClary, A.C.; Calhoun, K.; Bowman, E.D.; Vähäkangas, K.; Greathouse, K.L.; Wang, Y.; Olivo-Marston, S.; et al. A DRD1 polymorphism predisposes to lung cancer among those exposed to secondhand smoke during childhood. Cancer Prev. Res. Phila. Pa. 2014, 7, 1210–1218. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.-L.; Wang, X.; Xing, J.-S.; Lou, J.-C.; Ma, X.-C.; Zhang, B. The potential roles of dopamine in malignant glioma. Acta Neurol. Belg. 2017, 117, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Basu, B.; Shome, S.; Jadhav, T.; Roy, S.; Majumdar, J.; Dasgupta, P.S.; Basu, S. Dopamine, by acting through its D2 receptor, inhibits insulin-like growth factor-I (IGF-I)-induced gastric cancer cell proliferation via up-regulation of Krüppel-like factor 4 through down-regulation of IGF-IR and AKT phosphorylation. Am. J. Pathol. 2010, 177, 2701–2707. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Chakroborty, D.; Chowdhury, U.R.; Dasgupta, P.S.; Basu, S. Dopamine increases the efficacy of anticancer drugs in breast and colon cancer preclinical models. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 2502–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Smith, M.; Lu, C.; Shahzad, M.M.; Pena, G.N.; Allen, J.K.; Stone, R.L.; Mangala, L.S.; Han, H.D.; Kim, H.S.; Farley, D.; et al. Dopamine blocks stress-mediated ovarian carcinoma growth. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 3649–3659. [Google Scholar] [CrossRef] [Green Version]
- Asada, M.; Ebihara, S.; Numachi, Y.; Okazaki, T.; Yamanda, S.; Ikeda, K.; Yasuda, H.; Sora, I.; Arai, H. Reduced tumor growth in a mouse model of schizophrenia, lacking the dopamine transporter. Int. J. Cancer 2008, 123, 511–518. [Google Scholar] [CrossRef]
- Teunis, M.A.; Kavelaars, A.; Voest, E.; Bakker, J.M.; Ellenbroek, B.A.; Cools, A.R.; Heijnen, C.J. Reduced tumor growth, experimental metastasis formation, and angiogenesis in rats with a hyperreactive dopaminergic system. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 1465–1467. [Google Scholar] [CrossRef] [Green Version]
- Garau, L.; Govoni, S.; Stefanini, E.; Trabucchi, M.; Spano, P.F. Dopamine receptors: Pharmacological and anatomical evidences indicate that two distinct dopamine receptor populations are present in rat striatum. Life Sci. 1978, 23, 1745–1750. [Google Scholar] [CrossRef]
- Kebabian, J.W.; Calne, D.B. Multiple receptors for dopamine. Nature 1979, 277, 93–96. [Google Scholar] [CrossRef]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Structure-function of the G protein-coupled receptor superfamily. Annu. Rev. Pharm. Toxicol. 2013, 53, 531–556. [Google Scholar] [CrossRef] [Green Version]
- Perreault, M.L.; Hasbi, A.; O’Dowd, B.F.; George, S.R. Heteromeric dopamine receptor signaling complexes: Emerging neurobiology and disease relevance. Neuropsychopharmacology 2014, 39, 156–168. [Google Scholar] [CrossRef] [Green Version]
- Perreault, M.L.; O’Dowd, B.F.; George, S.R. Dopamine D₁-D₂ receptor heteromer regulates signaling cascades involved in addiction: Potential relevance to adolescent drug susceptibility. Dev. Neurosci. 2014, 36, 287–296. [Google Scholar] [CrossRef]
- Baik, J.-H. Dopamine signaling in reward-related behaviors. Front. Neural Circuits 2013, 7, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingrich, J.A.; Caron, M.G. Recent advances in the molecular biology of dopamine receptors. Annu. Rev. Neurosci. 1993, 16, 299–321. [Google Scholar] [CrossRef]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.M.; Westlind-Danielsson, A. Dopamine receptors: Molecular biology, biochemistry and behavioural aspects. Pharmacol. Ther. 1994, 64, 291–370. [Google Scholar] [CrossRef]
- Greengard, P. The neurobiology of dopamine signaling. Biosci. Rep. 2001, 21, 247–269. [Google Scholar] [CrossRef] [PubMed]
- Stipanovich, A.; Valjent, E.; Matamales, M.; Nishi, A.; Ahn, J.-H.; Maroteaux, M.; Bertran-Gonzalez, J.; Brami-Cherrier, K.; Enslen, H.; Corbillé, A.-G.; et al. A phosphatase cascade by which rewarding stimuli control nucleosomal response. Nature 2008, 453, 879–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bibb, J.A.; Snyder, G.L.; Nishi, A.; Yan, Z.; Meijer, L.; Fienberg, A.A.; Tsai, L.H.; Kwon, Y.T.; Girault, J.A.; Czernik, A.J.; et al. Phosphorylation of DARPP-32 by Cdk5 modulates dopamine signalling in neurons. Nature 1999, 402, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Kopljar, M.; Patrlj, L.; Korolija-Marinic, D.; Horzic, M.; Cupurdija, K.; Bakota, B. High Expression of DARPP-32 in Colorectal Cancer Is Associated with Liver Metastases and Predicts Survival for Dukes A and B Patients: Results of a Pilot Study. Int. Surg. 2015, 100, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.K.; Astone, M.; Liu, P.; Hall, S.R.; Coyle, A.M.; Dankert, E.N.; Hoffman, D.K.; Zhang, W.; Kuang, R.; Roden, A.C.; et al. DARPP-32 and t-DARPP promote non-small cell lung cancer growth through regulation of IKKα-dependent cell migration. Commun. Biol. 2018, 1, 43. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Soutto, M.; Chen, Z.; Blanca Piazuelo, M.; Kay Washington, M.; Belkhiri, A.; Zaika, A.; Peng, D.; El-Rifai, W. Activation of IGF1R by DARPP-32 promotes STAT3 signaling in gastric cancer cells. Oncogene 2019, 38, 5805–5816. [Google Scholar] [CrossRef]
- Reis, H.J.; Rosa, D.V.F.; Guimarães, M.M.; Souza, B.R.; Barros, A.G.A.; Pimenta, F.J.; Souza, R.P.; Torres, K.C.L.; Romano-Silva, M.A. Is DARPP-32 a potential therapeutic target? Expert Opin. Ther. Targets 2007, 11, 1649–1661. [Google Scholar] [CrossRef]
- Li, L.; Miyamoto, M.; Ebihara, Y.; Mega, S.; Takahashi, R.; Hase, R.; Kaneko, H.; Kadoya, M.; Itoh, T.; Shichinohe, T.; et al. DRD2/DARPP-32 expression correlates with lymph node metastasis and tumor progression in patients with esophageal squamous cell carcinoma. World J. Surg. 2006, 30, 1672–1679, discussion 1680-1671. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, Y.; Miyamoto, M.; Fukunaga, A.; Kato, K.; Shichinohe, T.; Kawarada, Y.; Kurokawa, T.; Cho, Y.; Murakami, S.; Uehara, H.; et al. DARPP-32 expression arises after a phase of dysplasia in oesophageal squamous cell carcinoma. Br. J. Cancer 2004, 91, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimenta, F.J.; Horta, M.C.; Vidigal, P.V.; De Souza, B.R.; De Marco, L.; Romano-Silva, M.A.; Gomez, R.S. Decreased expression of DARPP-32 in oral premalignant and malignant lesions. Anticancer Res. 2007, 27, 2339–2343. [Google Scholar]
- Felder, R.A.; Felder, C.C.; Eisner, G.M.; Jose, P.A. The dopamine receptor in adult and maturing kidney. Am. J. Physiol. 1989, 257, F315–F327. [Google Scholar] [CrossRef]
- Jose, P.A.; Yu, P.Y.; Yamaguchi, I.; Eisner, G.M.; Mouradian, M.M.; Felder, C.C.; Felder, R.A. Dopamine D1 receptor regulation of phospholipase C. Hypertens Res. 1995, 18 (Suppl. 1), S39–S42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskoski, R., Jr. Targeting ERK1/2 protein-serine/threonine kinases in human cancers. Pharmacol. Res. 2019, 142, 151–168. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Song, T.; Li, C.; Mao, W. GSK-3β in DNA repair, apoptosis, and resistance of chemotherapy, radiotherapy of cancer. Biochim. Et Biophys. Acta. Mol. Cell Res. 2020, 1867, 118659. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Bertrand, F.E.; Davis, N.M.; Sokolosky, M.; Abrams, S.L.; Montalto, G.; D’Assoro, A.B.; Libra, M.; Nicoletti, F.; et al. GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 2014, 5, 2881–2911. [Google Scholar] [CrossRef] [Green Version]
- Windham, P.F.; Tinsley, H.N. cGMP signaling as a target for the prevention and treatment of breast cancer. Semin. Cancer Biol. 2015, 31, 106–110. [Google Scholar] [CrossRef]
- Borcherding, D.C.; Tong, W.; Hugo, E.R.; Barnard, D.F.; Fox, S.; LaSance, K.; Shaughnessy, E.; Ben-Jonathan, N. Expression and therapeutic targeting of dopamine receptor-1 (D1R) in breast cancer. Oncogene 2016, 35, 3103–3113. [Google Scholar] [CrossRef]
- Yang, L.; Yao, Y.; Yong, L.; Feng, Y.; Su, H.; Yao, Q.; Xue, J.; Lu, W.; Zhou, T. Dopamine D1 receptor agonists inhibit lung metastasis of breast cancer reducing cancer stemness. Eur. J. Pharmacol. 2019, 859, 172499. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Ochieng, J.; Evans, S.L. The growth inhibitory properties of a dopamine agonist (SKF 38393) on MCF-7 cells. Anti-Cancer Drugs 1995, 6, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Minami, K.; Liu, S.; Liu, Y.; Chen, A.; Wan, Q.; Na, S.; Li, B.Y.; Matsuura, N.; Koizumi, M.; Yin, Y.; et al. Inhibitory Effects of Dopamine Receptor D1 Agonist on Mammary Tumor and Bone Metastasis. Sci. Rep. 2017, 7, 45686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Mou, Z.; Ma, Y.; Li, J.; Li, J.; Ji, X.; Wu, K.; Li, L.; Lu, W.; Zhou, T. Dopamine enhances the response of sunitinib in the treatment of drug-resistant breast cancer: Involvement of eradicating cancer stem-like cells. Biochem. Pharmacol. 2015, 95, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Wang, S.Y.; Ren, Y.P.; Li, J.; Guo, T.J.; Lu, W.; Zhou, T.Y. Antitumor effect of axitinib combined with dopamine and PK-PD modeling in the treatment of human breast cancer xenograft. Acta Pharmacol. Sin. 2019, 40, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Kline, C.L.B.; Ralff, M.D.; Lulla, A.R.; Wagner, J.M.; Abbosh, P.H.; Dicker, D.T.; Allen, J.E.; El-Deiry, W.S. Role of Dopamine Receptors in the Anticancer Activity of ONC201. Neoplasia 2018, 20, 80–91. [Google Scholar] [CrossRef]
- Yang, K.; Wei, M.; Yang, Z.; Fu, Z.; Xu, R.; Cheng, C.; Chen, X.; Chen, S.; Dammer, E.; Le, W. Activation of dopamine receptor D1 inhibits glioblastoma tumorigenicity by regulating autophagic activity. Cell. Oncol. Dordr. 2020. [Google Scholar] [CrossRef]
- Schrell, U.M.; Fahlbusch, R.; Adams, E.F.; Nomikos, P.; Reif, M. Growth of cultured human cerebral meningiomas is inhibited by dopaminergic agents. Presence of high affinity dopamine-D1 receptors. J. Clin. Endocrinol. Metab. 1990, 71, 1669–1671. [Google Scholar] [CrossRef]
- Carroll, R.S.; Schrell, U.M.; Zhang, J.; Dashner, K.; Nomikos, P.; Fahlbusch, R.; Black, P.M. Dopamine D1, dopamine D2, and prolactin receptor messenger ribonucleic acid expression by the polymerase chain reaction in human meningiomas. Neurosurgery 1996, 38, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Ruf, C.G.; Linbecker, M.; Port, M.; Riecke, A.; Schmelz, H.U.; Wagner, W.; Meineke, V.; Abend, M. Predicting metastasized seminoma using gene expression. BJU Int. 2012, 110, E14–E20. [Google Scholar] [CrossRef]
- Gray, J.M.; Ziemian, L. Antiestrogen binding sites in brain and pituitary of ovariectomized rats. Brain Res. 1992, 578, 55–60. [Google Scholar] [CrossRef]
- Watts, C.K.; Sutherland, R.L. Studies on the ligand specificity and potential identity of microsomal antiestrogen-binding sites. Mol. Pharmacol. 1987, 31, 541–551. [Google Scholar] [PubMed]
- Martin, Y.C. The Discovery of Novel Selective D1 Dopaminergic Agonists: A-68930, A-77636, A-86929, and ABT-413. Int. J. Med. Chem. 2011, 2011, 424535. [Google Scholar] [CrossRef] [PubMed]
- Neve, K.A.; Seamans, J.K.; Trantham-Davidson, H. Dopamine receptor signaling. J. Recept. Signal Transduct. Res. 2004, 24, 165–205. [Google Scholar] [CrossRef] [PubMed]
- Bibb, J.A. Decoding dopamine signaling. Cell 2005, 122, 153–155. [Google Scholar] [CrossRef] [Green Version]
- Zaccolo, M.; Movsesian, M.A. cAMP and cGMP signaling cross-talk: Role of phosphodiesterases and implications for cardiac pathophysiology. Circ. Res. 2007, 100, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, D.; Han, Y.; Wang, W.; Wang, N.; Yang, J.; Zeng, C. Dopamine D1-Like Receptors Suppress the Proliferation of Macrophages Induced by Ox-LDL. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 38, 415–426. [Google Scholar] [CrossRef]
- Huang, C.; Wu, J.; Liao, R.; Zhang, W. SKF83959, an agonist of phosphatidylinositol-linked D(1)-like receptors, promotes ERK1/2 activation and cell migration in cultured rat astrocytes. PLoS ONE 2012, 7, e49954. [Google Scholar] [CrossRef]
- Chen, J.; Rusnak, M.; Luedtke, R.R.; Sidhu, A. D1 dopamine receptor mediates dopamine-induced cytotoxicity via the ERK signal cascade. J. Biol. Chem. 2004, 279, 39317–39330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seong, S.B.; Ha, D.S.; Min, S.Y.; Ha, T.S. Autophagy Precedes Apoptosis in Angiotensin II-Induced Podocyte Injury. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2019, 53, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Clemens, M.J. Initiation factor eIF2 alpha phosphorylation in stress responses and apoptosis. Prog. Mol. Subcell. Biol. 2001, 27, 57–89. [Google Scholar]
- Hamamura, K.; Tanjung, N.; Yokota, H. Suppression of osteoclastogenesis through phosphorylation of eukaryotic translation initiation factor 2 alpha. J. Bone Miner. Metab. 2013, 31, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Yue, K.; Chen, L.; Tian, X.; Ru, Q.; Gan, Y.; Wang, D.; Jin, G.; Li, C. L-stepholidine, a natural dopamine receptor D1 agonist and D2 antagonist, inhibits heroin-induced reinstatement. Neurosci. Lett. 2014, 559, 67–71. [Google Scholar] [CrossRef]
- Hartmann, J.T.; Haap, M.; Kopp, H.G.; Lipp, H.P. Tyrosine kinase inhibitors—A review on pharmacology, metabolism and side effects. Curr. Drug Metab. 2009, 10, 470–481. [Google Scholar] [CrossRef]
- Elgebaly, A.; Menshawy, A.; El Ashal, G.; Osama, O.; Ghanem, E.; Omar, A.; Negida, A. Sunitinib alone or in combination with chemotherapy for the treatment of advanced breast cancer: A systematic review and meta-analysis. Breast Dis. 2016, 36, 91–101. [Google Scholar] [CrossRef]
- Joosten, S.C.; Hamming, L.; Soetekouw, P.M.; Aarts, M.J.; Veeck, J.; van Engeland, M.; Tjan-Heijnen, V.C. Resistance to sunitinib in renal cell carcinoma: From molecular mechanisms to predictive markers and future perspectives. Biochim. Et. Biophys. Acta 2015, 1855, 1–16. [Google Scholar] [CrossRef]
- Sierra, J.R.; Cepero, V.; Giordano, S. Molecular mechanisms of acquired resistance to tyrosine kinase targeted therapy. Mol. Cancer 2010, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and roles in cancer cell biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.F.; Hoffmann, E.K.; Novak, I. Cell volume regulation in epithelial physiology and cancer. Front. Physiol. 2013, 4, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, J.-M.; Rouzaire-Dubois, B. Roles of cell volume in molecular and cellular biology. Prog. Biophys. Mol. Biol. 2012, 108, 93–97. [Google Scholar] [CrossRef]
- He, W.; Li, H.; Min, X.; Liu, J.; Hu, B.; Hou, S.; Wang, J. Activation of volume-sensitive Cl(-) channel is involved in carboplatin-induced apoptosis in human lung adenocarcinoma cells. Cancer Biol. Ther. 2010, 9, 885–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Rycaj, K.; Chen, X.; Tang, D.G. Cancer stem cells and cell size: A causal link? Semin. Cancer Biol. 2015, 35, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.L.; Shimizu, T.; Ise, T.; Numata, T.; Kohno, K.; Okada, Y. Impaired activity of volume-sensitive Cl− channel is involved in cisplatin resistance of cancer cells. J. Cell. Physiol. 2007, 211, 513–521. [Google Scholar] [CrossRef]
- Morishita, K.; Watanabe, K.; Ichijo, H. Cell volume regulation in cancer cell migration driven by osmotic water flow. Cancer Sci. 2019, 110, 2337–2347. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chai, N.; Sha, B.; Guo, M.; Zhuang, J.; Xu, F.; Li, F. The effect of substrate stiffness on cancer cell volume homeostasis. J. Cell. Physiol. 2018, 233, 1414–1423. [Google Scholar] [CrossRef]
- Widler, L.; Jaeggi, K.A.; Glatt, M.; Muller, K.; Bachmann, R.; Bisping, M.; Born, A.R.; Cortesi, R.; Guiglia, G.; Jeker, H.; et al. Highly potent geminal bisphosphonates. From pamidronate disodium (Aredia) to zoledronic acid (Zometa). J. Med. Chem. 2002, 45, 3721–3738. [Google Scholar] [CrossRef]
- Onder, T.T.; Gupta, P.B.; Mani, S.A.; Yang, J.; Lander, E.S.; Weinberg, R.A. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008, 68, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.K.; Matchett, K.B.; McEnhill, P.M.; Dakir el, H.; McMullin, M.F.; El-Tanani, Y.; Patterson, L.; Faheem, A.; Rudland, P.S.; McCarron, P.A.; et al. Protein deregulation associated with breast cancer metastasis. Cytokine Growth Factor Rev. 2015, 26, 415–423. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X.; Chen, G.; Dong, C.; Zhang, D. The impact of matrix metalloproteinase 2 on prognosis and clinicopathology of breast cancer patients: A systematic meta-analysis. PLoS ONE 2015, 10, e0121404. [Google Scholar] [CrossRef] [PubMed]
- DuPre, S.A.; Hunter, K.W., Jr. Murine mammary carcinoma 4T1 induces a leukemoid reaction with splenomegaly: Association with tumor-derived growth factors. Exp. Mol. Pathol. 2007, 82, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Kowanetz, M.; Wu, X.; Lee, J.; Tan, M.; Hagenbeek, T.; Qu, X.; Yu, L.; Ross, J.; Korsisaari, N.; Cao, T.; et al. Granulocyte-colony stimulating factor promotes lung metastasis through mobilization of Ly6G+Ly6C+ granulocytes. Proc. Natl. Acad. Sci. USA 2010, 107, 21248–21255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhu, X.; Han, M.; Hao, F.; Lu, W.; Zhou, T. Mechanistic Pharmacokinetic/Pharmacodynamic Model of Sunitinib and Dopamine in MCF-7/Adr Xenografts: Linking Cellular Heterogeneity to Tumour Burden. AAPS J. 2020, 22, 45. [Google Scholar] [CrossRef]
- Chen, R.C.; Lin, N.U.; Golshan, M.; Harris, J.R.; Bellon, J.R. Internal mammary nodes in breast cancer: Diagnosis and implications for patient management—A systematic review. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2008, 26, 4981–4989. [Google Scholar] [CrossRef] [PubMed]
- Rukanskiene, D.; Veikutis, V.; Jonaitiene, E.; Baseviciute, M.; Kunigiskis, D.; Paukstaitiene, R.; Cepuliene, D.; Poskiene, L.; Bogusevicius, A. Preoperative Axillary Ultrasound versus Sentinel Lymph Node Biopsy in Patients with Early Breast Cancer. Med. KaunasLith. 2020, 56, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paula, A.C.; Lopes, C. Implications of Different Cancer Stem Cell Phenotypes in Breast Cancer. Anticancer Res. 2017, 37, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Raha, D.; Wilson, T.R.; Peng, J.; Peterson, D.; Yue, P.; Evangelista, M.; Wilson, C.; Merchant, M.; Settleman, J. The cancer stem cell marker aldehyde dehydrogenase is required to maintain a drug-tolerant tumor cell subpopulation. Cancer Res. 2014, 74, 3579–3590. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wersinger, C.; Sidhu, A. Chronic stimulation of D1 dopamine receptors in human SK-N-MC neuroblastoma cells induces nitric-oxide synthase activation and cytotoxicity. J. Biol. Chem. 2003, 278, 28089–28100. [Google Scholar] [CrossRef] [Green Version]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Bossi, O.; Gartsbein, M.; Leitges, M.; Kuroki, T.; Grossman, S.; Tennenbaum, T. UV irradiation increases ROS production via PKCdelta signaling in primary murine fibroblasts. J. Cell. Biochem. 2008, 105, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Uryu, K.; Wang, H.Y.; Friedman, E. D1 dopamine receptor agonists mediate activation of p38 mitogen-activated protein kinase and c-Jun amino-terminal kinase by a protein kinase A-dependent mechanism in SK-N-MC human neuroblastoma cells. Mol. Pharmacol. 1998, 54, 453–458. [Google Scholar] [CrossRef]
- Yan, J.; Yang, R. Dopamine receptor D1 promotes the proliferation, invasion and migration of gliomas by inhibiting cAMP signaling pathway. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi Chin. J. Cell. Mol. Immunol. 2018, 34, 1116–1121. [Google Scholar]
- Chi, A.S.; Tarapore, R.S.; Hall, M.D.; Shonka, N.; Gardner, S.; Umemura, Y.; Sumrall, A.; Khatib, Z.; Mueller, S.; Kline, C.; et al. Pediatric and adult H3 K27M-mutant diffuse midline glioma treated with the selective DRD2 antagonist ONC201. J. Neuro Oncol. 2019, 145, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Mondal, A.C.; Majumder, J.; Basu, S.; Dasgupta, P.S. Physiological concentrations of dopamine inhibit the proliferation and cytotoxicity of human CD4+ and CD8+ T cells in vitro: A receptor-mediated mechanism. Neuroimmunomodulation 2001, 9, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.C.; Mondal, A.C.; Basu, S.; Banerjee, S.; Majumder, J.; Bhattacharya, D.; Dasgupta, P.S. Dopamine inhibits cytokine release and expression of tyrosine kinases, Lck and Fyn in activated T cells. Int. Immunopharmacol. 2003, 3, 1019–1026. [Google Scholar] [CrossRef]
- Basu, B.; Sarkar, C.; Chakroborty, D.; Ganguly, S.; Shome, S.; Dasgupta, P.S.; Basu, S. D1 and D2 dopamine receptor-mediated inhibition of activated normal T cell proliferation is lost in jurkat T leukemic cells. J. Biol. Chem. 2010, 285, 27026–27032. [Google Scholar] [CrossRef] [Green Version]
- Epstein, P.M.; Mills, J.S.; Ross, C.P.; Strada, S.J.; Hersh, E.M.; Thompson, W.J. Increased cyclic nucleotide phosphodiesterase activity associated with proliferation and cancer in human and murine lymphoid cells. Cancer Res. 1977, 37, 4016–4023. [Google Scholar]
- Ekholm, D.; Mulloy, J.C.; Gao, G.; Degerman, E.; Franchini, G.; Manganiello, V.C. Cyclic nucleotide phosphodiesterases (PDE) 3 and 4 in normal, malignant, and HTLV-I transformed human lymphocytes. Biochem. Pharmacol. 1999, 58, 935–950. [Google Scholar] [CrossRef]
- Sachlos, E.; Risueno, R.M.; Laronde, S.; Shapovalova, Z.; Lee, J.H.; Russell, J.; Malig, M.; McNicol, J.D.; Fiebig-Comyn, A.; Graham, M.; et al. Identification of drugs including a dopamine receptor antagonist that selectively target cancer stem cells. Cell 2012, 149, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Yuan, L.B. Dopamine inhibits proliferation, induces differentiation and apoptosis of K562 leukaemia cells. Chin. Med J. 2007, 120, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Scozzi, D.; Mariotta, S.; Cherubini, E.; Bruno, P.; Tabbì, L.; Fioretti, F.; Sciannamea, A.; Piraino, A.; Pietrangeli, V.; Ricci, A. Increased levels of plasmatic dopamine in human small cell lung cancer. Eur. Respir. J. 2012, 40, P4218. [Google Scholar]
- Cherubini, E.; Di Napoli, A.; Noto, A.; Osman, G.A.; Esposito, M.C.; Mariotta, S.; Sellitri, R.; Ruco, L.; Cardillo, G.; Ciliberto, G.; et al. Genetic and Functional Analysis of Polymorphisms in the Human Dopamine Receptor and Transporter Genes in Small Cell Lung Cancer. J. Cell. Physiol. 2016, 231, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Marcellino, D.; Rivera, A.; Diaz-Cabiale, Z.; Filip, M.; Gago, B.; Roberts, D.C.; Langel, U.; Genedani, S.; Ferraro, L.; et al. Receptor-receptor interactions within receptor mosaics. Impact on neuropsychopharmacology. Brain Res. Rev. 2008, 58, 415–452. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Wang, S.; Zhu, X.; Xue, J.; Li, J.; Wang, L.; Li, J.; Lu, W.; Zhou, T. Pharmacokinetic-Pharmacodynamic Modeling of the Anti-Tumor Effect of Sunitinib Combined with Dopamine in the Human Non-Small Cell Lung Cancer Xenograft. Pharm. Res. 2017, 34, 408–418. [Google Scholar] [CrossRef]
- Su, H.; Xue, Z.; Feng, Y.; Xie, Y.; Deng, B.; Yao, Y.; Tian, X.; An, Q.; Yang, L.; Yao, Q.; et al. N-arylpiperazine-containing compound (C2): An enhancer of sunitinib in the treatment of pancreatic cancer, involving D1DR activation. Toxicol. Appl. Pharmacol. 2019, 384, 114789. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, C.; Gao, F.; Li, H. The effect and mechanism of dopamine D1 receptors on the proliferation of osteosarcoma cells. Mol. Cell. Biochem. 2017, 430, 31–36. [Google Scholar] [CrossRef]
- Martini, M.; De Santis, M.C.; Braccini, L.; Gulluni, F.; Hirsch, E. PI3K/AKT signaling pathway and cancer: An updated review. Ann. Med. 2014, 46, 372–383. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Gao, J.; Gao, F. Dopamine D1 receptors induce apoptosis of osteosarcoma cells via changes of MAPK pathway. Clin. Exp. Pharmacol. Physiol. 2017, 44, 1166–1168. [Google Scholar] [CrossRef]
- Ramjiawan, R.R.; Griffioen, A.W.; Duda, D.G. Anti-angiogenesis for cancer revisited: Is there a role for combinations with immunotherapy? Angiogenesis 2017, 20, 185–204. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis Primers 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Nagy, J.A.; Pal, S.; Vasile, E.; Eckelhoefer, I.A.; Bliss, V.S.; Manseau, E.J.; Dasgupta, P.S.; Dvorak, H.F.; Mukhopadhyay, D. The neurotransmitter dopamine inhibits angiogenesis induced by vascular permeability factor/vascular endothelial growth factor. Nat. Med. 2001, 7, 569–574. [Google Scholar] [CrossRef]
- Chakroborty, D.; Chowdhury, U.R.; Sarkar, C.; Baral, R.; Dasgupta, P.S.; Basu, S. Dopamine regulates endothelial progenitor cell mobilization from mouse bone marrow in tumor vascularization. J. Clin. Investig. 2008, 118, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Sarkar, C.; Chakroborty, D.; Nagy, J.; Mitra, R.B.; Dasgupta, P.S.; Mukhopadhyay, D. Ablation of peripheral dopaminergic nerves stimulates malignant tumor growth by inducing vascular permeability factor/vascular endothelial growth factor-mediated angiogenesis. Cancer Res. 2004, 64, 5551–5555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Smith, M.; Lee, S.J.; Lu, C.; Nagaraja, A.S.; He, G.; Rupaimoole, R.; Han, H.D.; Jennings, N.B.; Roh, J.W.; Nishimura, M.; et al. Biologic effects of dopamine on tumor vasculature in ovarian carcinoma. Neoplasia 2013, 15, 502–510. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Kraft, K. Beta-endorphin and catecholamine concentrations during chronic and acute stress in intensive care patients. Eur J. Med. Res. 1996, 1, 528–532. [Google Scholar]
- Lutgendorf, S.K.; DeGeest, K.; Sung, C.Y.; Arevalo, J.M.; Penedo, F.; Lucci, J., 3rd; Goodheart, M.; Lubaroff, D.; Farley, D.M.; Sood, A.K.; et al. Depression, social support, and beta-adrenergic transcription control in human ovarian cancer. BrainBehav. Immun. 2009, 23, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Thaker, P.H.; Han, L.Y.; Kamat, A.A.; Arevalo, J.M.; Takahashi, R.; Lu, C.; Jennings, N.B.; Armaiz-Pena, G.; Bankson, J.A.; Ravoori, M.; et al. Chronic stress promotes tumor growth and angiogenesis in a mouse model of ovarian carcinoma. Nat. Med. 2006, 12, 939–944. [Google Scholar] [CrossRef]
- Jain, R.K. Normalization of tumor vasculature: An emerging concept in antiangiogenic therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef]
- Winkler, F.; Kozin, S.V.; Tong, R.T.; Chae, S.S.; Booth, M.F.; Garkavtsev, I.; Xu, L.; Hicklin, D.J.; Fukumura, D.; di Tomaso, E.; et al. Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response to radiation: Role of oxygenation, angiopoietin-1, and matrix metalloproteinases. Cancer Cell 2004, 6, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian, X.; Hakansson, J.; Stahlberg, A.; Lindblom, P.; Betsholtz, C.; Gerhardt, H.; Semb, H. Pericytes limit tumor cell metastasis. J. Clin. Investig. 2006, 116, 642–651. [Google Scholar] [CrossRef] [Green Version]
- Felsing, D.E.; Jain, M.K.; Allen, J.A. Advances in Dopamine D1 Receptor Ligands for Neurotherapeutics. Curr. Top. Med. Chem. 2019, 19, 1365–1380. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Holmes, C. Neuronal source of plasma dopamine. Clin. Chem. 2008, 54, 1864–1871. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, D.S.; Swoboda, K.J.; Miles, J.M.; Coppack, S.W.; Aneman, A.; Holmes, C.; Lamensdorf, I.; Eisenhofer, G. Sources and physiological significance of plasma dopamine sulfate. J. Clin. Endocrinol. Metab. 1999, 84, 2523–2531. [Google Scholar] [CrossRef]
- Kovacs, Z.; Jung, I.; Gurzu, S. Arylsulfatases A and B: From normal tissues to malignant tumors. Pathol. Res. Pract. 2019, 215, 152516. [Google Scholar] [CrossRef]
- Laidler, P.; Kowalski, D.; Silberring, J. Arylsulfatase A in serum from patients with cancer of various organs. Clin. Chim. Acta Int. J. Clin. Chem. 1991, 204, 69–77. [Google Scholar] [CrossRef]
- Van den Berghe, G.; de Zegher, F.; Wouters, P.; Schetz, M.; Verwaest, C.; Ferdinande, P.; Lauwers, P. Dehydroepiandrosterone sulphate in critical illness: Effect of dopamine. Clin. Endocrinol. 1995, 43, 457–463. [Google Scholar] [CrossRef]
- Schnuelle, P.; Schmitt, W.H.; Weiss, C.; Habicht, A.; Renders, L.; Zeier, M.; Drüschler, F.; Heller, K.; Pisarski, P.; Banas, B.; et al. Effects of Dopamine Donor Pretreatment on Graft Survival after Kidney Transplantation: A Randomized Trial. Clin. J. Am. Soc. Nephrol. 2017, 12, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef]
- Kouvelas, D.; Goulas, A.; Papazisis, G.; Sardeli, C.; Pourzitaki, C. PDE5 inhibitors: In vitro and in vivo pharmacological profile. Curr. Pharm. Des. 2009, 15, 3464–3475. [Google Scholar] [CrossRef]
- Chen, W.; Qin, Y.; Liu, S. Cytokines, breast cancer stem cells (BCSCs) and chemoresistance. Clin. Transl. Med. 2018, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Fahrmann, J.F.; Lee, H.; Li, Y.J.; Tripathi, S.C.; Yue, C.; Zhang, C.; Lifshitz, V.; Song, J.; Yuan, Y.; et al. JAK/STAT3-Regulated Fatty Acid β-Oxidation Is Critical for Breast Cancer Stem Cell Self-Renewal and Chemoresistance. Cell Metab. 2018, 27, 136e135–150e135. [Google Scholar] [CrossRef] [Green Version]
- Flaim, K.E.; Gessner, G.W.; Crooke, S.T.; Sarau, H.M.; Weinstock, J. Binding of a novel dopaminergic agonist radioligand [3H]-fenoldopam (SKF 82526) to D-1 receptors in rat striatum. Life Sci. 1985, 36, 1427–1436. [Google Scholar] [CrossRef]
- Ng, S.S.; Pang, C.C. In vivo venodilator action of fenoldopam, a dopamine D(1)-receptor agonist. Br. J. Pharmacol. 2000, 129, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Weber, R.R.; McCoy, C.E.; Ziemniak, J.A.; Frederickson, E.D.; Goldberg, L.I.; Murphy, M.B. Pharmacokinetic and pharmacodynamic properties of intravenous fenoldopam, a dopamine1-receptor agonist, in hypertensive patients. Br. J. Clin. Pharm. 1988, 25, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Vedachalam, S.; Luo, D.; Antonio, T.; Reith, M.E.; Dutta, A.K. Development of a Highly Potent D2/D3 Agonist and a Partial Agonist from Structure-Activity Relationship Study of N(6)-(2-(4-(1H-Indol-5-yl)piperazin-1-yl)ethyl)-N(6)-propyl-4,5,6,7-tetrahydrobenzo[d]thiazole-2,6-diamine Analogues: Implication in the Treatment of Parkinson’s Disease. J. Med. Chem. 2015, 58, 9179–9195. [Google Scholar] [CrossRef]
- Chen, X.; McCorvy, J.D.; Fischer, M.G.; Butler, K.V.; Shen, Y.; Roth, B.L.; Jin, J. Discovery of G Protein-Biased D2 Dopamine Receptor Partial Agonists. J. Med. Chem. 2016, 59, 10601–10618. [Google Scholar] [CrossRef] [Green Version]
- Sohur, U.S.; Gray, D.L.; Duvvuri, S.; Zhang, Y.; Thayer, K.; Feng, G. Phase 1 Parkinson’s Disease Studies Show the Dopamine D1/D5 Agonist PF-06649751 is Safe and Well Tolerated. Neurol. Ther. 2018, 7, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, L.E.; Smyk-Randall, E.M.; Sibley, D.R. Cyclic AMP-mediated desensitization of D1 dopamine receptor-coupled adenylyl cyclase in NS20Y neuroblastoma cells. Mol. Cell Neurosci. 1994, 5, 567–575. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Premont, R.T.; Bohn, L.M.; Lefkowitz, R.J.; Caron, M.G. Desensitization of G protein-coupled receptors and neuronal functions. Annu. Rev. Neurosci. 2004, 27, 107–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazu, S.; Shimatsu, A.; Yamada, S.; Inoshita, N.; Nagamura, Y.; Usui, T.; Tsukada, T. Resistance to dopamine agonists in prolactinoma is correlated with reduction of dopamine D2 receptor long isoform mRNA levels. Eur. J. Endocrinol. 2012, 166, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse models of metastasis: Progress and prospects. Dis. Models Mech. 2017, 10, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Abbaszadegan, M.R.; Bagheri, V.; Razavi, M.S.; Momtazi, A.A.; Sahebkar, A.; Gholamin, M. Isolation, identification, and characterization of cancer stem cells: A review. J. Cell. Physiol. 2017, 232, 2008–2018. [Google Scholar] [CrossRef]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Lewis, M.T.; Huang, J.; Gutierrez, C.; Osborne, C.K.; Wu, M.F.; Hilsenbeck, S.G.; Pavlick, A.; Zhang, X.; Chamness, G.C.; et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J. Natl. Cancer Inst. 2008, 100, 672–679. [Google Scholar] [CrossRef]
- Conley, S.J.; Gheordunescu, E.; Kakarala, P.; Newman, B.; Korkaya, H.; Heath, A.N.; Clouthier, S.G.; Wicha, M.S. Antiangiogenic agents increase breast cancer stem cells via the generation of tumor hypoxia. Proc. Natl. Acad. Sci. USA 2012, 109, 2784–2789. [Google Scholar] [CrossRef] [Green Version]
- Chinchar, E.; Makey, K.L.; Gibson, J.; Chen, F.; Cole, S.A.; Megason, G.C.; Vijayakumar, S.; Miele, L.; Gu, J.W. Sunitinib significantly suppresses the proliferation, migration, apoptosis resistance, tumor angiogenesis and growth of triple-negative breast cancers but increases breast cancer stem cells. Vasc. Cell 2014, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, C.; Chakroborty, D.; Basu, S. Neurotransmitters as regulators of tumor angiogenesis and immunity: The role of catecholamines. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2013, 8, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, Q.; Liao, Q.; Zhao, Y. Potential Roles of Peripheral Dopamine in Tumor Immunity. J. Cancer 2017, 8, 2966–2973. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, R.; Tang, N.; Gong, Z.; Zhou, J.; Chen, Y.; Chen, K.; Cai, W. Dopamine inhibits the function of Gr-1+CD115+ myeloid-derived suppressor cells through D1-like receptors and enhances anti-tumor immunity. J. Leukoc. Biol. 2015, 97, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Pan, P.-Y.; Li, Q.; Sato, A.I.; Levy, D.E.; Bromberg, J.; Divino, C.M.; Chen, S.-H. Gr-1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006, 66, 1123–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, V.; Hu, X.; Weber, R.; Nagibin, V.; Groth, C.; Altevogt, P.; Utikal, J.; Umansky, V. Targeting Myeloid-Derived Suppressor Cells to Bypass Tumor-Induced Immunosuppression. Front. Immunol. 2018, 9, 398. [Google Scholar] [CrossRef]
- Saha, B.; Mondal, A.C.; Basu, S.; Dasgupta, P.S. Circulating dopamine level, in lung carcinoma patients, inhibits proliferation and cytotoxicity of CD4+ and CD8+ T cells by D1 dopamine receptors: An in vitro analysis. Int. Immunopharmacol. 2001, 1, 1363–1374. [Google Scholar] [CrossRef]
- Ng, W.; Kennar, R.; Uetrecht, J. Effect of clozapine and olanzapine on neutrophil kinetics: Implications for drug-induced agranulocytosis. Chem. Res. Toxicol. 2014, 27, 1104–1108. [Google Scholar] [CrossRef]
- Pollmächer, T.; Haack, M.; Schuld, A.; Kraus, T.; Hinze-Selch, D. Effects of antipsychotic drugs on cytokine networks. J. Psychiatr. Res. 2000, 34, 369–382. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | D1R Expression | Clinical Effects | Ref. |
|---|---|---|---|
| breast cancer |
|
| [52,53,54,55,56,57] |
| glioblastoma |
|
| [58,59] |
| meningioma |
|
| [60,61] |
| non-small cell lung carcinoma | NA |
| [19] |
| seminoma | − mRNA (patients’ samples)—upregulation |
| [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobczuk, P.; Łomiak, M.; Cudnoch-Jędrzejewska, A. Dopamine D1 Receptor in Cancer. Cancers 2020, 12, 3232. https://doi.org/10.3390/cancers12113232
Sobczuk P, Łomiak M, Cudnoch-Jędrzejewska A. Dopamine D1 Receptor in Cancer. Cancers. 2020; 12(11):3232. https://doi.org/10.3390/cancers12113232
Chicago/Turabian StyleSobczuk, Paweł, Michał Łomiak, and Agnieszka Cudnoch-Jędrzejewska. 2020. "Dopamine D1 Receptor in Cancer" Cancers 12, no. 11: 3232. https://doi.org/10.3390/cancers12113232
APA StyleSobczuk, P., Łomiak, M., & Cudnoch-Jędrzejewska, A. (2020). Dopamine D1 Receptor in Cancer. Cancers, 12(11), 3232. https://doi.org/10.3390/cancers12113232