Flavonoids Targeting HIF-1: Implications on Cancer Metabolism

 , ,
, ,  ,
,  , ,
, ,  ,
, 
 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Aim of the Study
1.2. Source of the Data
2. Hypoxic Conditions
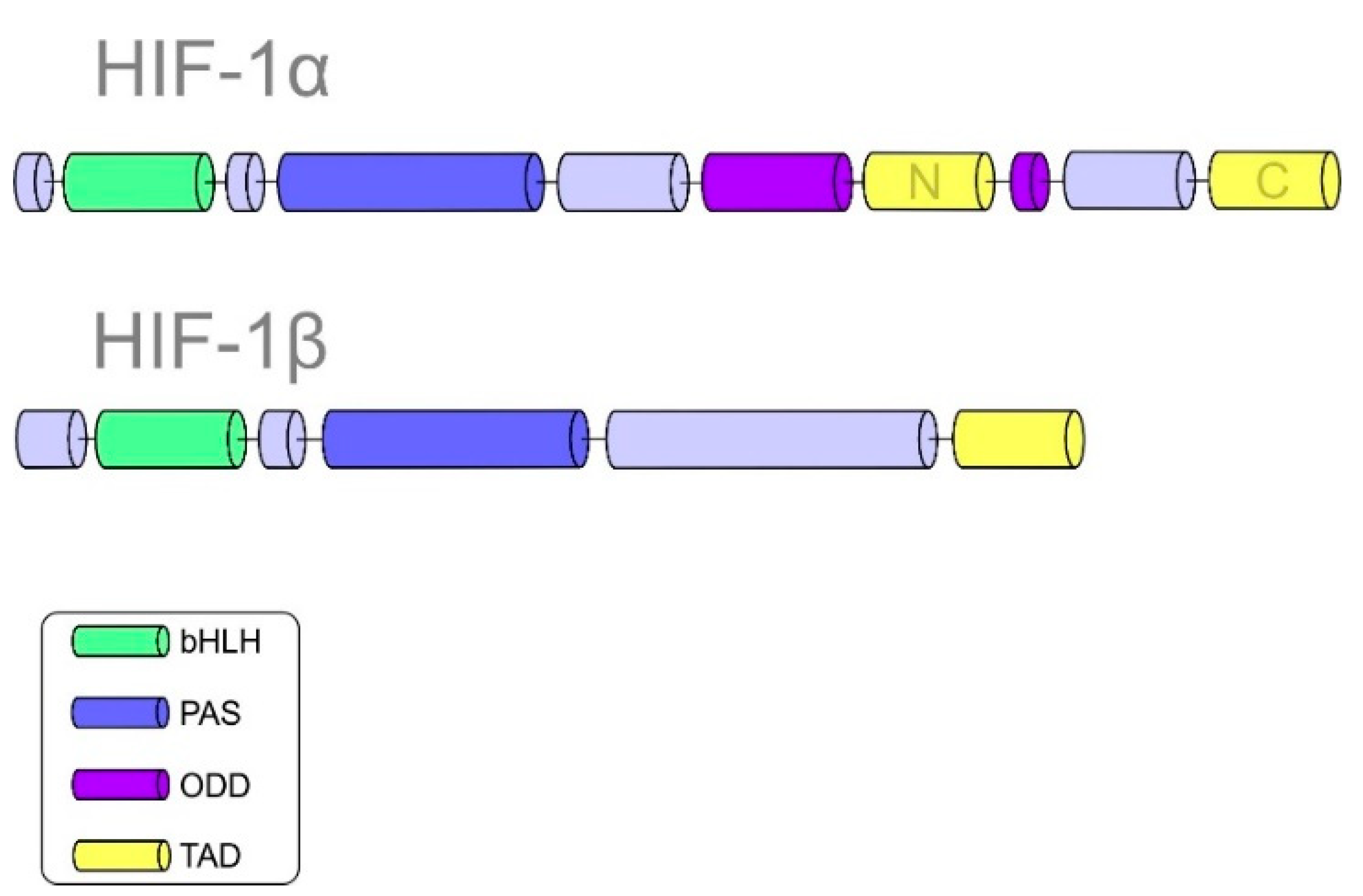
2.1. Structure of Hypoxia-Inducible Factor 1
2.2. Regulation of HIF-1
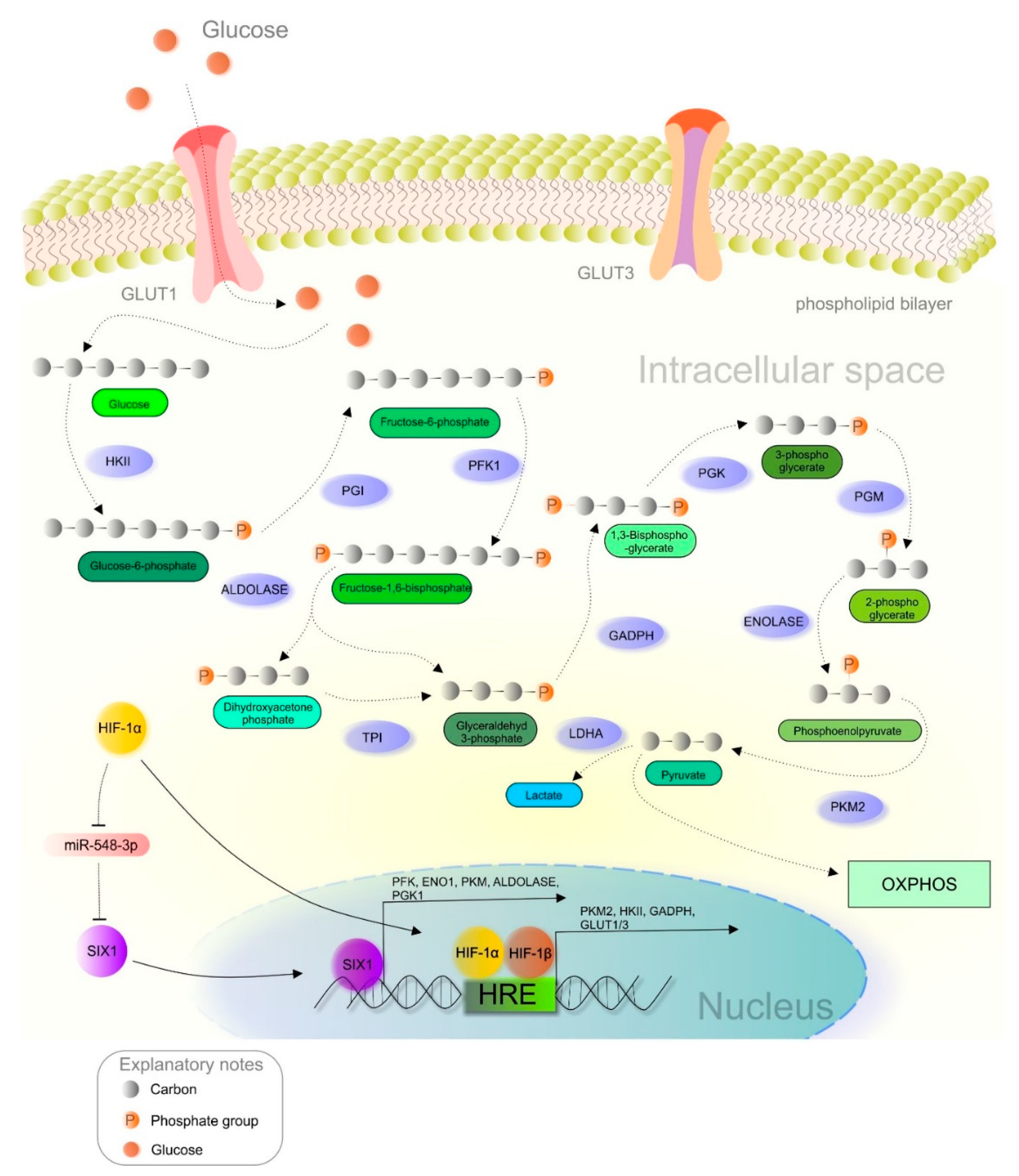
2.3. A Brief Introduction to the Warburg Phenotype
2.4. Implementation of HIF-1 in the Modulation of Cancer Metabolism
2.5. Therapeutic Interventions Based on HIF-1 Regulation: Current Status and Future Directions
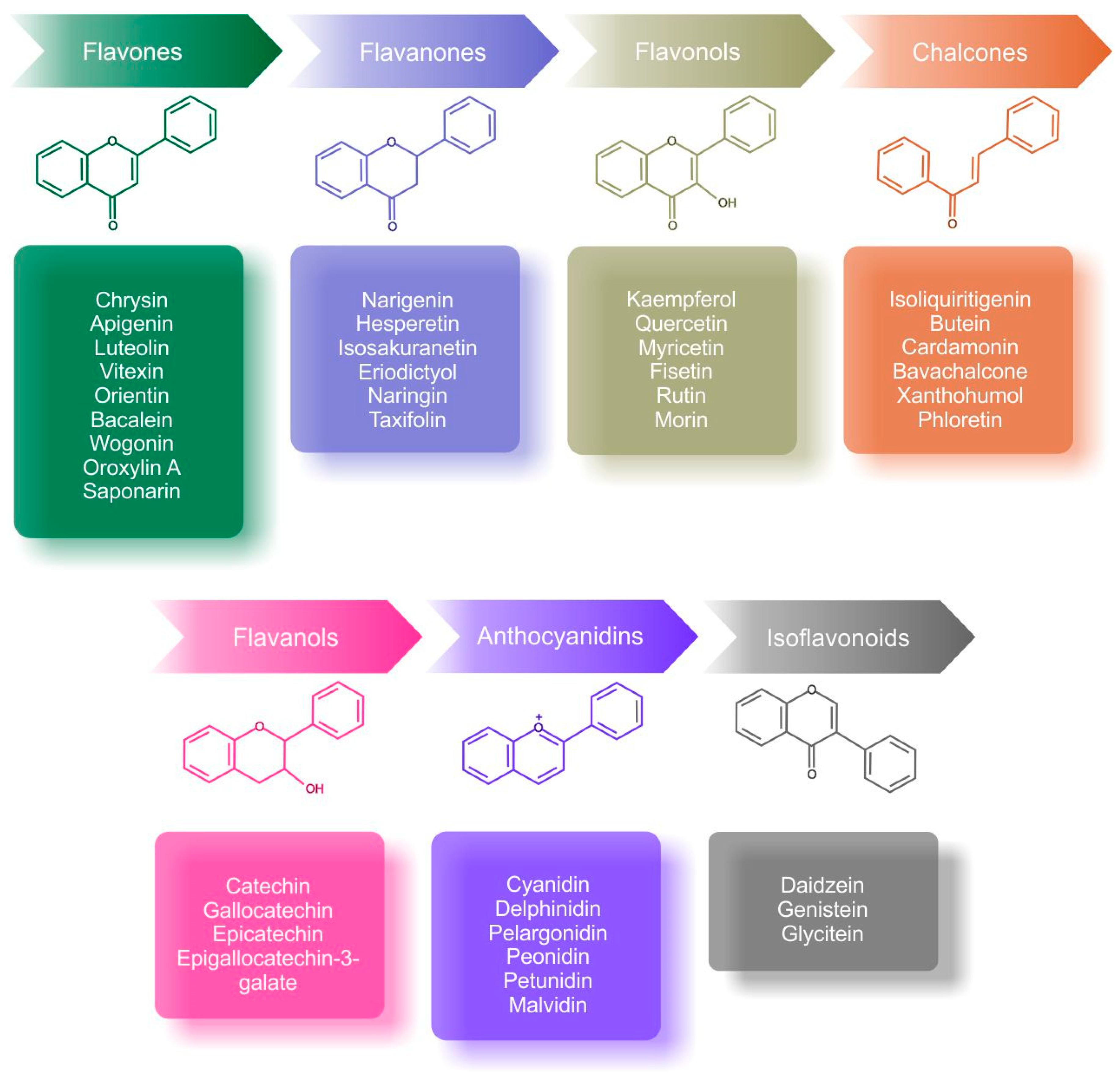
3. Beneficial Effects of Flavonoids in the Regulation of Hypoxic Molecular Cascades and the Warburg Effect
3.1. Regulation of HIF-1 Activity by Flavonoids
3.2. Flavonoids Targeting HIF-1 and Glucose Metabolism: Connections between Hypoxia and the Warburg Effect
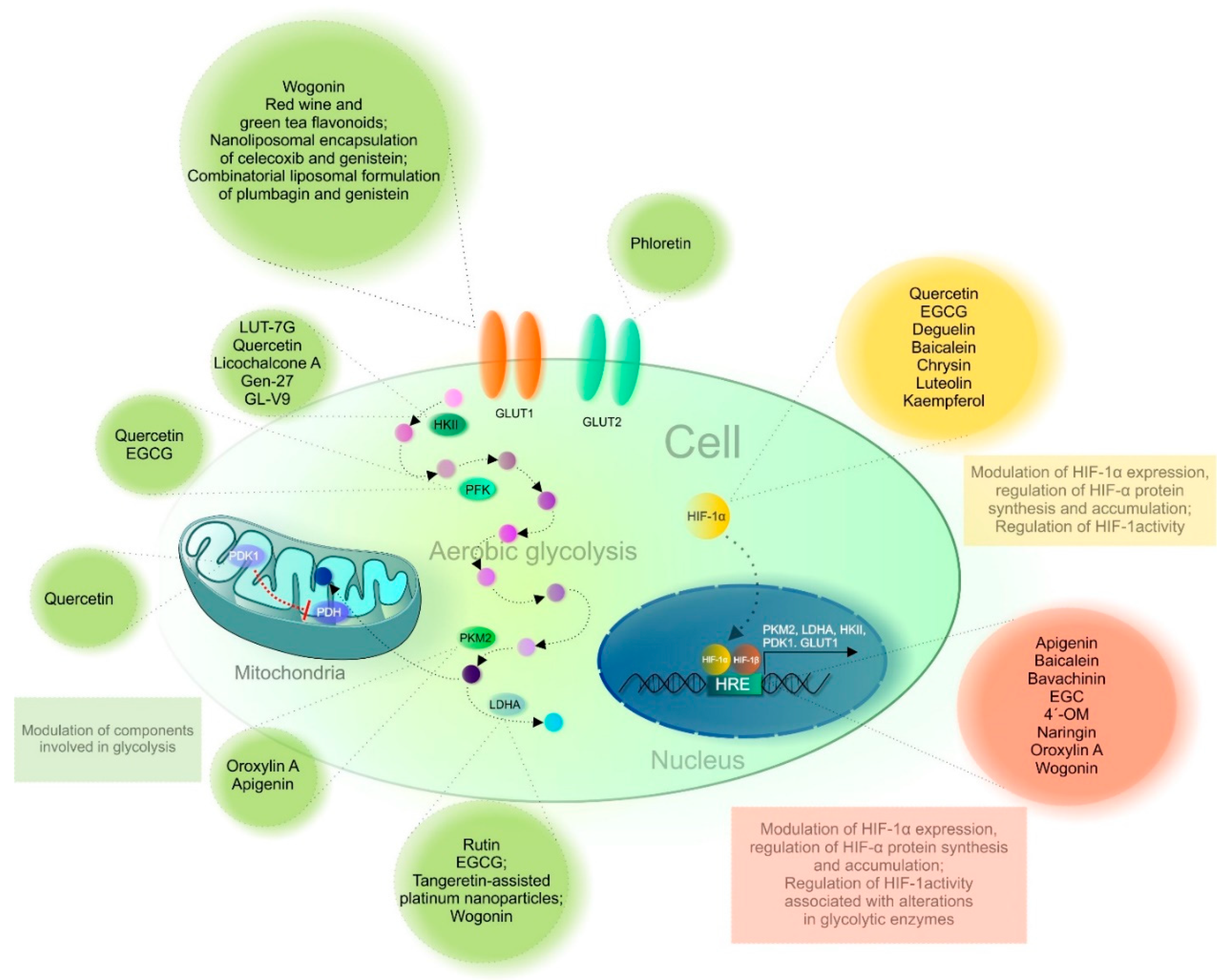
3.3. Anticancer Effects of Flavonoids Mediated through Glucose Transporters and Enzymes of Glucose Metabolism
3.3.1. Glucose Transporter (GLUT)
3.3.2. Hexokinase II
3.3.3. Pyruvate Kinase
3.3.4. Lactate Dehydrogenase
3.3.5. Phosphofructokinase-1
3.3.6. Pyruvate Dehydrogenase Kinase
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Biemar, F.; Foti, M. Global Progress against Cancer—Challenges and Opportunities. Cancer Biol. Med. 2013, 10, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Kim, J.H.; Prasad, S.; Aggarwal, B.B. Regulation of Survival, Proliferation, Invasion, Angiogenesis, and Metastasis of Tumor Cells Through Modulation of Inflammatory Pathways by Nutraceuticals. Cancer Metastasis Rev. 2010, 29, 405–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Peng, Y.; Zhao, J.; Lei, X.; Zheng, X.; Xie, Z.; Tang, G. Anticancer Activity of Natural Flavonoids: Inhibition of HIF-1α Signaling Pathway. Curr. Org. Chem. 2020, 23, 2945–2959. [Google Scholar] [CrossRef]
- Masoud, G.N.; Li, W. HIF-1α Pathway: Role, Regulation and Intervention for Cancer Therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Jögi, A.; Ehinger, A.; Hartman, L.; Alkner, S. Expression of HIF-1α Is Related to a Poor Prognosis and Tamoxifen Resistance in Contralateral Breast Cancer. PLoS ONE 2019, 14, e0226150. [Google Scholar] [CrossRef] [Green Version]
- Pezzuto, A.; Carico, E. Role of HIF-1 in Cancer Progression: Novel Insights. Available online: https://www.eurekaselect.com/167204/article (accessed on 4 November 2020).
- Nagao, A.; Kobayashi, M.; Koyasu, S.; Chow, C.C.T.; Harada, H. HIF-1-Dependent Reprogramming of Glucose Metabolic Pathway of Cancer Cells and Its Therapeutic Significance. Int. J. Mol. Sci. 2019, 20, 238. [Google Scholar] [CrossRef] [Green Version]
- Al Tameemi, W.; Dale, T.P.; Al-Jumaily, R.M.K.; Forsyth, N.R. Hypoxia-Modified Cancer Cell Metabolism. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Kozłowska, A.; Szostak-Wegierek, D. Flavonoids—Food Sources and Health Benefits. Rocz. Państwowego Zakładu Hig. 2014, 65, 79–85. [Google Scholar]
- Samec, M.; Liskova, A.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Buhrmann, C.; Varghese, E.; Abotaleb, M.; Qaradakhi, T.; Zulli, A.; et al. Flavonoids against the Warburg Phenotype—Concepts of Predictive, Preventive and Personalised Medicine to Cut the Gordian Knot of Cancer Cell Metabolism. EPMA J. 2020, 11, 377–398. [Google Scholar] [CrossRef]
- Liskova, A.; Koklesova, L.; Samec, M.; Smejkal, K.; Samuel, S.M.; Varghese, E.; Abotaleb, M.; Biringer, K.; Kudela, E.; Danko, J.; et al. Flavonoids in Cancer Metastasis. Cancers 2020, 12, 1498. [Google Scholar] [CrossRef] [PubMed]
- Liskova, A.; Koklesova, L.; Samec, M.; Varghese, E.; Abotaleb, M.; Samuel, S.M.; Smejkal, K.; Biringer, K.; Petras, M.; Blahutova, D.; et al. Implications of Flavonoids as Potential Modulators of Cancer Neovascularity. J. Cancer Res. Clin. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Carnero, A.; Lleonart, M. The Hypoxic Microenvironment: A Determinant of Cancer Stem Cell Evolution. Cell 2016, 1, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Finger, E.C.; Giaccia, A.J. Hypoxia, Inflammation, and the Tumor Microenvironment in Metastatic Disease. Cancer Metastasis Rev. 2010, 29, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Lequeux, A.; Noman, M.Z.; Xiao, M.; Sauvage, D.; Van Moer, K.; Viry, E.; Bocci, I.; Hasmim, M.; Bosseler, M.; Berchem, G.; et al. Impact of Hypoxic Tumor Microenvironment and Tumor Cell Plasticity on the Expression of Immune Checkpoints. Cancer Lett. 2019, 458, 13–20. [Google Scholar] [CrossRef]
- Rankin, E.B.; Nam, J.-M.; Giaccia, A.J. Hypoxia: Signaling the Metastatic Cascade. Trends Cancer 2016, 2, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, K. HIF-1 at the Crossroads of Hypoxia, Inflammation, and Cancer. Int. J. Cancer 2016, 138, 1058–1066. [Google Scholar] [CrossRef]
- Burslem, G.M.; Kyle, H.F.; Nelson, A.; Edwards, T.A.; Wilson, A.J. Hypoxia Inducible Factor (HIF) as a Model for Studying Inhibition of Protein-Protein Interactions. Chem. Sci. 2017, 8, 4188–4202. [Google Scholar] [CrossRef] [Green Version]
- Courtnay, R.; Ngo, D.C.; Malik, N.; Ververis, K.; Tortorella, S.M.; Karagiannis, T.C. Cancer Metabolism and the Warburg Effect: The Role of HIF-1 and PI3K. Mol. Biol. Rep. 2015, 42, 841–851. [Google Scholar] [CrossRef]
- Feng, Z.; Zou, X.; Chen, Y.; Wang, H.; Duan, Y.; Bruick, R.K. Modulation of HIF-2α PAS-B Domain Contributes to Physiological Responses. Proc. Natl. Acad. Sci. USA 2018, 115, 13240–13245. [Google Scholar] [CrossRef] [Green Version]
- Ziello, J.E.; Jovin, I.S.; Huang, Y. Hypoxia-Inducible Factor (HIF)-1 Regulatory Pathway and Its Potential for Therapeutic Intervention in Malignancy and Ischemia. Yale J. Biol. Med. 2007, 80, 51–60. [Google Scholar] [PubMed]
- Zhao, L.; Liu, Z.; Yang, F.; Zhang, Y.; Xue, Y.; Miao, H.; Liao, X.; Huang, H.; Li, G. Intrabody against Prolyl Hydroxylase 2 Promotes Angiogenesis by Stabilizing Hypoxia-Inducible Factor-1α. Sci. Rep. 2019, 9, 11861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Xiao, Z.; Yang, L.; Gao, Y.; Zhu, Q.; Hu, L.; Huang, D.; Xu, Q. Hypoxia-Inducible Factors in Hepatocellular Carcinoma (Review). Oncol. Rep. 2020, 43, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-Y.; He, Y.-L.; Zhu, L.-L. Possible Role of PHD Inhibitors as Hypoxia-Mimicking Agents in the Maintenance of Neural Stem Cells’ Self-Renewal Properties. Front. Cell Dev. Biol. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Snell, C.E.; Turley, H.; McIntyre, A.; Li, D.; Masiero, M.; Schofield, C.J.; Gatter, K.C.; Harris, A.L.; Pezzella, F. Proline-Hydroxylated Hypoxia-Inducible Factor 1α (HIF-1α) Upregulation in Human Tumours. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Zhang, C.; Polavaram, N.; Liu, F.; Wu, G.; Schroeder, M.A.; Lau, J.S.; Mukhopadhyay, D.; Jiang, S.-W.; O’Neill, B.P.; et al. The Role of Factor Inhibiting HIF (FIH-1) in Inhibiting HIF-1 Transcriptional Activity in Glioblastoma Multiforme. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Kataoka, N. Regulation of Gene Expression under Hypoxic Conditions. Int. J. Mol. Sci. 2019, 20, 3278. [Google Scholar] [CrossRef] [Green Version]
- Iommarini, L.; Porcelli, A.M.; Gasparre, G.; Kurelac, I. Non-Canonical Mechanisms Regulating Hypoxia-Inducible Factor 1 Alpha in Cancer. Front. Oncol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.V.; Hubbi, M.E.; Pan, F.; McDonald, K.R.; Mansharamani, M.; Cole, R.N.; Liu, J.O.; Semenza, G.L. Calcineurin Promotes Hypoxia-Inducible Factor 1α Expression by Dephosphorylating RACK1 and Blocking RACK1 Dimerization. J. Biol. Chem. 2007, 282, 37064–37073. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Arora, R.; Kaur, P.; Singh, B.; Mannan, R.; Arora, S. Overexpression of Hypoxia-Inducible Factor and Metabolic Pathways: Possible Targets of Cancer. Cell Biosci. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Tran, L.; Park, Y.; Chen, I.; Lan, J.; Xie, Y.; Semenza, G.L. Reciprocal Regulation of DUSP9 and DUSP16 Expression by HIF1 Controls ERK and P38 MAP Kinase Activity and Mediates Chemotherapy-Induced Breast Cancer Stem Cell Enrichment. Cancer Res. 2018, 78, 4191–4202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yao, L.; Yang, J.; Wang, Z.; Du, G. PI3K/Akt and HIF-1 Signaling Pathway in Hypoxia-Ischemia. Mol. Med. Rep. 2018, 18, 3547–3554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, E.D.; Semenchenko, K.; Wasylyk, B. Crosstalk between Mdm2, P53 and HIF1-α: Distinct Responses to Oxygen Stress and Implications for Tumour Hypoxia. Subcell. Biochem. 2014, 85, 199–214. [Google Scholar] [CrossRef]
- Kataria, N.; Kerr, B.; Zaiter, S.S.; McAlpine, S.; Cook, K.M. C-Terminal HSP90 Inhibitors Block the HSP90:HIF-1α Interaction and Inhibit the Cellular Hypoxic Response. bioRxiv 2019, 521989. [Google Scholar] [CrossRef]
- Luo, W.; Wang, Y. Epigenetic Regulators: Multifunctional Proteins Modulating Hypoxia-Inducible Factor-α Protein Stability and Activity. Cell. Mol. Life Sci. 2018, 75, 1043–1056. [Google Scholar] [CrossRef]
- Batie, M.; Frost, J.; Frost, M.; Wilson, J.W.; Schofield, P.; Rocha, S. Hypoxia Induces Rapid Changes to Histone Methylation and Reprograms Chromatin. Science 2019, 363, 1222–1226. [Google Scholar] [CrossRef]
- Kim, Y.; Nam, H.J.; Lee, J.; Park, D.Y.; Kim, C.; Yu, Y.S.; Kim, D.; Park, S.W.; Bhin, J.; Hwang, D.; et al. Methylation-Dependent Regulation of HIF-1α Stability Restricts Retinal and Tumour Angiogenesis. Nat. Commun. 2016, 7, 10347. [Google Scholar] [CrossRef] [Green Version]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional Regulation by Hypoxia Inducible Factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lafleur, V.N.; Richard, S.; Richard, D.E. Transcriptional Repression of Hypoxia-Inducible Factor-1 (HIF-1) by the Protein Arginine Methyltransferase PRMT1. Mol. Biol. Cell 2014, 25, 925–935. [Google Scholar] [CrossRef]
- Qin, Y.; Hu, Q.; Xu, J.; Ji, S.; Dai, W.; Liu, W.; Xu, W.; Sun, Q.; Zhang, Z.; Ni, Q.; et al. PRMT5 Enhances Tumorigenicity and Glycolysis in Pancreatic Cancer via the FBW7/CMyc Axis. Cell Commun. Signal. 2019, 17, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoepflin, Z.R.; Shapiro, I.M.; Risbud, M.V. Class I and IIa HDACs Mediate HIF-1α Stability Through PHD2-Dependent Mechanism While HDAC6, a Class IIb Member, Promotes HIF-1α Transcriptional Activity in Nucleus Pulposus Cells of the Intervertebral Disc. J. Bone Miner. Res. 2016, 31, 1287–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, S.; Ku, S.; Ciamporcero, E.; Miles, K.M.; Attwood, K.; Chintala, S.; Shen, L.; Ellis, L.; Sotomayor, P.; Swetzig, W.; et al. HDAC 1 and 6 Modulate Cell Invasion and Migration in Clear Cell Renal Cell Carcinoma. BMC Cancer 2016, 16, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, H.; Harvey, C.T.; Pittsenbarger, J.; Liu, Q.; Beer, T.M.; Xue, C.; Qian, D.Z. HDAC4 Protein Regulates HIF1α Protein Lysine Acetylation and Cancer Cell Response to Hypoxia. J. Biol. Chem. 2011, 286, 38095–38102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Sang, N. Histone Deacetylase Inhibitors: The Epigenetic Therapeutics That Repress Hypoxia-Inducible Factors. Available online: https://www.hindawi.com/journals/bmri/2011/197946/ (accessed on 11 October 2020).
- Robinson, C.M.; Lefebvre, F.; Poon, B.P.; Bousard, A.; Fan, X.; Lathrop, M.; Tost, J.; Kim, W.Y.; Riazalhosseini, Y.; Ohh, M. Consequences of VHL Loss on Global DNA Methylome. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.M.; Schmid, S.; Rudolph, T.; Anlauf, M.; Prinz, C.; Klöppel, G.; Moch, H.; Heitz, P.U.; Komminoth, P.; Perren, A. VHL Inactivation Is an Important Pathway for the Development of Malignant Sporadic Pancreatic Endocrine Tumors. Endocr. Relat. Cancer 2009, 16, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Holubekova, V.; Mendelova, A.; Jasek, K.; Mersakova, S.; Zubor, P.; Lasabova, Z. Epigenetic Regulation by DNA Methylation and MiRNA Molecules in Cancer. Future Oncol. 2017, 13, 2217–2222. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yang, C.; Wang, K.; Liu, X.; Liu, Q. MicroRNA-33b Inhibits the Proliferation and Migration of Osteosarcoma Cells via Targeting Hypoxia-Inducible Factor-1α. Oncol. Res. 2017, 25, 397–405. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, L.; Fang, Q.; Sun, J.; Zhang, S.; Zhan, C.; Liu, S.; Zhang, Y. MiR-338-3p Inhibits Hepatocarcinoma Cells and Sensitizes These Cells to Sorafenib by Targeting Hypoxia-Induced Factor 1α. PLoS ONE 2014, 9, e115565. [Google Scholar] [CrossRef] [Green Version]
- Yeh, Y.-M.; Chuang, C.-M.; Chao, K.-C.; Wang, L.-H. MicroRNA-138 Suppresses Ovarian Cancer Cell Invasion and Metastasis by Targeting SOX4 and HIF-1α. Int. J. Cancer 2013, 133, 867–878. [Google Scholar] [CrossRef]
- Hu, Q.; Liu, F.; Yan, T.; Wu, M.; Ye, M.; Shi, G.; Lv, S.; Zhu, X. MicroRNA-576-3p Inhibits the Migration and Proangiogenic Abilities of Hypoxia-treated Glioma Cells Through Hypoxia-Inducible Factor-1α. Int. J. Mol. Med. 2019, 43, 2387–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Zhan, M.; Chen, W.; Xu, S.; Long, M.; Shen, H.; Shi, Y.; Liu, Q.; Mohan, M.; Wang, J. MiR-143-5p Deficiency Triggers EMT and Metastasis by Targeting HIF-1α in Gallbladder Cancer. Cell. Physiol. Biochem. 2017, 42, 2078–2092. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.-M.; Tao, L.; Zhang, M.; Zhang, J.; Liu, X.; Chen, G.-F.; Zhu, Y.-J.; Zhang, P.-J. Clinicopathological Significance of MicroRNA-20b Expression in Hepatocellular Carcinoma and Regulation of HIF-1α and VEGF Effect on Cell Biological Behaviour. Dis. Markers 2015, 2015, 325176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Wang, Z.; Wang, S.; Weng, M.; Zhou, D.; Li, C.; Wang, J.; Chen, E.; Quan, Z. Long Non-Coding RNA LINC00152 Promotes Gallbladder Cancer Metastasis and Epithelial-Mesenchymal Transition by Regulating HIF-1α via MiR-138. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.-Z.; Zhang, L.; Wu, D.; Cai, G.-Y.; He, J.C.; Chen, X.-M. LncRNA HOTAIR Regulates HIF-1α/AXL Signaling Through Inhibition of MiR-217 in Renal Cell Carcinoma. Cell Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.; Zhao, L. LncRNA Nuclear-Enriched Abundant Transcript 1 Promotes Cell Proliferation and Invasion by Targeting MiR-186-5p/HIF-1α in Osteosarcoma. J. Cell. Biochem. 2019, 120, 6502–6514. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Liu, H.W.; Chen, J.Q.; Wang, S.H.; Hao, L.Q.; Liu, M.; Wang, B. The Long Noncoding RNA PVT1 Functions as a Competing Endogenous RNA by Sponging MiR-186 in Gastric Cancer. Biomed. Pharmacother. 2017, 88, 302–308. [Google Scholar] [CrossRef]
- Abdel-Haleem, A.M.; Lewis, N.E.; Jamshidi, N.; Mineta, K.; Gao, X.; Gojobori, T. The Emerging Facets of Non-Cancerous Warburg Effect. Front. Endocrinol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Vaupel, P.; Schmidberger, H.; Mayer, A. The Warburg Effect: Essential Part of Metabolic Reprogramming and Central Contributor to Cancer Progression. Int. J. Radiat. Biol. 2019, 95, 912–919. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does It Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Ruiz, R.; Rigoulet, M.; Devin, A. The Warburg and Crabtree Effects: On the Origin of Cancer Cell Energy Metabolism and of Yeast Glucose Repression. Biochim. Biophys. Acta BBA Bioenerg. 2011, 1807, 568–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetzman, E.S.; Prochownik, E.V. The Role for Myc in Coordinating Glycolysis, Oxidative Phosphorylation, Glutaminolysis, and Fatty Acid Metabolism in Normal and Neoplastic Tissues. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K–AKT Network at the Interface of Oncogenic Signalling and Cancer Metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Gonçalves, V.; Lameirinhas, A.; Henrique, R.; Jerónimo, C. Metabolism and Epigenetic Interplay in Cancer: Regulation and Putative Therapeutic Targets. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkie, M.D.; Anaam, E.A.; Lau, A.S.; Rubbi, C.P.; Jones, T.M.; Boyd, M.T.; Vlatković, N. TP53 Mutations in Head and Neck Cancer Cells Determine the Warburg Phenotypic Switch Creating Metabolic Vulnerabilities and Therapeutic Opportunities for Stratified Therapies. Cancer Lett. 2020, 478, 107–121. [Google Scholar] [CrossRef]
- Varghese, E.; Samuel, S.M.; Líšková, A.; Samec, M.; Kubatka, P.; Büsselberg, D. Targeting Glucose Metabolism to Overcome Resistance to Anticancer Chemotherapy in Breast Cancer. Cancers 2020, 12, 2252. [Google Scholar] [CrossRef]
- Dabral, S.; Muecke, C.; Valasarajan, C.; Schmoranzer, M.; Wietelmann, A.; Semenza, G.L.; Meister, M.; Muley, T.; Seeger-Nukpezah, T.; Samakovlis, C.; et al. A RASSF1A-HIF1α Loop Drives Warburg Effect in Cancer and Pulmonary Hypertension. Nat. Commun. 2019, 10, 2130. [Google Scholar] [CrossRef] [Green Version]
- Sadlecki, P.; Bodnar, M.; Grabiec, M.; Marszalek, A.; Walentowicz, P.; Sokup, A.; Zegarska, J.; Walentowicz-Sadlecka, M. The Role of Hypoxia-Inducible Factor-1α, Glucose Transporter-1, (GLUT-1) and Carbon Anhydrase IX in Endometrial Cancer Patients. Available online: https://www.hindawi.com/journals/bmri/2014/616850/ (accessed on 13 October 2020).
- Liu, Y.; Li, Y.; Tian, R.; Liu, W.; Fei, Z.; Long, Q.; Wang, X.; Zhang, X. The Expression and Significance of HIF-1α and GLUT-3 in Glioma. Brain Res. 2009, 1304, 149–154. [Google Scholar] [CrossRef]
- Fujino, M.; Aishima, S.; Shindo, K.; Oda, Y.; Morimatsu, K.; Tsutsumi, K.; Otsuka, T.; Tanaka, M.; Oda, Y. Expression of Glucose Transporter-1 Is Correlated with Hypoxia-Inducible Factor 1 (Alpha) and Malignant Potential in Pancreatic Neuroendocrine Tumors. Oncol. Lett. 2016, 12, 3337–3344. [Google Scholar] [CrossRef] [Green Version]
- Demaria, M.; Poli, V. PKM2, STAT3 and HIF-1α. JAK STAT 2012, 1, 194–196. [Google Scholar] [CrossRef] [Green Version]
- Dong, G.; Mao, Q.; Xia, W.; Xu, Y.; Wang, J.; Xu, L.; Jiang, F. PKM2 and Cancer: The Function of PKM2 beyond Glycolysis (Review). Oncol. Lett. 2016, 11, 1980–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palsson-McDermott, E.M.; Curtis, A.M.; Goel, G.; Lauterbach, M.A.; Sheedy, F.J.; Gleeson, L.E.; van den Bosch, M.W.; Quinn, S.R.; Domingo-Fernandez, R.; Johnson, D.G.; et al. Pyruvate Kinase M2 Regulates Hif-1α Activity and IL-1β Induction, and Is a Critical Determinant of the Warburg Effect in LPS-Activated Macrophages. Cell Metab. 2015, 21, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, V.P.; Miyamoto, S. HK2/Hexokinase-II Integrates Glycolysis and Autophagy to Confer Cellular Protection. Autophagy 2015, 11, 963–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, K.; Kirito, K.; Kawashima, I.; Koshiishi, M.; Kumagai, T.; Mitsumori, T. Aberrant Activation of NF-ΚB and Hypoxia Enhances Glycolytic Enzyme Hexokinase II Expression in B-Cell Lymphoma Through Activation of HIF-1. Blood 2017, 130, 4025. [Google Scholar] [CrossRef]
- Tarrado-Castellarnau, M.; Diaz-Moralli, S.; Polat, I.H.; Sanz-Pamplona, R.; Alenda, C.; Moreno, V.; Castells, A.; Cascante, M. Glyceraldehyde-3-Phosphate Dehydrogenase Is Overexpressed in Colorectal Cancer Onset. Transl. Med. Commun. 2017, 2, 6. [Google Scholar] [CrossRef]
- Liu, K.; Tang, Z.; Huang, A.; Chen, P.; Liu, P.; Yang, J.; Lu, W.; Liao, J.; Sun, Y.; Wen, S.; et al. Glyceraldehyde-3-Phosphate Dehydrogenase Promotes Cancer Growth and Metastasis Through Upregulation of SNAIL Expression. Int. J. Oncol. 2017, 50, 252–262. [Google Scholar] [CrossRef]
- Jadhav, S.; Krynetskaia, N.; Krynetskiy, E. Cellular Response to Anticancer Drugs in Melanoma Cells: Inhibition of Glyceraldehyde 3-Phosphate Dehydrogenase Results in Cell Cycle Arrest and Increased Chemoresistance to Chemotherapeutic Agents. Cancer Res. 2007, 67, 3238. [Google Scholar]
- Higashimura, Y.; Nakajima, Y.; Yamaji, R.; Harada, N.; Shibasaki, F.; Nakano, Y.; Inui, H. Up-Regulation of Glyceraldehyde-3-Phosphate Dehydrogenase Gene Expression by HIF-1 Activity Depending on Sp1 in Hypoxic Breast Cancer Cells. Arch. Biochem. Biophys. 2011, 509, 1–8. [Google Scholar] [CrossRef]
- Li, L.; Liang, Y.; Kang, L.; Liu, Y.; Gao, S.; Chen, S.; Li, Y.; You, W.; Dong, Q.; Hong, T.; et al. Transcriptional Regulation of the Warburg Effect in Cancer by SIX1. Cancer Cell 2018, 33, 368–385. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Tang, B.; Sun, X. Development of Inhibitors Targeting Hypoxia-Inducible Factor 1 and 2 for Cancer Therapy. Yonsei Med. J. 2017, 58, 489–496. [Google Scholar] [CrossRef]
- Albadari, N.; Deng, S.; Li, W. The Transcriptional Factors HIF-1 and HIF-2 and Their Novel Inhibitors in Cancer Therapy. Expert Opin. Drug Discov. 2019, 14, 667–682. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Rapisarda, A.; Park, S.R.; Kinders, R.J.; Chen, A.; Melillo, G.; Turkbey, B.; Steinberg, S.M.; Choyke, P.; Doroshow, J.H.; et al. Pilot Trial of EZN-2968, an Antisense Oligonucleotide Inhibitor of Hypoxia-Inducible Factor-1 Alpha (HIF-1α), in Patients with Refractory Solid Tumors. Cancer Chemother. Pharmacol. 2014, 73, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Wigerup, C.; Påhlman, S.; Bexell, D. Therapeutic Targeting of Hypoxia and Hypoxia-Inducible Factors in Cancer. Pharmacol. Ther. 2016, 164, 152–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beppu, K.; Nakamura, K.; Linehan, W.M.; Rapisarda, A.; Thiele, C.J. Topotecan Blocks Hypoxia-Inducible Factor-1α and Vascular Endothelial Growth Factor Expression Induced by Insulin-like Growth Factor-I in Neuroblastoma Cells. Cancer Res. 2005, 65, 4775–4781. [Google Scholar] [CrossRef] [Green Version]
- Aquino-Gálvez, A.; González-Ávila, G.; Delgado-Tello, J.; Castillejos-López, M.; Mendoza-Milla, C.; Zúñiga, J.; Checa, M.; Maldonado-Martínez, H.A.; Trinidad-López, A.; Cisneros, J.; et al. Effects of 2-Methoxyestradiol on Apoptosis and HIF-1α and HIF-2α Expression in Lung Cancer Cells under Normoxia and Hypoxia. Oncol. Rep. 2016, 35, 577–583. [Google Scholar] [CrossRef]
- Narita, T.; Yin, S.; Gelin, C.F.; Moreno, C.S.; Yepes, M.; Nicolaou, K.C.; Van Meir, E.G. Identification of a Novel Small Molecule HIF-1α Translation Inhibitor. Clin. Cancer Res. 2009, 15, 6128–6136. [Google Scholar] [CrossRef] [Green Version]
- Womeldorff, M.; Gillespie, D.; Jensen, R.L. Hypoxia-Inducible Factor-1 and Associated Upstream and Downstream Proteins in the Pathophysiology and Management of Glioblastoma. Neurosurg. Focus 2014, 37, E8. [Google Scholar] [CrossRef]
- Papale, M.; Ferretti, E.; Battaglia, G.; Bellavia, D.; Mai, A.; Tafani, M. EZH2, HIF-1, and Their Inhibitors: An Overview on Pediatric Cancers. Front. Pediatr. 2018, 6. [Google Scholar] [CrossRef]
- Fallah, J.; Rini, B.I. HIF Inhibitors: Status of Current Clinical Development. Curr. Oncol. Rep. 2019, 21, 6. [Google Scholar] [CrossRef]
- Ibrahim, N.O.; Hahn, T.; Franke, C.; Stiehl, D.P.; Wirthner, R.; Wenger, R.H.; Katschinski, D.M. Induction of the Hypoxia-Inducible Factor System by Low Levels of Heat Shock Protein 90 Inhibitors. Cancer Res. 2005, 65, 11094–11100. [Google Scholar] [CrossRef] [Green Version]
- Hutt, D.M.; Roth, D.M.; Vignaud, H.; Cullin, C.; Bouchecareilh, M. The Histone Deacetylase Inhibitor, Vorinostat, Represses Hypoxia Inducible Factor 1 Alpha Expression Through Translational Inhibition. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, E.; Nordgren, I.K.; Male, A.L.; Lawrence, C.E.; Hoakwie, F.; Cuda, F.; Court, W.; Fox, K.R.; Townsend, P.A.; Packham, G.K.; et al. A Cyclic Peptide Inhibitor of HIF-1 Heterodimerization That Inhibits Hypoxia Signaling in Cancer Cells. J. Am. Chem. Soc. 2013, 135, 10418–10425. [Google Scholar] [CrossRef] [PubMed]
- Mangraviti, A.; Raghavan, T.; Volpin, F.; Skuli, N.; Gullotti, D.; Zhou, J.; Asnaghi, L.; Sankey, E.; Liu, A.; Wang, Y.; et al. HIF-1α- Targeting Acriflavine Provides Long Term Survival and Radiological Tumor Response in Brain Cancer Therapy. Sci. Rep. 2017, 7, 14978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Qian, D.Z.; Rey, S.; Wei, H.; Liu, J.O.; Semenza, G.L. Anthracycline Chemotherapy Inhibits HIF-1 Transcriptional Activity and Tumor-Induced Mobilization of Circulating Angiogenic Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2353–2358. [Google Scholar] [CrossRef] [Green Version]
- Viziteu, E.; Grandmougin, C.; Goldschmidt, H.; Seckinger, A.; Hose, D.; Klein, B.; Moreaux, J. Chetomin, Targeting HIF-1α/P300 Complex, Exhibits Antitumour Activity in Multiple Myeloma. Br. J. Cancer 2016, 114, 519–523. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Available online: https://www.hindawi.com/journals/tswj/2013/162750/ (accessed on 25 October 2020).
- Hernández-Rodríguez, P.; Baquero, L.P.; Larrota, H.R. Chapter 14—Flavonoids: Potential Therapeutic Agents by Their Antioxidant Capacity. In Bioactive Compounds; Campos, M.R.S., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 265–288. ISBN 978-0-12-814774-0. [Google Scholar]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant Properties of Flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-Inflammatory Effects of Flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial Activity of Flavonoids and Their Structure-Activity Relationship: An Update Review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [Green Version]
- Al Aboody, M.S.; Mickymaray, S. Anti-Fungal Efficacy and Mechanisms of Flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Ninfali, P.; Antonelli, A.; Magnani, M.; Scarpa, E.S. Antiviral Properties of Flavonoids and Delivery Strategies. Nutrients 2020, 12, 2534. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Gaforio, J.J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghaddam, G.; Ebrahimi, S.A.; Rahbar-Roshandel, N.; Foroumadi, A. Antiproliferative Activity of Flavonoids: Influence of the Sequential Methoxylation State of the Flavonoid Structure. Phytother. Res. 2012, 26, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Ungaro, P. Chapter 9—Epigenetic Mechanisms of Quercetin and Other Flavonoids in Cancer Therapy and Prevention. In Epigenetics of Cancer Prevention; Translational Epigenetics; Bishayee, A., Bhatia, D., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 8, pp. 187–202. [Google Scholar]
- Zimna, A.; Kurpisz, M. Hypoxia-Inducible Factor-1 in Physiological and Pathophysiological Angiogenesis: Applications and Therapies. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Unwith, S.; Zhao, H.; Hennah, L.; Ma, D. The Potential Role of HIF on Tumour Progression and Dissemination. Int. J. Cancer 2015, 136, 2491–2503. [Google Scholar] [CrossRef]
- Hubbi, M.E.; Semenza, G.L. Regulation of Cell Proliferation by Hypoxia-Inducible Factors. Am. J. Physiol. Cell Physiol. 2015, 309, C775–C782. [Google Scholar] [CrossRef] [Green Version]
- Kilic, M.; Kasperczyk, H.; Fulda, S.; Debatin, K.-M. Role of Hypoxia Inducible Factor-1 Alpha in Modulation of Apoptosis Resistance. Oncogene 2007, 26, 2027–2038. [Google Scholar] [CrossRef]
- Barreca, D.; Gattuso, G.; Bellocco, E.; Calderaro, A.; Trombetta, D.; Smeriglio, A.; Laganà, G.; Daglia, M.; Meneghini, S.; Nabavi, S.M. Flavanones: Citrus Phytochemical with Health-Promoting Properties. BioFactors 2017, 43, 495–506. [Google Scholar] [CrossRef]
- Jiang, N.; Doseff, A.I.; Grotewold, E. Flavones: From Biosynthesis to Health Benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Wannatung, T.; Lee, S.; Yang, W.K.; Chung, S.H.; Lim, J.-S.; Choe, W.; Kang, I.; Kim, S.-S.; Ha, J. Quercetin Enhances Hypoxia-Mediated Apoptosis via Direct Inhibition of AMPK Activity in HCT116 Colon Cancer. Apoptosis 2012, 17, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Lee, Y.J. Quercetin Suppresses Hypoxia-Induced Accumulation of Hypoxia-Inducible Factor-1α (HIF-1α) Through Inhibiting Protein Synthesis. J. Cell. Biochem. 2008, 105, 546–553. [Google Scholar] [CrossRef]
- Du, G.; Lin, H.; Wang, M.; Zhang, S.; Wu, X.; Lu, L.; Ji, L.; Yu, L. Quercetin Greatly Improved Therapeutic Index of Doxorubicin against 4T1 Breast Cancer by Its Opposing Effects on HIF-1α in Tumor and Normal Cells. Cancer Chemother. Pharmacol. 2010, 65, 277–287. [Google Scholar] [CrossRef]
- Granja, A.; Pinheiro, M.; Reis, S. Epigallocatechin Gallate Nanodelivery Systems for Cancer Therapy. Nutrients 2016, 8, 307. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, Y.; Liu, Z.; Wang, F.; Zhao, Q. Inhibitory Effects of Epigallocatechin-3-Gallate on Cell Proliferation and the Expression of HIF-1α and P-Gp in the Human Pancreatic Carcinoma Cell Line PANC-1. Oncol. Rep. 2012, 27, 1567–1572. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Tang, X.; Lu, Q.; Zhang, Z.; Rao, J.; Le, A.D. Green Tea Extract and (-)-Epigallocatechin-3-Gallate Inhibit Hypoxia- and Serum-Induced HIF-1α Protein Accumulation and VEGF Expression in Human Cervical Carcinoma and Hepatoma Cells. Mol. Cancer Ther. 2006, 5, 1227–1238. [Google Scholar] [CrossRef] [Green Version]
- Jagadeeshan, S.; Prasad, M.M.; Nair, S.A. Chapter 14—Role of Deguelin in Chemoresistance. In Role of Nutraceuticals in Cancer Chemosensitization; Cancer Sensitizing Agents for Chemotherapy; Bharti, A.C., Aggarwal, B.B., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 2, pp. 287–296. [Google Scholar]
- Oh, S.-H.; Woo, J.K.; Jin, Q.; Kang, H.-J.; Jeong, J.-W.; Kim, K.-W.; Hong, W.K.; Lee, H.-Y. Identification of Novel Antiangiogenic Anticancer Activities of Deguelin Targeting Hypoxia-Inducible Factor-1 Alpha. Int. J. Cancer 2008, 122, 5–14. [Google Scholar] [CrossRef]
- Oh, S.H.; Woo, J.K.; Yazici, Y.D.; Myers, J.N.; Kim, W.-Y.; Jin, Q.; Hong, S.S.; Park, H.-J.; Suh, Y.-G.; Kim, K.-W.; et al. Structural Basis for Depletion of Heat Shock Protein 90 Client Proteins by Deguelin. J. Natl. Cancer Inst. 2007, 99, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-Y.; Oh, S.H.; Woo, J.-K.; Hong, W.K.; Lee, H.-Y. Targeting Heat Shock Protein 90 Overrides the Resistance of Lung Cancer Cells by Blocking Radiation-Induced Stabilization of Hypoxia-Inducible Factor-1α. Cancer Res. 2009, 69, 1624–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-J.; Wu, C.-S.; Shieh, J.-J.; Wu, J.-H.; Chen, H.-Y.; Chung, T.-W.; Chen, Y.-K.; Lin, C.-C. Baicalein Triggers Mitochondria-Mediated Apoptosis and Enhances the Antileukemic Effect of Vincristine in Childhood Acute Lymphoblastic Leukemia CCRF-CEM Cells. Available online: https://www.hindawi.com/journals/ecam/2013/124747/ (accessed on 27 October 2020).
- Dou, J.; Wang, Z.; Ma, L.; Peng, B.; Mao, K.; Li, C.; Su, M.; Zhou, C.; Peng, G. Baicalein and Baicalin Inhibit Colon Cancer Using Two Distinct Fashions of Apoptosis and Senescence. Oncotarget 2018, 9, 20089–20102. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhang, Y.; Zhang, W.; Guo, Q. Baicalein Suppresses the Proliferation and Invasiveness of Colorectal Cancer Cells by Inhibiting Snail-Induced Epithelial-Mesenchymal Transition. Mol. Med. Rep. 2020, 21, 2544–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Peng, B.; Nayak, Y.; Wang, C.; Si, F.; Liu, X.; Dou, J.; Xu, H.; Peng, G. Baicalein and Baicalin Promote Melanoma Apoptosis and Senescence via Metabolic Inhibition. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Wang, F.-R.; Jiang, Y.-S. Effect of Treatment with Baicalein on the Intracerebral Tumor Growth and Survival of Orthotopic Glioma Models. J. Neurooncol. 2015, 124, 5–11. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.; Chen, A.Y.; Ye, X.; Luo, H.; Rankin, G.O.; Chen, Y.C. Inhibitory Effect of Baicalin and Baicalein on Ovarian Cancer Cells. Int. J. Mol. Sci. 2013, 14, 6012–6025. [Google Scholar] [CrossRef] [Green Version]
- Balam, F.H.; Ahmadi, Z.S.; Ghorbani, A. Inhibitory Effect of Chrysin on Estrogen Biosynthesis by Suppression of Enzyme Aromatase (CYP19): A Systematic Review. Heliyon 2020, 6. [Google Scholar] [CrossRef]
- Moghadam, E.R.; Ang, H.L.; Asnaf, S.E.; Zabolian, A.; Saleki, H.; Yavari, M.; Esmaeili, H.; Zarrabi, A.; Ashrafizadeh, M.; Kumar, A.P. Broad-Spectrum Preclinical Antitumor Activity of Chrysin: Current Trends and Future Perspectives. Biomolecules 2020, 10, 1374. [Google Scholar] [CrossRef]
- Fu, B.; Xue, J.; Li, Z.; Shi, X.; Jiang, B.-H.; Fang, J. Chrysin Inhibits Expression of Hypoxia-Inducible Factor-1α Through Reducing Hypoxia-Inducible Factor-1α Stability and Inhibiting Its Protein Synthesis. Mol. Cancer Ther. 2007, 6, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Fang, B.; Chen, X.; Wu, M.; Kong, H.; Chu, G.; Zhou, Z.; Zhang, C.; Chen, B. Luteolin Inhibits Angiogenesis of the M2-like TAMs via the Downregulation of Hypoxia Inducible Factor-1α and the STAT3 Signalling Pathway under Hypoxia. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent Progress Regarding Kaempferol for the Treatment of Various Diseases (Review). Exp. Ther. Med. 2019, 18, 2759–2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonis, I.; Lakka, A.; Tsakalof, A.; Simos, G. The Dietary Flavonoid Kaempferol Effectively Inhibits HIF-1 Activity and Hepatoma Cancer Cell Viability under Hypoxic Conditions. Biochem. Biophys. Res. Commun. 2010, 398, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, A.; Mylonis, I.; Simos, G.; Bonanou, S.; Tsakalof, A. Flavonoids Induce HIF-1α but Impair Its Nuclear Accumulation and Activity. Free Radic. Biol. Med. 2008, 44, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.; Bender-Sigel, J.; Schrenk, D.; Flügel, D.; Kietzmann, T. The Antioxidant Quercetin Inhibits Cellular Proliferation via HIF-1-Dependent Induction of P21WAF. Antioxid. Redox Signal. 2010, 13, 437–448. [Google Scholar] [CrossRef]
- Triantafyllou, A.; Liakos, P.; Tsakalof, A.; Chachami, G.; Paraskeva, E.; Molyvdas, P.-A.; Georgatsou, E.; Simos, G.; Bonanou, S. The Flavonoid Quercetin Induces Hypoxia-Inducible Factor-1alpha (HIF-1α) and Inhibits Cell Proliferation by Depleting Intracellular Iron. Free Radic. Res. 2007, 41, 342–356. [Google Scholar] [CrossRef]
- Park, S.-S.; Bae, I.; Lee, Y.J. Flavonoids-Induced Accumulation of Hypoxia-Inducible Factor (HIF)-1α/2α Is Mediated Through Chelation of Iron. J. Cell. Biochem. 2008, 103, 1989–1998. [Google Scholar] [CrossRef]
- Gao, J.-L.; Chen, Y.-G. Natural Compounds Regulate Glycolysis in Hypoxic Tumor Microenvironment. Available online: https://www.hindawi.com/journals/bmri/2015/354143/ (accessed on 28 October 2020).
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed Res. Int. 2019, 2019. [Google Scholar] [CrossRef]
- Melstrom, L.G.; Salabat, M.R.; Ding, X.-Z.; Strouch, M.J.; Grippo, P.J.; Mirzoeva, S.; Pelling, J.C.; Bentrem, D.J. Apigenin Down-Regulates the Hypoxia Response Genes: HIF-1α, GLUT-1, and VEGF in Human Pancreatic Cancer Cells. J. Surg. Res. 2011, 167, 173–181. [Google Scholar] [CrossRef]
- Chen, F.; Zhuang, M.; Zhong, C.; Peng, J.; Wang, X.; Li, J.; Chen, Z.; Huang, Y. Baicalein Reverses Hypoxia-Induced 5-FU Resistance in Gastric Cancer AGS Cells Through Suppression of Glycolysis and the PTEN/Akt/HIF-1α Signaling Pathway. Oncol. Rep. 2015, 33, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Nepal, M.; Choi, H.J.; Choi, B.-Y.; Kim, S.L.; Ryu, J.-H.; Kim, D.H.; Lee, Y.-H.; Soh, Y. Anti-Angiogenic and Anti-Tumor Activity of Bavachinin by Targeting Hypoxia-Inducible Factor-1α. Eur. J. Pharmacol. 2012, 691, 28–37. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, D.; Han, S.; Wang, N.; Mo, F.; Loo, T.Y.; Shen, J.; Huang, H.; Chen, J. Bioactivity-Guided Identification and Cell Signaling Technology to Delineate the Lactate Dehydrogenase A Inhibition Effects of Spatholobus Suberectus on Breast Cancer. PLoS ONE 2013, 8, e56631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Veena, C.K.; Morgan, J.B.; Mohammed, K.A.; Jekabsons, M.B.; Nagle, D.G.; Zhou, Y.-D. Methylalpinumisoflavone Inhibits Hypoxia-Inducible Factor-1 (HIF-1) Activation by Simultaneously Targeting Multiple Pathways. J. Biol. Chem. 2009, 284, 5859–5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacanlı, M.; Başaran, A.A.; Başaran, N. Chapter 4—The Major Flavonoid of Grapefruit: Naringin. In Polyphenols: Prevention and Treatment of Human Disease (Second Edition); Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 37–44. ISBN 978-0-12-813008-7. [Google Scholar]
- Tajaldini, M.; Samadi, F.; Khosravi, A.; Ghasemnejad, A.; Asadi, J. Protective and Anticancer Effects of Orange Peel Extract and Naringin in Doxorubicin Treated Esophageal Cancer Stem Cell Xenograft Tumor Mouse Model. Biomed. Pharmacother. 2020, 121, 109594. [Google Scholar] [CrossRef] [PubMed]
- Deenonpoe, R.; Prayong, P.; Thippamom, N.; Meephansan, J.; Na-Bangchang, K. Anti-Inflammatory Effect of Naringin and Sericin Combination on Human Peripheral Blood Mononuclear Cells (HPBMCs) from Patient with Psoriasis. BMC Complement. Altern. Med. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dwivedi, P.; Singh Rawat, A.K.; Dwivedi, A.K. 3—Nutrition Nutraceuticals: A Proactive Approach for Healthcare. In Nutraceuticals; Nanotechnology in the Agri-Food Industry; Grumezescu, A.M., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 79–116. ISBN 978-0-12-804305-9. [Google Scholar]
- Guo, B.; Zhang, Y.; Hui, Q.; Wang, H.; Tao, K. Naringin Suppresses the Metabolism of A375 Cells by Inhibiting the Phosphorylation of C-Src. Tumor Biol. 2016, 37, 3841–3850. [Google Scholar] [CrossRef]
- Wei, L.; Zhou, Y.; Qiao, C.; Ni, T.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Oroxylin A Inhibits Glycolysis-Dependent Proliferation of Human Breast Cancer via Promoting SIRT3-Mediated SOD2 Transcription and HIF1α Destabilization. Cell Death Dis. 2015, 6, e1714. [Google Scholar] [CrossRef] [Green Version]
- Suvarna, V.; Sarkar, M.; Chaubey, P.; Murahari, M.; Sangave, P.C. 13—Role of Natural Products in Glaucoma Management. In Handbook of Nutrition, Diet, and the Eye, 2nd ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 221–230. ISBN 978-0-12-815245-4. [Google Scholar]
- Wang, H.; Zhao, L.; Zhu, L.-T.; Wang, Y.; Pan, D.; Yao, J.; You, Q.-D.; Guo, Q.-L. Wogonin Reverses Hypoxia Resistance of Human Colon Cancer HCT116 Cells via Downregulation of HIF-1α and Glycolysis, by Inhibiting PI3K/Akt Signaling Pathway. Mol. Carcinog. 2014, 53, E107–E118. [Google Scholar] [CrossRef]
- Jung, K.-H.; Lee, J.H.; Thien Quach, C.H.; Paik, J.-Y.; Oh, H.; Park, J.W.; Lee, E.J.; Moon, S.-H.; Lee, K.-H. Resveratrol Suppresses Cancer Cell Glucose Uptake by Targeting Reactive Oxygen Species-Mediated Hypoxia-Inducible Factor-1α Activation. J. Nucl. Med. 2013, 54, 2161–2167. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.-H.; Ho, C.-T.; Chen, Z.-F.; Chen, L.-C.; Whang-Peng, J.; Lin, T.-N.; Ho, Y.-S. The Apple Polyphenol Phloretin Inhibits Breast Cancer Cell Migration and Proliferation via Inhibition of Signals by Type 2 Glucose Transporter. J. Food Drug Anal. 2018, 26, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, L.; Wu, Y.; Dai, Q.; Zhou, Y.; Li, Z.; Yang, L.; Guo, Q.; Lu, N. Selective Anti-Tumor Activity of Wogonin Targeting the Warburg Effect through Stablizing P53. Pharmacol. Res. 2018, 135, 49–59. [Google Scholar] [CrossRef]
- Ojelabi, O.A.; Lloyd, K.P.; De Zutter, J.K.; Carruthers, A. Red Wine and Green Tea Flavonoids Are Cis-Allosteric Activators and Competitive Inhibitors of Glucose Transporter 1 (GLUT1)-Mediated Sugar Uptake. J. Biol. Chem. 2018, 293, 19823–19834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, J.; Guo, F.; Chen, Y.; Li, Y.; Yu, B.; Li, Y. Nanoliposomal Formulation Encapsulating Celecoxib and Genistein Inhibiting COX-2 Pathway and Glut-1 Receptors to Prevent Prostate Cancer Cell Proliferation. Cancer Lett. 2019, 448, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-Y.; Yuan, Y.; Shi, X.; Che, Y.-Y. Improved Drug Delivery and Anti-Tumor Efficacy of Combinatorial Liposomal Formulation of Genistein and Plumbagin by Targeting Glut1 and Akt3 Proteins in Mice Bearing Prostate Tumor. Colloids Surf. B Biointerfaces 2020, 190, 110966. [Google Scholar] [CrossRef] [PubMed]
- Palombo, R.; Caporali, S.; Falconi, M.; Iacovelli, F.; Morozzo Della Rocca, B.; Lo Surdo, A.; Campione, E.; Candi, E.; Melino, G.; Bernardini, S.; et al. Luteolin-7-O-β-d-Glucoside Inhibits Cellular Energy Production Interacting with HEK2 in Keratinocytes. Int. J. Mol. Sci. 2019, 20, 2689. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Pan, L.; Gao, C.; Xu, H.; Li, Y.; Zhang, L.; Ma, L.; Meng, L.; Sun, X.; Qin, H. Quercetin Inhibits the Proliferation of Glycolysis-Addicted HCC Cells by Reducing Hexokinase 2 and Akt-MTOR Pathway. Molecules 2019, 24, 1993. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, X.; Wang, Y.; Sun, Q.; Chen, M.; Liu, S.; Zou, X. Licochalcone A Suppresses Hexokinase 2-Mediated Tumor Glycolysis in Gastric Cancer via Downregulation of the Akt Signaling Pathway. Oncol. Rep. 2018, 39, 1181–1190. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Wei, L.; Liu, Y.; Ding, Y.; Liu, X.; Zhang, X.; Wang, X.; Yao, Y.; Lu, J.; Wang, Q.; et al. Gen-27, a Newly Synthesized Flavonoid, Inhibits Glycolysis and Induces Cell Apoptosis via Suppression of Hexokinase II in Human Breast Cancer Cells. Biochem. Pharmacol. 2017, 125, 12–25. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, L.; Zhou, Y.; Lu, N.; Tang, X.; Li, Z.; Wang, X. Flavonoid GL-V9 Induces Apoptosis and Inhibits Glycolysis of Breast Cancer via Disrupting GSK-3β-Modulated Mitochondrial Binding of HKII. Free Radic. Biol. Med. 2020, 146, 119–129. [Google Scholar] [CrossRef]
- Aslan, E.; Guler, C.; Adem, S. In Vitro Effects of Some Flavonoids and Phenolic Acids on Human Pyruvate Kinase Isoenzyme M2. J. Enzym. Inhib. Med. Chem. 2016, 31, 314–317. [Google Scholar] [CrossRef] [Green Version]
- Hasan, D.; Gamen, E.; Abu Tarboush, N.; Ismail, Y.; Pak, O.; Azab, B. PKM2 and HIF-1α Regulation in Prostate Cancer Cell Lines. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Wei, L.; Dai, Y.; Zhou, Y.; He, Z.; Yao, J.; Zhao, L.; Guo, Q.; Yang, L. Oroxylin A Activates PKM1/HNF4 Alpha to Induce Hepatoma Differentiation and Block Cancer Progression. Cell Death Dis. 2017, 8, e2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.; Shi, J.; Yang, P.; Jia, B.; Wu, H.; Zhang, X.; Li, Z. Apigenin Restrains Colon Cancer Cell Proliferation via Targeted Blocking of Pyruvate Kinase M2-Dependent Glycolysis. J. Agric. Food Chem. 2017, 65, 8136–8144. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate Dehydrogenase A: A Key Player in Carcinogenesis and Potential Target in Cancer Therapy. Cancer Med. 2018, 7, 6124–6136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-J.; Zhao, J.-K.; Ren, S.; Sun, W.-W.; Zhang, W.-J.; Zhang, J.-N. Wogonin Affects Proliferation and the Energy Metabolism of SGC-7901 and A549 Cells. Exp. Ther. Med. 2019, 17, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.-F.; Liu, X.; Li, W.-J.; Shi, Z.-W.; Yan, Y.-X.; Wang, L.-F.; Chen, M.; Xie, M.-Y. (-)-Epigallocatechin-3-Gallate Alleviates Doxorubicin-Induced Cardiotoxicity in Sarcoma 180 Tumor-Bearing Mice. Life Sci. 2017, 180, 151–159. [Google Scholar] [CrossRef]
- Gurunathan, S.; Jeyaraj, M.; Kang, M.-H.; Kim, J.-H. Tangeretin-Assisted Platinum Nanoparticles Enhance the Apoptotic Properties of Doxorubicin: Combination Therapy for Osteosarcoma Treatment. Nanomaterials 2019, 9, 1089. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wu, L.; Feng, J.; Li, J.; Liu, T.; Zhang, R.; Xu, S.; Cheng, K.; Zhou, Y.; Zhou, S.; et al. In Vitro and in Vivo Study of Epigallocatechin-3-Gallate-Induced Apoptosis in Aerobic Glycolytic Hepatocellular Carcinoma Cells Involving Inhibition of Phosphofructokinase Activity. Sci. Rep. 2016, 6, 28479. [Google Scholar] [CrossRef]
- Umar, S.M.; Kashyap, A.; Kahol, S.; Mathur, S.R.; Gogia, A.; Deo, S.V.S.; Prasad, C.P. Prognostic and Therapeutic Relevance of Phosphofructokinase Platelet-Type (PFKP) in Breast Cancer. Exp. Cell Res. 2020, 396, 112282. [Google Scholar] [CrossRef]
- Wei, R.; Hackman, R.M.; Wang, Y.; Mackenzie, G.G. Targeting Glycolysis with Epigallocatechin-3-Gallate Enhances the Efficacy of Chemotherapeutics in Pancreatic Cancer Cells and Xenografts. Cancers 2019, 11, 1496. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, R.; Mohammad, T.; Roy, S.; Anwar, S.; Gupta, P.; Haque, A.; Khan, P.; Kazim, S.N.; Islam, A.; Ahmad, F.; et al. Investigation of Inhibitory Potential of Quercetin to the Pyruvate Dehydrogenase Kinase 3: Towards Implications in Anticancer Therapy. Int. J. Biol. Macromol. 2019, 136, 1076–1085. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agents. | Effect on HIF-1 | References |
|---|---|---|
| EZN-2968 | Regulation of HIF-1α mRNA expression | [85] |
| Topotecan | Regulation of HIF-1α mRNA expression | [86,87] |
| 2-Methoxyestradiol | Modulation of HIF-1α and HIF-2α protein synthesis | [88] |
| KC7F2 | Modulation of HIF-1α protein synthesis | [89,90] |
| Geldanamycin, tanespimycin, and alvespimycin | Disruption of HIF-1α protein stabilization and accumulation | [91,92,93] |
| Vorinostat (HDACi) | Disruption of HIF-1α protein stabilization and accumulation | [94] |
| Cyclo-CLLFVY | Inhibition of HIF-1 α heterodimerization | [19,95] |
| Acriflavine | Inhibition of HIF-1α heterodimerization | [96] |
| Doxorubicin, daunorubicin | Inhibition of binding of HIF-1 to its target gene sequences | [97] |
| Chetomin | Inhibition of the HIF-1 transcriptional activity by targeting HIF-1α/p300 | [98] |
| Flavonoid | Study Details | Mechanism of Action | Effect on HIF-1 | Reference |
|---|---|---|---|---|
| Quercetin | Colon cancer cells (HCT116), BALB nu/nu mice | Inhibition of AMPK. Probability of cross-connection between suppression of AMPK and decrease in HIF-1 activity | Decrease in HIF-1 activity | [119] |
| Prostate cancer cells (LNCaP), breast cancer cells (SkBr3), and colon cancer cells (CX-1) | Modulation of the balance between HIF-1α translation and degradation | Inhibition of HIF-1α protein synthesis and accumulation | [120] | |
| Murine mammary carcinoma (4T1) | Promotion of HIF-1α degradation in cells. | Suppression of intra-tumoral HIF-1α | [121] | |
| EGCG | Pancreatic cancer cells (PANC-1) | Promotion of HIF-1α protein degradation and/or interaction with HIF-1α protein translational pathway | Decrease in HIF-1α protein expression | [123] |
| Human liver cancer cells (HepG2) and cervical cancer cells (HeLa) | Inhibition of HIF-1α expression by interfering with PI3K/Akt/mTOR pathway and translational apparatus of cancer cells | Inhibition of HIF-1α protein accumulation | [124] | |
| Deguelin | Lung cancer cells (H1299), human squamous cell carcinoma (UMSCC38), prostate cancer cells (PC3), gastric cancer cells (MKN-45), breast cancer cells (MCF-7), and vascular endothelial cells | Suppression of HIF-1α expression by the inhibition of protein synthesis, and its degradation through the ubiquitin- and proteasomal-dependent manner | Reduction of HIF-1α expression | [126] |
| Lung cancer (H1299) xenografts | Interaction with the ATP-binding pocket of Hsp90 and disruption of its function resulting in HIF-1α degradation via ubiquitin-mediated manner | Degradation of HIF-1α | [127] | |
| Lung cancer cells (H1299 and H226B); athymic nude mice bearing H1299 and H226B cells | Suppression of interaction between Hsp90 and HIF-1α | Suppression of HIF-1α expression | [128] | |
| Baicalein | Mice with human glioblastoma cells (U87) | Suppression of HIF-1α/VEGF pathway | Decrease in HIF-1α protein expression | [133] |
| Ovarian cancer cells (OVCAR-3a and CP-70) | Inhibition of HIF-1α expression by baicalein at concentration 20-μM and 40-μM. | Suppression of HIF-1α expression | [134] | |
| Chrysin | Prostate cancer cells (DU145) | Enhancing HIF-1α degradation via promotion of the prolyl hydroxylation of HIF-1α ODD, resulting in proteasomal-mediated degradation of HIF-1α. Regulation of HIF-1α expression via PI3K/Akt pathway | Inhibition of HIF-1α expression and protein stability | [137] |
| Luteolin | The mouse macrophage cell line RAW264.7 | Regulation of HIF-1α-VEGF/MMP9 signaling pathway | Suppression of HIF-1 activation | [138] |
| Kaempferol | hepatocellular cancer cells (Huh7) | HIF-1α mislocalization into the cytoplasm due to p44/42 MAPK inactivation, resulting in the suppression of HIF-1 activity | Decrease in HIF-1 activity | [140] |
| Quercetin, baicalein, luteolin, and fisetin | cervical cancer cells (HeLa) | Flavonoids affect HIF-1 transcriptional activity via impairing the MAPK pathway resulting in inhibition of phosphorylation and nuclear accumulation of HIF-1α | Inhibition of HIF-1α protein accumulation and HIF-1 transcriptional activity | [141] |
| Quercetin | Human hepatoma cells (HepG2) | Quercetin prolongs HIF-1α protein half time. Knock-down of the HIF-1α disrupts quercetin-mediated inhibition of cell proliferation | Induction of the HIF-1α expression and HIF-1 activity | [142] |
| cervical cancer cells (HeLa) | Quercetin induces HIF-1α and inhibits cell proliferation via iron chelation | Induction of the HIF-1α expression | [143] | |
| Human prostate adenocarcinoma cells (LNCaP, DU-145 and PC-3) | Quercetin containing iron-chelating moieties chelates cellular irons that are cofactors of HIF-1α/2α PHD leading to HIF-1a accumulation | Induction of HIF-1α/2α accumulation | [144] |
| Flavonoids | Study Details | Mechanism of Action | Effect on HIF-1/Glycolysis Components | Reference |
|---|---|---|---|---|
| Apigenin | Human pancreatic cancer cells (S2-013 and CD18) | Downregulation of HIF-1α and GLUT-1 mRNA expression. Repression of any hypoxia-mediated induction of GLUT-1 expression. Significant reduction of the HIF-1 protein level | Inhibition of HIF-1α and GLUT1 | [147] |
| Baicalein | Human gastric cancer cells (AGS) | Inhibition of glycolysis through the regulation of PTEN/Akt/HIF-1α signaling pathway | Suppression of HIF-1α expression; suppression of HKII, PDK1, LDHA | [148] |
| Bavachinin | HeLa derivatives (KB), Human osteosarcoma cells (HOS), KB xenografts | Promotion of VHL-HIF-1α interaction as a consequence of the elevated PHD2 activity. A decrease in glucose metabolism and energy level modulated by hypoxia due to bavachinin intervention | Inhibition of HIF-1 activity; decreased transcription of HKII and GLUT1 | [149] |
| EGC | Breast cancer cells (MCF-7 and MDA-MB-231) and MCF-7, MDA-MB-231 xenografts | Promotion of HIF-1α proteasomal-mediated degradation via Hsp90. EGC modulates the interaction of Hsp90/HIF-1α. Acceleration in HIF-1α proteasomal degradation correlates with regulation of LDHA | Inhibition of LDHA activity; induction of HIF-1α proteasomal degradation; decrease in HIF-1α/LDHA expression in vivo | [150] |
| ALP, 4’-OM | Breast cancer cells (T47D) | Inhibition of hypoxia-induced HIF-1 activation by ALP and 4’-OM. 4’-OM inhibits HIF-1 activation via suppression of mitochondrial electron transport chain and inhibition of protein synthesis | Inhibition of HIF-1 activity; inhibition of hypoxic induction of GLUT1 | [151] |
| Naringin | Human melanoma cells (A375) | Significant anticancer impact of naringin on HIF-1α, PKM2, LDHA expression mediated by suppression of phosphorylation of Tyr418 of c-Src | Inhibition of PKM2, LDHA, and HIF-1α | [156] |
| Oroxylin A | Breast cancer cells (MDA-MB-231) | Downregulation of HIF-1α via increasing PHD activity mediated by SIRT3. Glycolysis was inhibited by suppression of HIF-1 activity | HIF-1α destabilization; suppression of HKII expression | [157] |
| Wogonin | Colon cancer cells (HTC116) and Balb/C mice | Inhibition of HIF-1α and glycolysis-related proteins was mediated by suppression of PI3K/Akt signaling pathway leading to downregulation of PI3K/Akt-dependent transcriptional activity | Suppression of HIF-1α expression; decreases in HKII, PDK1, and LDHA expression | [159] |
| Enzyme | Flavonoid | Study Details | Mechanism of Action | Effects | Reference |
|---|---|---|---|---|---|
| Glucose transporters | |||||
| GLUT2 | Phloretin | Breast cancer cells (MDA-MB-231) | Inhibition of GLUT2 → accumulation of MDA-MB231 cells in the G0/G1 phase | suppression of migration and proliferation | [161] |
| GLUT1 | Wogonin | Colon cancer (HCT-116), ovarian cancer (A2780), and liver cancer (HepG2) cells, A2780 xenografts | Suppression of glucose metabolism followed by upregulated p53 mRNA and protein level (wt-p53 cancer cells) and regulation of p53 downstream glycolytic factors | Upregulated p53 and p53-inducible glycolysis in cancer cells and decreased GLUT1 in cells expressing wild type, but not mutated p53. Inhibition of glycolysis was accompanied by the downregulation of GLUT1 in xenografts | [162] |
| Red wine and green tea flavonoids | The evaluation of structure–function relationships in flavonoid–GLUT1 interactions | Stimulation of GLUT1-mediated sugar uptake at low concentrations → transport inhibition as the concentration raises (suggesting that at least two flavonoid-binding sites modulate GLUT1 function) | Act as: cis-allosteric activators of sugar uptake at low concentrations; and competitive inhibitors of GLUT1-mediated sugar uptake at higher concentrations | [163] | |
| Nanoliposomal encapsulation of celecoxib and genistein | Prostate cancer cells (PC-3, LNCaP) | Key processes behind the inhibition of prostate cancer cells: enhanced reactive oxygen species, decreased cellular GSH concentration, inhibited COX-2 synthesis and Glut-1 receptors | Suppressed GLUT1 receptors → prevention of prostate cancer cell proliferation | [164] | |
| Combinatorial liposomal formulation of plumbagin and genistein | Genistein (Glut-1 transporter protein inhibitor) induces high reactive oxygen species generation associated with AMPK signaling pathway. Low uptake of glucose → decreased metabolism of prostate cancer cells and simultaneous generation of reactive oxygen species and low GSH concentration → cell death | Decreased population of GLUT1 transporters | [165] | ||
| Enzymes of glucose metabolism | |||||
| HKII | LUT-7G | Keratinocytes | LUT-7G suggested to represent a strong HKII inhibitor via the binding in the active sites | HKII suppression → repression of the glycolytic pathway | [166] |
| Quercetin | Hepatocellular carcinoma cells (SMMC-7721 and Bel-7402) and murine xenograft model | Quercetin suppresses glycolysis through Akt-mTOR pathway-mediated HKII regulation | Inhibition of glycolysis and proliferation of glycolysis-addicted HCC cells (by reduced HKII) and decrease of HKII expression in vivo | [167] | |
| Licochalcone A | Gastric cancer cells (MKN45 and SGC7901) | Licochalcone A inhibits glycolysis mainly through the blockade of Akt signaling pathway | Suppression of HKII-mediated tumor glycolysis | [168] | |
| Gen-27 | Breast cancer cells (1H-I, MDA-MB-231, MCF-7 and MDA-MB-468) | The potential of Gen-27 to inhibit glycolysis and displaced HKII from mitochondrial membrane to the cytosol → blockage of its preferential access to ATP for glucose phosphorylation or preventing mechanism of cancer growth and immortality | Inhibition of glycolysis and induction of apoptosis (through HKII suppression accompanied by weakened interactions of HKII and VDAC) | [169] | |
| GL-V9 | Breast cancer cells (MDA-MB-231, MCF-7) | GL-V9 disrupts GSK-3β-modulated mitochondrial binding of HKII | Downregulation of HKII and disruption of mitochondrial binding of HKII resulting in apoptosis | [170] | |
| PKM1, PKM2 | Oroxylin A | Liver cancer model | Oroxylin A enhanced the protein expression of HNF-4α and its binding to the promoter region of HNF-1α and promoted direct interaction between PKM1 and HNF-4α in the nucleus | Increased PKM1/PKM2 ratio → HNF-4α activation → induction of hepatoma differentiation and suppression of cancer progression | [173] |
| Apigenin | Colon cancer cells (HCT116) | The potential of apigenin to ensure a low PKM2/PKM1 ratio through blockage of the β-catenin/c-Myc/PTBP1 signal pathway | Apigenin → allosteric PKM2 inhibitor (can ensure a low PKM2/PKM1 ratio and restrain the proliferation of colon cancer cells through a blockade of PKM2-dependent glycolysis) | [174] | |
| LDHA | Wogonin | Human gastric cancer cells (SGC-7901) and human lung adenocarcinoma cells (A549) | Effects of wogonin on energy metabolism: affecting ATP generation and the activities of energy associated with metabolism | Reduced LDHA activity | [176] |
| EGCG | Evaluation of effects of EGCG on doxorubicin-induced cardiotoxicity in Sarcoma 180 tumor bearing mice | EGCG-exerted heart benefits related to reduced LDH release | Attenuation of LDHA release. | [177] | |
| Tangeretin-assisted platinum nanoparticles | Osteosarcoma cells (U2OS) | Tangeretin-assisted platinum nanoparticles promote LDHA leakage | Increase of LDHA leakage and cell death | [178] | |
| PFK | Quercetin | Breast cancer cells (MDA-MB-231) | The ability of quercetin to impair PFKP-LDHA signaling → inhibiting migration of cancer cells mediated by aerobic glycolysis | Impairment of the PFKP-LDHA signaling axis → inhibition of cell migration induced by aerobic glycolysis | [180] |
| EGCG | Hepatocellular carcinoma cells (HCC-LM3 and HepG2) | EGCG inhibits glycolysis (especially PFK activity) in aerobic glycolytic HCC cell lines | Inhibition of PFK expression and activity | [179] | |
| Pancreatic cancer cells (Panc-1 and MIA PaCa-2) | EGCG inhibits glycolysis through repressing rate-limiting enzymes (PFK and PKM2) | Suppression of PFKP and PKM2 levels | [181] | ||
| PDK | Quercetin | Hepatocellular carcinoma cells (HepG2) and liver cancer (A549) cells | Quercetin binds with PDK3 and significantly inhibits its kinase activity | Interaction with residues of the active site cavity of PDK3 (conformational fitting). PDK3 inhibitory potential in cancer cells | [182] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samec, M.; Liskova, A.; Koklesova, L.; Mersakova, S.; Strnadel, J.; Kajo, K.; Pec, M.; Zhai, K.; Smejkal, K.; Mirzaei, S.; et al. Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers 2021, 13, 130. https://doi.org/10.3390/cancers13010130
Samec M, Liskova A, Koklesova L, Mersakova S, Strnadel J, Kajo K, Pec M, Zhai K, Smejkal K, Mirzaei S, et al. Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers. 2021; 13(1):130. https://doi.org/10.3390/cancers13010130
Chicago/Turabian StyleSamec, Marek, Alena Liskova, Lenka Koklesova, Sandra Mersakova, Jan Strnadel, Karol Kajo, Martin Pec, Kevin Zhai, Karel Smejkal, Sepideh Mirzaei, and et al. 2021. "Flavonoids Targeting HIF-1: Implications on Cancer Metabolism" Cancers 13, no. 1: 130. https://doi.org/10.3390/cancers13010130
APA StyleSamec, M., Liskova, A., Koklesova, L., Mersakova, S., Strnadel, J., Kajo, K., Pec, M., Zhai, K., Smejkal, K., Mirzaei, S., Hushmandi, K., Ashrafizadeh, M., Saso, L., Brockmueller, A., Shakibaei, M., Büsselberg, D., & Kubatka, P. (2021). Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers, 13(1), 130. https://doi.org/10.3390/cancers13010130