
Oncogenic Functions and Clinical Significance of Circular RNAs in Colorectal Cancer

 ,
, 
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
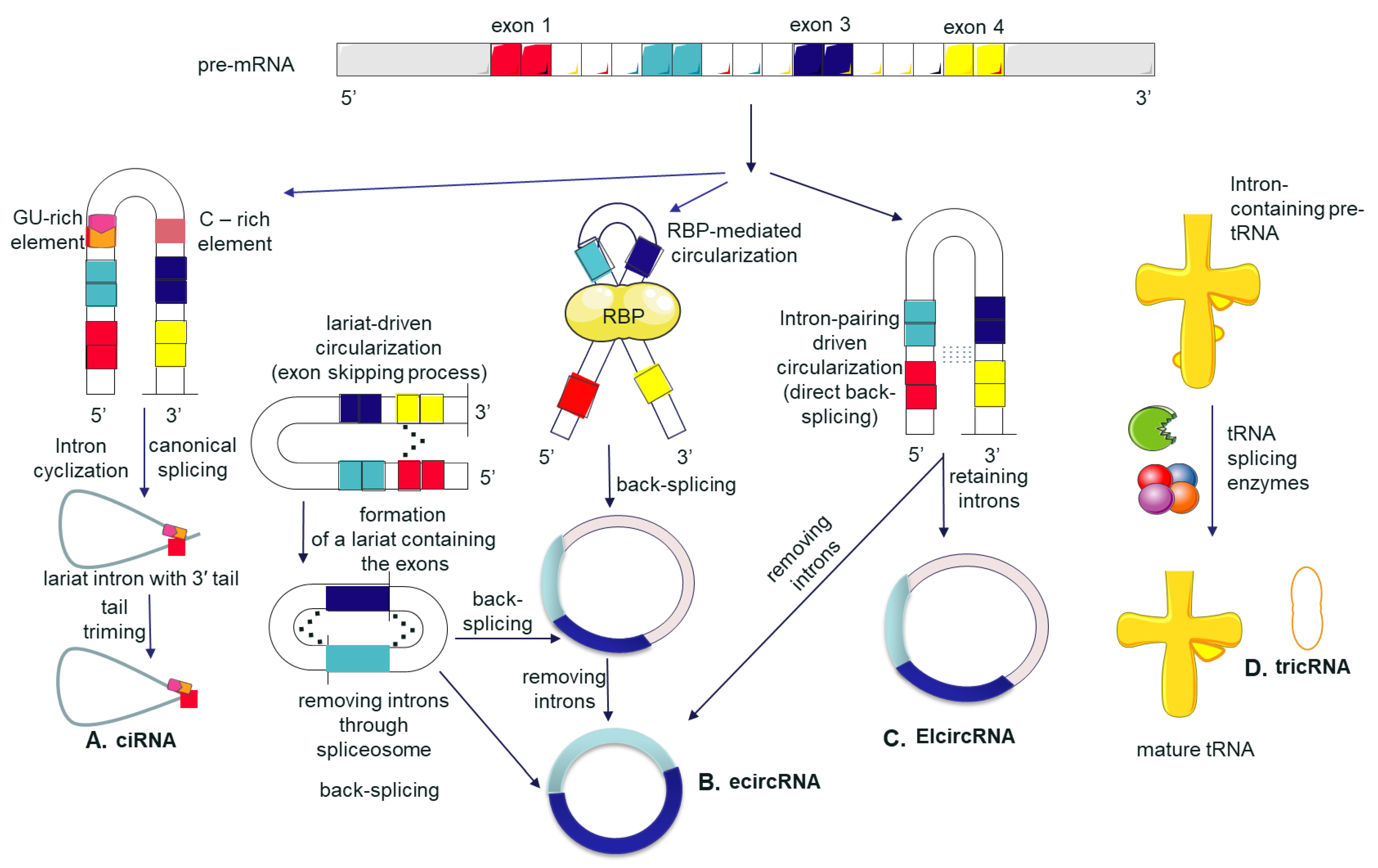
2. Discovery, Biogenesis, and Classification of Circular RNAs
3. CircRNA Resources Onlines

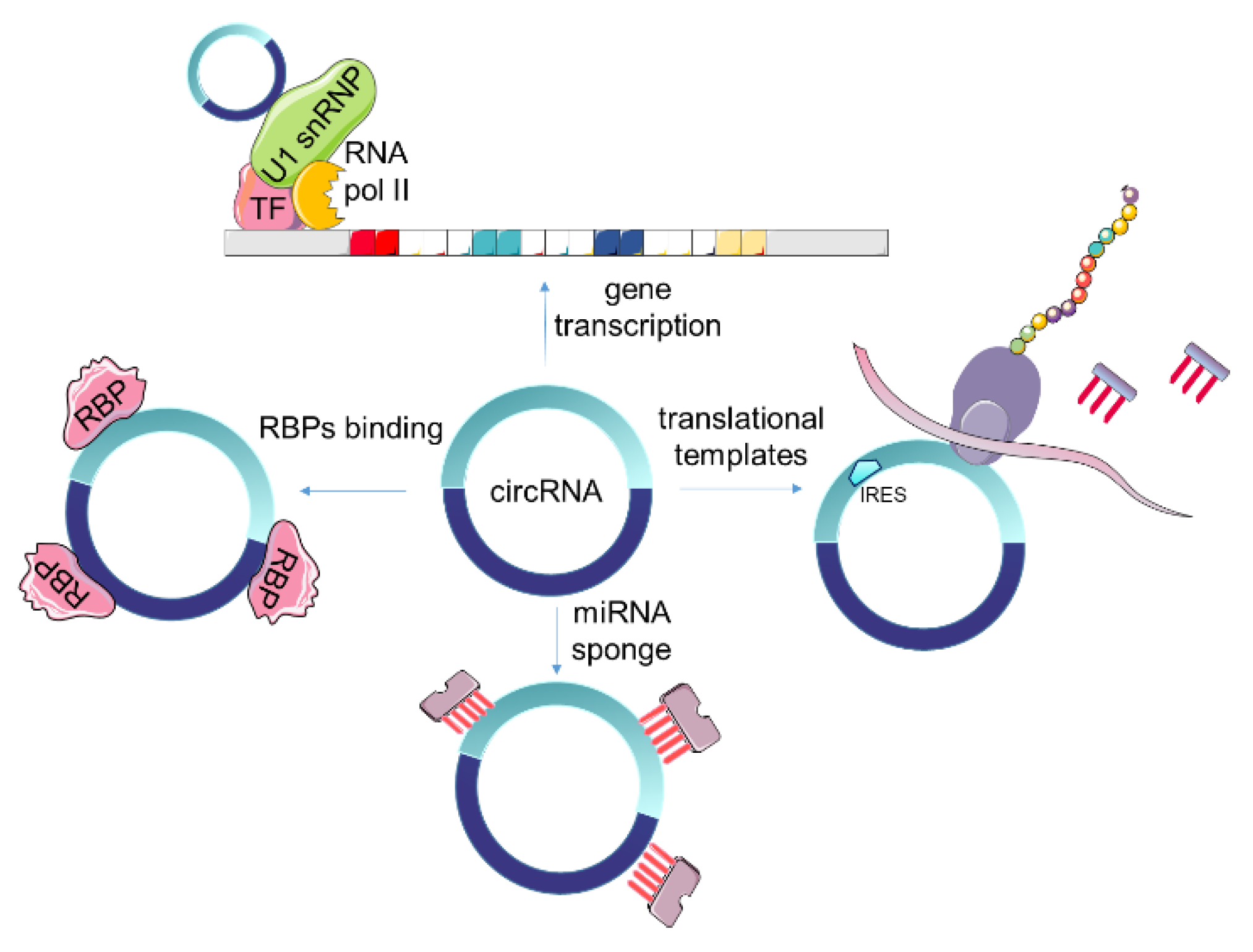
4. Function and Properties of Circular RNAs
4.1. Regulation of Genes Transcription and Alternative Splicing
4.2. Competing Inhibition of miRNAs
4.3. Interaction with RNA Binding Proteins (RBPs)
4.4. Translating Proteins
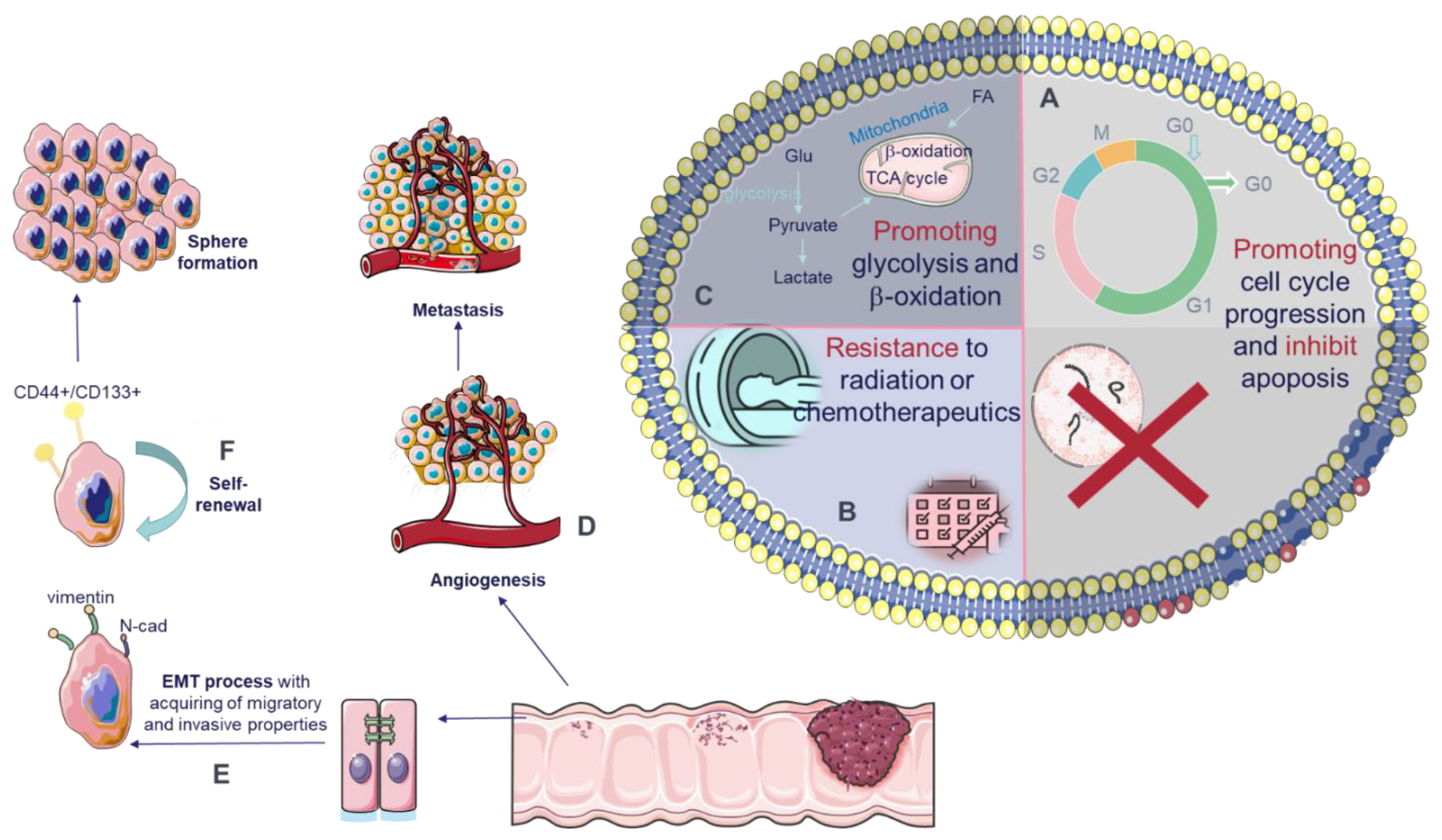
5. Oncogenic Functions of circRNAs
5.1. Impact of circRNAs on Tumor Progression in CRC
5.2. Impact of circRNAs on Invasion/Migration and Metastasis in CRC
5.3. Impact of circRNAs on Cell Cycle and Apoptosis in CRC
5.4. Impact of circRNAs on Angiogenesis in CRC
5.5. Impact of circRNAs on Chemo- and Radioresistance in CRC
5.6. Impact of circRNAs on Anti-Tumor Immunity in CRC
5.7. Impact of circRNAs on Metabolic Pathways in CRC
5.8. CircRNAs with Bioinformatically Predicted Impact on CRC Tumorogenesis
5.9. CircRNAs as Biomarkers in CRC

{kind=link}
{kind=link}
{kind=link}
| CircRNA (Alias) | Gene Symbol | Chromosome: Position | Effects of circRNA Silencing | Possible Mechanism | Association with Clinicopathological Features/biomarker | Reference |
|---|---|---|---|---|---|---|
| circABCC1 (hsa_circ_0000677 hsa_circ_001569) | ABCC1 | chr16:16101672- 16162159 | proliferation (–) invasion (–) | binds with β-catenin, sponges miR-145 | III, IV TNM stages, lymph node, distant metastasis **, poor tumor differentiation | [94,149,253] ** |
| circABCB10 (hsa_circ_0008717) | ABCB10 | chr1:229665945- 229678118 | ferroptosis (+) apoptosis (+) tumor growth (–) | sponges miR-326 | - | [198] |
| circACAP2 (hsa_circ_0007331) | ACAP2 | chr3:195101737- 195112876 | proliferation (–) migration (–) invasion (–) | sponges miR-21-5p | - | [151] |
| circACC1 (hsa_circ_0000759 hsa_circ_001391) | ACACA | chr17:35640167- 35646430 | proliferation (–) | probably via promoting AMPK holoenzyme stability and activation | - | [234] |
| circAGFG1 (hsa_circ_0058514) | AGFG1 | chr2:228356262- 228389631 | proliferation (–) migration (–) invasion (–) stemness (–) apoptosis (+) | sponges miR-4262, miR-185-5p | elevated in CRC patients with liver metastasis | [91] |
| circANKS1B (hsa_circ_0007294) | ANKS1B | chr12:100166699- 100175875 | migration (–) invasion (–) | sponges miR-149 | lymph node metastasis distance metastasis | [143] |
| circAPLP2 (hsa_circ_0000372) | APLP2 | chr11:129979323- 129980556 | proliferation (–) migration (–) invasion (–) | sponges miR-101-3p, miR-495, miR-485-5p | prognostic | [175,176,240] |
| circBANP (hsa_circ_0040824) | BANP | chr16:88061088- 88098938 | proliferation (–) | - | - | [100] |
| circCAMSAP1 (hsa_circ_0001900) | CAMSAP1 | chr9:138773478- 138774924 | proliferation (–) | sponges miR-328-5p | advanced T stage and clinical stage, better prediction performance than CEA and CA19-9 prognostic | [113] |
| circCCDC66 (hsa_circ_0001313 hsa_circ_000374) | CCDC66 | chr3:56626997- 56628056 | proliferation (–) colony formation (–) migration (–) invasion (–) apoptosis (+) radio-sensitivity (+) chemo resistance (–) | sponges miR-33b, miR-93, miR-185, miR-510-5p, miR-338-3p | diagnostic,*** prognostic | [102,210,211,212,253] *** |
| circCCT3 (hsa_circ_0004680) | CCT3 | chr1:156303337- 156304709 | invasion (–) apoptosis (+) | sponges miR-613 | advanced TNM stage prognostic | [202] |
| circCER (hsa_circ_100876 hsa_circ_0023404) | RNF121 | chr11:71668272- 71671937:+ | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-516b | tumor size and differentiation, lymph node and distant metastasis, vascular invasion, clinical T stage, prognostic | [140,256] |
| circCSNK1G1 (hsa_circ_101555 hsa_circ_0001955) | CSNK1G1 | chr15:64495280- 64508912:- | proliferation (–) apoptosis (+) migration (–) invasion (–) | sponges miR-597-5p, miR-455-3p | ≥5cm tumor size, lymph node and distant metastases, III and IV stages, prognostic | [183,184] |
| circCSPP1 (hsa_circ_0001806 hsa_circ_001780) | CSPP1 | chr8:68018139- 68028357 | stemness (–) migration (–) invasion (–) apoptosis (+) chemo sensitivity (+) | sponges miR-193-5p, miR-944 | distant metastasis, prognostic | [153,154,220] |
| circCTNNA1 (hsa_circ_0074169) | CTNNA1 | chr5:138223178- 138260399 | proliferation (–) migration (–) invasion (–) | sponges miR-149-5p, miR-363-3p | advanced TNM stage; prognostic | [147,187] |
| circCTNNB1 (hsa_circ_0123778) | CTNNB1 | chr3:41276543- 41276921 | β-catenin activity (–) | binds the Ia domain of DDX3 protein | - | [90] |
| circDENND4C (hsa_circ_0005684) | DENND4C | chr9:19286766- 19305525 | proliferation (–) migration (–) glycolysis (–) | sponges miR-760 | - | [241] |
| circERBIN (hsa_circ_0001492 hsa_circ_000729) | ERBB2IP | chr5:65284462- 65290692 | proliferation (–) migration (–) invasion (–) | sponges of miR-125a-5p, miR-138-5p | - | [110] |
| circFARSA (hsa_circ_0000896 hsa_circ_000263) | FARSA | chr19:13039155- 13039661 | proliferation (–) migration (–) invasion (–) | sponges miR-330-5p | prognostic | [106] |
| circFAT1 (hsa_circ_0001461 hsa_circ_000713) | FAT1 | chr4:187627716- 187630999 | proliferation (–) glycolysis (–) apoptosis (+) | sponges miR-520b and miR-302c-3p | - | [122] |
| circFMN2 (hsa_circ_0005100) | FMN2 | chr1:240458121- 240497529 | proliferation (–) migration (–) | sponges miR-1182 | histological grade, lymph nodes metastasis and TNM stage, diagnostic, prognostic | [117] |
| circGLIS2 (hsa_circ_101692) | GLIS2 | - | migration (–) leucocyte recruitment (–) | sponges miR-671 | - | [177] |
| circHIPK3 (hsa_circ_0000284 hsa_circ_100782 hsa_circ_000016 circPIK3) | HIPK3 | chr11:33307958- 33309057 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges for miR-7, miR-1207-5p | distant metastasis, TNM stage, liver metastasis, prognostic | [89,150] |
| circHUWE1 (hsa_circ_0140388) | HUWE1 | chrX:53641494- 53644407 | proliferation (–) colony formation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-486 | lymphovascular invasion, lymph node and distant metastasis, advanced TNM stage, prognostic | [96] |
| circIFT80 (hsa_circ_0067835) | IFT80 | chr3:160073800- 160099506 | cell growth (–) proliferation (–) colony formation (–) migration (–) invasion (–) apoptosis (+) radiosensitivity (+) | sponges miR-1236-3p, miR-296-5p | tumor size and advanced stage, prognostic | [115,222] |
| circKRT6C (hsa_circ_0026416) | KRT6C | chr12:52863194- 52865516 | proliferation (–) migration (–) invasion (–) | sponges miR-346 | tumor differentiation, TNM stage distant metastasis, lymphovascular and perineural invasion, diagnostic, prognostic | [144] |
| circLgr4 (hsa_circ_02276) | SPTAN1 | chr9: 131369882- 131375764 | self-renewal (–) invasion (–) | encodes peptide | prognostic | [78] |
| circLMNB1 (hsa_circ_0127801) | LMNB1 | chr5:126153227- 126153886 | proliferation (–) migration (–) invasion (–) apoptosis (+) | upregulates MMP2 and MMP-9 expression | histological grade, lymph node metastasis and TNM stage, diagnostic, prognostic | [174] |
| circLONP2 (hsa_circ_0008558) | LONP2 | chr16:48311248- 48337216 | migration (–) invasion (–) | promotes the processing of primary miR-17 | lymph node and distant metastasis, clinical stages (III+IV), prognostic | [170] |
| circMAT2B (hsa_circ_0128498) | MAT2B | chr5:162939007- 162943717 | proliferation (–) | sponges miR-610 | tumor size, lymph node and distant metastasis, TNM stage | [114] |
| circMBOAT2 (hsa_circ_0007334) | MBOAT2 | chr2:9083315- 9098771 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-519d-3p | TNM stage, distant metastasis, lymphovascular invasion, diagnostic, prognostic | [195] |
| circ-MDM2 (hsa_circ_0027492) | MDM2 | chr12:69210591- 69222711 | p53 levels (+) E2F targets’ levels (–) growth defects (+) G1-S progression (–) | p53 (unknown mechanism) | - | [190] |
| circNOX4 (hsa_circ_0023990) | NOX4 | chr11:89165951- 89185063 | proliferation (–) migration (–) invasion (–) glycolysis (–) | sponges miR-485-5p | tumor size, TNM stage, lymph node and distant metastasis, prognostic | [235] |
| circNRIP1 (hsa_circ_0004771) | NRIP1 | chr21:16386664- 16415895 | - | - | diagnostic | [254] |
| circNSD2 (hsa_circ_0008460) | WHSC1 | chr4:1902352- 1906105 | migration (–) invasion (–) | sponges miR-199b-5p | - | [127] |
| circNSUN2 (circRNA_103783 hsa_circ_0007380) | NSUN2 | chr5: 6623326- 6625782 | migration (–) invasion (–) | interacts with IGF2BP2 and HMGA2 forming a complex | prognostic | [167] |
| circPACRGL (hsa_circ_0069313) | PACRGL | chr4:20702035- 20729980 | proliferation (–) migration (–) invasion (–) | sponges miR-142-3p, miR-506-3p | - | [112] |
| circPIP5K1A (hsa_circ_0014130) | PIP5K1A | chr1:151206672- 151212515 | migration (–) invasion (–) apoptosis (+) | sponges miR-1273a | - | [188] |
| circPNN (hsa_circ_0101802) | PNN | chr14:39648294- 39648666 | - | miR6873-3p, miR-6738-3p, miR-6833-3p, let-7i-3p, miR-1301-3p (in silico prediction) | diagnostic | [246] |
| circPPP1R12A (hsa_circ_0000423 hsa_circ_001676) | PPP1R12A | chr12:80180153- 80183460 | proliferation (–) migration (–) invasion (–) via its encoded product apoptosis (+) | encodes functional peptide, sponges miR-375 | prognostic | [77,92] |
| circPRKDC (hsa_circ_0136666) | PRKDC | chr8:48715866- 48730122 | proliferation (–) migration (–) invasion (–) apoptosis (+) glycolysis (–) colony formation (–) | sponges miR-136, miR-198, miR-375, miR-383 | TNM stage, tumor size, lymph node and distant metastasis, prognostic | [124,125,214,237] |
| circPRMT5 (hsa_circ_0031242) | PRMT5 | chr14:23389732- 23392044 | proliferation (–) | sponges miR-377 | prognostic | [186] |
| circPTK2 (hsa_circ_0005273) | PTK2 | chr8:141710989- 141716304 | proliferation (–) migration (–) invasion (–) | binds to vimentin protein | prognostic | [169] |
| circPVT1 (hsa_circ_0001821 hsa_circ_000006) | TCONS | chr8:128902834- 128903244 | migration (–) invasion (–) | sponges miR-145 | advanced TNM stage, liver metastasis, prognostic | [142] |
| circRAE1 (hsa_circ_0060967) | RAE1 | chr20:55931552- 55943868 | proliferation (–) migration (–) invasion (–) | sponges miR-338-3p | tumor size, advanced tumor stage, lymph node metastasis | [119] |
| circRUNX1 (hsa_circ_0002360) | RUNX1 | chr21:36206706- 36231875 | proliferation (–) migration (–) apoptosis (+) | sponges miR-145-5p | lymph node and distant metastasis | [104] |
| circ_SMAD2 (hsa_circ_0000847 hsa_circ_000640) | SMAD2 | chr18:45391429- 45423180 | tumor growth (–) proliferation (–) invasion (–) | sponges miR-1258 | prognostic | [133] |
| circSMARCC1 (hsa_circ_0003602) | SMARCC1 | chr3:47702783- 47719801 | proliferation (–) migration (–) invasion (–) | sponges miR140-3p | - | [171] |
| circUBAP2 (hsa_circ_0001846 hsa_circ_001335) | UBAP2 | chr9:33944362- 33956144 | proliferation (–) migration (–) invasion (–) | sponges miR-199a | - | [200] |
| circVAPA (hsa_circ_0006990) | VAPA | chr18:9931806- 9937063 | proliferation (–) colony formation (–) migration (–) invasion (–) apoptosis (+) glycolysis (–) | sponges miR-19b-1-5p, miR-132-3p, miR-342-3p, miR-101-3p (in silico prediction) miR-101, miR-125a | lymphovascular invasion, lymph node and distant metastasis, TNM stage, diagnostic | [199,239] |
| circZNF609 (hsa_circ_0000615 hsa_circ_000193) | ZNF609 | chr15:64791491- 64792365 | migration (–) | sponges miR-150 | histological grade, lymph nodes metastasis, TNM stage, diagnostic, prognostic | [165] |
| ciRS-7 (hsa_circ_0001946 hsa_circRNA_105055 CDR1as) | CDR1 | chrX:139865339- 139866824:+ | proliferation (–) migration (–) invasion (–) | sponges miR-7, miR-135a-5p; CMTM4, CMTM6 | advanced TNM stages, lymph node metastasis, low histologic grade, larger tumors size, prognostic | [86,87,88,139,141,228] |
| ciRS-122 (hsa_circ_0005963) | TMEM128 | chr4:4239553- 4248070 | glycolysis (–) drug resistance (–) | sponges miR-122 | - | [208] |
| hsa_circ_0000069 (hsa_circ_001061, circSTIL) | STIL | chr1:47745912- 47748131 | proliferation (–) migration (–) invasion (–) | - | age, II, III TNM stages, dignostic*** | [191,253] *** |
| hsa_circ_0000218 (hsa_circ_001348) | DCLRE1C | chr10:14987103- 15066248 | proliferation (–) migration (–) invasion (–) | sponges miR-139-3p | T staging, local lymph node metastasis | [131] |
| hsa_circ_0000370 (hsa_circ_001553) | FLI1 | chr11:128628009- 128651918 | - | sponges miR-128-3p, miR-502-5p, miR-658 (in silico prediction) | lymph node metastasis, diagnostic | [245] |
| hsa_circ_0000392 (hsa_circ_000139) | YAF2 | chr12:42604156- 42604482 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-193a-5p | TNM stage, lymph node and distant metastasis, diagnostic | [101] |
| hsa_circ_0000504 (hsa_circ_001373) | TUBGCP3 | chr13:113170753- 113181798 | - | sponges miR-92a-3p, miR-485-5p (in silico prediction) | - | [215] |
| hsa_circ_0000511 (hsa_circ_002144) | RPPH1 | chr14:20811282- 20811431 | cell viability (–) proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-615-5p | tumor size, lymph node and distant metastasis, TNM stage, prognostic | [103] |
| hsa_circ_0000512 (hsa_circ_000166) | RPPH1 | chr14:20811282- 20811436 | proliferation (–) apoptosis (+) cell viability (–) colony formation (–) migration (–) invasion (–) | sponges miR-326, miR-330-5p, miR-296-5p | prognostic | [107,146,192] |
| hsa_circ_0000826 (hsa_circ_002032) | ANKRD12 | chr18:9182379- 9221997 | - | interact with RNA binding proteins (in silico prediction), sponges miR-103a-3p, miR-122-5p, miR-1178-3p, miR-1206, miR-107 (in silico prediction) | biomarker for liver metastasis, diagnostic | [162,163] |
| hsa_circ_0001178 (hsa_circ_001637) | USP25 | chr21:17135209- 17138460 | migration (–) invasion (–) | sponges miR-382, miR-587, miR-616 | lymph node and distant metastasis, III and IV stages, biomarker for liver metastasis, diagnostic, prognostic | [161,162] |
| hsa_circ_0004277 | WDR37 | chr10:1125950- 1126416 | proliferation (–) apoptosis (+) | sponges miR-512-5p | tumor size >5cm, III and IV stages, prognostic | [135] |
| hsa_circ_0004585 | KIAA1199 | chr15:81166204- 81212640 | - | sponges more than 20 miRNAs (in silico prediction) | tumor size, diagnostic | [244] |
| hsa_circ_0004831 | RSF1 | chr11:77402203- 77404656 | - | sponges miR-4326 (in silico prediction) | distant metastasis, differentiation grade, prognostic | [247] |
| hsa_circ_0005075 | EIF4G3 | chr1:21377358- 21415706 | proliferation (–) migration (–) invasion (–) | increases expression levels of β-catenin, cyclin D1 and c-myc, thus modulating Wnt/β-catenin pathway | histology differentiation, tumor size, distal metastasis, advanced TNM stage, prognostic | [98,257] |
| hsa_circ_0005615 (circ5615) | NFATC3 | chr16:68155889- 68157024 | proliferation (–) | sponges miR-149-5p | higher T stage, prognostic | [95] |
| hsa_circ_0005927 | VDAC3 | chr8:42259305- 42260979 | - | - | tumor size, diagnostic | [252] |
| hsa_circ_0006174 | RAD23B | chr9:110064315- 110068928 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-138-5p | TNM stage III, lymph node metastasis, prognostic | [155] |
| hsa_circ_0007006 | DYM | chr18:46783379- 46808545 | - | sponges miR-628-5p, miR-653-5p (in silico prediction) | - | [215] |
| hsa_circ_0007031 | TUBGCP3 | chr13:113158345- 113181798 | proliferation (–) apoptosis (+) sensitivity to 5-FU and radiation (+) | sponges miR-760, miR-133b, miR-885-3p, miR-324-5p (in silico prediction) | tumor size, TNM stage, and CEA | [215,216,217] |
| hsa_circ_0007142 | DOCK1 | chr10:128768965- 128788867 | proliferation (–) migration (–) invasion (–) colony formation (–) apoptosis (+) | sponges miR-103a-2-5p, and miR-122-5p, miR-455-5p | poor differentiation, tumor size >5cm, lymph node and distal metastasis, advanced TNM stage | [180,181,182] |
| hsa_circ_0007534 | DDX42 | chr17:61869771- 61877977 | proliferation (–) apoptosis (+) | - | III, IV stages, lymph node and distant metastasis, histological differentiation, diagnostic prognostic | [255] |
| hsa_circ_0007843 | ARHGAP32 | chr11:128993340- 129034322 | proliferation (–) migration (–) invasion (–) colony formation (–) | sponges miR-518c-5p | - | [172] |
| hsa_circ_000984 (hsa_circ_0001724) | CDK6 | chr7:92462409- 92463134 | cell cycle progression (–) proliferation (–) migration (–) invasion (–) | sponges miR-106b | III and IV stages | [185] |
| hsa_circ_0010522 (circ-133) | RAP1GAP | chr1:21939668- 21940582 | metastasis (–) | sponges miR133a | - | [164] |
| hsa_circ_0011385 (circ_100146) | EIF3I | chr1:32691771- 32692131 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-149 | - | [166] |
| hsa_circ_001680 (hsa_circ_0000598) | B2M | chr15:45009906- 45009989 | proliferation (–) migration (–) | sponges miR-340 | - | [221] |
| hsa_circ_001971 (hsa_circ_0001060) | UXS1 | chr2:106774513- 106782539 | proliferation (–) invasion (–) | sponges miR-29c-3p | TNM stage, diagnostic, prognostic | [201] |
| hsa_circ_0020095 | ATRNL1 | chr10:116975454- 117075246 | proliferation (–) migration (–) invasion (–) cisplatin resistance (–) | sponges miR-487a-3p | - | [157] |
| hsa_circ_0020397 | DOCK1 | chr10:128768965- 128926028 | cell viability (–) invasion (–) apoptosis (+) | inhibits miR-138 via upregulation of its targets | - | [226] |
| hsa_circ_0032833 | STON2 | chr14:81837331- 81864924 | colony formation (–) migration (–) invasion (–) apoptosis (+) 5-FU, oxaliplatin sensitivity (+) | sponges miR-125-5p | - | [206] |
| hsa_circ_0038646 | PRKCB | chr16:23999828- 24166178 | proliferation (–) migration (–) | sponges miR-331-3p | III and IV stages | [123] |
| hsa_circ_0053277 | NRBP1 | chr2:27663993- 27665124 | proliferation (–) migration (–) epithelial–mesenchymal transition (EMT) (–) | sponges miR-2467-3p | - | [173] |
| hsa_circ_0055625 | DUSP2 | chr2:96808907- 96810112 | proliferation (–) migration (–) invasion (–) | sponges miR-106b-5p | larger tumor size, TNM stage, histological differentiation, and lymph node metastasis | [193] |
| hsa_circ_0056618 | SPOPL | chr2:139259349- 139307864 | proliferation (–) migration (–) angiogenesis (–) | sponges miR-206 | tumor size, III and IV stage, lymph node and distant metastasis, prognostic | [203] |
| hsa_circ_0071589 | FAT1 | chr4:187517693- 187518946 | proliferation (–) migration (–) invasion (–) | sponges miR-600 | grade III and IV, lymph node metastasis and advanced clinical stage, prognostic | [121] |
| hsa_circ_0079993 | POLR2J4 | chr7:44043644- 44044934 | proliferation (–) | sponges miR-203a-3p.1 | III and IV stages, metastasis, prognostic | [238] |
| hsa_circ_0082182 | FAM71F2 | chr7:128317617- 128323309 | - | sponges miR-767-3p, miR-609, miR-2682-5p (in silico prediction) | lymph node metastasis | [245] |
| hsa_circ_0104631 | IREB2 | chr15:78757592- 78778182 | proliferation (–) invasion (–) | inhibits PTEN expression and promotes the AKT/mTOR pathway | advanced TNM stage, lymph node and distant metastasis, prognostic | [99] |
| hsa_circ_0115744 | TCONS | chr21:29818610- 29899681 | invasion (–) | sponges miR144 | liver metastasis, diagnostic | [120] |
| hsa_circ_0128846 | ZFR | chr5:32379220- 32420208 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-1184 | - | [105] |
| hsa_circ_100290 (has_circ_0013339) | SLC30A7 | chr1:101372407- 101379362 | proliferation (–) migration (–) invasion (–) apoptosis (+) | sponges miR-516b | observed in metastatic patients, prognostic | [93] |
| hsa_circ_100859 (hsa_circ_0023064) | KDM2A | chr11:66985201- 66986874 | proliferation (–) apoptosis (+) | sponges miR-217 | TNM stage, histological grade, KRAS mutations, diagnostic, prognostic | [111] |
| hsa_circ_101951 | - | - | proliferation (–) colony formation (–) migration (–) invasion (–) apoptosis (+) | activates the KIF3A-mediated EMT signaling pathway | tumor size, TNM stage, metastasis, prognostic | [168] |
| hsa_circ_102209 (has_circ_0045890) | PS1 | chr17: 76388556- 76411108 | proliferation (–) migration (–) invasion (–) EMT (–) apoptosis (+) | sponges miR-761 | histology grade III and IV, liver metastasis, prognostic | [129] |
| hsa_circ_102958 (hsa_circ_0003854) | PER2 | chr2: 239184383- 239186596 | proliferation (–) migration (–) invasion (–) | sponges miR-585 | III/IV clinical stages, lymph node metastasis, prognostic | [179] |
| hsa_circ_32883 | EML5 | - | - | sponges miR-130b-5p, miR-367-5p, miR128-3p, miR-501-5p, miR-381-3p (in silico prediction) | - | [218] |
| CircRNA (Alias) | Gene Symbol | Chromosome: | Biomarker | Reference |
|---|---|---|---|---|
| Position | ||||
| hsa_circ_0082182, | FAM71F2 | chr7:128317617-128323309 | diagnostic | [245] |
| hsa_circ_0000370, | FLI1 | chr11:128628009-128651918 | ||
| hsa_circ_0035445 | ALDH1A2 | chr15:58302846-58306479 | ||
| hsa_circ_0122319, | PLOD2 | chr3:145838898-145842016 | prognostic | [251] |
| hsa_circ_0079480, | CRPPA | chr7:16298014-16317851 | ||
| hsa_circ_0087391 | AGTPBP1 | chr9:88284399-88327481 | ||
| hsa_circ_0001900, | CAMSAP1 | chr9:138773478-138774924 | diagnostic | [252] |
| hsa_circ_0001178, | USP25 | chr21:17135209-17138460 | ||
| hsa_circ_0005927 | VDAC3 | chr8:42259305-42260979 | ||
| circCCDC66, | CCDC66 | chr3:56626997-56628056 | diagnostic | [253] |
| circABCC1, | ABCC1 | chr16:16101672-16162159 | ||
| circSTIL | STIL | chr1:47745912-47748131 |
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kuiper, R.P.; Weren, R.D.; van Kessel, A.G. Colorectal Cancer. In Genomic and Precision Medicine; Elsevier: Amsterdam, The Netherlands, 2017; pp. 195–209. [Google Scholar]
- De Falco, V.; Napolitano, S.; Roselló, S.; Huerta, M.; Cervantes, A.; Ciardiello, F.; Troiani, T. How we treat metastatic colorectal cancer. ESMO Open 2019, 4, e000813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, W.; Zhou, Q.; Chen, C.; Yuan, W.; Liu, J.; Li, X.; Sun, Z. Roles of circRNAs in the tumour microenvironment. Mol. Cancer 2020, 19, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artemaki, P.I.; Scorilas, A.; Kontos, C.K. Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers 2020, 12, 2464. [Google Scholar] [CrossRef]
- Suzuki, H.; Tsukahara, T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int. J. Mol. Sci. 2014, 15, 9331–9342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Zhang, Y.; Ke, X.; Liu, J.; Ma, X.; Liu, Y.; Liang, D.; Wang, L.; Guo, C.; Luo, Y. Characterization of circRNA-associated ceRNA networks in patients with nonvalvular persistent atrial fibrillation. Mol. Med. Rep. 2019, 19, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E. Translation of circRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Nie, H.; Wang, Y.; Liao, Z.; Zhou, J.; Ou, C. The function and mechanism of circular RNAs in gastrointestinal tumours. Cell Prolif. 2020, 53, e12815. [Google Scholar] [CrossRef]
- Naeli, P.; Pourhanifeh, M.H.; Karimzadeh, M.R.; Shabaninejad, Z.; Movahedpour, A.; Tarrahimofrad, H.; Mirzaei, H.R.; Bafrani, H.H.; Savardashtaki, A.; Mirzaei, H. Circular RNAs and gastrointestinal cancers: Epigenetic regulators with a prognostic and therapeutic role. Crit. Rev. Oncol. Hematol. 2020, 145, 102854. [Google Scholar] [CrossRef]
- Long, F.; Lin, Z.; Li, L.; Ma, M.; Lu, Z.; Jing, L.; Li, X.; Lin, C. Comprehensive landscape and future perspectives of circular RNAs in colorectal cancer. Mol. Cancer 2021, 20, 26. [Google Scholar] [CrossRef]
- Zeng, K.; Wang, S. Circular RNAs: The crucial regulatory molecules in colorectal cancer. Pathol.-Res. Pract. 2020, 216, 152861. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; Xu, S.; Guo, J. Novel potential tumor biomarkers: Circular RNAs and exosomal circular RNAs in gastrointestinal malignancies. J. Clin. Lab. Anal. 2020, 34, e23359. [Google Scholar] [CrossRef]
- Wang, H.Y.; Wang, Y.P.; Zeng, X.; Zheng, Y.; Guo, Q.H.; Ji, R.; Zhou, Y.N. Circular RNA is a popular molecule in tumors of the digestive system. Int. J. Oncol. 2020, 57, 21–42. [Google Scholar] [CrossRef]
- Li, A.; Wang, W.C.; McAlister, V.; Zhou, Q.; Zheng, X. Circular RNA in colorectal cancer. J. Cell. Mol. Med. 2021. [Google Scholar] [CrossRef]
- Hao, S.; Cong, L.; Qu, R.; Liu, R.; Zhang, G.; Li, Y. Emerging roles of circular RNAs in colorectal cancer. OncoTargets Ther. 2019, 12, 4765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopczynski, C.C.; Muskavitch, M. Introns excised from the Delta primary transcript are localized near sites of Delta transcription. J. Cell Biol. 1992, 119, 503–512. [Google Scholar] [CrossRef]
- Qian, L.; Vu, M.N.; Carter, M.; Wilkinson, M.F. A spliced intron accumulates as a lariat in the nucleus of T cells. Nucleic Acids Res. 1992, 20, 5345–5350. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Papavasileiou, P.; Peters, O.; Rajewsky, N. Identification and characterization of circular RNAs as a new class of putative biomarkers in human blood. PLoS ONE 2015, 10, e0141214. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, F.; Wu, W.; Zhang, Y.; Chang, W.; Ponnusamy, M.; Wang, K.; Li, P. Circular RNAs: A novel type of non-coding RNA and their potential implications in antiviral immunity. Int. J. Biol. Sci. 2017, 13, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Wang, J.; Guo, Z.; Li, M.; Li, M.; Liu, S.; Liu, H.; Li, W.; Yin, X.; Tao, J. Emerging function and potential diagnostic value of circular RNAs in cancer. Mol. Cancer 2018, 17, 123. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.; Greenman, C.; Cook, P.R.; Papantonis, A. Exon skipping is correlated with exon circularization. J. Mol. Biol. 2015, 427, 2414–2417. [Google Scholar] [CrossRef]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.-H.; Bindereif, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Wilusz, J.E. Circular RNAs: Unexpected outputs of many protein-coding genes. RNA Biol. 2017, 14, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Aucamp, J.; Bronkhorst, A.J.; Pretorius, P.J. A historical and evolutionary perspective on circulating nucleic acids and extracellular vesicles: Circulating nucleic acids as homeostatic genetic entities. Adv. Exp. Med. Biol. 2016. [Google Scholar] [CrossRef]
- Monat, C.; Quiroga, C.; Laroche-Johnston, F.; Cousineau, B. The Ll. LtrB intron from Lactococcus lactis excises as circles in vivo: Insights into the group II intron circularization pathway. RNA 2015, 21, 1286–1293. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Huang, G.; Li, S.; Yang, N.; Zou, Y.; Zheng, D.; Xiao, T. Recent progress in circular RNAs in human cancers. Cancer Lett. 2017, 404, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, mechanism, and function in human cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Sun, H.; Wang, H.; Jiang, W.; Tang, W.; Lu, C.; Zhang, W.; Chen, Z.; Lv, C. Star circular RNAs in human cancer: Progress and perspectives. OncoTargets Ther. 2019, 12, 8249–8261. [Google Scholar] [CrossRef] [Green Version]
- Salgia, S.R.; Singh, S.K.; Gurha, P.; Gupta, R. Two reactions of Haloferax volcanii RNA splicing enzymes: Joining of exons and circularization of introns. RNA 2003, 9, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Bogard, B.; Francastel, C.; Hubé, F. A new method for the identification of thousands of circular RNAs. Non-Coding RNA Investig. 2018, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Qu, S.; Zhong, Y.; Shang, R.; Zhang, X.; Song, W.; Kjems, J.; Li, H. The emerging landscape of circular RNA in life processes. RNA Biol. 2017, 14, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.-Z.; Guo, C.; Zou, M.-M.; Meng, P.; Zhang, T.-T. circRNA-miRNA-mRNA regulatory network in human lung cancer: An update. Cancer Cell Int. 2020, 20, 173. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.-M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X. The landscape of circular RNA in cancer. Cell 2019, 176, 869–881.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocheleau, L.; Pelchat, M. The Subviral RNA Database: A toolbox for viroids, the hepatitis delta virus and satellite RNAs research. BMC Microbiol. 2006, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.P.; Neeno-Eckwall, E.C.; Adhikari, B.N.; Perna, N.T.; Tisserat, N.; Leach, J.E.; Levesque, C.A.; Buell, C.R. The Comprehensive Phytopathogen Genomics Resource: A web-based resource for data-mining plant pathogen genomes. Database 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, S.; Das, S.; Sen, R.; Basak, P.; Chakrabarti, J. Circ2Traits: A comprehensive database for circular RNA potentially associated with disease and traits. Front. Genet. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-C.; Li, J.-R.; Sun, C.-H.; Andrews, E.; Chao, R.-F.; Lin, F.-M.; Weng, S.-L.; Hsu, S.-D.; Huang, C.-C.; Cheng, C. CircNet: A database of circular RNAs derived from transcriptome sequencing data. Nucleic Acids Res. 2016, 44, D209–D215. [Google Scholar] [CrossRef] [Green Version]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Han, P.; Zhou, T.; Guo, X.; Song, X.; Li, Y. circRNADb: A comprehensive database for human circular RNAs with protein-coding annotations. Sci. Rep. 2016, 6, 34985. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, S.P.; Wang, P.L.; Salzman, J. Circular RNA biogenesis can proceed through an exon-containing lariat precursor. eLife 2015, 4, e07540. [Google Scholar] [CrossRef]
- Schindewolf, C.; Braun, S.; Domdey, H. In vitro generation of a circular exon from a linear pre-mRNA transcript. Nucleic Acids Res. 1996, 24, 1260–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, X.-O.; Chen, T.; Xiang, J.-F.; Yin, Q.-F.; Xing, Y.-H.; Zhu, S.; Yang, L.; Chen, L.-L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumtaz, P.T.; Taban, Q.; Dar, M.A.; Mir, S.; Haq, Z.u.; Zargar, S.M.; Shah, R.A.; Ahmad, S.M. Deep Insights in Circular RNAs: From biogenesis to therapeutics. Biol. Proced. Online 2020, 22, 10. [Google Scholar] [CrossRef]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R. Circular RNAs in the mammalian brain are highly abundant, conserved, and dynamically expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-O.; Wang, H.-B.; Zhang, Y.; Lu, X.; Chen, L.-L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Hayne, C.K.; Schmidt, C.A.; Haque, M.I.; Matera, A.G.; Stanley, R.E. Reconstitution of the human tRNA splicing endonuclease complex: Insight into the regulation of pre-tRNA cleavage. Nucleic Acids Res. 2020, 48, 7609–7622. [Google Scholar] [CrossRef]
- Schmidt, C.A.; Giusto, J.D.; Bao, A.; Hopper, A.K.; Matera, A.G. Molecular determinants of metazoan tricRNA biogenesis. Nucleic Acids Res. 2019, 47, 6452–6465. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, T.; Wang, X.; He, A. Circles reshaping the RNA world: From waste to treasure. Mol. Cancer 2017, 16, 58. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Yuan, Z.; Du, K.Y.; Fang, L.; Lyu, J.; Zhang, C.; He, A.; Eshaghi, E.; Zeng, K.; Ma, J. Translation of yes-associated protein (YAP) was antagonized by its circular RNA via suppressing the assembly of the translation initiation machinery. Cell Death Differ. 2019, 26, 2758–2773. [Google Scholar] [CrossRef] [Green Version]
- Conn, V.M.; Hugouvieux, V.; Nayak, A.; Conos, S.A.; Capovilla, G.; Cildir, G.; Jourdain, A.; Tergaonkar, V.; Schmid, M.; Zubieta, C. A circRNA from SEPALLATA3 regulates splicing of its cognate mRNA through R-loop formation. Nat. Plants 2017, 3, 17053. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.O.; Mikkelsen, J.G. miRNA sponges: Soaking up miRNAs for regulation of gene expression. Wiley Interdiscip. Rev. RNA 2014, 5, 317–333. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Yan, M.; Li, H. Emerging role of circular RNAs in cancer. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA expression: Its potential regulation and function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, L.; Chen, L.-L. The biogenesis, functions, and challenges of circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [Green Version]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Yang, Q.; He, A.T.; Yang, B.B. Circular RNAs in cancer: Limitations in functional studies and diagnostic potential. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Karousi, P.; Artemaki, P.I.; Sotiropoulou, C.D.; Christodoulou, S.; Scorilas, A.; Kontos, C.K. Identification of Two Novel Circular RNAs Deriving from BCL2L12 and Investigation of Their Potential Value as a Molecular Signature in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 8867. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.-L.; Wang, Y. Extensive translation of circular RNAs driven by N 6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudi, E.; Kiltschewskij, D.; Fitzsimmons, C.; Cairns, M.J. Depolarization-associated CircRNA regulate neural gene expression and in some cases may function as templates for translation. Cells 2020, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, Y.; Wang, C.; Zhang, H.; Zhang, H.; Jiang, B.; Guo, X.; Song, X. IRESbase: A comprehensive database of experimentally validated internal ribosome entry sites. Genom. Proteom. Bioinform. 2020, 18, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Z. IRES-mediated cap-independent translation, a path leading to hidden proteome. J. Mol. Cell Biol. 2019, 11, 911–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Chen, L.; Zhou, Y.; Wang, Q.; Zheng, Z.; Xu, B.; Wu, C.; Zhou, Q.; Hu, W.; Wu, C. A novel protein encoded by a circular RNA circPPP1R12A promotes tumor pathogenesis and metastasis of colon cancer via Hippo-YAP signaling. Mol. Cancer 2019, 18, 47. [Google Scholar] [CrossRef] [Green Version]
- Zhi, X.; Zhang, J.; Cheng, Z.; Bian, L.; Qin, J. circLgr4 drives colorectal tumorigenesis and invasion through Lgr4-targeting peptide. Int. J. Cancer 2019. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.A. Dynamics of Cancer: Incidence, Inheritance, and Evolution; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Sun, S.; Klebaner, F.; Tian, T. A new model of time scheme for progression of colorectal cancer. BMC Syst. Biol. 2014, 8, S2. [Google Scholar] [CrossRef] [Green Version]
- European Society for Medical Oncology. Essentials for Clinicians: Gastrointestinal Tract Tumours, 2nd ed.; Rickmansworth: Hertfordshire, UK, 2021. [Google Scholar]
- La Vecchia, S.; Sebastián, C. Metabolic pathways regulating colorectal cancer initiation and progression. Semin. Cell Dev. Biol. 2020. [Google Scholar] [CrossRef]
- Koveitypour, Z.; Panahi, F.; Vakilian, M.; Peymani, M.; Forootan, F.S.; Esfahani, M.H.N.; Ghaedi, K. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Wan, M.-l.; Wang, Y.; Zeng, Z.; Deng, B.; Zhu, B.-s.; Cao, T.; Li, Y.-k.; Xiao, J.; Han, Q.; Wu, Q. Colorectal cancer (CRC) as a multifactorial disease and its causal correlations with multiple signaling pathways. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yang, J.; Fei, X.; Wang, X.; Wang, K. CircRNA ciRS-7: A Novel Oncogene in Multiple Cancers. Int. J. Biol. Sci. 2021, 17, 379–389. [Google Scholar] [CrossRef]
- Tang, W.; Ji, M.; He, G.; Yang, L.; Niu, Z.; Jian, M.; Wei, Y.; Ren, L.; Xu, J. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. OncoTargets Ther. 2017, 10, 2045–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, W.; Wei, Q.; Toden, S.; Yoshida, K.; Nagasaka, T.; Fujiwara, T.; Cai, S.; Qin, H.; Ma, Y.; Goel, A. Circular RNA ciRS-7—A promising prognostic biomarker and a potential therapeutic target in colorectal cancer. Clin. Cancer Res. 2017, 23, 3918–3928. [Google Scholar] [CrossRef] [Green Version]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Yang, F.; Fang, E.; Mei, H.; Chen, Y.; Li, H.; Li, D.; Song, H.; Wang, J.; Hong, M.; Xiao, W. Cis-acting circ-CTNNB1 promotes β-catenin signaling and cancer progression via DDX3-mediated transactivation of YY1. Cancer Res. 2019, 79, 557–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Dong, X.; Yan, B.; Yu, W.; Shan, L. CircAGFG1 drives metastasis and stemness in colorectal cancer by modulating YY1/CTNNB1. Cell Death Dis. 2020, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Tian, Z.; Zhang, L. CircPPP1R12A promotes the progression of colon cancer through regulating CTNNB1 via sponging miR-375. Anti-Cancer Drugs 2021. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Ye, B.-L.; Hu, B.-R.; Ruan, X.-J.; Shi, Y.-X. CircRNA_100290 promotes colorectal cancer progression through miR-516b-induced downregulation of FZD4 expression and Wnt/β-catenin signaling. Biochem. Biophys. Res. Commun. 2018, 504, 184–189. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, S.; Fu, Q. Exosomes from CD133+ cells carrying circ-ABCC1 mediate cell stemness and metastasis in colorectal cancer. J. Cell. Biochem. 2020, 121, 3286–3297. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Han, C.; Xia, W.; Li, X.; Fang, P.; Yin, R.; Xu, L.; Yang, L. circ5615 functions as a ceRNA to promote colorectal cancer progression by upregulating TNKS. Cell Death Dis. 2020, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-Y.; Li, X.-N.; Ye, C.-X.; Chen, Z.-L.; Wang, Z.-J. Circular RNA circHUWE1 is upregulated and promotes cell proliferation, migration and invasion in colorectal cancer by sponging miR-486. OncoTargets Ther. 2020, 13, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Chen, X.; Zeng, K.; Xu, M.; He, B.; Pan, Y.; Sun, H.; Pan, B.; Xu, X.; Xu, T. DNA-methylation-mediated silencing of miR-486-5p promotes colorectal cancer proliferation and migration through activation of PLAGL2/IGF2/β-catenin signal pathways. Cell Death Dis. 2018, 9, 1037. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Ren, Y.; Gao, Y.; Zhang, L.; Ding, Z. Hsa_circ_0005075 predicts a poor prognosis and acts as an oncogene in colorectal cancer via activating Wnt/β-catenin pathway. Eur. Rev. Med. Pharm. Sci. 2019, 23, 3311–3319. [Google Scholar] [CrossRef]
- Cao, J.; Ma, L.; Zhang, Y.; Guo, H.; Niu, X.; Zhao, T. Circ-0104631 promotes cell proliferation and invasion in colorectal cancer and predicts poor prognosis. Eur. Rev. Med. Pharm. Sci. 2019, 23, 4730–4737. [Google Scholar] [CrossRef]
- Zhu, M.; Xu, Y.; Chen, Y.; Yan, F. Circular BANP, an upregulated circular RNA that modulates cell proliferation in colorectal cancer. Biomed. Pharmacother. 2017, 88, 138–144. [Google Scholar] [CrossRef]
- Xu, H.; Liu, Y.; Cheng, P.; Wang, C.; Liu, Y.; Zhou, W.; Xu, Y.; Ji, G. CircRNA_0000392 promotes colorectal cancer progression through the miR-193a-5p/PIK3R3/AKT axis. J. Exp. Clin. Cancer Res. 2020, 39, 283. [Google Scholar] [CrossRef]
- Tu, F.-L.; Guo, X.-Q.; Wu, H.-X.; He, Z.-Y.; Wang, F.; Sun, A.-J.; Dai, X.-D. Circ-0001313/miRNA-510-5p/AKT2 axis promotes the development and progression of colon cancer. Am. J. Transl. Res. 2020, 12, 281–291. [Google Scholar]
- Wu, M.; Kong, C.; Cai, M.; Huang, W.; Chen, Y.; Wang, B.; Liu, X. Hsa_circRNA_002144 promotes growth and metastasis of colorectal cancer through regulating miR-615-5p/LARP1/mTOR pathway. Carcinogenesis 2021, 42, 601–610. [Google Scholar] [CrossRef]
- Chen, Z.-L.; Li, X.-N.; Ye, C.-X.; Chen, H.-Y.; Wang, Z.-J. Elevated levels of circRUNX1 in colorectal cancer promote cell growth and metastasis via miR-145-5p/IGF1 signalling. OncoTargets Ther. 2020, 13, 4035–4048. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Liu, W.; Liu, T.; Sun, D. Hsa_circ_0128846 promotes tumorigenesis of colorectal cancer by sponging hsa-miR-1184 and releasing AJUBA and inactivating Hippo/YAP signalling. J. Cell. Mol. Med. 2020, 24, 9908–9924. [Google Scholar] [CrossRef]
- Lu, C.; Fu, L.; Qian, X.; Dou, L.; Cang, S. Knockdown of circular RNA circ-FARSA restricts colorectal cancer cell growth through regulation of miR-330-5p/LASP1 axis. Arch. Biochem. Biophys. 2020, 689, 108434. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Zhang, Z. Hsa_circRNA_000166 facilitated cell growth and limited apoptosis through targeting miR-326/LASP1 axis in colorectal cancer. Gastroenterol. Res. Pract. 2020, 2020. [Google Scholar] [CrossRef]
- Chen, N.; Han, X.; Bai, X.; Yin, B.; Wang, Y. LASP1 induces colorectal cancer proliferation and invasiveness through Hippo signaling and Nanog mediated EMT. Am. J. Transl. Res. 2020, 12, 6490–6500. [Google Scholar] [PubMed]
- Zhou, R.; Shao, Z.; Liu, J.; Zhan, W.; Gao, Q.; Pan, Z.; Wu, L.; Xu, L.; Ding, Y.; Zhao, L. COPS5 and LASP1 synergistically interact to downregulate 14-3-3σ expression and promote colorectal cancer progression via activating PI3K/AKT pathway. Int. J. Cancer 2018, 142, 1853–1864. [Google Scholar] [CrossRef]
- Chen, L.-Y.; Wang, L.; Ren, Y.-X.; Pang, Z.; Liu, Y.; Sun, X.-D.; Tu, J.; Zhi, Z.; Qin, Y.; Sun, L.-N. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1α translation. Mol. Cancer 2020, 19, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; Huang, H.-L.; Huang, R.-Q.; Tian, C.; Zhu, H.-B.; Dai, Y.-H.; Li, Z.-Y. circRNA_100859 functions as an oncogene in colon cancer by sponging the miR-217-HIF-1α pathway. Aging 2020, 12, 13338–13353. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Gu, C.; Wang, W.; Wang, X.; Sun, J.; Zeng, B.; Chen, C.; Chang, W.; Ping, Y.; Ji, P. Exosomal circPACRGL promotes progression of colorectal cancer via the miR-142-3p/miR-506-3p-TGF-β1 axis. Mol. Cancer 2020, 19, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, H.-S.; Wang, F.-W.; Hu, T.; Liang, Z.-X.; Lan, N.; He, X.-W.; Zheng, X.-B.; Wu, X.-J.; Xie, D. circCAMSAP1 promotes tumor growth in colorectal cancer via the miR-328-5p/E2F1 axis. Mol. Ther. 2020, 28, 914–928. [Google Scholar] [CrossRef]
- Zhao, J.P.; Chen, L.L. Circular RNA MAT2B Induces Colorectal Cancer Proliferation via Sponging miR-610, Resulting in an Increased E2F1 Expression. Cancer Manag. Res. 2020, 12, 7107–7116. [Google Scholar] [CrossRef]
- Feng, W.; Gong, H.; Wang, Y.; Zhu, G.; Xue, T.; Wang, Y.; Cui, G. circIFT80 functions as a ceRNA of miR-1236-3p to promote colorectal cancer progression. Mol. Ther.-Nucleic Acids 2019, 18, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.-T.; Jiang, D.; Yuan, J.; Cui, Y.-M.; Shi, X.-W.; Chen, C.-M.; Bian, X.-W.; Deng, Y.-J.; Ding, Y.-Q. HOXB7 as a prognostic factor and mediator of colorectal cancer progression. Clin. Cancer Res. 2011, 17, 3569–3578. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, C.; Xu, R.; Wang, Y.; Li, D.; Zhang, B. A novel circFMN2 promotes tumor proliferation in CRC by regulating the miR-1182/hTERT signaling pathways. Clin. Sci. 2019, 133, 2463–2479. [Google Scholar] [CrossRef] [Green Version]
- Bertorelle, R.; Rampazzo, E.; Pucciarelli, S.; Nitti, D.; De Rossi, A. Telomeres, telomerase and colorectal cancer. World J. Gastroenterol. 2014, 20, 1940–1950. [Google Scholar] [CrossRef]
- Du, J.; Xu, J.; Chen, J.; Liu, W.; Wang, P.; Ye, K. circRAE1 promotes colorectal cancer cell migration and invasion by modulating miR-338-3p/TYRO3 axis. Cancer Cell Int. 2020, 20, 430. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Lv, L.; Xing, C. Circ_ 0115744 acts as miR-144 sponge to promote and predict the metastasis of colorectal cancer. Aging 2021, 13, 5892–5905. [Google Scholar] [CrossRef]
- Yong, W.; Zhuoqi, X.; Baocheng, W.; Dongsheng, Z.; Chuan, Z.; Yueming, S. Hsa_circ_0071589 promotes carcinogenesis via the miR-600/EZH2 axis in colorectal cancer. Biomed. Pharmacother. 2018, 102, 1188–1194. [Google Scholar] [CrossRef]
- Hu, B.; Xian, Z.; Zou, Q.; Zhang, D.; Su, D.; Yao, J.; Ren, D. CircFAT1 suppresses colorectal cancer development through regulating miR-520b/UHRF1 Axis or miR-302c-3p/UHRF1 Axis. Cancer Biother. Radiopharm. 2021, 36, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; He, Z.; Feng, F.; Chen, D.; Zhang, L.; Bai, J.; Wu, H.; Han, E.; Zhang, J. Hsa_circ_0038646 promotes cell proliferation and migration in colorectal cancer via miR-331-3p/GRIK3. Oncol. Lett. 2020, 20, 266–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.; Wang, A.; Liu, L.; Wang, G.; Li, G. Hsa_circ_0136666 promotes the proliferation and invasion of colorectal cancer through miR-136/SH2B1 axis. J. Cell. Physiol. 2019, 234, 7247–7256. [Google Scholar] [CrossRef]
- Wang, G.; Li, Y.; Zhu, H.; Huo, G.; Bai, J.; Gao, Z. Circ-PRKDC Facilitates the Progression of Colorectal Cancer Through miR-198/DDR1 Regulatory Axis. Cancer Manag. Res. 2020, 12, 12853–12865. [Google Scholar] [CrossRef]
- Borza, C.M.; Pozzi, A. Discoidin domain receptors in disease. Matrix Biol. 2014, 34, 185–192. [Google Scholar] [CrossRef]
- Chen, L.Y.; Zhi, Z.; Wang, L.; Zhao, Y.Y.; Deng, M.; Liu, Y.H.; Qin, Y.; Tian, M.M.; Liu, Y.; Shen, T. NSD2 circular RNA promotes metastasis of colorectal cancer by targeting miR-199b-5p-mediated DDR1 and JAG1 signalling. J. Pathol. 2019, 248, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Rodilla, V.; Villanueva, A.; Obrador-Hevia, A.; Robert-Moreno, À.; Fernández-Majada, V.; Grilli, A.; López-Bigas, N.; Bellora, N.; Albà, M.M.; Torres, F. Jagged1 is the pathological link between Wnt and Notch pathways in colorectal cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 6315–6320. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhou, H. Circular RNA hsa_circRNA_102209 promotes the growth and metastasis of colorectal cancer through miR-761-mediated Ras and Rab interactor 1 signaling. Cancer Med. 2020, 9, 6710–6725. [Google Scholar] [CrossRef]
- Zhang, Q.; Sui, Y.; Sui, X. MicroRNA-761 inhibits the metastasis of gastric cancer by negatively regulating Ras and Rab interactor 1. Oncol. Lett. 2019, 18, 3097–3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, F.-L.; Cao, M.-Z.; Li, Y.-F. Circ_0000218 plays a carcinogenic role in colorectal cancer progression by regulating miR-139-3p/RAB1A axis. J. Biochem. 2020, 167, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Shao, X.; Xu, M.; Wang, J.; Kuai, X.; Zhang, L.; Wu, J.; Zhou, C.; Mao, J. Rab1A promotes proliferation and migration abilities via regulation of the HER2/AKT-independent mTOR/S6K1 pathway in colorectal cancer. Oncol. Rep. 2019, 41, 2717–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wu, G.; Sun, P.; Zhu, Y.; Zhang, H. circ_SMAD2 regulate colorectal cancer cells proliferation through targeting miR-1258/RPN2 signaling pathway. J. Cancer 2021, 12, 1678–1686. [Google Scholar] [CrossRef]
- Bi, C.; Jiang, B. Downregulation of RPN2 induces apoptosis and inhibits migration and invasion in colon carcinoma. Oncol. Rep. 2018, 40, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Sun, H.; Liu, X.; Chen, J.; Tian, Z.; Xu, J.; Xiang, B.; Qin, B. Circular RNA hsa_circ_0004277 contributes to malignant phenotype of colorectal cancer by sponging miR-512-5p to upregulate the expression of PTMA. J. Cell. Physiol. 2020. [Google Scholar] [CrossRef]
- Zhang, M.; Cui, F.; Lu, S.; Lu, H.; Jiang, T.; Chen, J.; Zhang, X.; Jin, Y.; Peng, Z.; Tang, H. Increased expression of prothymosin-α, independently or combined with TP53, correlates with poor prognosis in colorectal cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 4867–4876. [Google Scholar]
- Banyard, J.; Bielenberg, D.R. The role of EMT and MET in cancer dissemination. Connect. Tissue Res. 2015, 56, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Viralippurath Ashraf, J.; Sasidharan Nair, V.; Saleh, R.; Elkord, E. Role of circular RNAs in colorectal tumor microenvironment. Biomed. Pharmacother. 2021, 137, 111351. [Google Scholar] [CrossRef]
- Deng, Z.; Li, X.; Wang, H.; Geng, Y.; Cai, Y.; Tang, Y.; Wang, Y.; Yu, X.; Li, L.; Li, R. Dysregulation of circRNA_0001946 contributes to the proliferation and metastasis of colorectal cancer cells by targeting MicroRNA-135a-5p. Front. Genet. 2020, 11, 357. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Wu, K.; Zhan, F.; Zeng, H. Dysregulated circRNA_100876 contributes to proliferation and metastasis of colorectal cancer by targeting microRNA-516b (miR-516b). Cancer Biol. Ther. 2020, 21, 733–740. [Google Scholar] [CrossRef]
- Zeng, Y.; Xu, Y.; Shu, R.; Sun, L.; Tian, Y.; Shi, C.; Zheng, Z.; Wang, K.; Luo, H. Altered expression profiles of circular RNA in colorectal cancer tissues from patients with lung metastasis. Int. J. Mol. Med. 2017, 40, 1818–1828. [Google Scholar] [CrossRef]
- Wang, Z.; Su, M.; Xiang, B.; Zhao, K.; Qin, B. Circular RNA PVT1 promotes metastasis via miR-145 sponging in CRC. Biochem. Biophys. Res. Commun. 2019, 512, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yang, R.; Yang, L.; Wang, D.; Zhou, X.; Sun, Y. circANKS1B regulates FOXM1 expression and promotes cell migration and invasion by functioning as a sponge of the miR-149 in colorectal cancer. OncoTargets Ther. 2019, 12, 4065–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Shi, J.; He, Q.; Sun, G.; Gao, L.; Ye, J.; Tang, X.; Qu, H. Hsa_circ_0026416 promotes proliferation and migration in colorectal cancer via miR-346/NFIB axis. Cancer Cell Int. 2020, 20, 494. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, J.; Yuan, W.; Ruan, H.; Shu, Y.; Ji, J.; Wu, L.; Tang, Q.; Zhou, Z.; Zhang, X. Nuclear factor I/B promotes colorectal cancer cell proliferation, epithelial-mesenchymal transition and 5-fluorouracil resistance. Cancer Sci. 2019, 110, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Dai, G.J. Hsa_circRNA_000166 promotes cell proliferation, migration and invasion by regulating miR-330-5p/ELK1 in colon cancer. OncoTargets Ther. 2020, 13, 5529–5539. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, S.; Liao, N.; Huang, H.; Chen, W.; Wu, Z.; Wu, D. CircCTNNA1 acts as a ceRNA for miR-363-3p to facilitate the progression of colorectal cancer by promoting CXCL5 expression. J. Biol. Res.-Thessalon. 2021, 28, 7. [Google Scholar] [CrossRef]
- Zhao, J.; Ou, B.; Han, D.; Wang, P.; Zong, Y.; Zhu, C.; Liu, D.; Zheng, M.; Sun, J.; Feng, H. Tumor-derived CXCL5 promotes human colorectal cancer metastasis through activation of the ERK/Elk-1/Snail and AKT/GSK3β/β-catenin pathways. Mol. Cancer 2017, 16, 70. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Ren, X.; Xin, S.; Lan, X.; Lu, G.; Lin, Y.; Yang, S.; Zeng, Z.; Liao, W.; Ding, Y.-Q. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget 2016, 7, 26680–26691. [Google Scholar] [CrossRef]
- Yan, Y.; Su, M.; Qin, B. CircHIPK3 promotes colorectal cancer cells proliferation and metastasis via modulating of miR-1207-5p/FMNL2 signal. Biochem. Biophys. Res. Commun. 2020, 524, 839–846. [Google Scholar] [CrossRef]
- He, J.-H.; Li, Y.-G.; Han, Z.-P.; Zhou, J.-B.; Chen, W.-M.; Lv, Y.-B.; He, M.-L.; Zuo, J.-D.; Zheng, L. The CircRNA-ACAP2/Hsa-miR-21-5p/Tiam1 regulatory feedback circuit affects the proliferation, migration, and invasion of colon cancer SW480 cells. Cell. Physiol. Biochem. 2018, 49, 1539–1550. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Y.; Huang, J.; Wang, S.; Ni, W.; Guan, J.; Li, Q.; Zhang, Y.; Ding, Y.; Chen, B. MiR-141 suppresses the migration and invasion of HCC cells by targeting Tiam1. PLoS ONE 2014, 9, e88393. [Google Scholar] [CrossRef]
- Sun, J.; Liu, J.; Zhu, Q.; Xu, F.; Kang, L.; Shi, X. Hsa_circ_0001806 Acts as a ceRNA to facilitate the stemness of colorectal cancer cells by increasing COL1A1. OncoTargets Ther. 2020, 13, 6315–6327. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, L.; Shi, K.; Yuan, B.; Cao, G.; Kong, C.; Fu, J.; Man, Z.; Li, X.; Zhang, X. CircCSPP1 functions as a ceRNA to promote colorectal carcinoma cell EMT and liver metastasis by upregulating COL1A1. Front. Oncol. 2020, 10, 850. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Lin, Y.; Wang, Z.; Liu, Y.; Guo, W. Circ_0006174 Accelerates Colorectal Cancer Progression through Regulating miR-138-5p/MACC1 Axis. Cancer Manag. Res. 2021, 13, 1673–1686. [Google Scholar] [CrossRef]
- Stein, U.; Walther, W.; Arlt, F.; Schwabe, H.; Smith, J.; Fichtner, I.; Birchmeier, W.; Schlag, P.M. MACC1, a newly identified key regulator of HGF-MET signaling, predicts colon cancer metastasis. Nat. Med. 2009, 15, 59–67. [Google Scholar] [CrossRef]
- Sun, Y.; Cao, Z.; Shan, J.; Gao, Y.; Liu, X.; Ma, D.; Li, Z. Hsa_circ_0020095 Promotes Oncogenesis and Cisplatin Resistance in Colon Cancer by Sponging miR-487a-3p and Modulating SOX9. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Shen, Z.; Deng, H.; Fang, Y.; Zhu, X.; Ye, G.-t.; Yan, L.; Liu, H.; Li, G. Identification of the interplay between SOX9 and S100P in the metastasis and invasion of colon carcinoma. Oncotarget 2015, 6, 20672–20684. [Google Scholar] [CrossRef] [Green Version]
- Blache, P.; Canterel-Thouennon, L.; Busson, M.; Verdié, P.; Subra, G.; Ychou, M.; Prévostel, C. A short SOX9 peptide mimics SOX9 tumor suppressor activity and is sufficient to inhibit colon cancer cell growth. Mol. Cancer Ther. 2019, 18, 1386–1395. [Google Scholar] [CrossRef] [Green Version]
- Prévostel, C.; Blache, P. The dose-dependent effect of SOX9 and its incidence in colorectal cancer. Eur. J. Cancer 2017, 86, 150–157. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, Z.; Wang, S.; Zhu, W.; Zheng, P.; Wang, W. Circular RNA hsa_circ_0001178 facilitates the invasion and metastasis of colorectal cancer through upregulating ZEB1 via sponging multiple miRNAs. Biol. Chem. 2020, 401, 487–496. [Google Scholar] [CrossRef]
- Xu, H.; Wang, C.; Song, H.; Xu, Y.; Ji, G. RNA-Seq profiling of circular RNAs in human colorectal Cancer liver metastasis and the potential biomarkers. Mol. Cancer 2019, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tao, C.; Tang, Y.; Xia, Y.; Li, X.; Wang, X. Hypoxia-induced hsa_circ_0000826 is linked to liver metastasis of colorectal cancer. J. Clin. Lab. Anal. 2020, 34, e23405. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, H.; Yang, Y.; Wang, X.; Deng, T.; Liu, R.; Ning, T.; Bai, M.; Li, H.; Zhu, K. Hypoxia induced exosomal circRNA promotes metastasis of Colorectal Cancer via targeting GEF-H1/RhoA axis. Theranostics 2020, 10, 8211–8226. [Google Scholar] [CrossRef]
- Wu, L.; Xia, J.; Yang, J.; Shi, Y.; Xia, H.; Xiang, X.; Yu, X. Circ-ZNF609 promotes migration of colorectal cancer by inhibiting Gli1 expression via microRNA-150. J. BUON 2018, 23, 1343–1349. [Google Scholar]
- Liu, K.; Mou, Y.; Shi, X.; Liu, T.; Chen, Z.; Zuo, X. Circular RNA 100146 Promotes Colorectal Cancer Progression by the MicroRNA 149/HMGA2 Axis. Mol. Cell. Biol. 2021, 41. [Google Scholar] [CrossRef]
- Chen, R.-X.; Chen, X.; Xia, L.-P.; Zhang, J.-X.; Pan, Z.-Z.; Ma, X.-D.; Han, K.; Chen, J.-W.; Judde, J.-G.; Deas, O. N 6-methyladenosine modification of circNSUN2 facilitates cytoplasmic export and stabilizes HMGA2 to promote colorectal liver metastasis. Nat. Commun. 2019, 10, 4695. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.F.; Pei, F.L.; Cao, M.Z. CircRNA_101951 promotes migration and invasion of colorectal cancer cells by regulating the KIF3A-mediated EMT pathway. Exp. Ther. Med. 2020, 19, 3355–3361. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Meng, Q.; Sun, H.; Wu, S.; Hu, W.; Liu, G.; Li, X.; Yang, Y.; Chen, R. CircPTK2 (hsa_circ_0005273) as a novel therapeutic target for metastatic colorectal cancer. Mol. Cancer 2020, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Han, K.; Wang, F.-W.; Cao, C.-H.; Ling, H.; Chen, J.-W.; Chen, R.-X.; Feng, Z.-H.; Luo, J.; Jin, X.-H.; Duan, J.-L. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol. Cancer 2020, 19, 60. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-S.; Lin, C.-H.; Huang, L.-Y.; Qiu, X.-M. CircRNA SMARCC1 Sponges MiR-140-3p to Regulate Cell Progression in Colorectal Cancer. Cancer Manag. Res. 2020, 12, 4899–4910. [Google Scholar] [CrossRef]
- He, J.H.; Han, Z.P.; Luo, J.G.; Jiang, J.W.; Zhou, J.B.; Chen, W.M.; Lv, Y.B.; He, M.L.; Zheng, L.; Li, Y.G. Hsa_Circ_0007843 acts as a mIR-518c-5p sponge to regulate the migration and invasion of colon cancer SW480 cells. Front. Genet. 2020, 11, 9. [Google Scholar] [CrossRef]
- Xiao, H.; Liu, M. Circular RNA hsa_circ_0053277 promotes the development of colorectal cancer by upregulating matrix metallopeptidase 14 via miR-2467-3p sequestration. J. Cell. Physiol. 2020, 235, 2881–2890. [Google Scholar] [CrossRef]
- He, C.; Huang, C.; Zhou, R.; Yu, H. CircLMNB1 promotes colorectal cancer by regulating cell proliferation, apoptosis and epithelial-mesenchymal transition. OncoTargets Ther. 2019, 12, 6349–6359. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-B.; Huang, S.-S.; Lu, C.-G.; Tian, S.-D.; Chen, M. CircAPLP2 regulates the proliferation and metastasis of colorectal cancer by targeting miR-101-3p to activate the Notch signalling pathway. Am. J. Transl. Res. 2020, 12, 2554–2569. [Google Scholar]
- Liu, X.; Qin, Y.; Tang, X.; Wang, Y.; Bian, C.; Zhong, J. Circular RNA circ_0000372 contributes to the proliferation, migration and invasion of colorectal cancer by elevating IL6 expression via sponging miR-495. Anti-Cancer Drugs 2021, 32, 296–305. [Google Scholar] [CrossRef]
- Chen, J.; Yang, X.; Liu, R.; Wen, C.; Wang, H.; Huang, L.; Li, W.; Zhu, Z.; Zhu, Y.; Liu, H. Circular RNA GLIS2 promotes colorectal cancer cell motility via activation of the NF-κB pathway. Cell Death Dis. 2020, 11, 788. [Google Scholar] [CrossRef]
- Celtikci, B. A Crosstalk between Dual-Specific Phosphatases and Dual-Specific Protein Kinases Can Be a Potential Therapeutic Target for Anti-cancer Therapy. In Protein Kinase-Mediated Decisions between Life and Death; Springer: Berlin/Heidelberg, Germany, 2021; pp. 357–382. [Google Scholar]
- Li, R.; Wu, B.; Xia, J.; Ye, L.; Yang, X. Circular RNA hsa_circRNA_102958 promotes tumorigenesis of colorectal cancer via miR-585/CDC25B axis. Cancer Manag. Res. 2019, 11, 6887–6893. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Xu, J.; Li, C.; Dai, X.; Wu, T.; Wen, J. Circular RNA circ_0007142 facilitates colorectal cancer progression by modulating CDC25A expression via miR-122-5p. OncoTargets Ther. 2020, 13, 3689–3701. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.-L.; Sha, X.; Wang, Y.; Li, J.; Zhang, M.-Y.; Guo, Z.-Y.; Sun, S.-A.; He, J.-D. Circular RNA hsa_circ_0007142 is upregulated and targets miR-103a-2-5p in colorectal cancer. J. Oncol. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, T.; Wu, H.; Zhang, L.; Li, K.; Xiao, X.; Zhang, L.; Zhang, Y. Circular RNA circ_0007142 regulates cell proliferation, apoptosis, migration and invasion via miR-455-5p/SGK1 axis in colorectal cancer. Anti-Cancer Drugs 2020, 32, 22–33. [Google Scholar] [CrossRef]
- Chen, Z.; Ren, R.; Wan, D.; Wang, Y.; Xue, X.; Jiang, M.; Shen, J.; Han, Y.; Liu, F.; Shi, J. Hsa_circ_101555 functions as a competing endogenous RNA of miR-597-5p to promote colorectal cancer progression. Oncogene 2019, 38, 6017–6034. [Google Scholar] [CrossRef]
- Huang, X.; Shen, X.; Peng, L.; Mai, W.; Wan, Y.; Zhang, H. CircCSNK1G1 Contributes to the Development of Colorectal Cancer by Increasing the Expression of MYO6 via Competitively Targeting miR-455-3p. Cancer Manag. Res. 2020, 12, 9563–9575. [Google Scholar] [CrossRef]
- Xu, X.-W.; Zheng, B.-A.; Hu, Z.-M.; Qian, Z.-Y.; Huang, C.-J.; Liu, X.-Q.; Wu, W.-D. Circular RNA hsa_circ_000984 promotes colon cancer growth and metastasis by sponging miR-106b. Oncotarget 2017, 8, 91674–91683. [Google Scholar] [CrossRef]
- Yang, B.; Du, K.; Yang, C.; Xiang, L.; Xu, Y.; Cao, C.; Zhang, J.; Liu, W. CircPRMT5 circular RNA promotes proliferation of colorectal cancer through sponging miR-377 to induce E2F3 expression. J. Cell. Mol. Med. 2020, 24, 3431–3437. [Google Scholar] [CrossRef]
- Chen, P.; Yao, Y.; Yang, N.; Gong, L.; Kong, Y.; Wu, A. Circular RNA circCTNNA1 promotes colorectal cancer progression by sponging miR-149-5p and regulating FOXM1 expression. Cell Death Dis. 2020, 11, 557. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, C.; Ma, J.-X.; Ren, H.; Sun, Y.; Xu, J.-Z. Circular RNA PIP5K1A promotes colon cancer development through inhibiting miR-1273a. World J. Gastroenterol. 2019, 25, 5300–5309. [Google Scholar] [CrossRef] [PubMed]
- Vaiopoulos, A.G.; Papachroni, K.K.; Papavassiliou, A.G. Colon carcinogenesis: Learning from NF-κB and AP-1. Int. J. Biochem. Cell Biol. 2010, 42, 1061–1065. [Google Scholar] [CrossRef]
- Chaudhary, R.; Muys, B.R.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Li, X.L.; Zhu, Y.; Daulatabad, S.V.; Tsitsipatis, D.; Meltzer, P.S. A circular RNA from the MDM2 locus controls cell cycle progression by suppressing p53 levels. Mol. Cell. Biol. 2020, 40. [Google Scholar] [CrossRef]
- Guo, J.-N.; Li, J.; Zhu, C.-L.; Feng, W.-T.; Shao, J.-X.; Wan, L.; Huang, M.-D.; He, J.-D. Comprehensive profile of differentially expressed circular RNAs reveals that hsa_circ_0000069 is upregulated and promotes cell proliferation, migration, and invasion in colorectal cancer. OncoTargets Ther. 2016, 9, 7451–7458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wu, H.; Chu, F.; Zhang, L.; Xiao, X. Knockdown of circ_0000512 Inhibits Cell Proliferation and Promotes Apoptosis in Colorectal Cancer by Regulating miR-296-5p/RUNX1 Axis. OncoTargets Ther. 2020, 13, 7357–7368. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Zhao, P.; Zhou, H.; Mao, T. Has_circ_0055625 from circRNA profile increases colon cancer cell growth by sponging miR-106b-5p. J. Cell. Biochem. 2019, 120, 3027–3037. [Google Scholar] [CrossRef] [PubMed]
- Cabodi, S.; del Pilar Camacho-Leal, M.; Di Stefano, P.; Defilippi, P. Integrin signalling adaptors: Not only figurants in the cancer story. Nat. Rev. Cancer 2010, 10, 858–870. [Google Scholar] [CrossRef]
- Tang, X.; Sun, G.; He, Q.; Wang, C.; Shi, J.; Gao, L.; Ye, J.; Liang, Y.; Qu, H. Circular noncoding RNA circMBOAT2 is a novel tumor marker and regulates proliferation/migration by sponging miR-519d-3p in colorectal cancer. Cell Death Dis. 2020, 11, 625. [Google Scholar] [CrossRef]
- Ye, X.; Lv, H. MicroRNA-519d-3p inhibits cell proliferation and migration by targeting TROAP in colorectal cancer. Biomed. Pharmacother. 2018, 105, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, J.; Zhao, J.; Wang, X. Silencing of hsa_circ_0007534 suppresses proliferation and induces apoptosis in colorectal cancer cells. Eur. Rev. Med. Pharm. Sci. 2018, 22, 118–126. [Google Scholar] [CrossRef]
- Xian, Z.; Hu, B.; Wang, T.; Cai, J.; Zeng, J.; Zou, Q.; Zhu, P. CircABCB10 silencing inhibits the cell ferroptosis and apoptosis by regulating the miR-326/CCL5 axis in rectal cancer. Neoplasma 2020, 67, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-N.; Wang, Z.-J.; Ye, C.-X.; Zhao, B.-C.; Huang, X.-X.; Yang, L. Circular RNA circVAPA is up-regulated and exerts oncogenic properties by sponging miR-101 in colorectal cancer. Biomed. Pharmacother. 2019, 112, 108611. [Google Scholar] [CrossRef]
- Dai, J.; Zhuang, Y.; Tang, M.; Qian, Q.; Chen, J. CircRNA UBAP2 facilitates the progression of colorectal cancer by regulating miR-199a/VEGFA pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7963–7971. [Google Scholar] [CrossRef]
- Chen, C.; Huang, Z.; Mo, X.; Song, Y.; Li, X.; Li, X.; Zhang, M. The circular RNA 001971/miR-29c-3p axis modulates colorectal cancer growth, metastasis, and angiogenesis through VEGFA. J. Exp. Clin. Cancer Res. 2020, 39, 91. [Google Scholar] [CrossRef]
- Li, W.; Xu, Y.; Wang, X.; Cao, G.; Bu, W.; Wang, X.; Fang, Z.; Xu, Y.; Dong, M.; Tao, Q. circCCT3 modulates vascular endothelial growth factor A and Wnt signaling to enhance colorectal cancer metastasis through sponging miR-613. DNA Cell Biol. 2020, 39, 118–125. [Google Scholar] [CrossRef]
- Zheng, X.; Ma, Y.; Zhang, X.; Li, Y.; Zhao, H.; Han, S. Circ_0056618 promoted cell proliferation, migration and angiogenesis through sponging with miR-206 and upregulating CXCR4 and VEGF-A in colorectal cancer. Eur. Rev. Med. Pharm. Sci. 2020, 24, 4190–4202. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Cervantes, A.; Nordlinger, B.; Arnold, D. Metastatic colorectal cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2014, 25, iii1–iii9. [Google Scholar] [CrossRef]
- de Gramont, A.d.; Figer, A.; Seymour, M.; Homerin, M.; Hmissi, A.; Cassidy, J.; Boni, C.; Cortes-Funes, H.; Cervantes, A.; Freyer, G. Leucovorin and fluorouracil with or without oxaliplatin as first-line treatment in advanced colorectal cancer. J. Clin. Oncol. 2000, 18, 2938–2947. [Google Scholar] [CrossRef]
- Li, S.; Zheng, S. Down-Regulation of Circ_0032833 Sensitizes Colorectal Cancer to 5-Fluorouracil and Oxaliplatin Partly Depending on the Regulation of miR-125-5p and MSI1. Cancer Manag. Res. 2020, 12, 11257–11269. [Google Scholar] [CrossRef] [PubMed]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef]
- Lai, M.; Liu, G.; Li, R.; Bai, H.; Zhao, J.; Xiao, P.; Mei, J. Hsa_circ_0079662 induces the resistance mechanism of the chemotherapy drug oxaliplatin through the TNF-α pathway in human colon cancer. J. Cell. Mol. Med. 2020, 24, 5021–5027. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-C.; Yu, Y.-S.; Lin, H.-H.; Hsiao, K.-Y. Oxaliplatin-induced DHX9 phosphorylation promotes oncogenic circular RNA CCDC66 expression and development of chemoresistance. Cancers 2020, 12, 697. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, K.-Y.; Lin, Y.-C.; Gupta, S.K.; Chang, N.; Yen, L.; Sun, H.S.; Tsai, S.-J. Noncoding effects of circular RNA CCDC66 promote colon cancer growth and metastasis. Cancer Res. 2017, 77, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Peng, X.; Lu, X.; Wei, Q.; Chen, M.; Liu, L. Inhibition of hsa_circ_0001313 (circCCDC66) induction enhances the radio-sensitivity of colon cancer cells via tumor suppressor miR-338-3p: Effects of cicr_0001313 on colon cancer radio-sensitivity. Pathol.-Res. Pract. 2019, 215, 689–696. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.; Cai, G.; Huang, P. Downregulation of Circ_0071589 Suppresses Cisplatin Resistance in Colorectal Cancer by Regulating the MiR-526b-3p/KLF12 Axis. Cancer Manag. Res. 2021, 13, 2717–2731. [Google Scholar] [CrossRef]
- Chen, H.; Pei, L.; Xie, P.; Guo, G. Circ-PRKDC contributes to 5-fluorouracil resistance of colorectal cancer cells by regulating miR-375/FOXM1 axis and Wnt/β-catenin pathway. Onco Targets Ther. 2020, 13, 5939–5953. [Google Scholar] [CrossRef]
- Xiong, W.; Ai, Y.-Q.; Li, Y.-F.; Ye, Q.; Chen, Z.-T.; Qin, J.-Y.; Liu, Q.-Y.; Wang, H.; Ju, Y.-H.; Li, W.-H. Microarray analysis of circular RNA expression profile associated with 5-fluorouracil-based chemoradiation resistance in colorectal cancer cells. BioMed. Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Zhang, J.; Chu, Z.; Liu, P.; Zhang, X.; Li, C.; Gu, X. Circ_0007031 Serves as a Sponge of miR-760 to Regulate the Growth and Chemoradiotherapy Resistance of Colorectal Cancer via Regulating DCP1A. Cancer Manag. Res. 2020, 12, 8465–8479. [Google Scholar] [CrossRef]
- He, X.; Ma, J.; Zhang, M.; Cui, J.; Yang, H. Circ_0007031 enhances tumor progression and promotes 5-fluorouracil resistance in colorectal cancer through regulating miR-133b/ABCC5 axis. Cancer Biomark. 2020. [Google Scholar] [CrossRef] [PubMed]
- Abu, N.; Hon, K.W.; Jeyaraman, S.; Yahaya, A.; Abdullah, N.M.; Mustangin, M.; Sulaiman, S.A.; Jamal, R.; Ab-Mutalib, N.-S. Identification of differentially expressed circular RNAs in chemoresistant colorectal cancer. Epigenomics 2019, 11, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zheng, W.; Li, N.; Su, Z.; Zhao, L.; Zhou, H.; Jia, L. MicroRNA-130b targets PTEN to mediate drug resistance and proliferation of breast cancer cells via the PI3K/Akt signaling pathway. Sci. Rep. 2017, 7, 41942. [Google Scholar] [CrossRef]
- Xi, L.; Liu, Q.; Zhang, W.; Luo, L.; Song, J.; Liu, R.; Wei, S.; Wang, Y. Circular RNA circCSPP1 knockdown attenuates doxorubicin resistance and suppresses tumor progression of colorectal cancer via miR-944/FZD7 axis. Cancer Cell Int. 2021, 21, 153. [Google Scholar] [CrossRef]
- Jian, X.; He, H.; Zhu, J.; Zhang, Q.; Zheng, Z.; Liang, X.; Chen, L.; Yang, M.; Peng, K.; Zhang, Z. Hsa_circ_001680 affects the proliferation and migration of CRC and mediates its chemoresistance by regulating BMI1 through miR-340. Mol. Cancer 2020, 19, 20. [Google Scholar] [CrossRef]
- Wang, P.; Sun, Y.; Yang, Y.; Chen, Y.; Liu, H. Circ_0067835 Knockdown Enhances the Radiosensitivity of Colorectal Cancer by miR-296-5p/IGF1R Axis. OncoTargets Ther. 2021, 14, 491–502. [Google Scholar] [CrossRef]
- Santer, L.; Bär, C.; Thum, T. Circular RNAs: A Novel Class of Functional RNA Molecules with a Therapeutic Perspective. Mol. Ther. 2019, 27, 1350–1363. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Jiang, C.; Li, S. The potential regulatory roles of circular RNAs in tumor immunology and immunotherapy. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Wang, X.; Teng, F.; Kong, L.; Yu, J. PD-L1 expression in human cancers and its association with clinical outcomes. OncoTargets Ther. 2016, 9, 5023–5039. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.l.; Xu, L.l.; Wang, F. Hsa_circ_0020397 regulates colorectal cancer cell viability, apoptosis and invasion by promoting the expression of the miR-138 targets TERT and PD-L1. Cell Biol. Int. 2017, 41, 1056–1064. [Google Scholar] [CrossRef]
- Liu, T.; Yuan, X.; Xu, D. Cancer-specific telomerase reverse transcriptase (TERT) promoter mutations: Biological and clinical implications. Genes 2016, 7, 38. [Google Scholar] [CrossRef]
- Tanaka, E.; Miyakawa, Y.; Kishikawa, T.; Seimiya, T.; Iwata, T.; Funato, K.; Odawara, N.; Sekiba, K.; Yamagami, M.; Suzuki, T. Expression of circular RNA CDR1-AS in colon cancer cells increases cell surface PD-L1 protein levels. Oncol. Rep. 2019, 42, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.E.; Short, S.P.; Williams, C.S. Colorectal cancer and metabolism. Curr. Colorectal Cancer Rep. 2018, 14, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Faes, S.; Duval, A.P.; Planche, A.; Uldry, E.; Santoro, T.; Pythoud, C.; Stehle, J.-C.; Horlbeck, J.; Letovanec, I.; Riggi, N. Acidic tumor microenvironment abrogates the efficacy of mTORC1 inhibitors. Mol. Cancer 2016, 15, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pate, K.T.; Stringari, C.; Sprowl-Tanio, S.; Wang, K.; TeSlaa, T.; Hoverter, N.P.; McQuade, M.M.; Garner, C.; Digman, M.A.; Teitell, M.A. Wnt signaling directs a metabolic program of glycolysis and angiogenesis in colon cancer. EMBO J. 2014, 33, 1454–1473. [Google Scholar] [CrossRef]
- Kawada, K.; Nakamoto, Y.; Kawada, M.; Hida, K.; Matsumoto, T.; Murakami, T.; Hasegawa, S.; Togashi, K.; Sakai, Y. Relationship between 18F-fluorodeoxyglucose accumulation and KRAS/BRAF mutations in colorectal cancer. Clin. Cancer Res. 2012, 18, 1696–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, K.; Kawada, K.; Iwamoto, M.; Inamoto, S.; Sasazuki, T.; Shirasawa, S.; Hasegawa, S.; Sakai, Y. Metabolic alterations caused by KRAS mutations in colorectal cancer contribute to cell adaptation to glutamine depletion by upregulation of asparagine synthetase. Neoplasia 2016, 18, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 regulates assembly and activation of AMPK complex under metabolic stress. Cell Metab. 2019, 30, 157–173.e7. [Google Scholar] [CrossRef]
- Wang, X.; Tao, G.; Huang, D.; Liang, S.; Zheng, D. Circular RNA NOX4 promotes the development of colorectal cancer via the microRNA-485-5p/CKS1B axis. Oncol. Rep. 2020, 44, 2009–2020. [Google Scholar] [CrossRef]
- Steven, A.; Heiduk, M.; Recktenwald, C.V.; Hiebl, B.; Wickenhauser, C.; Massa, C.; Seliger, B. Colorectal carcinogenesis: Connecting K-RAS–Induced transformation and CREB activity in vitro and in vivo. Mol. Cancer Res. 2015, 13, 1248–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zang, H.; Zhang, X.; Huang, G. circ_0136666 Facilitates the Progression of Colorectal Cancer via miR-383/CREB1 Axis. Cancer Manag. Res. 2020, 12, 6795–6806. [Google Scholar] [CrossRef]
- Lu, X.; Yu, Y.; Liao, F.; Tan, S. Homo Sapiens circular RNA 0079993 (hsa_circ_0079993) of the POLR2J4 gene acts as an oncogene in colorectal cancer through the microRNA-203a-3p. 1 and CREB1 axis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 6872–6883. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Y.; Yamaguchi, K.; Hu, J.; Zhang, L.; Wang, J.; Tian, J.; Chen, W. Circular RNA circVAPA knockdown suppresses colorectal cancer cell growth process by regulating miR-125a/CREB5 axis. Cancer Cell Int. 2020, 20, 103. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, J.; Wang, Z.; Xi, J.; Bai, L.; Zhang, Y. Knockdown of circAPLP2 Inhibits Progression of Colorectal Cancer by Regulating miR-485-5p/FOXK1 Axis. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Qin, X.; Wang, Y.; Fu, J. Circular RNA circDENND4C facilitates proliferation, migration and glycolysis of colorectal cancer cells through miR-760/GLUT1 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2387–2400. [Google Scholar] [CrossRef] [PubMed]
- Spitzner, M.; Roesler, B.; Bielfeld, C.; Emons, G.; Gaedcke, J.; Wolff, H.A.; Rave-Fränk, M.; Kramer, F.; Beissbarth, T.; Kitz, J. STAT3 inhibition sensitizes colorectal cancer to chemoradiotherapy in vitro and in vivo. Int. J. Cancer 2014, 134, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, L.; Lindner, A.; de Chaumont, C.; Kehoe, J.; Fay, J.; Bacon, O.; Toomey, S.; Huber, H.; Hennessy, B.; Kay, E. BCL2 protein signalling determines acute responses to neoadjuvant chemoradiotherapy in rectal cancer. J. Mol. Med. 2015, 93, 315–326. [Google Scholar] [CrossRef]
- Tian, J.; Xi, X.; Wang, J.; Yu, J.; Huang, Q.; Ma, R.; Zhang, X.; Li, H.; Wang, L. CircRNA hsa_circ_0004585 as a potential biomarker for colorectal cancer. Cancer Manag. Res. 2019, 11, 5413–5423. [Google Scholar] [CrossRef]
- Ye, D.-X.; Wang, S.-S.; Huang, Y.; Chi, P. A 3-circular RNA signature as a noninvasive biomarker for diagnosis of colorectal cancer. Cancer Cell Int. 2019, 19, 276. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, J.; Li, P.; Li, N.; Zhang, Y.; Binang, H.; Zhao, Y.; Duan, W.; Chen, Y.; Wang, Y. RNA-seq profiling of serum exosomal circular RNAs reveals Circ-PNN as a potential biomarker for human colorectal cancer. Front. Oncol. 2020, 10, 982. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Xia, M.; Jiao, X.; Fan, L. Hsa_circ_0004831 serves as a blood-based prognostic biomarker for colorectal cancer and its potentially circRNA-miRNA-mRNA regulatory network construction. Cancer Cell Int. 2020, 20, 557. [Google Scholar] [CrossRef] [PubMed]
- Bahn, J.H.; Zhang, Q.; Li, F.; Chan, T.-M.; Lin, X.; Kim, Y.; Wong, D.T.; Xiao, X. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin. Chem. 2015, 61, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Chen, S.; Chen, H.; Mo, X.; Li, T.; Shao, Y.; Xiao, B.; Guo, J. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clin. Chim. Acta 2015, 444, 132–136. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, H.Q.; Zhao, Q.; Wang, F.; Lan, P.; Wang, Z.; Zuo, Z.X.; Wu, Q.N.; Fan, X.J.; Mo, H.Y.; Chen, L. A circRNA signature predicts postoperative recurrence in stage II/III colon cancer. EMBO Mol. Med. 2019, 11, e10168. [Google Scholar] [CrossRef]
- Li, J.; Song, Y.; Wang, J.; Huang, J. Plasma circular RNA panel acts as a novel diagnostic biomarker for colorectal cancer detection. Am. J. Transl. Res. 2020, 12, 7395–7403. [Google Scholar]
- Lin, J.; Cai, D.; Li, W.; Yu, T.; Mao, H.; Jiang, S.; Xiao, B. Plasma circular RNA panel acts as a novel diagnostic biomarker for colorectal cancer. Clin. Biochem. 2019, 74, 60–68. [Google Scholar] [CrossRef]
- Pan, B.; Qin, J.; Liu, X.; He, B.; Wang, X.; Pan, Y.; Sun, H.; Xu, T.; Xu, M.; Chen, X. Identification of serum exosomal hsa-circ-0004771 as a novel diagnostic biomarker of colorectal cancer. Front. Genet. 2019, 10, 1096. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, S.; Liu, Y.; Wang, Y.; Lin, T.; Li, Y.; Zhang, R. Hsa_circ_0007534 as a blood-based marker for the diagnosis of colorectal cancer and its prognostic value. Int. J. Clin. Exp. Pathol. 2018, 11, 1399–1406. [Google Scholar] [PubMed]
- Zhou, G.R.; Huang, D.P.; Sun, Z.F.; Zhang, X.F. Characteristics and prognostic significance of circRNA-100876 in patients with colorectal cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11587–11593. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Li, P.; Gong, P.Y. Hsa_circ_0005075 promotes the proliferation and invasion of colorectal cancer cells. Int. J. Biol. Markers 2019, 34, 284–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radanova, M.; Mihaylova, G.; Nazifova-Tasinova, N.; Levkova, M.; Tasinov, O.; Ivanova, D.; Mihaylova, Z.; Donev, I. Oncogenic Functions and Clinical Significance of Circular RNAs in Colorectal Cancer. Cancers 2021, 13, 3395. https://doi.org/10.3390/cancers13143395
Radanova M, Mihaylova G, Nazifova-Tasinova N, Levkova M, Tasinov O, Ivanova D, Mihaylova Z, Donev I. Oncogenic Functions and Clinical Significance of Circular RNAs in Colorectal Cancer. Cancers. 2021; 13(14):3395. https://doi.org/10.3390/cancers13143395
Chicago/Turabian StyleRadanova, Maria, Galya Mihaylova, Neshe Nazifova-Tasinova, Mariya Levkova, Oskan Tasinov, Desislava Ivanova, Zhasmina Mihaylova, and Ivan Donev. 2021. "Oncogenic Functions and Clinical Significance of Circular RNAs in Colorectal Cancer" Cancers 13, no. 14: 3395. https://doi.org/10.3390/cancers13143395
APA StyleRadanova, M., Mihaylova, G., Nazifova-Tasinova, N., Levkova, M., Tasinov, O., Ivanova, D., Mihaylova, Z., & Donev, I. (2021). Oncogenic Functions and Clinical Significance of Circular RNAs in Colorectal Cancer. Cancers, 13(14), 3395. https://doi.org/10.3390/cancers13143395