
The Effects of Exercise Duration and Intensity on Breast Cancer-Related DNA Methylation: A Randomized Controlled Trial

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Design
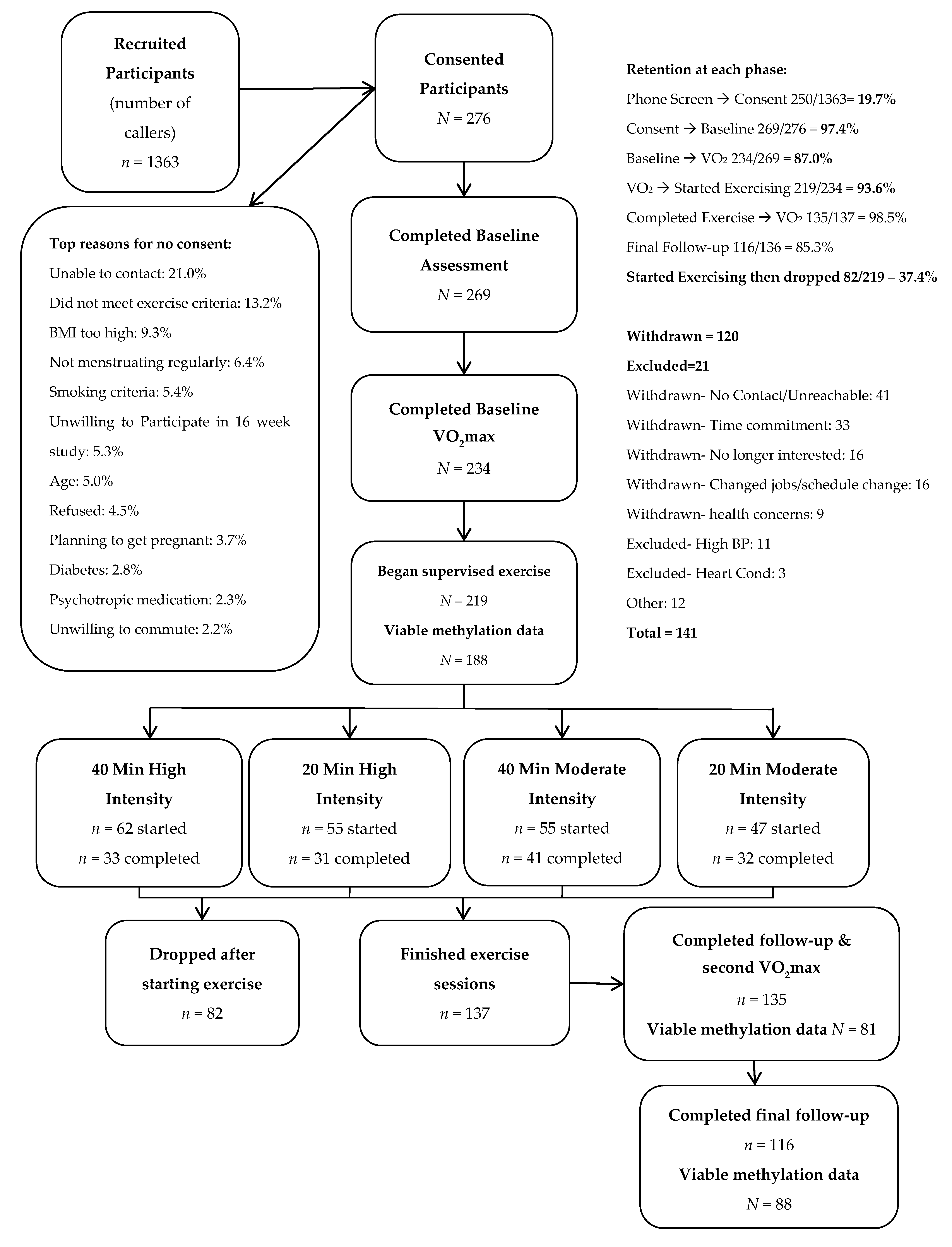
2.2. Participants
2.3. Measures
2.3.1. Demographics
2.3.2. VO2max
2.4. Exercise Volume Calculation
2.5. BMI
2.6. Gene Methylation
2.7. Follow-Up Exercise Behavior
2.8. Procedures
2.9. Statistical Analysis
3. Results
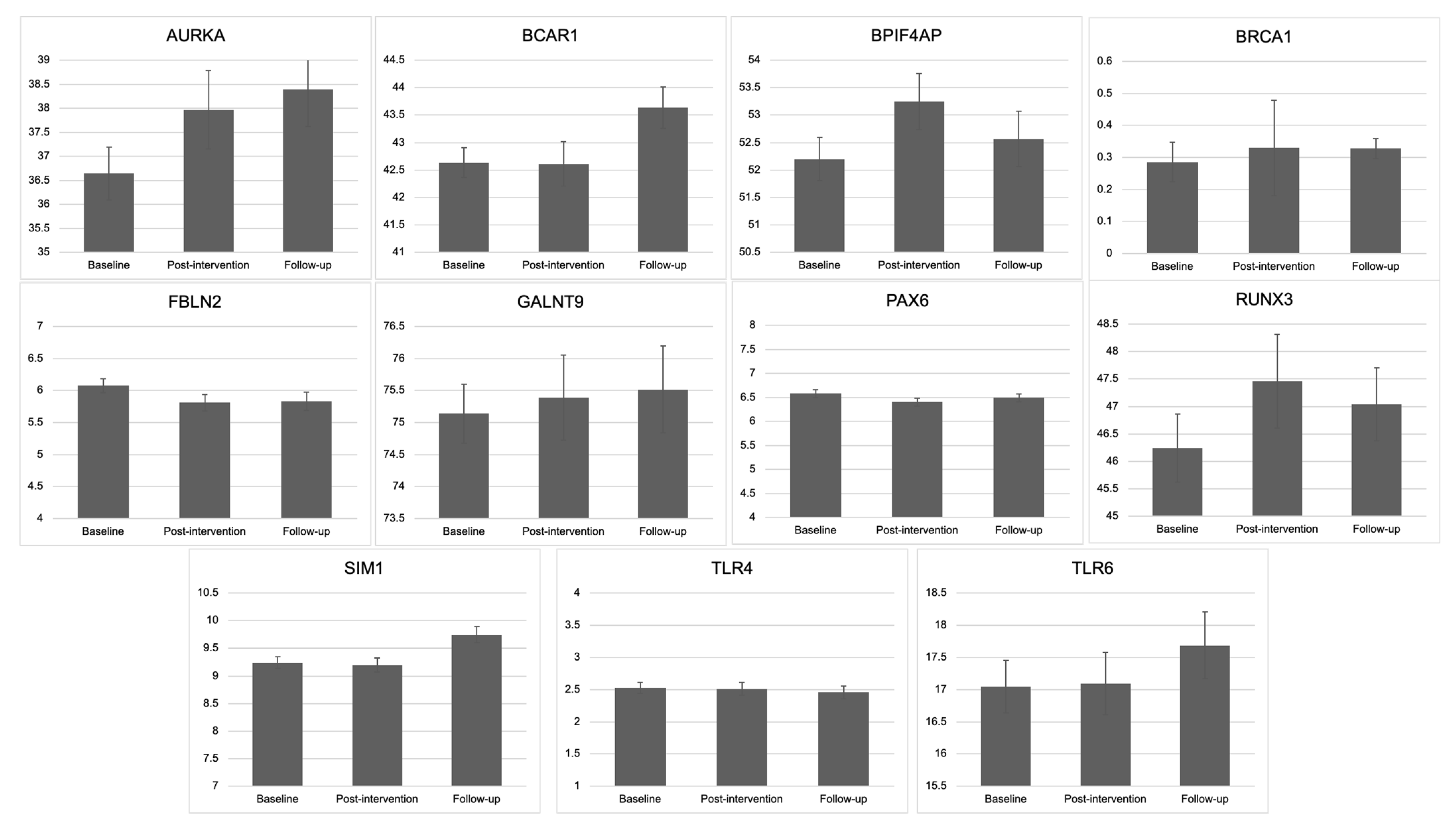
3.1. Change in Methylation: Baseline, Post-Intervention, and Follow-Up
3.2. Change in Methylation from Baseline to Post-Intervention Based on Quantity of Exercise Completed
3.2.1. Exercise Volume
3.2.2. VO2max
3.2.3. Exercise Condition
3.3. Six-Month Follow-Up Methylation Change as a Function of Level of Continued Exercise
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.; et al. SEER Cancer Statistics Review, 1975–2017; National Cancer Institute: Bethesda, MD, USA, 2020. [Google Scholar]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Neyman, N.; Aminou, R.; Altekruse, S.F.; Kosary, C.L.; Ruhl, J.; Tatalovich, Z.; Cho, H.; et al. SEER Cancer Statistics Review, 1975–2009 (Vintage 2009 Populations). Available online: https://seer.cancer.gov/archive/csr/1975_2009_pops09/ (accessed on 1 June 2021).
- Monninkhof, E.M.; Elias, S.G.; Vlems, F.A.; van der Tweel, I.; Schuit, A.J.; Voskuil, D.W.; van Leeuwen, F.E. Physical activity and breast cancer: A systematic review. Epidemiology 2007, 18, 137–157. [Google Scholar] [CrossRef] [Green Version]
- Friedenreich, C.M.; Thune, I. A review of physical activity and prostate cancer risk. Cancer Causes Control 2001, 12, 461–475. [Google Scholar] [CrossRef]
- Friedenreich, C.M.; Orenstein, M.R. Physical activity and cancer prevention: Etiologic evidence and biological mechanisms. J. Nutr. 2002, 132, 3456S–3464S. [Google Scholar] [CrossRef]
- Lynch, B.M.; Neilson, H.K.; Friedenreich, C.M. Physical activity and breast cancer prevention. In Physical Activity and Cancer; Springer: Berlin/Heidelberg, Germany, 2010; pp. 13–42. [Google Scholar]
- Rogers, C.J.; Colbert, L.H.; Greiner, J.W.; Perkins, S.N.; Hursting, S.D. Physical activity and cancer prevention. Sport Med. 2008, 38, 271–296. [Google Scholar] [CrossRef]
- Hojman, P.; Gehl, J.; Christensen, J.F.; Pedersen, B.K. Molecular Mechanisms Linking Exercise to Cancer Prevention and Treatment. Cell Metab. 2018, 27, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Boyne, D.J.; O’Sullivan, D.E.; Olij, B.F.; King, W.D.; Friedenreich, C.M.; Brenner, D.R. Physical activity, global DNA methylation, and breast cancer risk: A systematic literature review and meta-analysis. Cancer Epidemiol. Biomark. Prev. 2018, 27, 1320–1331. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Irwin, M.L. Effects of Physical Activity on DNA Methylation and Associations with Breast Cancer; Springer: Cham, Switzerland, 2016; pp. 251–264. [Google Scholar]
- Jones, P.A.; Baylin, S.B. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet 2002, 3, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M.; Corn, P.G.; Baylin, S.B.; Herman, J.G. A gene hypermethylation profile of human cancer. Cancer Res. 2001, 61, 3225–3229. [Google Scholar]
- Herman, J.G.; Baylin, S.B. Gene silencing in cancer in association with promoter hypermethylation. N. Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.; Esteller, M. Cancer epigenomics: Beyond genomics. Curr. Opin. Genet Dev. 2012, 22, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Sarkies, P.; Sale, J.E. Cellular epigenetic stability and cancer. Trends Genet. 2012, 28, 118–127. [Google Scholar] [CrossRef]
- Widschwendter, M.; Jones, A.; Teschendorff, A.E. Epigenetics makes its mark on women-specific cancers—an opportunity to redefine oncological approaches? Gynecol. Oncol. 2013, 128, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.-M.; Tan, J.-X.; Lu, X.-W.; Ding, Y.; Han, X.; Sun, Y.-J. BCSG1 methylation status and BCSG1 expression in breast tissues derived from Chinese women with breast cancer. Oncology 2008, 74, 61–68. [Google Scholar] [CrossRef]
- Veeck., J.; Esteller, M. Breast cancer epigenetics: From DNA methylation to microRNAs. J. Mammary Gland Biol. Neoplasia 2010, 15, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Tobias, D.K.; Akinkuolie, A.O.; Chandler, P.D.; Lawler, P.R.; Manson, J.E.; Buring, J.E.; Ridker, P.M.; Wang, L.; Lee, I.-M.; Mora, S. Markers of Inflammation and Incident Breast Cancer Risk in the Women’s Health Study. Am. J. Epidemiol. 2017, 187, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, Inflammation, and Cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, H.; Feng, P.; Zhou, X.; Wen, H.; Xie, X.; Shen, H.; Zhu, X. Reduced expression of Toll-like receptor 4 inhibits human breast cancer cells proliferation and inflammatory cytokines secretion. J. Exp. Clin. Cancer Res. 2010, 29, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinsky, S.A.; Palmisano, W.A.; Gilliland, F.D.; Crooks, L.A.; Divine, K.K.; Winters, S.A.; Grimes, M.J.; Harms, H.J.; Tellez, C.S.; Smith, T.M. Aberrant promoter methylation in bronchial epithelium and sputum from current and former smokers. Cancer Res. 2002, 62, 2370–2377. [Google Scholar]
- Ronn, T.; Volkov, P.; Gillberg, L.; Kokosar, M.; Perfilyev, A.; Jacobsen, A.L.; Jorgensen, S.W.; Brons, C.; Jansson, P.-A.; Eriksson, K.-F.; et al. Impact of age, BMI and HbA1c levels on the genome-wide DNA methylation and mRNA expression patterns in human adipose tissue and identification of epigenetic biomarkers in blood. Hum. Mol. Genet. 2015, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillman, A.S.; Gardiner, C.K.; Koljack, C.E.; Bryan, A.D. Body mass index, diet, and exercise: Testing possible linkages to breast cancer risk via DNA methylation. Breast Cancer Res. Treat. 2018, 168, 241–248. [Google Scholar] [CrossRef]
- Coyle, Y.M.; Xie, X.-J.; Lewis, C.M.; Bu, D.; Milchgrub, S.; Euhus, D.M. Role of Physical Activity in Modulating Breast Cancer Risk as Defined by APC and RASSF1A Promoter Hypermethylation in Nonmalignant Breast Tissue. Cancer Epidemiol. Prev. Biomark. 2007, 16, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, A.D.; Magnan, R.E.; Hooper, A.E.C.; Harlaar, N.; Hutchison, K.E. Physical Activity and Differential Methylation of Breast Cancer Genes Assayed from Saliva: A Preliminary Investigation. Ann. Behav. Med. 2013, 45, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.F.; Cardarelli, R.; Carroll, J.; Zhang, S.; Fulda, K.G.; Gonzalez, K.; Vishwanatha, J.K.; Morabia, A.; Santella, R.M. Physical activity and global genomic DNA methylation in a cancer-free population. Epigenetics 2011, 6, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyne, D.J.; King, W.D.; Brenner, D.R.; McIntyre, J.B.; Courneya, K.S.; Friedenreich, C.M. Aerobic exercise and DNA methylation in postmenopausal women: An ancillary analysis of the Alberta Physical Activity and Breast Cancer Prevention (ALPHA) Trial. PLoS ONE 2018, 13, e0198641. [Google Scholar] [CrossRef]
- Chen, L. Tumor suppressor function of RUNX3 in breast cancer. J. Cell Biochem. 2012, 113, 1470–1477. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Yin, W.; Zhang, X.; Yu, X.; Wang, C.; Xu, S.; Feng, W.; Yang, H. PAX6 overexpression is associated with the poor prognosis of invasive ductal breast cancer. Oncol. Lett. 2015, 10, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.R.M.; Hoessli, D.C.; Fang, M. N-acetylgalactosaminyltransferases in cancer. Oncotarget 2016, 7, 54067. [Google Scholar] [CrossRef] [Green Version]
- Kwak, H.-I.; Gustafson, T.; Metz, R.P.; Laffin, B.; Schedin, P.; Porter, W.W. Inhibition of breast cancer growth and invasion by single-minded 2s. Carcinogenesis 2007, 28, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Hill, V.K.; Hesson, L.B.; Dansranjavin, T.; Dallol, A.; Bieche, I.; Vacher, S.; Tommasi, S.; Dobbins, T.; Gentle, D.; Euhus, D.; et al. Identification of 5 novel genes methylated in breast and other epithelial cancers. Mol. Cancer 2010, 9, e51. [Google Scholar] [CrossRef] [Green Version]
- Du, R.; Huang, C.; Liu, K.; Li, X.; Dong, Z. Targeting AURKA in Cancer: Molecular mechanisms and opportunities for Cancer therapy. Mol. Cancer 2021, 20, e15. [Google Scholar] [CrossRef] [PubMed]
- Wallez, Y.; Riedl, S.J.; Pasquale, E.B. Association of the breast cancer antiestrogen resistance protein 1 (BCAR1) and BCAR3 scaffolding proteins in cell signaling and antiestrogen resistance. J. Biol. Chem. 2014, 289, 10431–10444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretschneider, N.; Brand, H.; Miller, N.; Lowery, A.J.; Kerin, M.J.; Gannon, F.; Denger, S. Estrogen induces repression of the breast cancer and salivary gland expression gene in an estrogen receptor α-dependent manner. Cancer Res. 2008, 68, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, Q.C.; Raja, E.; Salto-Tellez, M.; Liu, Q.; Ito, K.; Inoue, M.; Putti, T.C.; Loh, M.; Ko, T.K.; Huang, C. RUNX3 is frequently inactivated by dual mechanisms of protein mislocalization and promoter hypermethylation in breast cancer. Cancer Res. 2006, 66, 6512–6520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, H.; Yamashita, S.; Fujii, S.; Tanabe, K.; Mukai, H.; Ushijima, T. DNA methylation marker to estimate the breast cancer cell fraction in DNA samples. Med. Oncol. 2018, 35, 147. [Google Scholar] [CrossRef] [PubMed]
- Pangeni, R.P.; Channathodiyil, P.; Huen, D.S.; Eagles, L.W.; Johal, B.K.; Pasha, D.; Hadjistephanou, N.; Nevell, O.; Davies, C.L.; Adewumi, A.I. The GALNT9, BNC1 and CCDC8 genes are frequently epigenetically dysregulated in breast tumours that metastasise to the brain. Clin. Epigenetics 2015, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Vojta, A.; Samaržija, I.; Bočkor, L.; Zoldoš, V. Glyco-genes change expression in cancer through aberrant methylation. Biochim. Biophys. Acta-Gen. Subj. 2016, 1860, 1776–1785. [Google Scholar] [CrossRef]
- Medina-Aguilar, R.; Pérez-Plasencia, C.; Marchat, L.A.; Gariglio, P.; García Mena, J.; Rodríguez Cuevas, S.; Ruíz-García, E.; Astudillo-De La Vega, H.; Hernández Juárez, J.; Flores-Pérez, A.; et al. Methylation landscape of human breast cancer cells in response to dietary compound resveratrol. PLoS ONE 2016, 11, e0157866. [Google Scholar] [CrossRef]
- Harris, P.A.; Taylor, R.; Thielke, R.; Payne, J.; Gonzalez, N.; Conde, J.G. Research electronic data capture (REDCap)—a metadata-driven methodology and workflow process for providing translational research informatics support. J. Biomed. Inform. 2009, 42, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Pescatello, L.S.; Riebe, D.; Thompson, P.D. ACSM’s Guidelines for exercise Testing and Prescription; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- Reed, K.; Poulin, M.L.; Yan, L.; Parissenti, A.M. Comparison of bisulfite sequencing PCR with pyrosequencing for measuring differences in DNA methylation. Anal. Biochem. 2010, 397, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.N.; Haskell, W.L.; Ho, P.; Paffenbarger, R.S.; Vranizan, K.M.; Farquhar, J.W.; Wood, P.D. Assessment of habitual physical activity by a sevenday recall in a community survey and controlled experiments. Am. J. Epidemiol. 1985, 122, 794–804. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Nakajima, N.; Takeika, M.; Mori, M.; Hashimoto, S.; Sakurai, A.; Nose, H.; Itano, N.; Shiohara, M.; Oh, T.; Taniguchi, S. Exercise effects on methylation of ASC gene. Int. J. Sport Med. 2009, 30, 1–5. [Google Scholar]
- Teschendorff, A.E.; Menon, U.; Gentry-Maharaj, A.; Ramus, S.J.; Weisenberger, D.J.; Shen, H.; Campan, M.; Noushmehr, H.; Bell, C.G.; Maxwell, A.P. Age-dependent DNA methylation of genes that are suppressed in stem cells is a hallmark of cancer. Genome Res. 2010, 20, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteller, M.; Silva, J.M.; Dominguez, G.; Bonilla, F.; Matias-Guiu, X.; Lerma, E.; Bussaglia, E.; Prat, J.; Harkes, I.C.; Repasky, E.A. Promoter hypermethylation and BRCA1 inactivation in sporadic breast and ovarian tumors. J. Natl. Cancer Inst. 2000, 92, 564–569. [Google Scholar] [CrossRef]
- Dobrovic, A.; Simpfendorfer, D. Methylation of the BRCA1 gene in sporadic breast cancer. Cancer Res. 1997, 57, 3347–3350. [Google Scholar]
- Turner, N.C.; Reis-Filho, J.S.; Russell, A.M.; Springall, R.J.; Ryder, K.; Steele, D.; Savage, K.; Gillett, C.E.; Schmitt, F.C.; Ashworth, A. BRCA1 dysfunction in sporadic basal-like breast cancer. Oncogene 2007, 26, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, D.; Easton, D.F.; Stratton, M.; Narod, S.; Goldgar, D.; Devilee, P.; Bishop, D.T.; Weber, B.; Lenoir, G.; Chang-Claude, J. Genetic heterogeneity and penetrance analysis of the BRCA1 and BRCA2 genes in breast cancer families. Am. J. Hum. Genet. 1998, 62, 676–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easton, D.F.; Ford, D.; Bishop, D.T. Breast and ovarian cancer incidence in BRCA1-mutation carriers. Breast Cancer Linkage Consortium. Am. J. Hum. Genet. 1995, 56, 265. [Google Scholar]
- Chopra-Tandon, N.; Wu, H.; Arcaro, K.F.; Sturgeon, S.R. Relationships between global DNA methylation in circulating white blood cells and breast cancer risk factors. J. Cancer Epidemiol. 2017, 2017, e2705860. [Google Scholar] [CrossRef] [Green Version]
- Jabłońska, E.; Reszka, E. Selenium and Epigenetics in Cancer: Focus on DNA Methylation. In Advances in Cancer Research; Academic Press Inc.: Cambridge, MA, USA, 2017; pp. 193–234. [Google Scholar]
- Van Dijk, S.J.; Molloy, P.L.; Varinli, H.; Morrison, J.L.; Muhlhausler, B.S.; Buckley, M.; Clark, S.J.; McMillen, I.C.; Noakes, M.; Samaras, K.; et al. Epigenetics and human obesity. Int. J. Obes. 2015, 39, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Bewerunge-Hudler, M.; Schick, M.; Burwinkel, B.; Herpel, E.; Hoffmeister, M.; Brenner, H. Blood-derived DNA methylation predictors of mortality discriminate tumor and healthy tissue in multiple organs. Mol. Oncol. 2020, 14, 2111–2123. [Google Scholar] [CrossRef]
- Yuasa, Y.; Nagasaki, H.; Akiyama, Y.; Hashimoto, Y.; Takizawa, T.; Kojima, K.; Kawano, T.; Sugihara, K.; Imai, K.; Nakachi, K. DNA methylation status is inversely correlated with green tea intake and physical activity in gastric cancer patients. Int. J. Cancer 2009, 124, 2677–2682. [Google Scholar] [CrossRef]
- Troiano, R.P.; Berrigan, D.; Dodd, K.W.; Masse, L.C.; Tilert, T.; McDowell, M. Physical Activity in the United States Measured by Accelerometer. Med. Sci. Sport Exerc. 2008, 40, 181–188. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
|
|
| Characteristic | Completed Trial (n = 135) M (SD) | Enrolled, Not Completed (n = 141) M (SD) | Test Statistic for Group Differences |
|---|---|---|---|
| Age | 37.43 (4.71) | 37.08 (4.59) | t(267) = 0.60), p = 0.545 |
| Race (% white) | 59.7% | 51.5% | χ2(6) = 6.69, p = 0.035 |
| Education (% college degree or higher) | 71.6% | 54.1% | χ2(7) = 14.13, p = 0.05 |
| BMI (kg/m2) | 28.54 (5.62) | 29.38 (5.15) | t(261) = −1.25, p = 0.211 |
| BaselineVO2max (mL/kg/min) | 27.95 (5.80) | 26.46 (4.80) | t(229) = 2.08, p = 0.04 |
| Self-reported exercise minutes/week | 17.13 (32.61) | 15.56 (25.02) | t(266) = 0.441, p = 0.66 |
| Characteristic | Overall Sample (N = 81) | Low Intensity + 20 min (n = 21) | Low Intensity + 40 min (n = 23) | High Intensity + 20 min (n = 17) | High Intensity + 40 min (n = 20) |
|---|---|---|---|---|---|
| Age | 37.14 (4.71) | 37.00 (4.86) | 37.26 (4.60) | 37.82 (5.45) | 33.55 (4.29) |
| Race (% White) | 65.4% | 61.9% | 65.2% | 70.6% | 65.0% |
| BMI (kg/m2) | 29.60 (5.69) | 28.22 (6.02) | 29.36 (4.95) | 29.59 (5.69) | 31.32 (6.08) |
| VO2max (mL/kg/min) | 27.37 (5.29) | 28.18 (5.73) | 26.78 (5.38) | 27.70 (5.41) | 26.89 (4.85) |
| Self-reported exercise mins/week | 18.27 (31.94) | 12.86 (15.13) | 22.85 (48.14) | 17.65 (22.23) | 19.25 (29.92) |
| Gene | Gene Location of Methylation Profile | Region of Methylation Profile from Transcriptional Start Site (Number of Base Pairs) |
|---|---|---|
| BRCA1 | 5′ Untranslated Region | −76 to +23 |
| RUNX3 | 5′ Untranslated Region | −12 to +76 |
| SIM1 | 5′ Untranslated Region | +3 to +37 |
| AURKA | 5′ Untranslated Region | −1276 to −1241 |
| BCAR1 | 5′ Untranslated Region | −503 to −482 |
| BPIFA4P | 5′ Untranslated Region | −529 |
| GALNT9 | Intron 1 | +41,806 to +41,854 |
| FBLN2 | Intron 1 | +263 to +268 |
| PAX6 | Intron 4 | +12,543 to +12,609 |
| TLR4 | 5′ Untranslated Region | +27 to +51 |
| TLR6 | 5′ Untranslated Region | −1291 to −1269 |
| Variable | Coefficient | Std. Error | T | p |
|---|---|---|---|---|
| (Intercept) | 0.189 | 0.090 | 2.08 | 0.041 * |
| Baseline methylation | 0.974 | 0.050 | 19.60 | <0.001 |
| Age (centered) | 0.026 | 0.015 | 1.78 | 0.080 |
| BMI (centered) | −0.008 | 0.012 | −0.69 | 0.491 |
| Change in VO2max | −0.054 | 0.021 | −2.52 | 0.014 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gillman, A.S.; Helmuth, T.; Koljack, C.E.; Hutchison, K.E.; Kohrt, W.M.; Bryan, A.D. The Effects of Exercise Duration and Intensity on Breast Cancer-Related DNA Methylation: A Randomized Controlled Trial. Cancers 2021, 13, 4128. https://doi.org/10.3390/cancers13164128
Gillman AS, Helmuth T, Koljack CE, Hutchison KE, Kohrt WM, Bryan AD. The Effects of Exercise Duration and Intensity on Breast Cancer-Related DNA Methylation: A Randomized Controlled Trial. Cancers. 2021; 13(16):4128. https://doi.org/10.3390/cancers13164128
Chicago/Turabian StyleGillman, Arielle S., Timothy Helmuth, Claire E. Koljack, Kent E. Hutchison, Wendy M. Kohrt, and Angela D. Bryan. 2021. "The Effects of Exercise Duration and Intensity on Breast Cancer-Related DNA Methylation: A Randomized Controlled Trial" Cancers 13, no. 16: 4128. https://doi.org/10.3390/cancers13164128
APA StyleGillman, A. S., Helmuth, T., Koljack, C. E., Hutchison, K. E., Kohrt, W. M., & Bryan, A. D. (2021). The Effects of Exercise Duration and Intensity on Breast Cancer-Related DNA Methylation: A Randomized Controlled Trial. Cancers, 13(16), 4128. https://doi.org/10.3390/cancers13164128