The Role of Steroid Hormone Receptors in Urothelial Tumorigenesis



Abstract
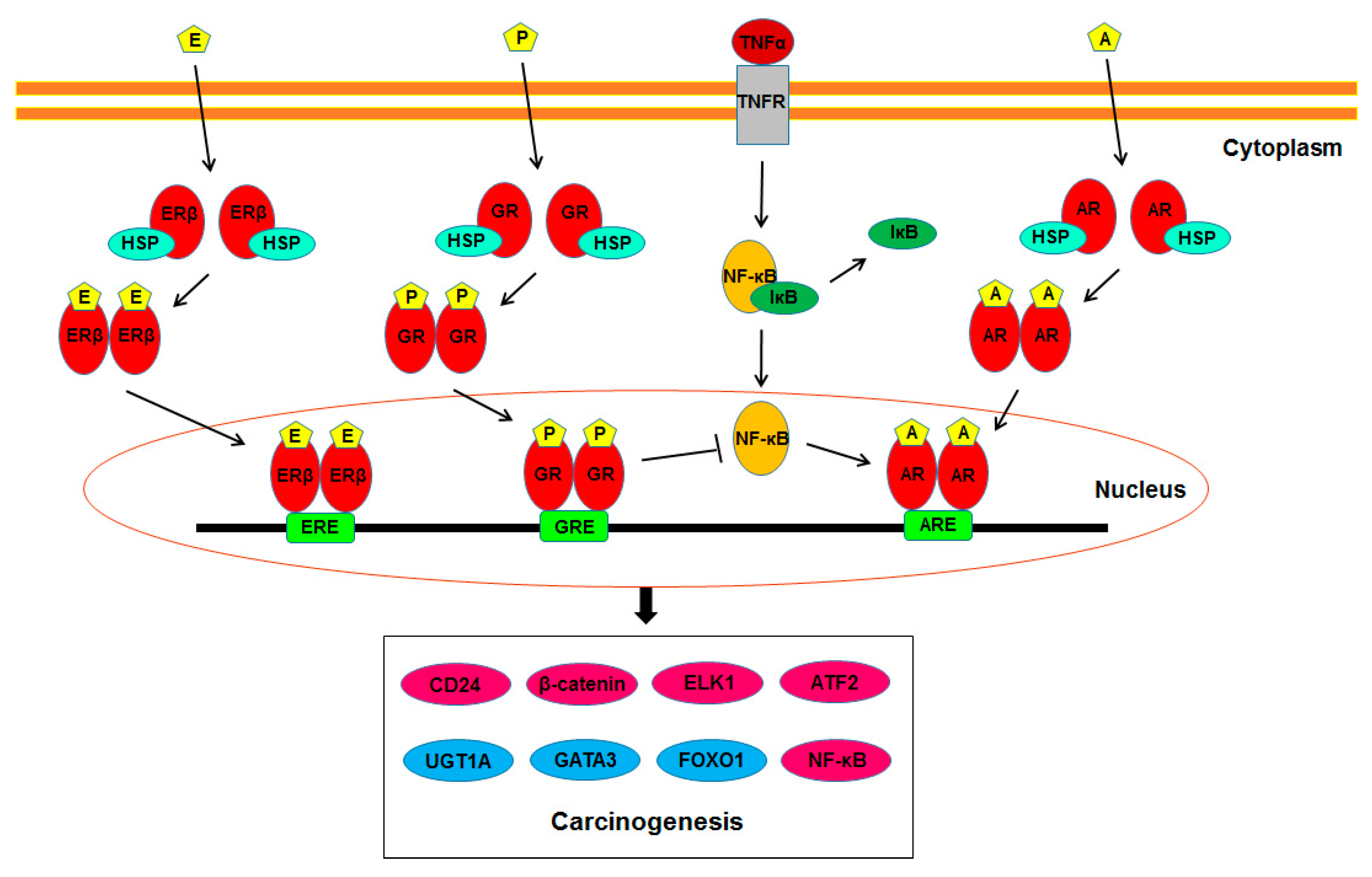
:1. Introduction
2. AR
3. ERs
4. GR
5. PR
6. VDR
7. Molecules Modulated by Steroid Hormone Receptor Signaling in Urothelial Cells
7.1. UDP-Glucuronosyltransferases (UGTs)
7.2. GATA3
7.3. FOXO1
7.4. CD24
7.5. β-Catenin
7.6. ELK1
7.7. ATF2
7.8. NF-κB
8. Conclusions
Funding
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Rouprêt, M.; Babjuk, M.; Compérat, E.; Zigeuner, R.; Sylvester, R.J.; Burger, M.; Cowan, N.C.; Gontero, P.; Van Rhijn, B.W.G.; Mostafid, A.H.; et al. European Association of Urology guidelines on upper urinary tract urothelial carcinoma: 2017 update. Eur. Urol. 2018, 73, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Yokomizo, A.; Takeuchi, A.; Imada, K.; Kiyoshima, K.; Inokuchi, J.; Tatsugami, K.; Ohga, S.; Nakamura, K.; Honda, H.; et al. Secondary bladder cancer after anticancer therapy for prostate cancer: Reduced comorbidity after androgen-deprivation therapy. Oncotarget 2015, 6, 14710–14719. [Google Scholar] [CrossRef] [Green Version]
- Izumi, K.; Taguri, M.; Miyamoto, H.; Hara, Y.; Kishida, T.; Chiba, K.; Murai, T.; Hirai, K.; Suzuki, K.; Fujinami, K.; et al. Androgen deprivation therapy prevents bladder cancer recurrence. Oncotarget 2014, 5, 12665–12674. [Google Scholar] [CrossRef] [Green Version]
- Shiota, M.; Kiyoshima, K.; Yokomizo, A.; Takeuchi, A.; Kashiwagi, E.; Dejima, T.; Takahashi, R.; Inokuchi, J.; Tatsugami, K.; Eto, M. Suppressed recurrent bladder cancer after androgen suppression with androgen deprivation therapy or 5α-reductase inhibitor. J. Urol. 2017, 197, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.; Ito, Y.; Miyamoto, H.; Miyoshi, Y.; Ota, J.; Moriyama, M.; Murai, T.; Hayashi, H.; Inayama, Y.; Ohashi, K.; et al. Expression of androgen receptor in non-muscle-invasive bladder cancer predicts the preventive effect of androgen deprivation therapy on tumor recurrence. Oncotarget 2016, 7, 14153–14160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boorjian, S.; Ugras, S.; Mongan, N.P.; Gudas, L.J.; You, X.; Tickoo, S.K.; Scherr, D.S. Androgen receptor expression is inversely correlated with pathologic tumor stage in bladder cancer. Urology 2004, 64, 383–388. [Google Scholar] [CrossRef]
- Kauffman, E.C.; Robinson, B.D.; Downes, M.J.; Powell, L.G.; Lee, M.M.; Scherr, D.S.; Gudas, L.J.; Mongan, N.P. Role of androgen receptor and associated lysine-demethylase coregulators, LSD1 and JMJD2A, in localized and advanced human bladder cancer. Mol. Carcinog. 2011, 50, 931–944. [Google Scholar] [CrossRef] [Green Version]
- Tuygun, C.; Kankaya, D.; Imamoglu, A.; Sertcelik, A.; Zengin, K.; Oktay, M.; Sertcelik, N. Sex-specific hormone receptors in urothelial carcinomas of the human urinary bladder: A comparative analysis of clinicopathological features and survival outcomes according to receptor expression. Urol. Oncol. 2011, 29, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, H.; Yao, J.L.; Chaux, A.; Zheng, Y.; Hsu, I.; Izumi, K.; Chang, C.; Messing, E.M.; Netto, G.J.; Yeh, S. Expression of androgen and oestrogen receptors and its prognostic significance in urothelial neoplasm of the urinary bladder. BJU Int. 2012, 109, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Mashhadi, R.; Pourmand, G.; Kosari, F.; Mehrsai, A.; Salem, S.; Pourmand, M.R.; Alatab, S.; Khonsari, M.; Heydari, F.; Beladi, L.; et al. Role of steroid hormone receptors in formation and progression of bladder carcinoma: A case-control study. Urol. J. 2014, 11, 1968–1973. [Google Scholar] [PubMed]
- Kashiwagi, E.; Fujita, K.; Yamaguchi, S.; Fushimi, H.; Ide, H.; Inoue, S.; Mizushima, T.; Reis, L.O.; Sharma, R.; Netto, G.J.; et al. Expression of steroid hormone receptors and its prognostic significance in urothelial carcinoma of the upper urinary tract. Cancer Biol. Ther. 2016, 17, 1188–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasui, M.; Kawahara, T.; Takamoto, D.; Izumi, K.; Uemura, H.; Miyamoto, H. Distribution of androgen receptor expression in the urinary bladder. Int. J. Urol. 2019, 26, 305–306. [Google Scholar] [CrossRef] [PubMed]
- Ide, H.; Inoue, S.; Miyamoto, H. Histopathological and prognostic significance of the expression of sex hormone receptors in bladder cancer: A meta-analysis of immunohistochemical studies. PLoS ONE 2017, 12, e0174746. [Google Scholar] [CrossRef] [PubMed]
- Sikic, D.; Breyer, J.; Hartmann, A.; Burger, M.; Erben, P.; Denzinger, D.; Eckstein, M.; Stöhr, R.; Wach, S.; Wullich, B.; et al. High androgen receptor mRNA expression is independently associated with prolonged cabcer-specific and recurrence-free survival in stage T1 bladder cancer. Trans. Oncol. 2017, 10, 340–345. [Google Scholar] [CrossRef]
- Yasui, M.; Kawahara, T.; Izumi, K.; Yao, M.; Ishiguro, Y.; Ishiguro, H.; Uemura, H.; Miyoshi, Y. Androgen receptor mRNA expression is a predictor for recurrence-free survival in non-muscle invasive bladder cancer. BMC Cancer 2019, 19, 331. [Google Scholar] [CrossRef]
- Liu, C.H.; Huang, J.D.; Huang, S.W.; Hour, T.C.; Huang, Y.K.; Hsueh, Y.M.; Chiou, H.Y.; Lee, T.C.; Jan, K.Y.; Chen, C.J.; et al. Androgen receptor gene polymorphism may affect the risk of urothelial carcinoma. J. Biomed. Sci. 2008, 15, 261–269. [Google Scholar] [CrossRef]
- Teng, X.Y.; Liu, G.Q.; Diao, X.L.; Wu, Z.Y.; Li, L.; Zhang, W.; Zhang, X.; Su, Q. CAG repeats in the androgen receptor gene are shorter in patients with pulmonary, esophageal or bladder carcinoma and longer in women with uterine leiomyoma. Oncol. Rep. 2010, 23, 811–818. [Google Scholar]
- Zhuang, Y.H.; Blauer, M.; Tammela, T.; Tuohimaa, P. Immunodetection of androgen receptor in human urinary bladder cancer. Histopathology 1997, 30, 556–562. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Noda, S.; Matsuyama, C.; Shimizu, A.; Kamai, T. Sex steroid hormone receptors in bladder cancer: Usefulness in differential diagnosis and implications in histogenesis of bladder cancer. Urol. Oncol. 2019, 37, 353.e9–353.e15. [Google Scholar] [CrossRef]
- Kontos, S.; Kominea, A.; Melachrinou, M.; Balampani, E.; Sotiropoulou-Bonikou, G. Inverse expression of estrogen receptor-β and nuclear factor-κB in urinary bladder carcinogenesis. Int. J. Urol. 2010, 17, 801–809. [Google Scholar] [CrossRef]
- Ishiguro, H.; Kawahara, T.; Zheng, Y.; Netto, G.J.; Miyamoto, H. Reduced glucocorticoid receptor expression predicts bladder tumor recurrence and progression. Am. J. Clin. Pathol. 2014, 142, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Sahin, M.O.; Canda, A.E.; Yorukoglu, K.; Mungan, M.U.; Sade, M.; Kirkali, Z. 1,25 Dihydroxyvitamin D3 receptor expression in superficial transitional cell carcinoma of the bladder: A possible prognostic factor? Eur. Urol. 2005, 47, 52–57. [Google Scholar]
- Jóźwicki, W.; Brozyna, A.A.; Siekiera, J.; Slominski, A.T. Expression of vitamin D receptor (VDR) positively correlates with survival of urothelial bladder cancer patients. Int. J. Mol. Sci. 2015, 16, 24369–24386. [Google Scholar] [CrossRef] [Green Version]
- Okajima, E.; Hiramatsu, T.; Iriya, K.; Ijuin, M.; Matsushima, S. Effects of sex hormones on development of urinary bladder tumours in rats induced by N-butyl-N-(4-hydroxybutyl) nitrosamine. Urol. Res. 1975, 3, 73–79. [Google Scholar] [CrossRef]
- Imada, S.; Akaza, H.; Ami, Y.; Koiso, K.; Ideyama, Y.; Takenaka, T. Promoting effects and mechanisms of action of androgen in bladder carcinogenesis in male rats. Eur. Urol. 1997, 31, 360–364. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Q.; Huang, X.; Yang, J.; Xu, Y.; Zhang, G. The effects of early versus delayed castration targeting androgen on prolonging survival in a mouse model of bladder cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 10283–10293. [Google Scholar]
- Miyamoto, H.; Yang, Z.; Chen, Y.T.; Ishiguro, H.; Uemura, H.; Kubota, Y.; Nagashima, Y.; Chang, Y.J.; Hu, Y.C.; Tsai, M.Y.; et al. Promotion of bladder cancer development and progression by androgen receptor signals. J. Natl. Cancer Inst. 2007, 99, 558–568. [Google Scholar] [CrossRef]
- Hsu, J.W.; Hsu, I.; Xu, D.; Miyamoto, H.; Liang, L.; Wu, X.R.; Shyr, C.R.; Chang, C. Decreased tumorigenesis and mortality from bladder cancer in mice lacking urothelial androgen receptor. Am. J. Pathol. 2013, 182, 1811–1820. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.T.; Hooker, E.; Luong, R.; Yu, E.J.; He, Y.; Gonzalgo, M.L.; Sun, Z. Conditional expression of the androgen receptor increases susceptibility of bladder cancer in mice. PLoS ONE 2016, 11, e0148851. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ishiguro, H.; Kawahara, T.; Miyamoto, Y.; Izumi, K.; Miyamoto, H. GATA3 in the urinary bladder: Suppression of neoplastic transformation and down-regulation by androgens. Am. J. Cancer Res. 2014, 4, 461–473. [Google Scholar]
- Kawahara, T.; Inoue, S.; Kashiwagi, E.; Chen, J.; Ide, H.; Mizushima, T.; Li, Y.; Zheng, Y.; Miyamoto, H. Enzalutamide as an androgen receptor inhibitor prevents urothelial tumorigenesis. Am. J. Cancer Res. 2017, 7, 2041–2050. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Ide, H.; Mizushima, T.; Jiang, G.; Kawahara, T.; Miyamoto, H. ELK1 promotes urothelial tumorigenesis in the presence of an activated androgen receptor. Am. J. Cancer Res. 2018, 8, 2325–2336. [Google Scholar]
- Shen, S.S.; Smith, C.L.; Hsieh, J.T.; Yu, J.; Kim, I.Y.; Jian, W.; Sonpavde, G.; Ayala, G.E.; Younes, M.; Lerner, S.P. Expression of estrogen receptors-alpha and -beta in bladder cancer cell lines and human bladder tumor tissue. Cancer 2006, 106, 2610–2616. [Google Scholar] [CrossRef]
- Teng, J.; Wang, Z.Y.; Jarrard, D.F.; Bjorling, D.E. Roles of estrogen receptor α and β in modulating urothelial cell proliferation. Endocr. Relat. Cancer 2008, 15, 351–364. [Google Scholar] [CrossRef]
- Sanchez-Carbayo, M. Hypermethylation in bladder cancer: Biological pathways and translational applications. Tumour Biol. 2012, 33, 347–361. [Google Scholar] [CrossRef]
- Brait, M.; Begum, S.; Carvalho, A.L.; Dasgupta, S.; Vettore, A.L.; Czerniak, B.; Caballero, O.L.; Westra, W.H.; Sidransky, D.; Hoque, M.O. Aberrant promoter methylation of multiple genes during pathogenesis of bladder cancer. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2786–2794. [Google Scholar] [CrossRef] [Green Version]
- Reid, L.M.; Leav, I.; Kwan, P.W.; Russell, P.; Merk, F.B. Characterization of a human, sex steroid-responsive transitional cell carcinoma maintained as a tumor line (R198) in athymic nude mice. Cancer Res. 1984, 44, 4560–4573. [Google Scholar]
- Waalkes, M.P.; Liu, J.; Ward, J.M.; Powell, D.A.; Diwan, B.A. Urogenital carcinogenesis in female CD1 mice induced by in utero arsenic exposure is exacerbated by postnatal diethylstilbestrol treatment. Cancer Res. 2006, 66, 1337–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, I.; Yeh, C.R.; Slavin, S.; Miyamoto, H.; Netto, G.J.; Tsai, Y.C.; Muyan, M.; Wu, X.R.; Messing, E.M.; Guancial, E.A.; et al. Estrogen receptor alpha prevents bladder cancer via INPP4B inhibited akt pathway in vitro and in vivo. Oncotarget 2014, 5, 7917–7935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, I.; Chuang, K.L.; Slavin, S.; Da, J.; Lim, W.X.; Pang, S.T.; O’Brien, J.H.; Yeh, S. Suppression of ERβ signaling via ERβ knockout or antagonist protects against bladder cancer development. Carcinogenesis 2014, 35, 651–661. [Google Scholar] [CrossRef] [PubMed]
- George, S.K.; Tovar-Sepulveda, V.; Shen, S.S.; Jian, W.; Zhang, Y.; Hilsenbeck, S.G.; Lerner, S.P.; Smith, C.L. Chemoprevention of BBN-induced bladder carcinogenesis by the selective estrogen receptor modulator tamoxifen. Transl. Oncol. 2013, 6, 244–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, R.; Wada, H.; Ito, K.; Adcock, I.A. Effects of glucocorticoids on gene transcription. Eur. J. Pharmacol. 2004, 500, 51–62. [Google Scholar] [CrossRef]
- Duma, D.; Jewell, C.M.; Cidlowski, J.A. Multiple glucocorticoid receptor isoforms and mechanisms of post-translational modification. J. Steroid Biochem. Mol. Biol. 2006, 102, 11–21. [Google Scholar] [CrossRef]
- Oakley, R.H.; Jewell, C.M.; Yudt, M.R.; Bofetiado, D.M.; Cidlowski, J.A. The dominant negative activity of the human glucocorticoid receptor β isoform. Specificity and mechanisms of action. J. Biol. Chem. 1999, 274, 27857–27866. [Google Scholar] [CrossRef] [Green Version]
- Ide, H.; Inoue, S.; Miyamoto, H. The role of glucocorticoid receptor signaling in bladder cancer progression. Cancers 2018, 10, 484. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, K.; Schned, A.; Fortuny, J.; Heaney, J.; Marsit, C.; Kelsey, K.T.; Karagas, M.R. Glucocorticoid therapy and risk of bladder cancer. Br. J. Cancer 2009, 101, 1316–1320. [Google Scholar] [CrossRef]
- Ide, H.; Inoue, S.; Mizushima, T.; Kashiwagi, E.; Zheng, Y.; Miyamoto, H. Role of glucocorticoid signaling in urothelial tumorigenesis: Inhibition by prednisone presumably through inducing glucocorticoid receptor transrepression. Mol. Carcinog. 2019, 58, 2297–2305. [Google Scholar] [CrossRef]
- Tanner, T.M.; Verrijdt, G.; Rombauts, W.; Louw, A.; Hapgood, J.P.; Claessens, F. Anti-androgenic properties of compound A, an analog of a non-steroidal plant compound. Mol. Cell. Endocrinol. 2003, 201, 155–164. [Google Scholar] [CrossRef]
- Ide, H.; Inoue, S.; Mizushima, T.; Jiang, G.; Nagata, Y.; Goto, T.; Kashiwagi, E.; Miyamoto, H. Compound A inhibits urothelial tumorigenesis via both the androgen receptor and glucocorticoid receptor signaling pathways. Am. J. Transl. Res. 2020, 12, 1779–1788. [Google Scholar] [PubMed]
- Zheng, Y.; Ishiguro, H.; Ide, H.; Inoue, S.; Kashiwagi, E.; Kawahara, T.; Jalalizadeh, M.; Reis, L.O.; Miyamoto, H. Compound A inhibits bladder cancer growth predominantly via glucocorticoid receptor transrepression. Mol. Endocrinol. 2015, 29, 1486–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.M.; O’Connell, M.J.; Messing, E.M.; Reeder, J.E. Decreased bladder cancer growth in parous mice. Urology 2008, 72, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, B.J.; Amr, S.; Ezzat, S.; Saleh, D.; Gouda, I.; Loay, I.; Hifnawy, T.; Mikhail, N.N.; Abdel-Hamid, M.; Zhan, M.; et al. Estrogen exposure and bladder cancer risk in Egyptian women. Maturitas 2010, 67, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.P.; Leng, J.Y.; Zhang, R.K.; Zhang, L.; Zhang, B.; Jiang, W.Y.; Tong, L. Functional analysis of gene expression profiling-based prediction in bladder cancer. Oncol. Lett. 2018, 15, 8417–8842. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Huang, J.L.; Qiu, M.X.; Ma, Z.W. Impact of serum vitamin D level on risk of bladder cancer: A systemic review and meta-analysis. Tumour Biol. 2015, 36, 1567–1572. [Google Scholar] [CrossRef]
- Mittal, R.D.; Manchanda, P.K.; Bhat, S.; Bid, H.K. Association of vitamin-D receptor (Fok-I) gene polymorphism with bladder cancer in an Indian population. BJU Int. 2007, 99, 933–937. [Google Scholar] [CrossRef]
- Konety, B.R.; Lavelle, J.P.; Pirtskalaishvili, G.; Dhir, R.; Meyers, S.A.; Nguyen, T.S.; Hershberger, P.; Shurin, M.R.; Johnson, C.S.; Trump, D.L.; et al. Effects of vitamin D (calcitriol) on transitional cell carcinoma of the bladder in vitro and in vivo. J. Urol. 2001, 165, 253–258. [Google Scholar] [CrossRef]
- Izumi, K.; Zheng, Y.; Hsu, J.W.; Chang, C.; Miyamoto, H. Androgen receptor signals regulate UDP-glucuronosyltransferases in the urinary bladder: A potential mechanism of androgen-induced bladder carcinogenesis. Mol. Carcinog. 2013, 52, 94–102. [Google Scholar] [CrossRef]
- Izumi, K.; Li, Y.; Ishiguro, H.; Zheng, Y.; Yao, J.L.; Netto, G.J.; Miyamoto, H. Expression of UDP-glucuronosyltransferase 1A in bladder cancer: Association with prognosis and regulation by estrogen. Mol. Carcinog. 2014, 53, 314–324. [Google Scholar] [CrossRef]
- Izumi, K.; Inoue, S.; Ide, H.; Fujita, K.; Mizushima, T.; Jiang, G.; Yamaguchi, S.; Fushimi, H.; Nonomura, N.; Miyamoto, H. Uridine 5′diphospho-glucuronosyltransferase 1A expression as an independent prognosticator in urothelial carcinoma of the upper urinary tract. Int. J. Urol. 2018, 25, 429–435. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Klaene, J.J.; Li, Y.; Paonessa, J.D.; Stablewski, A.B.; Vouros, P.; Zhang, Y. The inverse relationship between bladder and liver in 4-aminobiphenyl-induced DNA damage. Oncotarget 2015, 6, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.B.; Trpkov, K.; Lopez-Beltran, A.; Grignon, D.; Members of the ISUP Immunohistochemistry in Diagnostic Urologic Pathology Group. Best practices recommendations in the application of immunohistochemistry in the bladder lesions: Report from the International Society of Urologic Pathology consensus conference. Am. J. Surg. Pathol. 2014, 38, e20–e34. [Google Scholar] [CrossRef]
- Miyamoto, H.; Izumi, K.; Yao, J.L.; Li, Y.; Yang, Q.; McMahon, L.A.; Gonzalez-Roibon, N.; Hicks, D.G.; Netto, G.J. GATA binding protein 3 is down-regulated in bladder cacner yet strong expression is an independent predictor of poor prognosis in invasive tumor. Hum. Pathol. 2012, 43, 2033–2040. [Google Scholar] [CrossRef]
- Inoue, S.; Mizushima, T.; Fujita, K.; Meliti, A.; Ide, H.; Yamaguchi, S.; Fushimi, H.; Netto, G.J.; Nonomura, N.; Miyamoto, H. GATA3 immunohistochemistry in urothelial carcinoma of the upper urinary tract as a urothelial marker and a prognosticator. Hum. Pathol. 2017, 64, 83–90. [Google Scholar] [CrossRef]
- Ide, H.; Mizushima, T.; Jiang, G.; Goto, T.; Nagata, Y.; Teramoto, Y.; Inoue, S.; Li, Y.; Kashiwagi, E.; Baras, A.S.; et al. FOXO1 as a tumor suppressor inactivated via AR/ERβ signals in urothelial cells. Endocr. Relat. Cancer 2020, 27, 231–244. [Google Scholar] [CrossRef]
- Ide, H.; Jiang, G.; Mizushima, T.; Fujita, K.; Inoue, S.; Yamaguchi, S.; Fushimi, H.; Nonomura, N.; Miyamoto, H. Forkhead box O1 as an indicator of prognosis is inactivated in urothelial carcinoma of the upper urinary tract. Oncol. Lett. 2019, 17, 482–487. [Google Scholar] [CrossRef]
- Ooki, A.; VandenBussche, C.J.; Kates, M.; Hahn, N.M.; Matoso, A.; McConkey, D.J.; Bivalacqua, T.J.; Hoque, M.O. CD24 regulates cancer stem cell (CSC)-like traits and a panel of CSC-related molecules serves as a non-invasive urinary biomarker for the detection of bladder cancer. Br. J. Cancer 2018, 119, 961–970. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, S.; Shen, H.; Xu, K.; Chen, J.; Li, H.; Xu, Y.; Xu, A.; Chen, B.; Kaku, H.; et al. Clinical significance of CD24 as a predictor of bladder cancer recurrence. Oncol. Lett. 2013, 6, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Overdevest, J.B.; Knubel, K.H.; Duex, J.E.; Thomas, S.; Nitz, M.D.; Harding, M.A.; Smith, S.C.; Frierson, H.F.; Conaway, M.; Theodorescu, D. CD24 expression is important in male urothelial tumorigenesis and metastasis in mice and is androgen regulated. Proc. Natl. Acad. Sci. USA 2012, 109, E3588–E3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, N.; Dancik, G.M.; Goodspeed, A.; Costello, J.C.; Owens, C.; Duex, J.E.; Theodorescu, D. GON4L drives cancer growth through a YY1-androgen receptor-CD24 axis. Cancer Res. 2016, 76, 5175–5185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, H.; Igawa, M.; Shigeno, K.; Terashima, M.; Deguchi, M.; Yamanaka, M.; Ribeiro-Filho, L.; Kane, C.J.; Dahiya, R. β-Catenin mutations correlate with over expression of c-myc and cyclin D1 genes in bladder cancer. J. Urol. 2002, 168, 2220–2226. [Google Scholar] [CrossRef]
- Lin, C.; Yin, Y.; Stemler, K.; Humphrey, P.; Kibel, A.S.; Mysorekar, I.U.; Ma, L. Constitutive β-catenin activation induces male-specific tumorigenesis in the bladder urothelium. Cancer Res. 2013, 73, 5914–5925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, Y.; Izumi, K.; Ishiguro, H.; Ye, B.; Li, F.; Miyamoto, H. Androgen activates beta-catenin signaling in bladder cancer cells. Endocr. Relat. Cancer 2013, 20, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, T.; Shareef, H.K.; Aljarah, A.K.; Ide, H.; Li, Y.; Kashiwagi, E.; Netto, G.J.; Zheng, Y.; Miyamoto, H. ELK1 is up-regulated by androgen in bladder cancer cells and promotes tumor progression. Oncotarget 2015, 6, 29860–29876. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Ide, H.; Fujita, K.; Mizushima, T.; Jiang, G.; Kawahara, T.; Yamaguchi, S.; Fushimi, H.; Nonomura, N.; Miyamoto, H. Expression of phospho-ELK1 and its prognostic significance in urothelial carcinoma of the upper urinary tract. Int. J. Mol. Sci. 2018, 19, 777. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, T.; Ide, H.; Kashiwagi, E.; Patterson, J.D.; Inoue, S.; Shareef, H.K.; Aljarah, A.K.; Zheng, Y.; Barasm, A.S.; Miyamoto, H. Silodosin inhibits the growth of bladder cancer cells and enhances the cytotoxic activity of cisplatin via ELK1 inactivation. Am. J. Cancer Res. 2015, 5, 2959–2968. [Google Scholar]
- Inoue, S.; Mizushima, T.; Ide, H.; Jiang, G.; Goto, T.; Nagata, Y.; Netto, G.J.; Miyamoto, H. ATF2 promotes urothelial cancer outgrowth via cooperation with androgen receptor signaling. Endocr. Connect. 2018, 7, 1397–1408. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Ide, H.; Mizushima, T.; Jiang, G.; Netto, G.J.; Gotoh, M.; Miyamoto, H. Nuclear factor-κB promotes urothelial tumorigenesis and cancer progression via cooperation with androgen receptor signaling. Mol. Cancer Ther. 2018, 17, 1303–1314. [Google Scholar] [CrossRef] [Green Version]
- Nelius, T.; Filleur, S.; Yemelyanov, A.; Budunova, I.; Shroff, E.; Mirochnik, Y.; Aurora, A.; Veliceasa, D.; Xiao, W.; Wang, Z.; et al. Androgen receptor targets NFκB and TSP1 to suppress prostate tumor growth in vivo. Int. J. Cancer 2007, 121, 999–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Altuwaijri, S.; Deng, F.; Chen, L.; Lal, P.; Bhanot, U.K.; Korets, R.; Wenske, S.; Lilja, H.G.; Chang, C.; et al. NF-κB regulates androgen receptor expression and prostate cancer growth. Am. J. Pathol. 2009, 175, 489–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, N.; Cheng, H.; Lin, D.; Liu, L.; Yang, O.; Jia, L.; Fazli, L.; Gleave, M.E.; Wang, Y.; Rennie, P.; et al. The expression of glucocorticoid receptor is negatively regulated by active androgen receptor signaling in prostate tumors. Int. J. Cancer 2015, 136, E27–E38. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author, Year [Reference] | Receptor | Tumor Site | Positive/Total Cases | ||
|---|---|---|---|---|---|
| Non-Tumor | Tumor | p Value | |||
| Boorjian, 2004 [8] | AR | Bladder | 32/37 (86%) | 26/49 (53%) | 0.001 * |
| Kauffman, 2011 [9] | AR | Bladder | 50/59 (84%) | 30/59 (51%) | <0.001 |
| Tuygun, 2011 [10] | AR | Bladder | 0/58 (0%) (Male) | 71/139 (51%) | <0.001 * |
| Miyamoto, 2012 [11] | AR | Bladder | 113/141 (80%) | 79/188 (42%) | <0.001 |
| Mashhadi, 2014 [12] | AR | Bladder | 0/132 (0%) | 26/120 (22%) | <0.001 |
| Kashiwagi, 2015 [13] | AR | UUT | 46/80 (58%) | 20/99 (20%) | <0.001 |
| Miyamoto, 2012 [11] | ERα | Bladder | 70/141 (50%) | 51/188 (27%) | <0.001 |
| Mashhadi, 2014 [12] | ERα | Bladder | 2/132 (2%) | 3/120 (3%) | 0.67 |
| Kashiwagi, 2015 [13] | ERα | UUT | 32/80 (40%) | 18/99 (18%) | 0.001 |
| Imai, 2019 [22] | ERα | Bladder | 33/92 (36%) | 48/125 (38%) | 0.777 * |
| Kontos, 2010 [23] | ERβ | Bladder | 27/29 (93%) | 84/111 (76%) | 0.041 * |
| Miyamoto, 2012 [11] | ERβ | Bladder | 125/141 (89%) | 93/188 (49%) | <0.001 |
| Kashiwagi, 2015 [13] | ERβ | UUT | 68/80 (85%) | 62/99 (63%) | 0.001 |
| Ishiguro,2014 [24] | GR | Bladder | 90/94 (96%) | 129/149 (87%) | 0.026 |
| Kashiwagi, 2015 [13] | GR | UUT | 67/80 (84%) | 62/99 (63%) | 0.001 |
| Mashhadi, 2014 [12] | PR | Bladder | 3/132 (2%) | 5/120 (4%) | 0.48 |
| Kashiwagi, 2015 [13] | PR | UUT | 10/80 (13%) | 16/99 (16%) | 0.487 |
| Imai, 2019 [22] | PR | Bladder | 1/92 (1%) | 4/125 (3%) | 0.398 * |
| Sahin, 2005 [25] | VDR | Bladder | 70/105 (67%) | 90/105 (86%) | 0.02 |
| Jóźwicki, 2015 [26] | VDR | Bladder | 12/12 (100%) | 62/71 (87%) | 0.345 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ide, H.; Miyamoto, H. The Role of Steroid Hormone Receptors in Urothelial Tumorigenesis. Cancers 2020, 12, 2155. https://doi.org/10.3390/cancers12082155
Ide H, Miyamoto H. The Role of Steroid Hormone Receptors in Urothelial Tumorigenesis. Cancers. 2020; 12(8):2155. https://doi.org/10.3390/cancers12082155
Chicago/Turabian StyleIde, Hiroki, and Hiroshi Miyamoto. 2020. "The Role of Steroid Hormone Receptors in Urothelial Tumorigenesis" Cancers 12, no. 8: 2155. https://doi.org/10.3390/cancers12082155
APA StyleIde, H., & Miyamoto, H. (2020). The Role of Steroid Hormone Receptors in Urothelial Tumorigenesis. Cancers, 12(8), 2155. https://doi.org/10.3390/cancers12082155