
HCV Proteins Modulate the Host Cell miRNA Expression Contributing to Hepatitis C Pathogenesis and Hepatocellular Carcinoma Development

 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
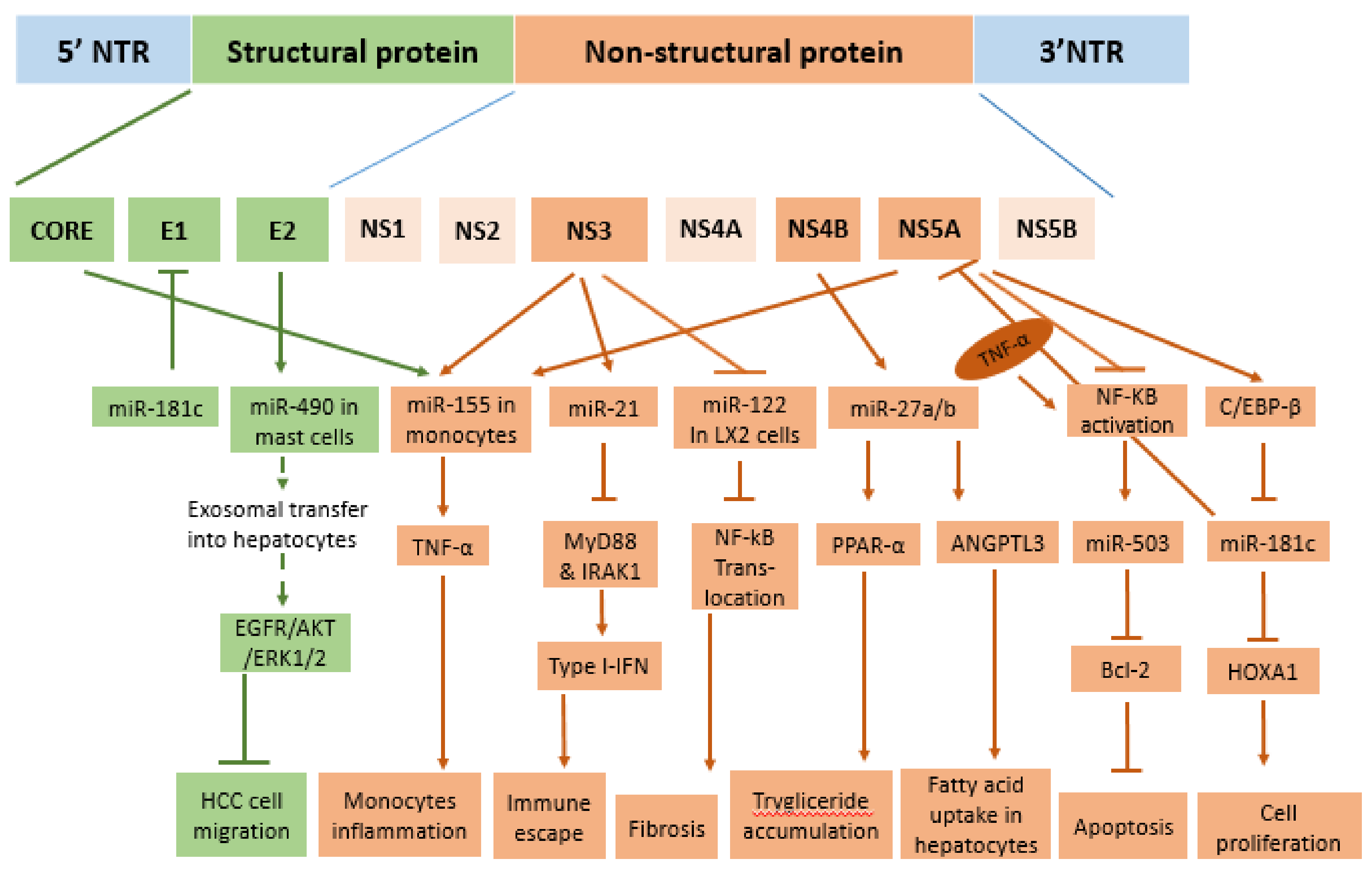
2. HCV Proteins and Their Role in Infection
3. HCV Core Protein Modulates Multiple miRNAs in Host Cells
3.1. MiRNAs Induced by HCV Core Protein Inhibit the Interferon Response and Promote Viral Replication
3.2. HCV Core Protein Alters Hepatic Lipid Metabolism Through miRNA-Dependent Mechanisms
3.3. From Liver Fibrosis to Liver Tumor: The Role of HCV Core Protein in the Dysregulation of Intracellular miRNAs
4. E2 Structural Protein Stimulates the Release of Exosomal miR-490 to Inhibit Cell Migration
5. Non-Structural Proteins in the Regulation of Cellular miRNA Expression
6. The Role of HCV-Induced miRNAs in Liver Carcinogenesis
6.1. HCV Proteins Determine the Up-Regulation of miRNA Involved in Liver Cancer Development
6.2. The Down-Regulation of Host Cell miRNAs Contribute to Liver Carcinogenesis
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- Lindenbach, B.D.; Rice, C.M. Unravelling Hepatitis C virus replication from genome to function. Nature 2005, 436, 933–938. [Google Scholar] [CrossRef]
- GBD 2015 Mortality and Causes of Death Collaborators. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the global burden of disease study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Hepatitis Report, 2017; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Farci, P.; Strazzera, R.; Alter, H.J.; Farci, S.; Degioannis, D.; Coiana, A.; Peddis, G.; Usai, F.; Serra, G.; Chessa, L.; et al. Early changes in Hepatitis C viral quasispecies during interferon therapy predict the therapeutic outcome. Proc. Natl. Acad. Sci. USA 2002, 99, 3081–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mchutchison, J.G.; Gordon, S.C.; Schiff, E.R.; Shiffman, M.L.; Lee, W.M.; Rustgi, V.K.; Goodman, Z.D.; Ling, M.-H.; Cort, S.; Albrecht, J.K. Interferon Alfa-2b alone or in combination with ribavirin as initial treatment for chronic Hepatitis C. N. Engl. J. Med. 1998, 339, 1485–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoratti, M.J.; Siddiqua, A.; Morassut, R.E.; Zeraatkar, D.; Chou, R.; van Holten, J.; Xie, F.; Druyts, E. Pangenotypic direct acting antivirals for the treatment of chronic Hepatitis C virus infection: A systematic literature review and meta-analysis. EClinicalMedicine 2020, 18, 100237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tato Marinho, R.A.R. The patient bomb: Sustained viral response after Hepatitis C in cirrhosis. GE Port. J. Gastroenterol. 2020, 27, 147–148. [Google Scholar] [CrossRef] [PubMed]
- Aleman, S.; Rahbin, N.; Weiland, O.; Davidsdottir, L.; Hedenstierna, M.; Rose, N.; Verbaan, H.; Stål, P.; Carlsson, T.; Norrgren, H.; et al. A risk for hepatocellular carcinoma persists long-term after sustained virologic response in patients with Hepatitis C-associated liver cirrhosis. Clin. Infect. Dis. 2013, 57, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Muzica, C.M.; Stanciu, C.; Huiban, L.; Singeap, A.-M.; Sfarti, C.; Zenovia, S.; Cojocariu, C.; Trifan, A. Hepatocellular carcinoma after direct-acting antiviral Hepatitis C virus therapy: A debate near the end. World J. Gastroenterol. 2020, 26, 6770–6781. [Google Scholar] [CrossRef] [PubMed]
- Pascut, D.; Pratama, M.Y.; Tiribelli, C. HCC occurrence after DAA treatments: Molecular tools to assess the post-treatment risk and surveillance. Hepat. Oncol. 2020, 7, HEP21. [Google Scholar] [CrossRef] [PubMed]
- Malespin, M.; Harris, C.; Kanar, O.; Jackman, K.; Smotherman, C.; Johnston, A.; Ferm, J.; de Melo, S.W.; Scolapio, J.S.; Nelson, D.R.; et al. Barriers to treatment of chronic Hepatitis C with direct acting antivirals in an urban clinic. Ann. Hepatol. 2019, 18, 304–309. [Google Scholar] [CrossRef]
- Randall, G.; Panis, M.; Cooper, J.D.; Tellinghuisen, T.L.; Sukhodolets, K.E.; Pfeffer, S.; Landthaler, M.; Landgraf, P.; Kan, S.; Lindenbach, B.D.; et al. Cellular cofactors affecting Hepatitis C virus infection and replication. Proc. Natl. Acad. Sci. USA 2007, 104, 12884–12889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jopling, C.L.; Norman, K.L.; Sarnow, P. Positive and negative modulation of viral and Cellular mRNAs by liver-specific microRNA miR-122. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 369–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’souza, S.; Lau, K.C.; Coffin, C.S.; Patel, T.R. Molecular mechanisms of viral hepatitis induced hepatocellular carcinoma. World J. Gastroenterol. 2020, 26, 5759–5783. [Google Scholar] [CrossRef]
- Mahmoudvand, S.; Shokri, S.; Taherkhani, R.; Farshadpour, F. Hepatitis C virus core protein modulates several signaling pathways involved in hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.; Lejamtel, C.; Benzoubir, N.; Battaglia, S.; Sidahmed-Adrar, N.; Desterke, C.; Lemasson, M.; Rosenberg, A.R.; Samuel, D.; Bréchot, C.; et al. Hepatitis C virus core protein targets 4E-BP1 expression and phosphorylation and potentiates myc-induced liver carcinogenesis in transgenic mice. Oncotarget 2017, 8, 56228–56242. [Google Scholar] [CrossRef] [Green Version]
- Niepmann, M.; Gerresheim, G.K. Hepatitis C virus translation regulation. Int. J. Mol. Sci. 2020, 21, 2328. [Google Scholar] [CrossRef] [Green Version]
- Lavie, M.; Goffard, A.; Dubuisson, J. HCV glycoproteins: Assembly of a functional E1–E2 heterodimer. In Hepatitis C Viruses: Genomes and Molecular Biology; Tan, S.-L., Ed.; Horizon Bioscience: Norfolk, UK, 2006; ISBN 978-1-904933-20-5. [Google Scholar]
- Ashfaq, U.A.; Javed, T.; Rehman, S.; Nawaz, Z.; Riazuddin, S. An overview of HCV molecular biology, replication and immune responses. Virol. J. 2011, 8, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gawlik, K.; Gallay, P.A. HCV core protein and virus assembly: What we know without structures. Immunol. Res. 2014, 60, 1–10. [Google Scholar] [CrossRef]
- Tong, Y.; Lavillette, D.; Li, Q.; Zhong, J. Role of Hepatitis C virus envelope glycoprotein E1 in virus entry and assembly. Front. Immunol. 2018, 9, 1411. [Google Scholar] [CrossRef]
- Wünschmann, S.; Medh, J.D.; Klinzmann, D.; Schmidt, W.N.; Stapleton, J.T. Characterization of Hepatitis C virus (HCV) and HCV E2 interactions with CD81 and the low-density lipoprotein receptor. J. Virol. 2000, 74, 10055–10062. [Google Scholar] [CrossRef] [Green Version]
- Douam, F.; Dao Thi, V.L.; Maurin, G.; Fresquet, J.; Mompelat, D.; Zeisel, M.B.; Baumert, T.F.; Cosset, F.-L.; Lavillette, D. Critical interaction between E1 and E2 glycoproteins determines binding and fusion properties of Hepatitis C virus during cell entry. Hepatology 2014, 59, 776–788. [Google Scholar] [CrossRef]
- Tscherne, D.M.; Jones, C.T.; Evans, M.J.; Lindenbach, B.D.; McKeating, J.A.; Rice, C.M. Time- and temperature-dependent activation of Hepatitis C virus for low-pH-triggered entry. J. Virol. 2006, 80, 1734–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, I.C. The Hepatitis C virus nonstructural protein 2 (NS2): An up-and-coming antiviral drug target. Viruses 2010, 2, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Dimitrova, M.; Imbert, I.; Kieny, M.P.; Schuster, C. Protein-protein interactions between Hepatitis C virus nonstructural proteins. J. Virol. 2003, 77, 5401–5414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolykhalov, A.A.; Mihalik, K.; Feinstone, S.M.; Rice, C.M. Hepatitis C virus-encoded enzymatic activities and conserved RNA elements in the 3′ nontranslated region are essential for virus replication in vivo. J. Virol. 2000, 74, 2046–2051. [Google Scholar] [CrossRef] [Green Version]
- Blight, K.J. Charged residues in Hepatitis C virus NS4B are critical for multiple NS4B functions in RNA replication. J. Virol. 2011, 85, 8158–8171. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, K.; Lange, C.M.; Gouttenoire, J.; Meylan, E.; Brass, V.; Penin, F.; Moradpour, D. Nonstructural protein 3-4A: The swiss army knife of Hepatitis C virus. J. Viral. Hepat. 2011, 18, 305–315. [Google Scholar] [CrossRef]
- Koch, J.O.; Bartenschlager, R. Modulation of Hepatitis C virus NS5A hyperphosphorylation by nonstructural proteins NS3, NS4A, and NS4B. J. Virol. 1999, 73, 7138–7146. [Google Scholar] [CrossRef] [Green Version]
- Paterson, M.; Laxton, C.D.; Thomas, H.C.; Ackrill, A.M.; Foster, G.R. Hepatitis C virus NS5A protein inhibits interferon antiviral activity, but the effects do not correlate with clinical response. Gastroenterology 1999, 117, 1187–1197. [Google Scholar] [CrossRef]
- Atoom, A.M.; Taylor, N.G.A.; Russell, R.S. The elusive function of the Hepatitis C virus P7 protein. Virology 2014, 462–463, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, M.B. MiR-122: A key factor and therapeutic target in liver disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunden, R.D.; Khan, J.Q.; Ghezelbash, S.; Wilson, J.A. The role of the liver-specific microRNA, miRNA-122 in the HCV replication cycle. Int. J. Mol. Sci. 2020, 21, 5677. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, I.M.; Cheng, G.; Wieland, S.; Volinia, S.; Croce, C.M.; Chisari, F.V.; David, M. Interferon modulation of cellular microRNAs as an antiviral mechanism. Nature 2007, 449, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Nicolas, E.; Marks, D.; Sander, C.; Lerro, A.; Buendia, M.A.; Xu, C.; Mason, W.S.; Moloshok, T.; Bort, R.; et al. MiR-122, a mammalian liver-specific microRNA, is processed from hcr mRNA and may downregulate the high affinity cationic amino acid transporter CAT-1. RNA Biol. 2004, 1, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Henke, J.I.; Goergen, D.; Zheng, J.; Song, Y.; Schüttler, C.G.; Fehr, C.; Jünemann, C.; Niepmann, M. microRNA-122 stimulates translation of Hepatitis C virus RNA. EMBO J. 2008, 27, 3300–3310. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Xing, X.; Yang, Q.; Xu, H.; He, J.; Chen, Z.; Zhu, H. The effects of Hepatitis C virus core protein on the expression of miR-122 in vitro. Virol. J. 2013, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez, R.T.; Bandyopadhyay, S.; Wendlandt, E.B.; Keck, K.; Hoffer, B.A.; Icardi, M.S.; Christensen, R.N.; Schmidt, W.N.; McCaffrey, A.P. Correlation between microRNA expression levels and clinical parameters associated with chronic Hepatitis C viral infection in humans. Lab. Investig. 2010, 90, 1727–1736. [Google Scholar] [CrossRef] [Green Version]
- Sarasin-Filipowicz, M.; Krol, J.; Markiewicz, I.; Heim, M.H.; Filipowicz, W. Decreased levels of microRNA miR-122 in individuals with Hepatitis C responding poorly to interferon therapy. Nat. Med. 2009, 15, 31–33. [Google Scholar] [CrossRef]
- Morita, K.; Taketomi, A.; Shirabe, K.; Umeda, K.; Kayashima, H.; Ninomiya, M.; Uchiyama, H.; Soejima, Y.; Maehara, Y. Clinical significance and potential of hepatic microRNA-122 expression in Hepatitis C. Liver Int. 2011, 31, 474–484. [Google Scholar] [CrossRef]
- Kim, G.-W.; Lee, S.-H.; Cho, H.; Kim, M.; Shin, E.-C.; Oh, J.-W. Hepatitis C virus core protein promotes miR-122 destabilization by inhibiting GLD-2. PLoS Pathog. 2016, 12, e1005714. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Eckmann, C.R.; Kadyk, L.C.; Wickens, M.; Kimble, J. A regulatory cytoplasmic poly(A) polymerase in caenorhabditis elegans. Nature 2002, 419, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Nakel, K.; Bonneau, F.; Eckmann, C.R.; Conti, E. Structural basis for the activation of the C. Elegans noncanonical cytoplasmic poly(A)-polymerase GLD-2 by GLD-3. Proc. Natl. Acad. Sci. USA 2015, 112, 8614–8619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chen, J.; Wang, H.; Shi, J.; Wu, K.; Liu, S.; Liu, Y.; Wu, J. HCV-induced miR-21 contributes to evasion of host immune system by targeting MyD88 and IRAK1. PLoS Pathog. 2013, 9, e1003248. [Google Scholar] [CrossRef]
- He, C.-L.; Liu, M.; Tan, Z.-X.; Hu, Y.-J.; Zhang, Q.-Y.; Kuang, X.-M.; Kong, W.-L.; Mao, Q. Hepatitis c virus core protein-induced miR-93-5p up-regulation inhibits interferon signaling pathway by targeting IFNAR1. World J. Gastroenterol. 2018, 24, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Liu, H.; Yin, Q.; Li, H.; Gao, H. MiR-93 enhances hepatocellular carcinoma invasion and metastasis by EMT via targeting PDCD4. Biotechnol. Lett. 2017, 39, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- de Weerd, N.A.; Matthews, A.Y.; Pattie, P.R.; Bourke, N.M.; Lim, S.S.; Vivian, J.P.; Rossjohn, J.; Hertzog, P.J. A hot spot on interferon α/β receptor subunit 1 (IFNAR1) underpins its interaction with interferon-β and dictates signaling. J. Biol. Chem. 2017, 292, 7554–7565. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Xu, Y.; Jiang, L.; Cao, X.; Liu, F.; Li, H.; Zhang, L.; Li, Z.; Li, J.; Ye, J.; et al. Differentially expressed microRNAs in huh-7 cells expressing HCV core genotypes 3a or 1b: Potential functions and downstream pathways. Int. J. Mol. Med. 2012, 30, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Balan, V.; Hernandez, J.; Rosati, M.; Williams, J.; Rodriguez-Luna, H.; Schwartz, J.; Harrison, E.; Anderson, M.; Byrne, T.; et al. Hepatic steatosis in Hepatitis C virus genotype 3 infection: Does it correlate with body mass index, fibrosis, and HCV risk factors? Dig. Dis. Sci. 2004, 49, 25–29. [Google Scholar] [CrossRef]
- Chan, A.; Patel, K.; Naggie, S. Genotype 3 Infection:The last stand of Hepatitis C virus. Drugs 2017, 77, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Soupene, E.; Kuypers, F.A. Mammalian long-chain Acyl-CoA synthetases. Exp. Biol. Med. 2008, 233, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Clément, S.; Sobolewski, C.; Gomes, D.; Rojas, A.; Goossens, N.; Conzelmann, S.; Calo, N.; Negro, F.; Foti, M. Activation of the oncogenic miR-21-5p promotes HCV replication and steatosis induced by the viral core 3a protein. Liver Int. 2019, 39, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, M.; Foti, M. PTEN at the crossroad of metabolic diseases and cancer in the liver. Ann. Hepatol. 2008, 7, 192–199. [Google Scholar] [CrossRef]
- Singaravelu, R.; Chen, R.; Lyn, R.K.; Jones, D.M.; O’Hara, S.; Rouleau, Y.; Cheng, J.; Srinivasan, P.; Nasheri, N.; Russell, R.S.; et al. Hepatitis C virus induced up-regulation of microRNA-27: A novel mechanism for hepatic steatosis. Hepatology 2014, 59, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Nasheri, N.; Joyce, M.; Rouleau, Y.; Yang, P.; Yao, S.; Tyrrell, D.L.; Pezacki, J.P. Modulation of fatty acid synthase enzyme activity and expression during Hepatitis C virus replication. Chem. Biol. 2013, 20, 570–582. [Google Scholar] [CrossRef]
- Dharancy, S.; Malapel, M.; Perlemuter, G.; Roskams, T.; Cheng, Y.; Dubuquoy, L.; Podevin, P.; Conti, F.; Canva, V.; Philippe, D.; et al. Impaired expression of the peroxisome proliferator-activated receptor alpha during Hepatitis C virus infection. Gastroenterology 2005, 128, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackel-Cram, C.; Qiao, L.; Xiang, Z.; Brownlie, R.; Zhou, Y.; Babiuk, L.; Liu, Q. Hepatitis C virus genotype-3a core protein enhances sterol regulatory element-binding protein-1 activity through the phosphoinositide 3-kinase-Akt-2 pathway. J. Gen. Virol. 2010, 91, 1388–1395. [Google Scholar] [CrossRef]
- Lyn, R.K.; Kennedy, D.C.; Sagan, S.M.; Blais, D.R.; Rouleau, Y.; Pegoraro, A.F.; Xie, X.S.; Stolow, A.; Pezacki, J.P. Direct imaging of the disruption of Hepatitis C virus replication complexes by inhibitors of lipid metabolism. Virology 2009, 394, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Hishiki, T.; Sugiyama, K.; Ogawa, K.; Funami, K.; Kato, A.; Ohsaki, Y.; Fujimoto, T.; Takaku, H.; Shimotohno, K. lipoprotein lipase and hepatic triglyceride lipase reduce the infectivity of Hepatitis C virus (HCV) through their catalytic activities on HCV-associated lipoproteins. Virology 2010, 407, 152–159. [Google Scholar] [CrossRef]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Eberlé, D.; Hegarty, B.; Bossard, P.; Ferré, P.; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef]
- Yang, H.-X.; Zhang, M.; Long, S.-Y.; Tuo, Q.-H.; Tian, Y.; Chen, J.-X.; Zhang, C.-P.; Liao, D.-F. Cholesterol in LDL receptor recycling and degradation. Clin. Chim. Acta 2020, 500, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Cholesterol Metabolism and the Pathogenesis of Non-Alcoholic Steatohepatitis. Prog Lipid Res 2013, 52, 175–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Wang, Q.; Liu, S.-A.; Zhang, J.-Q.; Ju, W.; Quan, M.; Feng, S.-H.; Dong, J.-L.; Gao, P.; Cheng, J. MicroRNA-185-5p mediates regulation of SREBP2 expression by Hepatitis C virus core protein. World J. Gastroenterol. 2015, 21, 4517–4525. [Google Scholar] [CrossRef] [PubMed]
- Rotllan, N.; Fernández-Hernando, C. MicroRNA Regulation of Cholesterol Metabolism. Cholesterol 2012, 2012, 847849. [Google Scholar] [CrossRef]
- Ramirez, C.M.; Dávalos, A.; Goedeke, L.; Salerno, A.G.; Warrier, N.; Cirera-Salinas, D.; Suárez, Y.; Fernández-Hernando, C. microRNA-758 regulates cholesterol efflux through posttranscriptional repression of ATP-binding cassette transporter A1. Arter. Thromb. Vasc. Biol. 2011, 31, 2707–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Fu, S.; Wang, J. Hepatitis C virus infection decreases the expression of toll-like receptors 3 and 7 via upregulation of miR-758. Arch. Virol. 2014, 159, 2997–3003. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Morgan, T.R. The natural history of Hepatitis C virus (HCV) infection. Int. J. Med. Sci. 2006, 3, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Gea, V.; Friedman, S.L. Pathogenesis of liver fibrosis. Annu. Rev. Pathol. 2011, 6, 425–456. [Google Scholar] [CrossRef]
- Chusri, P.; Kumthip, K.; Hong, J.; Zhu, C.; Duan, X.; Jilg, N.; Fusco, D.N.; Brisac, C.; Schaefer, E.A.; Cai, D.; et al. HCV induces transforming growth factor Β1 through activation of endoplasmic reticulum stress and the unfolded protein response. Sci. Rep. 2016, 6, 22487. [Google Scholar] [CrossRef]
- Lin, W.; Tsai, W.-L.; Shao, R.-X.; Wu, G.; Peng, L.F.; Barlow, L.L.; Chung, W.J.; Zhang, L.; Zhao, H.; Jang, J.-Y.; et al. Hepatitis C virus regulates transforming growth factor beta1 production through the generation of reactive oxygen species in a nuclear factor kappaB-dependent manner. Gastroenterology 2010, 138, 2509–2518.e1. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, C.H.; Lee, S.-W. Hepatitis C virus infection stimulates transforming growth factor-Β1 expression through up-regulating miR-192. J. Microbiol. 2016, 54, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Zhang, J.; Wang, M.; Lanting, L.; Yuan, H.; Rossi, J.J.; Natarajan, R. microRNA-192 in diabetic kidney glomeruli and its function in TGF-beta-induced collagen expression via inhibition of E-box repressors. Proc. Natl. Acad. Sci. USA 2007, 104, 3432–3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, M.; Putta, S.; Wang, M.; Yuan, H.; Lanting, L.; Nair, I.; Gunn, A.; Nakagawa, Y.; Shimano, H.; Todorov, I.; et al. TGF-beta activates Akt kinase through a microRNA-dependent amplifying circuit targeting PTEN. Nat. Cell Biol. 2009, 11, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Postigo, A.A. Opposing functions of ZEB proteins in the regulation of the TGFbeta/BMP signaling pathway. EMBO J. 2003, 22, 2443–2452. [Google Scholar] [CrossRef]
- Kato, M.; Dang, V.; Wang, M.; Park, J.T.; Deshpande, S.; Kadam, S.; Mardiros, A.; Zhan, Y.; Oettgen, P.; Putta, S.; et al. TGF-β induces acetylation of chromatin and of Ets-1 to alleviate repression of miR-192 in diabetic nephropathy. Sci. Signal. 2013, 6, ra43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Grunsven, L.A.; Schellens, A.; Huylebroeck, D.; Verschueren, K. SIP1 (smad interacting protein 1) and DeltaEF1 (delta-crystallin enhancer binding factor) are structurally similar transcriptional repressors. J. Bone Jt. Surg. Am. 2001, 83A (Suppl. 1), S40–S47. [Google Scholar]
- Li, H.-C.; Ma, H.-C.; Yang, C.-H.; Lo, S.-Y. Production and pathogenicity of Hepatitis C virus core gene products. World J. Gastroenterol. 2014, 20, 7104–7122. [Google Scholar] [CrossRef] [PubMed]
- Fukutomi, T.; Zhou, Y.; Kawai, S.; Eguchi, H.; Wands, J.R.; Li, J. Hepatitis C virus core protein stimulates hepatocyte growth: Correlation with upregulation of Wnt-1 expression. Hepatology 2005, 41, 1096–1105. [Google Scholar] [CrossRef]
- Li, B.; Xie, Z.; Li, B. MiR-152 functions as a tumor suppressor in colorectal cancer by targeting PIK3R3. Tumour Biol. 2016, 37, 10075–10084. [Google Scholar] [CrossRef]
- Yin, T.; Liu, M.-M.; Jin, R.-T.; Kong, J.; Wang, S.-H.; Sun, W.-B. MiR-152-3p modulates hepatic carcinogenesis by targeting cyclin-dependent kinase 8. Pathol. Res. Pract. 2019, 215, 152406. [Google Scholar] [CrossRef]
- Huang, S.; Xie, Y.; Yang, P.; Chen, P.; Zhang, L. HCV core protein-induced down-regulation of microRNA-152 promoted aberrant proliferation by regulating Wnt1 in HepG2 cells. PLoS ONE 2014, 9, e81730. [Google Scholar] [CrossRef]
- Xu, H.; Li, G.; Yue, Z.; Li, C. HCV core protein-induced upregulation of microRNA-196a promotes aberrant proliferation in hepatocellular carcinoma by targeting FOXO1. Mol. Med. Rep. 2016, 13, 5223–5229. [Google Scholar] [CrossRef] [PubMed]
- Myatt, S.S.; Lam, E.W.-F. The emerging roles of forkhead box (fox) proteins in cancer. Nat. Rev. Cancer 2007, 7, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Quan, H.; Zhou, F.; Nie, D.; Chen, Q.; Cai, X.; Shan, X.; Zhou, Z.; Chen, K.; Huang, A.; Li, S.; et al. Hepatitis C virus core protein epigenetically silences SFRP1 and enhances HCC aggressiveness by inducing epithelial-mesenchymal transition. Oncogene 2014, 33, 2826–2835. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wu, J.; Liu, M.; Yin, H.; He, J.; Zhang, B. Downregulation of miRNA-30c and miR-203a is associated with Hepatitis C virus core protein-induced epithelial-mesenchymal transition in normal hepatocytes and hepatocellular carcinoma cells. Biochem. Biophys. Res. Commun. 2015, 464, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Li, W.-C.; Hellem, M.R.; Rostad, K.; Popa, M.; McCormack, E.; Oyan, A.M.; Kalland, K.-H.; Ke, X.-S. MiR-182 and miR-203 induce mesenchymal to epithelial transition and self-sufficiency of growth signals via repressing SNAI2 in prostate cells. Int. J. Cancer 2013, 133, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Xia, Y.; Wang, P.; Liu, B.; Chen, Y. Low expression of microRNA-30c promotes invasion by inducing epithelial mesenchymal transition in non-small cell lung cancer. Mol. Med. Rep. 2014, 10, 2575–2579. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, H.; Liu, J.; Tu, X.; Zang, Y.; Zhu, J.; Chen, J.; Dong, L.; Zhang, J. MiR-30 inhibits TGF-Β1-induced epithelial-to-mesenchymal transition in hepatocyte by targeting snail1. Biochem. Biophys. Res. Commun. 2012, 417, 1100–1105. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, Y.; Kitamoto, M.; Nakanishi, T.; Yasui, W.; Tahara, E.; Nakayama, J.; Ishikawa, F.; Tahara, H.; Ide, T.; Kajiyama, G. Immuno-histochemical detection of human telomerase reverse transcriptase in human liver tissues. Oncogene 2000, 19, 3888–3893. [Google Scholar] [CrossRef] [Green Version]
- Shiu, T.-Y.; Shih, Y.-L.; Feng, A.-C.; Lin, H.-H.; Huang, S.-M.; Huang, T.-Y.; Hsieh, C.-B.; Chang, W.-K.; Hsieh, T.-Y. HCV core inhibits hepatocellular carcinoma cell replicative senescence through downregulating microRNA-138 expression. J. Mol. Med. 2017, 95, 629–639. [Google Scholar] [CrossRef]
- Xiong, L.; Zhen, S.; Yu, Q.; Gong, Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol. Lett. 2017, 14, 2141–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Liu, Y.; Gao, Q.; Han, Y.; Zhang, G.; Xu, S.; Cheng, K.; Zou, W. MiR-490-3p inhibits the growth and invasiveness in triple-negative breast cancer by repressing the expression of TNKS2. Gene 2016, 593, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, X.; Xiu, Y.-L.; Sun, K.-X.; Zhao, Y. MicroRNA-490-3P targets CDK1 and inhibits ovarian epithelial carcinoma tumorigenesis and progression. Cancer Lett. 2015, 362, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, R.; Li, Z.; Huang, N.; Wu, X.; Li, S.; Li, Y.; Wu, S. MicroRNA-490-3p inhibits colorectal cancer metastasis by targeting TGFβR1. BMC Cancer 2015, 15, 1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, M.; Li, X.; Tang, H. MiR-490-3p modulates cell growth and epithelial to mesenchymal transition of hepatocellular carcinoma cells by targeting endoplasmic reticulum-golgi intermediate compartment protein 3 (ERGIC3). J. Biol. Chem. 2013, 288, 4035–4047. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; He, J.; Liu, Y. MiR-490-3p inhibits autophagy via targeting ATG7 in hepatocellular carcinoma. IUBMB Life 2018, 70, 468–478. [Google Scholar] [CrossRef] [Green Version]
- Khanizadeh, S.; Ravanshad, M.; Hosseini, S.Y.; Davoodian, P.; Almasian, M.; Khanlari, Z. The effect of the Hepatitis C virus (HCV) NS3 protein on the expression of miR-150, miR-199a, miR-335, miR-194 and miR-27a. Microb. Pathog. 2017, 110, 688–693. [Google Scholar] [CrossRef]
- Zhang, J.; Ishigaki, Y.; Takegami, T. Hepatitis C virus NS3 protein modulates the biological behaviors of malignant hepatocytes by altering the expression of host cell microRNA. Mol. Med. Rep. 2015, 12, 5109–5115. [Google Scholar] [CrossRef] [Green Version]
- Heim, M.H.; Thimme, R. Innate and adaptive immune responses in HCV infections. J. Hepatol. 2014, 61, S14–S25. [Google Scholar] [CrossRef] [Green Version]
- Beutler, B.; Eidenschenk, C.; Crozat, K.; Imler, J.-L.; Takeuchi, O.; Hoffmann, J.A.; Akira, S. Genetic analysis of resistance to viral infection. Nat. Rev. Immunol. 2007, 7, 753–766. [Google Scholar] [CrossRef]
- Pompili, M.; Biolato, M.; Miele, L.; Grieco, A. Tumor necrosis factor-α inhibitors and chronic Hepatitis C: A comprehensive literature review. World J. Gastroenterol. 2013, 19, 7867–7873. [Google Scholar] [CrossRef]
- Nelson, D.R.; Lim, H.L.; Marousis, C.G.; Fang, J.W.S.; Davis, G.L.; Shen, L.; Urdea, M.S.; Kolberg, J.A.; Lau, J.Y.N. Activation of tumor necrosis factor-α system in chronic Hepatitis C virus infection. Dig. Dis. Sci. 1997, 42, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Cimino, A.; Costinean, S.; Dumitru, C.D.; Adair, B.; Fabbri, M.; Alder, H.; Liu, C.G.; Calin, G.A.; et al. Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock. J. Immunol. 2007, 179, 5082–5089. [Google Scholar] [CrossRef]
- Bala, S.; Tilahun, Y.; Taha, O.; Alao, H.; Kodys, K.; Catalano, D.; Szabo, G. Increased microRNA-155 expression in the serum and peripheral monocytes in chronic HCV Infection. J. Transl. Med. 2012, 10, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, J.; Lin, J.; He, L.; Huang, J.; Liu, Q. TNF-α/MiR-155 axis induces the transformation of osteosarcoma cancer stem cells independent of TP53INP1. Gene 2020, 726, 144224. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Xiao, Z.; Wang, F. Hepatitis C virus nonstructural 5a protein (HCV-NS5A) inhibits hepatocyte apoptosis through the NF-Κb/MiR-503/Bcl-2 pathway. Mol. Cells 2017, 40, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Caporali, A.; Meloni, M.; Nailor, A.; Mitić, T.; Shantikumar, S.; Riu, F.; Sala-Newby, G.B.; Rose, L.; Besnier, M.; Katare, R.; et al. P75(NTR)-dependent activation of NF-ΚB regulates microRNA-503 transcription and pericyte-endothelial crosstalk in diabetes after limb ischaemia. Nat. Commun. 2015, 6, 8024. [Google Scholar] [CrossRef] [Green Version]
- Sakata, K.; Hara, M.; Terada, T.; Watanabe, N.; Takaya, D.; Yaguchi, S.; Matsumoto, T.; Matsuura, T.; Shirouzu, M.; Yokoyama, S.; et al. HCV NS3 protease enhances liver fibrosis via binding to and activating TGF-β type I receptor. Sci. Rep. 2013, 3, 3243. [Google Scholar] [CrossRef] [PubMed]
- Selimovic, D.; El-Khattouti, A.; Ghozlan, H.; Haikel, Y.; Abdelkader, O.; Hassan, M. Hepatitis C virus-related hepatocellular carcinoma: An insight into molecular mechanisms and therapeutic strategies. World J. Hepatol. 2012, 4, 342–355. [Google Scholar] [CrossRef]
- Chen, C.; Wu, C.-Q.; Zhang, Z.-Q.; Yao, D.-K.; Zhu, L. Loss of expression of miR-335 is implicated in hepatic stellate cell migration and activation. Exp. Cell Res. 2011, 317, 1714–1725. [Google Scholar] [CrossRef]
- Venugopal, S.K.; Jiang, J.; Kim, T.-H.; Li, Y.; Wang, S.-S.; Torok, N.J.; Wu, J.; Zern, M.A. Liver fibrosis causes downregulation of miRNA-150 and miRNA-194 in hepatic stellate cells, and their overexpression causes decreased stellate cell activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G101–G106. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Zhang, J.; Huang, G.; Qian, J.; Wang, X.; Mei, S. Over-expressed microRNA-27a and 27b influence fat accumulation and cell proliferation during rat hepatic stellate cell activation. FEBS Lett. 2009, 583, 759–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanizadeh, S.; Ravanshad, M.; Hosseini, S.Y.; Davoodian, P.; Zadeh, A.N.; Sabahi, F.; Sarvari, J.; Khanlari, Z.; Hasani-Azad, M. The possible role of NS3 protease activity of Hepatitis C virus on fibrogenesis and miR-122 expression in hepatic stellate cells. Acta Virol. 2016, 60, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Kanda, T.; Sasaki, R.; Haga, Y.; Jiang, X.; Wu, S.; Nakamoto, S.; Yokosuka, O. MicroRNA-122 inhibits the production of inflammatory cytokines by targeting the PKR activator PACT in human hepatic stellate cells. PLoS ONE 2015, 10, e0144295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, K.; Miyaaki, H.; Ichikawa, T. Antitumor function of microRNA-122 against hepatocellular carcinoma. J. Gastroenterol. 2014, 49, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Gramantieri, L.; Ferracin, M.; Fornari, F.; Veronese, A.; Sabbioni, S.; Liu, C.-G.; Calin, G.A.; Giovannini, C.; Ferrazzi, E.; Grazi, G.L.; et al. Cyclin G1 is a target of miR-122a, a microRNA frequently down-regulated in human hepatocellular carcinoma. Cancer Res. 2007, 67, 6092–6099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.-C.; Hsu, P.W.-C.; Lai, T.-C.; Chau, G.-Y.; Lin, C.-W.; Chen, C.-M.; Lin, C.-D.; Liao, Y.-L.; Wang, J.-L.; Chau, Y.-P.; et al. MicroRNA-122, a tumor suppressor microRNA that regulates intrahepatic metastasis of hepatocellular carcinoma. Hepatology 2009, 49, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Shrivastava, S.; Bhanja Chowdhury, J.; Ray, R.; Ray, R.B. Transcriptional suppression of miR-181c by Hepatitis C virus enhances homeobox a1 expression. J. Virol. 2014, 88, 7929–7940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; de la Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The promising role of miR-21 as a cancer biomarker and its importance in RNA-based therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-H.; Tsao, C.-J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chu, Y.; Xu, M.; Zhang, X.; Zhou, Y. MiR-21 promotes cell migration and invasion of hepatocellular carcinoma by targeting KLF5. Oncol. Lett. 2019, 17, 2221–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demerdash, H.M.; Hussien, H.M.; Hassouna, E.; Arida, E.A. Detection of microRNA in hepatic cirrhosis and hepatocellular carcinoma in Hepatitis C genotype-4 in egyptian patients. BioMed Res. Int. 2017, 2017, e1806069. [Google Scholar] [CrossRef] [PubMed]
- Karakatsanis, A.; Papaconstantinou, I.; Gazouli, M.; Lyberopoulou, A.; Polymeneas, G.; Voros, D. Expression of microRNAs, miR-21, miR-31, miR-122, miR-145, miR-146a, miR-200c, miR-221, miR-222, and miR-223 in patients with hepatocellular carcinoma or intrahepatic cholangiocarcinoma and its prognostic significance. Mol. Carcinog. 2013, 52, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Pineau, P.; Volinia, S.; McJunkin, K.; Marchio, A.; Battiston, C.; Terris, B.; Mazzaferro, V.; Lowe, S.W.; Croce, C.M.; Dejean, A. MiR-221 overexpression contributes to liver tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 264–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; He, X.-X.; Chang, Y.; Sun, S.-Z.; Xu, C.-R.; Lin, J.-S. Downregulation of miR-93 expression reduces cell proliferation and clonogenicity of HepG2 Cells. Hepatogastroenterology 2012, 59, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Hoshino, H.; Wang, J.; Ono, S.; Iida, Y.; Hata, K.; Huang, S.K.; Colquhoun, S.; Hoon, D.S.B. MicroRNA-93 activates c-Met/PI3K/Akt pathway activity in hepatocellular carcinoma by directly inhibiting PTEN and CDKN1A. Oncotarget 2014, 6, 3211–3224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Liu, T.-T.; Yu, X.-N.; Balakrishnan, A.; Zhu, H.-R.; Guo, H.-Y.; Zhang, G.-C.; Bilegsaikhan, E.; Sun, J.-L.; Song, G.-Q.; et al. MicroRNA-93-5p promotes hepatocellular carcinoma progression via a microRNA-93-5p/MAP3K2/c-Jun positive feedback circuit. Oncogene 2020, 39, 5768–5781. [Google Scholar] [CrossRef]
- Li, X.; Xu, M.; Ding, L.; Tang, J. MiR-27a: A novel biomarker and potential therapeutic target in tumors. J. Cancer 2019, 10, 2836–2848. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Delogu, S.; Ho, C.; Lee, S.A.; Gui, B.; Jiang, L.; Ladu, S.; Cigliano, A.; Dombrowski, F.; Evert, M.; et al. Inactivation of Spry2 accelerates AKT-driven hepatocarcinogenesis via activation of MAPK and PKM2 pathways. J. Hepatol. 2012, 57, 577–583. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Zhang, J.; Lin, J.; Zhang, C.; Sun, S. Expression and function of microRNA-27b in hepatocellular carcinoma. Mol. Med. Rep. 2016, 13, 2801–2808. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Zheng, J.; Guo, X.; Gao, X.; Liu, H.; Tu, Y.; Zhang, Y. Reduced expression of SOCS2 and SOCS6 in hepatocellular carcinoma correlates with aggressive tumor progression and poor prognosis. Mol. Cell. Biochem. 2013, 378, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kaltenecker, D.; Themanns, M.; Mueller, K.M.; Spirk, K.; Suske, T.; Merkel, O.; Kenner, L.; Luís, A.; Kozlov, A.; Haybaeck, J.; et al. Hepatic growth hormone-JAK2-STAT5 signalling: Metabolic function, non-alcoholic fatty liver disease and hepatocellular carcinoma progression. Cytokine 2019, 124, 154569. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Wu, S.; Wu, Y.; Liu, T.; Zhao, X.; Li, Y. MicroRNA-196a/-196b regulate the progression of hepatocellular carcinoma through modulating the JAK/STAT pathway via targeting SOCS2. Cell Death Dis. 2019, 10, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-Y.; Chen, C.-L.; Hu, Y.-C.; Chi, Y.; Huang, Y.-H.; Su, C.-W.; Jeng, W.-J.; Liang, Y.-J.; Wu, J.-C. High expression of microRNA-196a is associated with progression of hepatocellular carcinoma in younger patients. Cancers 2019, 11, 1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdmanis, P.N.; Kim, H.K.; Chu, K.; Zhang, F.; Xu, J.; Munding, E.M.; Shen, J.; Kay, M.A. MiR-122 removal in the liver activates imprinted microRNAs and enables more effective microRNA-mediated gene repression. Nat. Commun. 2018, 9, 5321. [Google Scholar] [CrossRef]
- Thakral, S.; Ghoshal, K. MiR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr. Gene Ther. 2015, 15, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, H.L.A.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Fornari, F.; Gramantieri, L.; Giovannini, C.; Veronese, A.; Ferracin, M.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Tavolari, S.; et al. MiR-122/Cyclin G1 interaction modulates P53 activity and affects doxorubicin sensitivity of human hepatocarcinoma cells. Cancer Res. 2009, 69, 5761–5767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, C.; Wang, R.; Li, D.; Lin, X.-J.; Wei, Q.-K.; Yuan, Y.; Wang, Q.; Chen, W.; Zhuang, S.-M. A novel GSK-3 beta-C/EBP alpha-miR-122-insulin-like growth factor 1 receptor regulatory circuitry in human hepatocellular carcinoma. Hepatology 2010, 52, 1702–1712. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, X.; Wu, L.; Yang, R.; Yang, Z.; Wang, Q.; Wu, F. MicroRNA-122 suppresses cell proliferation and induces cell apoptosis in hepatocellular carcinoma by directly targeting Wnt/β-catenin pathway. Liver Int. 2012, 32, 752–760. [Google Scholar] [CrossRef]
- Zhi, Q.; Zhu, J.; Guo, X.; He, S.; Xue, X.; Zhou, J.; Hu, B.; Li, H.; Chen, S.; Zhao, H.; et al. Metastasis-related miR-185 is a potential prognostic biomarker for hepatocellular carcinoma in early stage. Biomed. Pharm. 2013, 67, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Liu, S.; Han, M.; Feng, S.; Liang, J.; Li, Z.; Li, Y.; Lu, H.; Liu, T.; Ma, Y.; et al. MicroRNA-185 induces potent autophagy via Akt signaling in hepatocellular carcinoma. Tumour Biol. 2017, 39, 1010428317694313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Jin, W.; Jin, H.; Wang, X. MTOR in viral hepatitis and hepatocellular carcinoma: Function and treatment. BioMed Res. Int. 2014, 2014, 735672. [Google Scholar] [CrossRef] [PubMed]
- Qadir, X.V.; Han, C.; Lu, D.; Zhang, J.; Wu, T. MiR-185 inhibits hepatocellular carcinoma growth by targeting the DNMT1/PTEN/Akt pathway. Am. J. Pathol. 2014, 184, 2355–2364. [Google Scholar] [CrossRef] [Green Version]
- Murakami, Y.; Tamori, A.; Itami, S.; Tanahashi, T.; Toyoda, H.; Tanaka, M.; Wu, W.; Brojigin, N.; Kaneoka, Y.; Maeda, A.; et al. The expression level of miR-18b in hepatocellular carcinoma is associated with the grade of malignancy and prognosis. BMC Cancer 2013, 13, 99. [Google Scholar] [CrossRef] [Green Version]
- Lian, J.; Jing, Y.; Dong, Q.; Huan, L.; Chen, D.; Bao, C.; Wang, Q.; Zhao, F.; Li, J.; Yao, M.; et al. MiR-192, a prognostic indicator, targets the SLC39A6/snail pathway to reduce tumor metastasis in human hepatocellular carcinoma. Oncotarget 2016, 7, 2672–2683. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wei, X.; Sun, Y.; Gao, H.; Zheng, X.; Wong, L.L.; Jin, L.; Liu, N.; Hernandez, B.; Peplowska, K.; et al. MiR-192-5p silencing by genetic aberrations is a key event in hepatocellular carcinomas with cancer stem cell features. Cancer Res. 2019, 79, 941–953. [Google Scholar] [CrossRef] [Green Version]
- El-Araby, R.E.; Khalifa, M.A.; Zoheiry, M.M.; Zahran, M.Y.; Rady, M.I.; Ibrahim, R.A.; El-Talkawy, M.D.; Essawy, F.M. The interaction between microRNA-152 and DNA methyltransferase-1 as an epigenetic prognostic biomarker in HCV-induced liver cirrhosis and hcc patients. Cancer Gene Ther. 2020, 27, 486–497. [Google Scholar] [CrossRef]
- Dang, Y.-W.; Zeng, J.; He, R.-Q.; Rong, M.-H.; Luo, D.-Z.; Chen, G. Effects of miR-152 on cell growth inhibition, motility suppression and apoptosis induction in hepatocellular carcinoma cells. Asian Pac. J. Cancer Prev. 2014, 15, 4969–4976. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, Y.; Qi, Y.; Yu, D.; Shao, Q.; Liang, J. MicroRNA-152 inhibits tumor cell growth by directly targeting RTKN in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Furuta, M.; Kozaki, K.; Tanaka, S.; Arii, S.; Imoto, I.; Inazawa, J. MiR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 2010, 31, 766–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, D.; Shen, S.; Fu, S.; Preston, B.; Brandon, C.; He, S.; Shen, C.; Wu, J.; Wang, S.; Xie, W.; et al. MiR-203 suppresses the proliferation and metastasis of hepatocellular carcinoma by targeting oncogene ADAM9 and oncogenic long non-coding RNA HULC. Anticancer. Agents Med. Chem. 2016, 16, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Wanjun, L.; Hui, S.; Dongyue, C.; Xinjun, Y.; Jisheng, Z. MiR-203 inhibits proliferation of HCC cells by targeting survivin. Cell Biochem. Funct. 2013, 31, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tong, D.; Guo, Q.; Wang, X.; Wu, F.; Li, Q.; Yang, J.; Zhao, L.; Qin, Y.; Liu, Y.; et al. HOXD3 targeted by miR-203a suppresses cell metastasis and angiogenesis through VEGFR in human hepatocellular carcinoma cells. Sci. Rep. 2018, 8, 2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksuz, Z.; Serin, M.S.; Kaplan, E.; Dogen, A.; Tezcan, S.; Aslan, G.; Emekdas, G.; Sezgin, O.; Altintas, E.; Tiftik, E.N. Serum microRNAs; miR-30c-5p, miR-223-3p, miR-302c-3p and miR-17-5p could be used as novel non-invasive biomarkers for HCV-positive cirrhosis and hepatocellular carcinoma. Mol. Biol. Rep. 2015, 42, 713–720. [Google Scholar] [CrossRef]
- McCann, J.V.; Xiao, L.; Kim, D.J.; Khan, O.F.; Kowalski, P.S.; Anderson, D.G.; Pecot, C.V.; Azam, S.H.; Parker, J.S.; Tsai, Y.S.; et al. Endothelial miR-30c suppresses tumor growth via inhibition of TGF-β-induced serpine1. J. Clin. Investig. 2019, 129, 1654–1670. [Google Scholar] [CrossRef]
- Zhao, D.-W.; Li, M.-M.; Han, J.-P.; Wang, Y.; Jiang, L.-X.; Chang, H.-L. MiR-30c exerts tumor suppressive functions in colorectal carcinoma by directly targeting BCL9. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3335–3343. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, L.-J.; Tan, Y.-X.; Ren, H.; Qi, Z.-T. MiR-138 induces cell cycle arrest by targeting cyclin D3 in hepatocellular carcinoma. Carcinogenesis 2012, 33, 1113–1120. [Google Scholar] [CrossRef]
- Yang, G.; Guo, S.; Liu, H.-T.; Yang, G. MiR-138-5p predicts negative prognosis and exhibits suppressive activities in hepatocellular carcinoma HCC by targeting FOXC1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8788–8800. [Google Scholar] [CrossRef]
- Huang, B.; Li, H.; Huang, L.; Luo, C.; Zhang, Y. Clinical significance of microRNA 138 and cyclin D3 in hepatocellular carcinoma. J. Surg. Res. 2015, 193, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.-Q.; Li, M.-C.; Zhang, Y.-Q.; Liu, X.-G. MiR-490-5p inhibits the metastasis of hepatocellular carcinoma by down-regulating E2F2 and ECT2. J. Cell. Biochem. 2018, 119, 8317–8324. [Google Scholar] [CrossRef]
- Xu, B.; Xu, T.; Liu, H.; Min, Q.; Wang, S.; Song, Q. MiR-490-5p suppresses cell proliferation and invasion by targeting BUB1 in hepatocellular carcinoma cells. Pharmacology 2017, 100, 269–282. [Google Scholar] [CrossRef]
- Yan, M.; Wang, C.; He, B.; Yang, M.; Tong, M.; Long, Z.; Liu, B.; Peng, F.; Xu, L.; Zhang, Y.; et al. Aurora-A kinase: A potent oncogene and target for cancer therapy. Med. Res. Rev. 2016, 36, 1036–1079. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-P.; Li, Z.-R. MiR-503-5p regulates cell epithelial-to-mesenchymal transition, metastasis and prognosis of hepatocellular carcinoma through inhibiting WEE1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2028–2037. [Google Scholar] [CrossRef]
- Li, B.; Liu, L.; Li, X.; Wu, L. MiR-503 suppresses metastasis of hepatocellular carcinoma cell by targeting PRMT1. Biochem. Biophys. Res. Commun. 2015, 464, 982–987. [Google Scholar] [CrossRef]
- Yang, X.; Zang, J.; Pan, X.; Yin, J.; Xiang, Q.; Yu, J.; Gan, R.; Lei, X. MiR-503 inhibits proliferation making human hepatocellular carcinoma cells susceptible to 5-fluorouracil by targeting EIF4E. Oncol. Rep. 2017, 37, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Gong, C.; Wu, J.; Gao, J.; Liu, W.; Liao, W.; Wu, L. MicroRNA-181c suppresses growth and metastasis of hepatocellular carcinoma by modulating NCAPG. Cancer Manag. Res. 2019, 11, 3455–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| miRNA | HCV Protein | miR-Expression | Cell Model | Animal Model | Effect |
|---|---|---|---|---|---|
| miR-122 [38,42] | Core | Down | Huh7.5.1, Huh7 | Positive regulator of infection at initial phases. The Inhibition of TENT-2, by core protein, affects the miR-122 maturation and function. HCV self-inhibitory effect. | |
| miR-21 [43] | Core Genotype 3a | Up | Huh-7 | mir21alox/lox (control) and Mir21a KO mice | Downregulates PTEN to promote lipid accumulation and steatosis. Promote viral replication. |
| miR-93 [44] | Core Genotype 1b | Up | Huh7 | IFNAR1 down-regulation to abolish IFN α pathway. | |
| miR-27a and miR-27b [45] | Core Genotype 1b | Up | Huh7.5 | SCID-beige/Alb-uPa mice infected with genotype 1a and 2b | Repression of PPAR-α, which led to triglycerides accumulation, and ANGPTL3, which caused an increase in lipoproteinlipase activity and fatty acid uptake. HCV self-inhibitory mechanism |
| miR-185 [46] | Core Genotype 1b | Down | HepG2 | Targets SREBP2, control of cholesterol homeostasis. | |
| miR-758 [47] | Core | Up | QSG-7701 | Regulation of the cholesterol metabolism, also controlling the cholesterol efflux through ABCA1 repression. TLR3 and TLR7 downregulation with a subsequent decrease in IFNα and IFNβ. | |
| miR-192 [48] | Core Genotype 1b | Up | Huh-7, Huh-7.5 | Downregulates ZEB1 responsible for the TGF-β1 inhibition. | |
| miR-152 [49] | Core | Down | HepG2 | WNT1 increase with a consequent promotion of cell growth and colony formation. | |
| miR-196a [50] | Core Genotype 1b | Up | HepG2, Huh-7 | FOXO1 down-regulation with a consequent proliferation. | |
| miR-203 [51] | Core Genotype 1b | Down | L02 normal human liver, HepG2 | Balb/C nude mice injected with L02 or HepG2 | Induced EMT, increase in cell viability, and a decreased apoptosis susceptibility, possibly by up-regulating SNAL2, the miR-203 target. |
| miR-30c [51] | Core Genotype 1b | Down | L02 normal human liver, HepG2 | Balb/C nude mice injected with L02 or HepG2 | Induced EMT, increase in cell viability, and a decreased apoptosis susceptibility, possibly by up-regulating SNAL1, the miR-30c target |
| miR-138 [52] | Core Genotype 1b | Down | HepG2, Huh-7 | TERT increased expression. Indefinite growth and suppression of the senescence process. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascut, D.; Hoang, M.; Nguyen, N.N.Q.; Pratama, M.Y.; Tiribelli, C. HCV Proteins Modulate the Host Cell miRNA Expression Contributing to Hepatitis C Pathogenesis and Hepatocellular Carcinoma Development. Cancers 2021, 13, 2485. https://doi.org/10.3390/cancers13102485
Pascut D, Hoang M, Nguyen NNQ, Pratama MY, Tiribelli C. HCV Proteins Modulate the Host Cell miRNA Expression Contributing to Hepatitis C Pathogenesis and Hepatocellular Carcinoma Development. Cancers. 2021; 13(10):2485. https://doi.org/10.3390/cancers13102485
Chicago/Turabian StylePascut, Devis, Minh Hoang, Nhu N. Q. Nguyen, Muhammad Yogi Pratama, and Claudio Tiribelli. 2021. "HCV Proteins Modulate the Host Cell miRNA Expression Contributing to Hepatitis C Pathogenesis and Hepatocellular Carcinoma Development" Cancers 13, no. 10: 2485. https://doi.org/10.3390/cancers13102485
APA StylePascut, D., Hoang, M., Nguyen, N. N. Q., Pratama, M. Y., & Tiribelli, C. (2021). HCV Proteins Modulate the Host Cell miRNA Expression Contributing to Hepatitis C Pathogenesis and Hepatocellular Carcinoma Development. Cancers, 13(10), 2485. https://doi.org/10.3390/cancers13102485