
A Rare Variant in ERF (rs144812092) Predisposes to Prostate and Bladder Cancers in an Extended Pedigree

 , , , , and
, , , , and 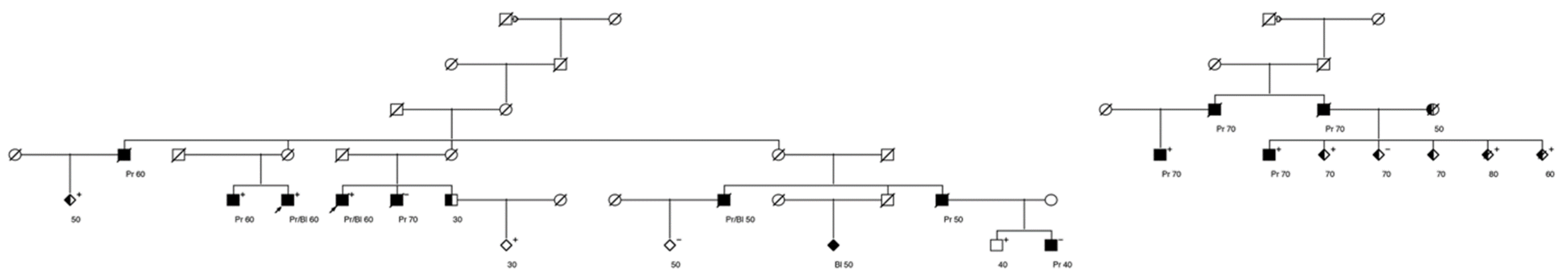{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Utah Population Data Base
2.2. Bladder Cancer Cases
2.3. High-Risk Bladder Cancer Pedigrees
2.4. Whole Exome Sequencing
2.5. Case-Control Association Analysis
2.6. Protein Prediction Modeling
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cannon-Albright, L.A.; Thomas, A.; Goldgar, D.E.; Gholami, K.; Rowe, K.; Jacobsen, M.; McWhorter, W.P.; Skolnick, M.H. Fa-miliality of cancer in Utah. Cancer Res. 1994, 54, 2378–2385. [Google Scholar] [PubMed]
- Albright, F.; Teerlink, C.; Werner, T.L.; Cannon-Albright, L.A. Significant evidence for a heritable contribution to cancer predisposition: A review of cancer familiality by site. BMC Cancer 2012, 12, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermejo, J.L.; Sundquist, J.; Hemminki, K. Sex-specific familial risks of urinary bladder cancer and associated neoplasms in Sweden. Int. J. Cancer 2009, 124, 2166–2171. [Google Scholar] [CrossRef]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nat. Cell Biol. 2009, 461, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Wijsman, E.M. The role of large pedigrees in an era of high-throughput sequencing. Qual. Life Res. 2012, 131, 1555–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, J.; Wang, J.; Leal, S.M. Genetic linkage analysis in the age of whole-genome sequencing. Nat. Rev. Genet. 2015, 16, 275–284. [Google Scholar] [CrossRef]
- Miki, Y.; Swensen, J.; Shattuck-Eidens, D.; Futreal, P.A.; Harshman, K.; Tavtigian, S.; Liu, Q.; Cochran, C.; Bennett, L.M.; Ding, W.; et al. A strong candidate for the breast and ovarian cancer susceptibility gene BRCA1. Science 1994, 266, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Tavtigian, S.V.; Simard, J.; Rommens, J.M.; Couch, F.J.; Shattuck-Eidens, D.; Neuhausen, S.L.; Merajver, S.D.; Thorlacius, S.; Offit, K.; Stoppalyonnet, D.; et al. The complete BRCA2 gene and mutations in chromosome 13q-linked kindreds. Nat. Genet. 1996, 12, 333–337. [Google Scholar] [CrossRef]
- Kamb, A.; Shattuck-Eidens, D.; Eeles, R.; Liu, Q.; Gruis, N.A.; Ding, W.; Hussey, C.; Tran, T.; Miki, Y.; Weaver-Feldhaus, J.; et al. Analysis of the p16 gene (CDKN2) as a candidate for the chromosome 9p mela-noma susceptibility locus. Nat. Genet. 1994, 8, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Teerlink, C.C.; Huff, C.; Stevens, J. A non-synonymous variant in GOLM1 in cutaneous malignant melanoma. JNCI J. Natl. Cancer Inst. 2018, 110, 1380–1385. [Google Scholar]
- Thompson, B.A.; Snow, A.K.; Koptiuch, C.; Kohlmann, W.K.; Mooney, R.; Johnson, S.; Huff, C.D.; Yu, Y.; Teerlink, C.C.; Feng, B.-J.; et al. A novel ribosomal protein S20 variant in a family with unexplained colo-rectal cancer and polyposis. Clin. Genet. 2020, 97. [Google Scholar] [CrossRef] [PubMed]
- Albright, L.A.C. Utah Family-Based Analysis: Past, Present and Future. Hum. Hered. 2007, 65, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Sudlow, C.; Gallacher, J.; Allen, N.; Beral, V.; Burton, P.; Danesh, J.; Downey, P.; Elliott, P.; Green, J.; Landray, M.; et al. UK Biobank: An Open Access Resource for Identifying the Causes of a Wide Range of Complex Diseases of Middle and Old Age. PLoS Med. 2015, 12, e1001779. [Google Scholar] [CrossRef] [Green Version]
- Abraham, G.; Qiu, Y.; Inouye, M. FlashPCA2: Principal component analysis of Biobank-scale genotype datasets. Bioinformatics 2017, 33, 2776–2778. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Deelen, P.; Bonder, M.J.; Van Der Velde, K.J.; Westra, H.-J.; Winder, E.; Hendriksen, D.; Franke, L.; Swertz, M.A. Genotype harmonizer: Automatic strand alignment and format conversion for genotype data integration. BMC Res. Notes 2014, 7, 901. [Google Scholar] [CrossRef] [Green Version]
- Loh, P.-R.; Palamara, P.F.; Price, A.L. Fast and accurate long-range phasing in a UK Biobank cohort. Nat. Genet. 2016, 48, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Forer, L.; Schönherr, S.; Sidore, C.; Locke, A.E.; Kwong, A.; Vrieze, S.I.; Chew, E.Y.; Levy, S.; McGue, M.; et al. Next-generation genotype imputation service and methods. Nat. Genet. 2016, 48, 1284–1287. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, R.; Facelli, J.C. Understanding protein structural chanes for oncogenic missense variants. Heliyon 2021, 7, e06013. [Google Scholar] [CrossRef]
- Hernandez, R.; Facelli, J.C. Structure analysis of the proteins associated with polyA repeat expansion disorders. J. Biomol. Struct. Dyn. 2021, 18, 1–11. [Google Scholar] [CrossRef]
- Teerlink, C.C.; Jurynec, M.J.; Hernande, R.; Stevens, J.; Hughes, D.C.; Brunker, C.P.; Rowe, K.; Grunwald, D.J.; Facelli, J.C.; Can-non-Albright, L.A. A role for the MEGF6 gene in predisposition to osteoporosis. Ann. Hum. Genet. 2020, 85, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, T.; Liu, B.; Hernandez, R.; Facelli, J.C.; Grossman, D. A novel CDKN 2A variant (p16 L117P ) in a patient with familial and multiple primary melanomas. Pigment. Cell Melanoma Res. 2019, 32, 734–738. [Google Scholar] [CrossRef]
- UNIProtKB – P50548. Available online: https://www.uniprot.org/uniprot/P50548#function (accessed on 13 May 2021).
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Post, R.S.; Kiemeney, L.A.; Ligtenberg, M.J.L.; Witjes, J.A.; Hulsbergen-van de Kaa, C.A.; Bodmer, D.; Schaap, L.; Kets, C.M.; van Krieken, J.H.J.M.; Hoogerbrugge, N. Risk of urothelial bladder cancer in Lynch syndrome is increased, in particular among MSH2 mutation carriers. J. Med. Genet. 2010, 47, 464–470. [Google Scholar] [CrossRef] [PubMed]
- ERF ETS2 Repressor Factor. Available online: https://www.ncbi.nlm.nih.gov/gene/2077 (accessed on 13 May 2021).
- Huber, C.D.; Kim, B.Y.; Lohmueller, K.E. Population genetic models of GERP scores suggest pervasive turnover of constrained sites across mammalian evolution. PLoS Genet. 2020, 16, e1008827. [Google Scholar] [CrossRef]
- Huang, F.W.; Mosquera, J.M.; Garofalo, A.; Oh, C.; Baco, M.; Amin-Mansour, A.; Rabasha, B.; Bahl, S.; Mullane, S.A.; Robinson, B.D.; et al. Exome Sequencing of African-American Prostate Cancer Reveals Loss-of-Function ERF Mutations. Cancer Discov. 2017, 7, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.; Van Allen, E.M.; Wu, Y.-M.; Schultz, N.; Lonigro, R.J.; Mosquera, J.-M.; Montgomery, B.; Taplin, M.-E.; Pritchard, C.C.; Attard, G.; et al. Integrative Clinical Genomics of Advanced Prostate Cancer. Cell 2015, 161, 1215–1228. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Coleman, I.; Morrissey, C.; Zhang, X.; True, L.D.; Gulati, R.; Etzioni, R.; Bolouri, H.; Montgomery, B.; White, T.; et al. Substantial interindividual and limited intraindividual genomic diversity among tumors from men with metastatic prostate cancer. Nat. Med. 2016, 22, 369–378. [Google Scholar] [CrossRef]
- Bose, R.; Karthaus, W.R.; Armenia, J.; Abida, W.; Iaquinta, P.J.; Zhang, Z.; Wongvipat, J.; Wasmuth, E.V.; Shah, N.; Sullivan, P.S.; et al. ERF mutations reveal a balance of ETS factors controlling prostate oncogenesis. Nat. Cell Biol. 2017, 546, 671–675. [Google Scholar] [CrossRef]
- Cannon-Albright, L.A.; Farnham, J.M.; Thomas, A.; Camp, N.J. Identification and study of Utah pseudo-isolate popula-tions-prospects for gene identification. Am. J. Med. Genet. A 2005, 137, 269–275. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cannon-Albright, L.A.; Teerlink, C.C.; Stevens, J.; Huang, F.W.; Sipeky, C.; Schleutker, J.; Hernandez, R.; Facelli, J.; Agarwal, N.; Trump, D.L. A Rare Variant in ERF (rs144812092) Predisposes to Prostate and Bladder Cancers in an Extended Pedigree. Cancers 2021, 13, 2399. https://doi.org/10.3390/cancers13102399
Cannon-Albright LA, Teerlink CC, Stevens J, Huang FW, Sipeky C, Schleutker J, Hernandez R, Facelli J, Agarwal N, Trump DL. A Rare Variant in ERF (rs144812092) Predisposes to Prostate and Bladder Cancers in an Extended Pedigree. Cancers. 2021; 13(10):2399. https://doi.org/10.3390/cancers13102399
Chicago/Turabian StyleCannon-Albright, Lisa Anne, Craig Carl Teerlink, Jeff Stevens, Franklin W. Huang, Csilla Sipeky, Johanna Schleutker, Rolando Hernandez, Julio Facelli, Neeraj Agarwal, and Donald L. Trump. 2021. "A Rare Variant in ERF (rs144812092) Predisposes to Prostate and Bladder Cancers in an Extended Pedigree" Cancers 13, no. 10: 2399. https://doi.org/10.3390/cancers13102399
APA StyleCannon-Albright, L. A., Teerlink, C. C., Stevens, J., Huang, F. W., Sipeky, C., Schleutker, J., Hernandez, R., Facelli, J., Agarwal, N., & Trump, D. L. (2021). A Rare Variant in ERF (rs144812092) Predisposes to Prostate and Bladder Cancers in an Extended Pedigree. Cancers, 13(10), 2399. https://doi.org/10.3390/cancers13102399