The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers


 ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Narrative Review
1.1. Introduction
1.2. Circular RNAs
1.3. Characteristics
1.4. Biogenesis and Degradation
1.5. Biological Functions
1.5.1. Competing Endogenous RNA Network
1.5.2. Interaction with RNA-Binding Proteins (RBPs)
1.5.3. Protein and Peptide Regulation
1.5.4. Pseudogene Generation
1.6. CircRNA Research Techniques
1.6.1. CircRNA Sequencing and Profiling
1.6.2. Bioinformatics
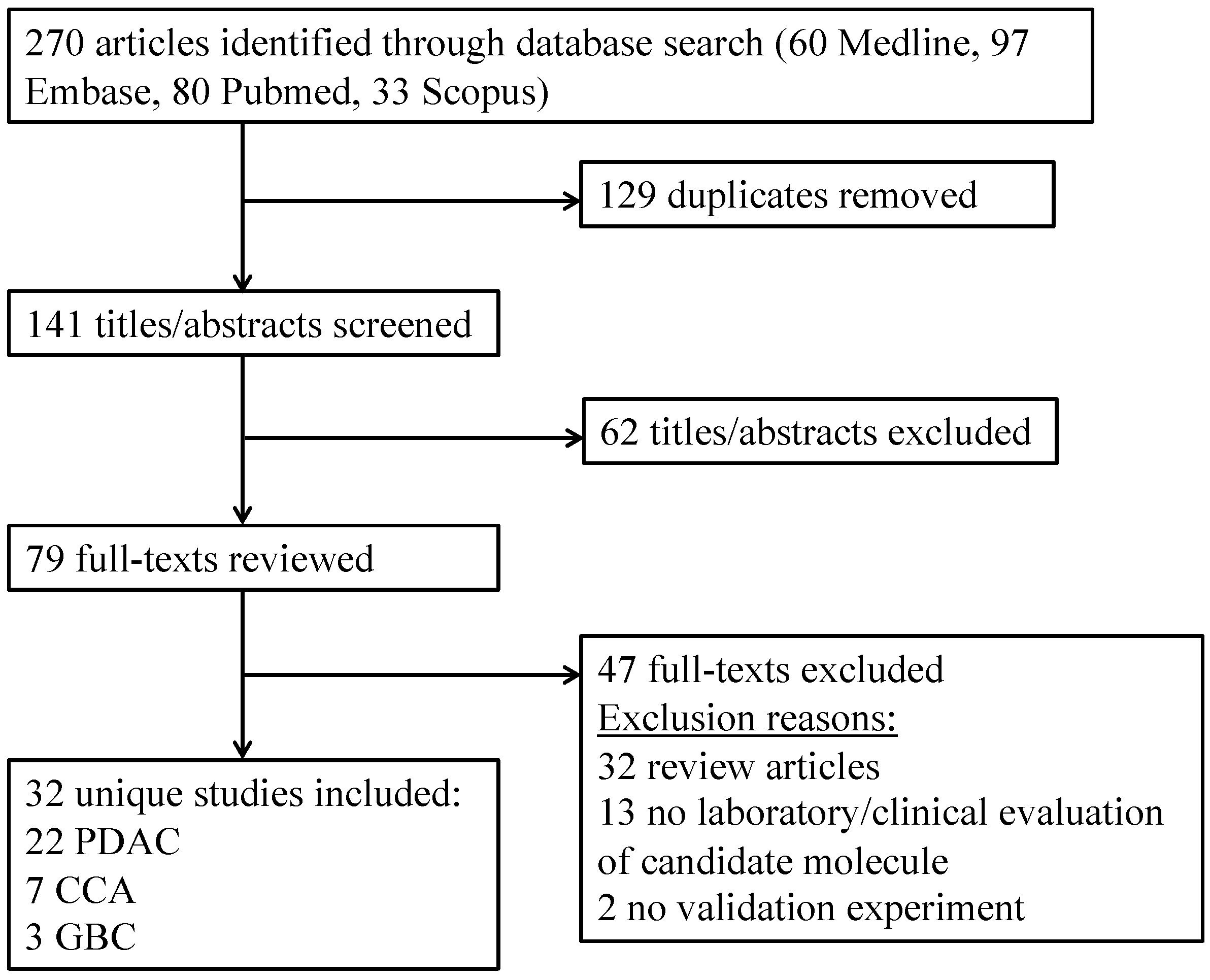
2. Results
2.1. Studies Included
2.2. Circular RNA Expression
2.3. In Vitro and In Vivo Characteristics
2.4. Clinical Disease Characteristics
2.5. Results for Biliary Tract Malignancies
3. Discussion
3.1. Biological Role of circRNA in PDAC
3.1.1. Increasing Cell Proliferation
3.1.2. Enhancing Tumour Invasion and Metastasis
3.1.3. Promoting Angiogenesis and Lymphangiogenesis
3.1.4. Attenuating the Immune Response
3.1.5. Epithelial to Mesenchymal Transition and Cancer Stem Cells
3.1.6. Chemotherapy Resistance
3.2. Clinical Utilisations of circRNAs
3.2.1. Diagnostic Biomarkers
3.2.2. Prognostic and Predictive Biomarkers
3.2.3. CircRNAs as Therapeutic Targets
3.3. Future Research
4. Systematic Review Methodology
4.1. Search
4.2. Eligibility Criteria
4.3. Data Collection
4.4. Results Reporting
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UK CR. Pancreatic and Biliary Tract Cancer Statistics. 2020. Available online: https://www.cancerresearchuk.org/ (accessed on 10 October 2020).
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Coupland, V.H.; Kocher, H.M.; Berry, D.P.; Allum, W.; Linklater, K.M.; Konfortion, J.; Moller, H.; Davies, E.A. Incidence and survival for hepatic, pancreatic and biliary cancers in England between 1998 and 2007. Cancer Epidemiol. 2012, 36, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci. Rep. 2015, 5, 12453. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Circular RNA (circRNA) in Alzheimer’s disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.C.S.; Elhassan, S.A.M.; Lim, G.P.L.; Kok, W.H.; Tan, S.W.; Leong, E.N.; Tan, S.H.; Chan, E.W.L.; Bhattamisra, S.K.; Rajendran, R.; et al. The roles of circular RNAs in human development and diseases. Biomed. Pharmacother. 2019, 111, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Hann, S.S. Biological roles and mechanisms of circular RNA in human cancers. Oncol. Targets Ther. 2020, 13, 2067–2092. [Google Scholar] [CrossRef] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome. Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V.; The, C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [Green Version]
- Kos, A.; Dijkema, R.; Arnberg, A.C.; van der Meide, P.H.; Schellekens, H. The hepatitis delta (delta) virus possesses a circular RNA. Nature 1986, 323, 558–560. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Memczak, S.; Papavasileiou, P.; Peters, O.; Rajewsky, N. Identification and characterization of circular RNAs as a new class of putative biomarkers in human blood. PLoS ONE 2015, 10, e0141214. [Google Scholar] [CrossRef] [PubMed]
- Nigro, J.M.; Cho, K.R.; Fearon, E.R.; Kern, S.E.; Ruppert, J.M.; Oliner, J.D.; Kinzler, K.W.; Vogelstein, B. Scrambled exons. Cell 1991, 64, 607–613. [Google Scholar] [CrossRef]
- Tufekci, K.U.; Oner, M.G.; Meuwissen, R.L.J.; Genc, S. The role of microRNAs in human diseases. Methods Mol. Biol. 2014, 1107, 33–50. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-L.; Yang, L. Regulation of circRNA biogenesis. RNA Biol. 2015, 12, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Cortés-López, M.; Miura, P. Emerging functions of circular RNAs. Yale J. Biol. Med. 2016, 89, 527–537. [Google Scholar]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Enuka, Y.; Lauriola, M.; Feldman, M.E.; Sas-Chen, A.; Ulitsky, I.; Yarden, Y. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res. 2016, 44, 1370–1383. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sharp, P.A. Molecular biology. A circuitous route to noncoding RNA. Science 2013, 340, 440–441. [Google Scholar] [CrossRef] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hetuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1993, 7, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Kai, D.; Yannian, L.; Yitian, C.; Dinghao, G.; Xin, Z.; Wu, J. Circular RNA HIPK3 promotes gallbladder cancer cell growth by sponging microRNA-124. Biochem. Biophys. Res. Commun. 2018, 503, 863–869. [Google Scholar] [CrossRef]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.-H.; Bindereif, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Haddad, G.; Lorenzen, J.M. Biogenesis and function of circular RNAs in health and in disease. Front. Pharmacol. 2019, 10, 428. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Bentley, D.L. Coupling mRNA processing with transcription in time and space. Nat. Rev. Genet. 2014, 15, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-O.; Wang, H.-B.; Zhang, Y.; Lu, X.; Chen, L.-L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; He, M.; Huang, S.; Lin, R.; Zhan, M.; Yang, D.; Shen, H.; Xu, S.; Cheng, W.; Yu, J.; et al. Circular RNA circERBB2 promotes gallbladder cancer progression by regulating PA2G4-dependent rDNA transcription. Mol. Cancer 2019, 18, 166. [Google Scholar] [CrossRef] [Green Version]
- Aktas, T.; Avsar, I.I.; Maticzka, D.; Bhardwaj, V.; Pessoa, R.C.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, S.P.; Wang, P.L.; Salzman, J. Circular RNA biogenesis can proceed through an exon-containing lariat precursor. Elife 2015, 4, e07540. [Google Scholar] [CrossRef]
- Hansen, T.B.; Wiklund, E.D.; Bramsen, J.B.; Villadsen, S.B.; Statham, A.L.; Clark, S.J.; Kjems, J. miRNA-dependent gene silencing involving Ago2-mediated cleavage of a circular antisense RNA. EMBO J. 2011, 30, 4414–4422. [Google Scholar] [CrossRef] [Green Version]
- Park, O.H.; Ha, H.; Lee, Y.; Boo, S.H.; Kwon, D.H.; Song, H.K.; Kim, Y.K. Endoribonucleolytic cleavage of m(6)A-containing RNAs by RNase P/MRP complex. Mol. Cell 2019, 74, 494–507. [Google Scholar] [CrossRef]
- Fischer, J.W.; Busa, V.F.; Shao, Y.; Leung, A.K.L. Structure-mediated RNA decay by UPF1 and G3BP1. Mol. Cell 2020, 78, 70–84. [Google Scholar] [CrossRef]
- Liu, C.-X.; Li, X.; Nan, F.; Jiang, S.; Gao, X.; Guo, S.-K.; Xue, W.; Cui, Y.; Dong, K.; Ding, H.; et al. Structure and degradation of circular RNAs regulate PKR activation in innate immunity. Cell 2019, 177, 865–880. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, X.; Peng, Y. Structure-mediated degradation of CircRNAs. Trends Cell Biol. 2020, 30, 501–503. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Qu, S.; Hao, X.; Song, W.; Niu, K.; Yang, X.; Zhang, X.; Shang, R.; Wang, Q.; Li, H.; Liu, Z. Circular RNA circRHOT1 is upregulated and promotes cell proliferation and invasion in pancreatic cancer. Epigenomics 2019, 11, 53–63. [Google Scholar] [CrossRef]
- Li, Z.; Yanfang, W.; Li, J.; Jiang, P.; Peng, T.; Chen, K.; Zhao, X.; Zhang, Y.; Zhen, P.; Zhu, J.; et al. Tumor-released exosomal circular RNA PDE8A promotes invasive growth via the miR-338/MACC1/MET pathway in pancreatic cancer. Cancer Lett. 2018, 432, 237–250. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Zhen, T.; Dong, Y.; Pei, X.; Shi, H. Hsa_circ_001653 Implicates in the development of pancreatic ductal adenocarcinoma by regulating microRNA-377-Mediated HOXC6 Axis. Mol Ther. Nucleic Acids 2020. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef] [PubMed]
- Denzler, R.; Agarwal, V.; Stefano, J.; Bartel, D.P.; Stoffel, M. Assessing the ceRNA hypothesis with quantitative measurements of miRNA and target abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, eaam8526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, Y.; Cai, Q.; Ma, M.; Jin, L.Y.; Weng, M.; Zhou, D.; Tang, Z.; Wang, J.D.; Quan, Z. Circular RNA FOXP1 promotes tumor progression and Warburg effect in gallbladder cancer by regulating PKLR expression. Mol. Cancer 2019, 18, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, L.; Rong, W.; Bai, L.; Cui, H.; Zhang, S.; Li, Y.; Chen, D.; Meng, X. Upregulated circular RNA circ_0007534 indicates an unfavorable prognosis in pancreatic ductal adenocarcinoma and regulates cell proliferation, apoptosis, and invasion by sponging miR-625 and miR-892b. J. Cell Biochem. 2019, 120, 3780–3789. [Google Scholar] [CrossRef]
- Abdelmohsen, K.; Panda, A.C.; Munk, R.; Grammatikakis, I.; Dudekula, D.B.; De, S.; Kim, J.; Noh, J.H.; Kim, K.M.; Martindale, J.L.; et al. Identification of HuR target circular RNAs uncovers suppression of PABPN1 translation by CircPABPN1. RNA Biol. 2017, 14, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, X.; Jia, Y.; Zhang, Y.; Shi, L.; Li, Q.; Zang, A.; Wang, H. Circular RNA: Biogenesis, degradation, functions and potential roles in mediating resistance to anticarcinogens. Epigenomics 2020, 12, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.-O.; Chen, T.; Xiang, J.-F.; Yin, Q.-F.; Xing, Y.-H.; Zhu, S.; Yang, L.; Chen, L.-L. Circular intronic long noncoding RNAs. Mol. Cell. 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Schneider, T.; Bindereif, A. Circular RNAs: Coding or noncoding? Cell Res. 2017, 27, 724–725. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Sarnow, P. Initiation of protein synthesis by the eukaryotic translational apparatus on circular RNAs. Science 1995, 268, 415–417. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol. Cell 2017, 66, 22–37. [Google Scholar] [CrossRef] [Green Version]
- Dong, R.; Zhang, X.-O.; Zhang, Y.; Ma, X.-K.; Chen, L.-L.; Yang, L. CircRNA-derived pseudogenes. Cell Res. 2016, 26, 747–750. [Google Scholar] [CrossRef]
- Feng, Z.; Meng, S.; Zhou, H.; Xu, Z.; Tang, Y.; Li, P.; Liu, C.; Huang, Y.; Wu, M. Functions and potential applications of circular RNAs in cancer stem cells. Front. Oncol. 2019, 9, 500. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Hao, X.; Wang, H.; Liu, Z.; He, Y.; Pu, M.; Zhang, H.; Yu, H.; Duan, J.; Qu, S. Circular RNA Expression profile of pancreatic ductal adenocarcinoma revealed by microarray. Cell Physiol. Biochem. 2016, 40, 1334–1344. [Google Scholar] [CrossRef]
- Guo, S.; Xu, X.; Ouyang, Y.; Wang, Y.; Yang, J.; Yin, L.; Ge, J.; Wang, H. Microarray expression profile analysis of circular RNAs in pancreatic cancer. Mol. Med. Rep. 2018, 17, 7661–7671. [Google Scholar] [CrossRef]
- Seimiya, T.; Otsuka, M.; Iwata, T.; Tanaka, E.; Sekiba, K.; Shibata, C.; Moriyama, M.; Nakagawa, R.; Maruyama, R.; Koike, K. Aberrant expression of a novel circular RNA in pancreatic cancer. J. Hum. Genet. 2020. [Google Scholar] [CrossRef]
- Szabo, L.; Salzman, J. Detecting circular RNAs: Bioinformatic and experimental challenges. Nat. Rev. Genet. 2016, 17, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Barrett, S.P.; Salzman, J. Circular RNAs: Analysis, expression and potential functions. Development 2016, 143, 1838–1847. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ji, P.; Chen, S.; Hou, L.; Zhao, F. Reconstruction of full-length circular RNAs enables isoform-level quantification. Genome Med. 2019, 11, 2. [Google Scholar] [CrossRef]
- Zhang, F.; Zhao, X.; Dong, H.; Xu, J. circRNA expression analysis in lung adenocarcinoma: Comparison of paired fresh frozen and formalin-fixed paraffin-embedded specimens. Biochem. Biophys. Res. Commun. 2018, 500, 738–743. [Google Scholar] [CrossRef]
- Thangaraj, M.P.; Furber, K.L.; Gan, J.K.; Ji, S.; Sobchishin, L.; Doucette, J.R.; Nazarali, A.J. RNA-binding protein quaking stabilizes Sirt2 mRNA during oligodendroglial differentiation. J. Biol. Chem. 2017, 292, 5166–5182. [Google Scholar] [CrossRef] [Green Version]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for sequence-based miRNA target prediction: What to choose? Int. J. Mol. Sci. 2016, 17, 1987. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Ni, J.; Lu, S.; Jiang, S.; You, L.; Liu, H.; Shou, J.; Zhai, C.; Zhang, W.; Shao, S.; et al. CircUBAP2-mediated competing endogenous RNA network modulates tumorigenesis in pancreatic adenocarcinoma. Aging 2019, 11, 8484–8501. [Google Scholar] [CrossRef]
- Zhou, J.-Z.; Hu, M.-R.; Diao, H.-L.; Wang, Q.-W.; Huang, Q.; Ge, B.-J. Comprehensive analysis of differentially expressed circRNAs revealed a ceRNA network in pancreatic ductaladenocarcinoma. Arch. Med. Sci. 2019, 15, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.-M.; Jiang, M.-T.; Lin, P.; Yang, H.; Dang, Y.-W.; Yu, Q.; Liao, D.-Y.; Luo, D.-Z.; Chen, G. Potential ceRNA networks involved in autophagy suppression of pancreatic cancer caused by chloroquine diphosphate: A study based on differentiallyexpressed circRNAs, lncRNAs, miRNAs and mRNAs. Int. J. Oncol. 2019, 54, 600–626. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chen, G.; Shi, Y.; Zhang, Y.; Sun, J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. Oncol. Targets Ther. 2017, 10, 5783–5794. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Liu, D.-Y.; Guo, J.-T.; Ge, N.; Zhu, P.; Liu, X.; Wang, S.; Wang, G.-X.; Sun, S.-Y. Circular RNA circ-LDLRAD3 as a biomarker in diagnosis of pancreatic cancer. World J. Gastroenterol. 2017, 23, 8345–8354. [Google Scholar] [CrossRef]
- An, Y.; Cai, H.; Zhang, Y.; Liu, S.; Duan, Y.; Sun, D.; Chen, X.; He, X. circZMYM2 Competed endogenously with miR-335-5p to regulate JMJD2C in pancreatic cancer. Cell Physiol. Biochem. 2018, 51, 2224–2236. [Google Scholar] [CrossRef]
- Zhu, P.; Ge, N.; Liu, D.; Yang, F.; Zhang, K.; Guo, J.; Liu, X.; Wang, S.; Wang, G.; Sun, S. Preliminary investigation of the function of hsa_circ_0006215 in pancreatic cancer. Oncol. Lett. 2018, 16, 603–611. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Jiang, P.; Peng, M.; Zhang, X.; Chen, K.; Liu, H.; Bi, H.; Liu, X.; Li, X. Circular RNA IARS (circ-IARS) secreted by pancreatic cancer cells and located within exosomes regulates endothelial monolayer permeability to promote tumor metastasis. J. Exp. Clin. Cancer Res. 2018, 37, 177. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wang, T.; Yan, L.; Qu, L. A novel prognostic biomarker for pancreatic ductal adenocarcinoma: Hsa_circ_0001649. Gene 2018, 675, 88–93. [Google Scholar] [CrossRef]
- Xu, Y.; Yao, Y.; Gao, P.; Cui, Y. Upregulated circular RNA circ_0030235 predicts unfavorable prognosis in pancreatic ductal adenocarcinoma and facilitates cell progression by sponging miR-1253 and miR-1294. Biochem. Biophys. Res. Commun. 2019, 509, 138–142. [Google Scholar] [CrossRef]
- Liu, L.; Liu, F.-B.; Huang, M.; Xie, K.; Xie, Q.-S.; Liu, C.-H.; Shen, M.-J.; Huang, Q. Circular RNA ciRS-7 promotes the proliferation and metastasis of pancreatic cancer by regulating miR-7-mediated EGFR/STAT3 signaling pathway. Hepatobiliary Pancreat. Dis. Int. 2019. [Google Scholar] [CrossRef]
- Yang, J.; Cong, X.; Ren, M.; Sun, H.; Liu, T.; Chen, G.; Wang, Q.; Li, Z.; Yu, S.; Yang, Q. Circular RNA hsa_circRNA_0007334 is predicted to promote MMP7 and COL1A1 expression by functioning as a miRNA sponge in pancreatic ductal adenocarcinoma. J. Oncol. 2019, 2019, 7630894. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhang, C.; Chen, Y.; Gao, S. Downregulation of circular RNA circ-LDLRAD3 suppresses pancreatic cancer progression through miR-137-3p/PTN axis. Life Sci. 2019, 116871. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Z.; Zhang, M.; Wang, B.; Ye, J.; Zhang, Y.; Tang, D.; Ma, D.; Jin, W.; Li, X.; et al. Circ-ASH2L promotes tumor progression by sponging miR-34a to regulate Notch1 in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 466. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Ye, H.; Wang, W.; Sun, M.; Zhang, J.; Zhao, Z.; Jiang, G. Circular RNA ADAM9 facilitates the malignant behaviours of pancreatic cancer by sponging miR-217 and upregulating PRSS3 expression. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3920–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xia, L.; Dong, L.; Wang, J.; Xiao, Q.; Yu, X.; Zhu, H. CircHIPK3 promotes Gemcitabine (GEM) resistance in pancreatic cancer cells by sponging miR-330-5p and targets RASSF1. Cancer Manag. Res. 2020, 12, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.H.; Lou, U.K.; Li, Y.; Chan, S.L.; Tong, J.H.M.; To, K.-F.; Chen, Y. CircFOXK2 promotes growth and metastasis of pancreatic ductal adenocarcinoma by complexing with RNA binding proteins and sponging MiR-942. Cancer Res. 2020, 80, 2138–2149. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhou, Q.; Su, D.; Luo, Y.; Fu, Z.; Huang, L.; Li, Z.; Jiang, D.; Kong, Y.; Li, Z.; et al. Circular RNA circBFAR promotes the progression of pancreatic ductal adenocarcinoma via the miR-34b-5p/MET/Akt axis. Mol. Cancer 2020, 19. [Google Scholar] [CrossRef]
- Kong, Y.; Li, Y.; Luo, Y.; Zhu, J.; Zheng, H.; Gao, B.; Guo, X.; Li, Z.; Chen, R.; Chen, C. circNFIB1 inhibits lymphangiogenesis and lymphatic metastasis via the miR-486-5p/PIK3R1/VEGF-C axis in pancreatic cancer. Mol. Cancer 2020, 19. [Google Scholar] [CrossRef]
- Guo, W.; Zhao, L.; Wei, G.; Liu, P.; Zhang, Y.; Fu, L. Blocking circ_0013912 suppressed cell growth, migration and invasion of pancreatic ductal adenocarcinoma cells in vitro and in vivo partially through sponging miR-7-5p. Cancer Manag. Res. 2020, 12, 7291–7303. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, P.; Zhuang, Y.; Du, L. Hsa_circRNA_001587 upregulates the expression of SLC4A4 to inhibit migration and invasion and angiogenesis of pancreatic cancer cells via binding to microRNA. Gastrointest. Liver Physiol. 2020. Online ahead of print. [Google Scholar] [CrossRef]
- Jiang, X.-M.; Li, Z.-L.; Li, J.-L.; Xu, Y.; Leng, K.-M.; Cui, Y.-F.; Sun, D.-J. A novel prognostic biomarker for cholangiocarcinoma: CircRNA Cdr1as. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 365–371. [Google Scholar] [CrossRef]
- Xu, Y.; Yao, Y.; Liu, Y.; Wang, Z.; Hu, Z.; Su, Z.; Li, C.; Wang, H.; Jiang, X.; Kang, P.; et al. Elevation of circular RNA circ_0005230 facilitates cell growth and metastasis via sponging miR-1238 and miR-1299 in cholangiocarcinoma. Aging (Albany NY) 2019, 11, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yao, Y.; Zhong, X.; Leng, K.; Qin, W.; Qu, L.; Cui, Y.; Jiang, X. Downregulated circular RNA hsa_circ_0001649 regulates proliferation, migration and invasion in cholangiocarcinoma cells. Biochem. Biophys. Res. Commun. 2018, 496, 455–461. [Google Scholar] [CrossRef]
- Li, D.; Tang, Z.; Gao, Z.; Shen, P.; Liu, Z.; Dang, X. Circular RNA CDR1as exerts oncogenic properties partially through regulating miR-641 in cholangiocarcinoma. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef]
- Wang, S.; Hu, Y.; Lv, X.; Li, B.; Gu, D.; Li, Y.; Sun, Y.; Su, Y. Circ-0000284 arouses malignant phenotype of cholangiocarcinoma cells and regulates the biological functions of peripheral cells through cellular communication. Clin. Sci. 2019, 133, 1935–1953. [Google Scholar] [CrossRef]
- Xu, Y.; Leng, K.; Yao, Y.; Kang, P.; Liao, G.; Han, Y.; Shi, G.; Ji, D.; Huang, P.; Zheng, W.; et al. A novel circular RNA, circ-CCAC1, contributes to CCA progression, induces angiogenesis, and disrupts vascular endothelial barriers. Hepatology 2020. [Google Scholar] [CrossRef]
- Li, X.; He, M.; Guo, J.; Cao, T. Upregulation of circular RNA circ-ERBB2 predicts unfavorable prognosis and facilitates the progression of gastric cancer via miR-503/CACUL1 and miR-637/MMP-19 signaling. Biochem. Biophys. Res. Commun. 2019, 511, 926–930. [Google Scholar] [CrossRef]
- Lu, Q.; Fang, T. Circular RNA SMARCA5 correlates with favorable clinical tumor features and prognosis, and increases chemotherapy sensitivity in intrahepatic cholangiocarcinoma. J. Clin. Lab. Anal. 2019, 34, e23138. [Google Scholar] [CrossRef]
- Junttila, M.R.; De Sauvage, F.J. Influence of tumour micro-environment heterogeneity on therapeutic response. Nat. Cell Biol. 2013, 501, 346–354. [Google Scholar] [CrossRef]
- Ma, Z.; Shuai, Y.; Gao, X.; Wen, X.; Ji, J. Circular RNAs in the tumour microenvironment. Mol. Cancer 2020, 19, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Zheng, S.; Deng, X.; Yang, A.; Xie, X.; Tang, H. The role of circular RNA CDR1as/ciRS-7 in regulating tumor microenvironment: A pan-cancer analysis. Biomolecules 2019, 9, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Cao, F.; Ren, G.; Gao, D.; Bhakta, V.; Zhang, Y.; Cao, H.; Dong, Z.; Zang, W.; Zhang, S.; et al. PRSS3 promotes tumour growth and metastasis of human pancreatic cancer. Gut 2010, 59, 1535–1544. [Google Scholar] [CrossRef]
- Wang, G.; Kang, M.-X.; Lu, W.-J.; Chen, Y.; Zhang, B.; Wu, Y.-L. MACC1: A potential molecule associated with pancreatic cancer metastasis and chemoresistance. Oncol. Lett. 2012, 4, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, T.M.; Gleisner, A.L.; Cameron, J.L.; Winter, J.M.; Assumpcao, L.; Lillemoe, K.D.; Wolfgang, C.; Hruban, R.H.; Schulick, R.D.; Yeo, C.J.; et al. Prognostic relevance of lymph node ratio following pancreaticoduodenectomy for pancreatic cancer. Surgery 2007, 141, 610–618. [Google Scholar] [CrossRef]
- Ou, Z.-L.; Luo, Z.; Wei, W.; Liang, S.; Gao, T.-L.; Lu, Y.-B. Hypoxia-induced shedding of MICA and HIF1A-mediated immune escape of pancreatic cancer cells from NK cells: Role of circ_0000977/miR-153 axis. RNA Biol. 2019, 16, 1592–1603. [Google Scholar] [CrossRef]
- Wang, S.; Huang, S.; Sun, Y.L. Epithelial-mesenchymal transition in pancreatic cancer: A review. BioMed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lagunas-Rangel, F.A. Circular RNAs and their participation in stemness of cancer. Med. Oncol. 2020, 37, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, A.; Barilani, M.; Rossi, R.L.; Jalal, M.M.K.; Rusconi, F.; Buono, G.; Ragni, E.; Cantarella, G.; Simpson, H.A.R.W.; Péault, B.; et al. FOXP1 circular RNA sustains mesenchymal stem cell identity via microRNA inhibition. Nucleic Acids Res. 2019, 47, 5325–5340. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Wang, D.; Wei, A.; Ke, N.; Wang, Y.; Tang, J.; He, S.; Hu, W.; Liu, X. MicroRNA-410-3p attenuates gemcitabine resistance in pancreatic ductal adenocarcinoma by inhibiting HMGB1-mediated autophagy. Oncotarget 2017, 8, 107500–107512. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Yu, Y.; Ding, F. Microarray analysis of circular RNA expression profiles associated with gemcitabine resistance in pancreatic cancer cells. Oncol. Rep. 2018, 40, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Shao, F.; Huang, M.; Meng, F.; Huang, Q. Circular RNA Signature Predicts Gemcitabine Resistance of Pancreatic Ductal Adenocarcinoma. Front. Pharmacol. 2018, 9, 584. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhang, L.; Han, G.; Zuo, X.; Zhang, Y.; Zhu, Q.; Wu, J.; Wang, X. A meta-analysis of the diagnostic accuracy of circular RNAs in digestive system malignancy. Cell. Physiol. Biochem. 2018, 45, 962–972. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, H.; Lv, X.; Yang, Z.; Gao, M.; Bi, Y.; Zhang, Z.; Wang, S.; Cui, Z.; Zhou, B.; et al. Diagnostic performance of circular RNAs in human cancers: A systematic review and meta-analysis. Mol. Genet. Genom. Med. 2019, 7, e00749. [Google Scholar] [CrossRef]
- Bahn, J.H.; Zhang, Q.; Li, F.; Chan, T.-M.; Lin, X.; Kim, Y.; Wong, D.T.; Xiao, X. The landscape of MicroRNA, Piwi-Interacting RNA, and circular RNA in human saliva. Clin. Chem. 2015, 61, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Tuck, M.K.; Chan, D.W.; Chia, D.; Godwin, A.K.; Grizzle, W.E.; Krueger, K.E.; Rom, W.; Sanda, M.; Sorbara, L.; Stass, S.; et al. Standard operating procedures for serum and plasma collection: Early detection research network consensus statement standard operating procedure integration working group. J. Proteome Res. 2009, 8, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Lei, K.; Huang, F.; Jiang, Z.; Zhou, X. Exo-circRNAs: A new paradigm for anticancer therapy. Mol. Cancer 2019, 18, 56. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Tvall, J.O.L.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Seimiya, T.; Otsuka, M.; Liu, H.-Y.; Suzuki, T.; Sekiba, K.; Yamagami, M.; Tanaka, E.; Ishibashi, R.; Koike, K. Circular RNA and exosomes in pancreatic cancer progression. Transl. Cancer Res. 2018, 7, S745–S747. [Google Scholar] [CrossRef]
- Shengli, L. RNA-seq Reveals Abundant circRNA, lncRNA and mRNA in Blood Exosomes of Patients with Pancreatic Carcinoma. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE100232 (accessed on 10 October 2020).
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. exoRBase: A database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2017, 46, D106–D112. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Geng, S.; Yuan, H.; Li, Y.; Zhang, S.; Pu, L.; Ge, J.; Niu, X.; Li, Y.; Jiang, H. Circular RNA expression profiles in extracellular vesicles from the plasma of patients with pancreatic ductal adenocarcinoma. FEBS Open Bio 2019, 9, 2052–2062. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Sole, C.; Arnaiz, E.; Manterola, L.; Otaegui, D.; Lawrie, C.H. The circulating transcriptome as a source of cancer liquid biopsy biomarkers. Semin. Cancer Biol. 2019, 58, 100–108. [Google Scholar] [CrossRef]
- Reddi, K.K.; Holland, J.F. Elevated serum ribonuclease in patients with pancreatic cancer. Proc. Natl. Acad. Sci. USA 1976, 73, 2308–2310. [Google Scholar] [CrossRef] [Green Version]
- A Burris, H.; Moore, M.J.; Andersen, J.; Green, M.R.; Rothenberg, M.L.; Modiano, M.R.; Cripps, M.C.; Portenoy, R.K.; Storniolo, A.M.; Tarassoff, P.; et al. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: A randomized trial. J. Clin. Oncol. 1997, 15, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Von Hoff, D.D.; Ervin, T.; Arena, F.P.; Chiorean, E.G.; Infante, J.; Moore, M.; Seay, T.; Tjulandin, S.A.; Ma, W.W.; Saleh, M.N.; et al. Increased survival in pancreatic cancer with nab-paclitaxel plus gemcitabine. N. Engl. J. Med. 2013, 369, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Gzil, A.; Zarębska, I.; Bursiewicz, W.; Antosik, P.; Grzanka, D.; Szylberg, Ł. Markers of pancreatic cancer stem cells and their clinical and therapeutic implications. Mol. Biol. Rep. 2019, 46, 6629–6645. [Google Scholar] [CrossRef] [Green Version]
- Lyu, D.; Huang, S. The emerging role and clinical implication of human exonic circular RNA. RNA Biol. 2016, 14, 1000–1006. [Google Scholar] [CrossRef]
- Notta, F.; Chan-Seng-Yue, M.; Lemire, M.; Ilinca, L.; Wilson, G.W.; Connor, A.A.; Denroche, R.E.; Liang, S.-B.; Brown, A.M.K.; Kim, J.C.; et al. A renewed model of pancreatic cancer evolution based on genomic rearrangement patterns. Nat. Cell Biol. 2016, 538, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.-J.; Ma, Y.-S.; Jiang, X.-Q.; Wu, T.-M.; Wu, Z.-J.; Li, Z.-Z.; Wang, J.-H.; Gao, Q.-X.; Yi, B.; Shi, Y.; et al. Whole-transcriptome sequencing identifies key differentially expressed mRNAs, miRNAs, lncRNAs, and circRNAs associated with CHOL. Mol. Ther. Nucleic Acids 2020, 21, 592–603. [Google Scholar] [CrossRef]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [Green Version]
- Dong, R.; Ma, X.-K.; Li, G.-W.; Yang, L. CIRCpedia v2: An updated database for comprehensive circular RNA annotation and expression comparison. Genom. Proteom. Bioinform. 2018, 16, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Ji, P.; Zhao, F. CircAtlas: An integrated resource of one million highly accurate circular RNAs from 1070 vertebrate transcriptomes. Genome Biol. 2020, 21, 1–14. [Google Scholar] [CrossRef]
- Jacobson, R.; Munding, E.; Hayden, D.M.; Levy, M.A.; Kuzel, T.M.; Pappas, S.G.; Masood, A. Levy evolving clinical utility of liquid biopsy in gastrointestinal cancers. Cancers 2019, 11, 1164. [Google Scholar] [CrossRef] [Green Version]
- Kaczor-Urbanowicz, K.E.; Cheng, J.; King, J.C.; Sedarat, A.; Pandol, S.J.; Farrell, J.J.; Wong, D.T.; Kim, Y. Reviews on current liquid biopsy for detection and management of pancreatic cancers. Pancreas 2020, 49, 1141–1152. [Google Scholar] [CrossRef]
- Luchini, C.; Veronese, N.; Nottegar, A.; Cappelletti, V.; Daidone, M.G.; Smith, L.; Parris, C.; Brosens, L.A.A.; Caruso, M.G.; Cheng, L.; et al. Liquid biopsy as surrogate for tissue for molecular profiling in pancreatic cancer: A meta-analysis towards precision medicine. Cancers 2019, 11, 1152. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Author | Year | Cancer Type | circRNA | Expression in PDAC Samples | Association with Features of Malignancy Demonstrated |
|---|---|---|---|---|---|
| Chen G. at al. [78] * | 2017 | PDAC | hsa_circ_100782 | Upregulated | In vitro and in vivo |
| Yang F. et al. [79] | 2017 | PDAC | hsa_circ_0006988 | Upregulated | Clinical data |
| An Y. et. al. [80] | 2018 | PDAC | hsa_circ_0099999 (circZMYM2) | Upregulated | In vitro and in vivo |
| Zhu P. et al. [81] | 2018 | PDAC | hsa_circ_0006215 | Upregulated | In vitro |
| Li J. et al. [82] | 2018 | PDAC | circ-IARS | Upregulated | In vitro, in vivo and clinical data |
| Li Z. et al. [47] | 2018 | PDAC | circ-PDE8A | Upregulated | In vitro, in vivo and clinical data |
| Jiang Y. et al. [83] | 2018 | PDAC | hsa_circ_0001649 | Downregulated | In vitro and clinical data |
| Qu S. et al. [46] | 2019 | PDAC | hsa_circ_0005397 (circ-RHOT1) | Upregulated | In vitro |
| Xu Y. et al. [84] | 2019 | PDAC | hsa_circ_0030235 | Upregulated | In vitro and clinical data |
| Hao L. et al. [54] | 2019 | PDAC | hsa_circ_0007534 | Upregulated | In vitro, in vivo and clinical data |
| Liu L. et al. [85] | 2019 | PDAC | ciRS-7 (Cdr1as) | Upregulated | In vitro and clinical data |
| Yang J. et al. [86] | 2019 | PDAC | hsa_circ_0007334 | Upregulated | In vitro |
| Yao J. et al. [87] | 2019 | PDAC | circLDLRAD3 | Upregulated | In vitro and in vivo |
| Chen Y. et al. [88] | 2019 | PDAC | circASH2L | Upregulated | In vitro, in vivo and clinical data |
| Xing C. et al. [89] | 2019 | PDAC | circADAM9 | Upregulated | In vitro, in vivo and clinical data |
| Zhang X et al. [48] | 2020 | PDAC | hsa_circ_001653 | Upregulated | In vitro, in vivo and clinical data |
| Liu Y. et al. [90] | 2020 | PDAC | circHIPK3 | Upregulated | In vitro and in vivo |
| Wong C. et al. [91] | 2020 | PDAC | circFOXK2 | Upregulated | In vitro and in vivo |
| Guo X. et al. [92] | 2020 | PDAC | hsa_circ_0009065 (circBFAR) | Upregulated | In vitro and in vivo |
| Kong Y. et al. [93] | 2020 | PDAC | hsa_circ_0086375 (circNFIB1) | Downregulated | In vitro, in vivo and clinical data |
| Guo W. et al. [94] | 2020 | PDAC | hsa_circ_0013912 | Upregulated | In vitro, in vivo and clinical data |
| Zhang X. et al. [95] | 2020 | PDAC | hsa_circ_001587 | Downregulated | In vitro, in vivo and clinical data |
| circRNA | Expression in PDAC Tissue | Expression in Cell Lines | miRNA | Implicated Molecules and Pathways |
|---|---|---|---|---|
| hsa_circ_100782 | − | Upregulated | miR-124 | IL6/STAT3 |
| hsa_circ_0099999 (circZMYM2) | Upregulated | − | miR-335-5p | JMJD2C |
| hsa_circ_0006215 | Upregulated | − | miR-378a-3p | SERPINA4 |
| circ-IARS | Upregulated | Upregulated (and exosomes) | miR-122 | ZO-1, RhoA, F-actin |
| circ-PDE8A | Upregulated | Upregulated | miR-338 | MACC1/MET |
| hsa_circ_0005397 (circ-RHOT1) | Upregulated | Upregulated | miR-26b; miR-125a, miR-330; miR-382 | − |
| hsa_circ_0030235 | Upregulated | Upregulated | miR-1253; miR-1294 | − |
| hsa_circ_0007534 | Upregulated | Upregulated | miR-625; miR-892b | − |
| ciRS-7 (Cdr1as) | Upregulated | − | miR-7 | EGF/STAT3 |
| hsa_circ_0007334 | Upregulated | − | miR-144-3p; miR-577 | MMP7 |
| circ-LDLRAD3 | Upregulated | Upregulated | miR-137-3p | Pleiotrophin |
| circASH2L | Upregulated | Upregulated | miR-34a | Notch 1 |
| circADAM9 | Upregulated | Upregulated | miR-217 | PRSS3 |
| hsa_circ_001653 | Upregulated | Upregulated | miR-377 | HOXC6 |
| circHIPK3 | Upregulated | Upregulated | miR-330-5p | RASSF1 |
| circFOXK2 | Upregulated | Upregulated | miR-942 | YBX1 and hnRNPK; NUF2 and PDXK |
| hsa_circ_0009065 (circBFAR) | Upregulated | Upregulated | miR-34b-5p | MET |
| hsa_circ_0086375 (circNFIB1) | Downregulated | Downregulated | miR-486-5p | PIK3R1/VEGF-C |
| hsa_circ_0013912 | Upregulated | Upregulated | miR-7-5p | − |
| hsa_circ_001587 | Downregulated | Downregulated | miR-223 | SLC4A4 |
| circRNA | Study Type | Proliferation/Viability | Migration | Invasion | Apoptosis |
|---|---|---|---|---|---|
| hsa_circ_100782 | Silencing of upregulated circRNA | ↓ | − | − | − |
| hsa_circ_0099999 | ↓ | − | ↓ | ↑ | |
| hsa_circ_0006215 | ←→ | ↓ | − | − | |
| circ-IARS | − | ↓ | − | − | |
| hsa_circ_0005397 (circ-RHOT1) | ↓ | ↓ | ↓ | − | |
| hsa_circ_0030235 | ↓ | − | − | ↑ | |
| hsa_circ_0007534 | ↓ | ↓ | ↓ | ↑ | |
| ciRS-7 (Cdr1as) | ↓ | − | ↓ | − | |
| hsa_circ_0007334 | − | − | − | − | |
| circ-LDLRAD3 | ↓ | ↓ | ↓ | − | |
| hsa_circ_001653 | ↓ | − | ↓ | ↑ | |
| circHIPK3 | ↓ | ↓ | ↓ | ↑ | |
| circFOXK2 | ↓ | ↓ | ↓ | ↑ | |
| hsa_circ_0009065 (circBFAR) | ↓ | ↓ | ↓ | − | |
| hsa_circ_0013912 | ↓ | ↓ | ↓ | ↑ | |
| hsa_circ_0099999 | Overexpression of upregulated circRNA | ↑ | − | ↑ | ↓ |
| hsa_circ_0006215 | ↑ | ↑ | − | ↑ | |
| circ-IARS | − | ↑ | − | − | |
| circ-PDE8A | − | ↑ | ↑ | − | |
| hsa_circ_0030235 | ↑ | ↑ | − | ↓ | |
| hsa_circ_0007534 | ↑ | ↑ | ↑ | ↓ | |
| circASH2L | ↑ | ↑ | ↑ | − | |
| circADAM9 | ↑ | ↑ | ↑ | − | |
| hsa_circ_001653 | ↑ | − | ↑ | ↓ | |
| circFOXK2 | ↑ | ↑ | ↑ | − | |
| hsa_circ_0009065 (circBFAR) | ↑ | ↑ | ↑ | − | |
| hsa_circ_0086375 (circNFIB1) | Silencing of downregulated circRNA * | − | ↑ | − | − |
| hsa_circ_001587 | ↑ | ↑ | ↑ | − | |
| hsa_circ_0001649 | Overexpression of downregulated circRNA * | ↓ | − | − | ↑ |
| hsa_circ_0086375 (circNFIB1) | − | ↓ | − | − | |
| hsa_circ_001587 | ↓ | ↓ | ↓ | − |
| circRNA | Animal | Method | Findings |
|---|---|---|---|
| hsa_circ_100782 | Nude mice | circRNA knockdown | Decreased tumour size |
| hsa_circ_0099999 | Nude mice | circRNA knockdown | Decreased tumour size |
| circ-IARS | Nude mice | circRNA overexpression | Increased tumour size and metastatic disease |
| circ-PDE8A | Nude mice | circRNA overexpression | Increased peripheral blood exosomal GFP signals |
| hsa_circ_0007534 | Nude mice | circRNA knockdown | Decreased tumour size |
| circ-LDLRAD3 | Nude mice | circRNA knockdown | Decreased tumour size and weight |
| circASH2L | Nude mice | circRNA overexpression | Increased tumour size and metastatic disease |
| circADAM9 | Nude mice | circRNA knockdown | Decreased tumour size and weight |
| hsa_circ_001653 | Nude mice | circRNA knockdown | Decreased tumour size and weight |
| circHIPK3 | Nude mice | circRNA knockdown | Decreased tumour size and weight |
| circFOXK2 | Nude mice | circRNA knockdown | Decreased tumour size and metastasis |
| hsa_circ_0009065 (circBFAR) | Nude mice | circRNA overexpression | Increased tumour size |
| hsa_circ_0086375 (circNFIB1) | Nude mice | circRNA knockdown * | Increased lymph node metastasis |
| hsa_circ_0013912 | Nude mice | circRNA knockdown | Decreased tumour size and weight |
| hsa_circ_001587 | Nude mice | circRNA overexpression * | Decreased tumour size and weight |
| circRNA knockdown * | Increased tumour size and weight |
| circRNA | Direction of Dysregulation | Sample Assessed | Tumour Size | Duodenal Invasion | Neural Invasion | Lymphatic Spread | Vascular Spread | Metastatic Disease | Stage (TNM) | Differentiation Grade | Survival Time |
|---|---|---|---|---|---|---|---|---|---|---|---|
| hsa_circ_0006988 | Upregulated | Tissue | ←→ | − | − | ↑ | ↑ | ←→ | ←→ | − | − |
| hsa_circ_0006988 | Upregulated | Plasma | ←→ | − | − | ↑ | ↑ | ←→ | − | − | − |
| circ-IARS | Upregulated | Tissue | ←→ | ←→ | ↑ | ←→ | ↑ | ↑ | ↑ | − | ↓ |
| circ-PDE8A | Upregulated | Tissue | ←→ | ←→ | ←→ | ↑ | ←→ | ←→ | ↑ | ←→ | ↓ |
| circ-PDE8A | Upregulated | Plasma exosome | ←→ | ↑ | ←→ | ←→ | ↑ | ↑ | ↑ | ←→ | ↓ |
| hsa_circ_0001649 | Downregulated | Tissue | − | − | − | ←→ | − | − | ↑ | ↓ | ↓ |
| hsa_circ_0030235 | Upregulated | Tissue | − | − | − | ↑ | − | − | ↑ | ←→ | ↓ |
| hsa_circ_0007534 | Upregulated | Tissue | − | − | − | ↑ | − | − | ↑ | ←→ | − |
| ciRS-7 (Cdr1as) | Upregulated | Tissue | ←→ | − | − | ↑ | ↑ | − | − | − | − |
| circASH2L | Upregulated | Tissue | ←→ | ←→ | ←→ | ↑ | ←→ | ←→ | ↑ | ←→ | ↓ |
| circADAM9 | Upregulated | Tissue | − | − | − | ↑ | − | − | ↑ | − | ↓ |
| hsa_circ_001653 | Upregulated | Tissue | − | − | − | − | − | − | − | − | ↓ |
| hsa_circ_0009065 (circBFAR) | Upregulated | Tissue | ←→ | − | − | ←→ | − | − | ↑ | ←→ | ↓ |
| hsa_circ_0086375 (circNFIB1) | Downregulated | Tissue | ←→ | − | − | ↑ | − | − | ↑ | ←→ | ↓ |
| hsa_circ_0013912 | Upregulated | Tissue | ←→ | − | − | ↑ | − | − | ↑ | − | − |
| hsa_circ_0086375 (circNFIB1) | Downregulated | Tissue | − | − | − | ↑ | − | − | − | ↑ | ↓ |
| Author | Year | Cancer Type | circRNA | Expression in Tumour Tissue | Ass Association with Features of Malignancy Evaluated |
|---|---|---|---|---|---|
| Xu Y et al. [98] | 2018 | CCA | hsa_circ_0001649 | Down | In vitro, in vivo and clinical data |
| Jiang X et al. [96] | 2018 | CCA | ciRS-7 (Cdr1as) | Up | Clinical data |
| Kai D et al. [26] | 2018 | GBC | hsa_circ_0000284 (or circHIPK3) | Up | In vitro |
| Xu Y et al. [97] | 2019 | CCA | circ_0005230 | Up | In vitro, in vivo and clinical data |
| Wang et al. [53] | 2019 | GBC | hsa_circ_0008234 (or circFOXP1) | Up | In vitro, in vivo and clinical data |
| Lu Q and Fang T [103] | 2019 | CCA | circSMARCA5 | Down | In vitro and clinical data |
| Huang X et al. [33] | 2019 | GBC | circERBB2 | Up | In vitro, in vivo and clinical data |
| Wang S et al. [100] | 2019 | CCA | hsa_circ_0000284 | Up | In vitro and in vivo |
| Li D et al. [99] | 2020 | CCA | ciRS-7 (Cdr1as) | Up | In vitro and in vivo |
| Xu Y et al. [101] | 2020 | CCA | hsa_circ_102064 (or circ-CCAC1) | Up | In vitro, in vivo and clinical data |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limb, C.; Liu, D.S.K.; Veno, M.T.; Rees, E.; Krell, J.; Bagwan, I.N.; Giovannetti, E.; Pandha, H.; Strobel, O.; Rockall, T.A.; et al. The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers. Cancers 2020, 12, 3250. https://doi.org/10.3390/cancers12113250
Limb C, Liu DSK, Veno MT, Rees E, Krell J, Bagwan IN, Giovannetti E, Pandha H, Strobel O, Rockall TA, et al. The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers. Cancers. 2020; 12(11):3250. https://doi.org/10.3390/cancers12113250
Chicago/Turabian StyleLimb, Christopher, Daniel S. K. Liu, Morten T. Veno, Eleanor Rees, Jonathan Krell, Izhar N. Bagwan, Elisa Giovannetti, Hardev Pandha, Oliver Strobel, Timothy A. Rockall, and et al. 2020. "The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers" Cancers 12, no. 11: 3250. https://doi.org/10.3390/cancers12113250
APA StyleLimb, C., Liu, D. S. K., Veno, M. T., Rees, E., Krell, J., Bagwan, I. N., Giovannetti, E., Pandha, H., Strobel, O., Rockall, T. A., & Frampton, A. E. (2020). The Role of Circular RNAs in Pancreatic Ductal Adenocarcinoma and Biliary-Tract Cancers. Cancers, 12(11), 3250. https://doi.org/10.3390/cancers12113250