
The Emerging Influences of Alpha-Fetoprotein in the Tumorigenesis and Progression of Hepatocellular Carcinoma




Abstract
:Simple Summary
Abstract
1. Introduction
2. Biological Characteristics of AFP and Potential Biomarkers in Diagnosis of HCC
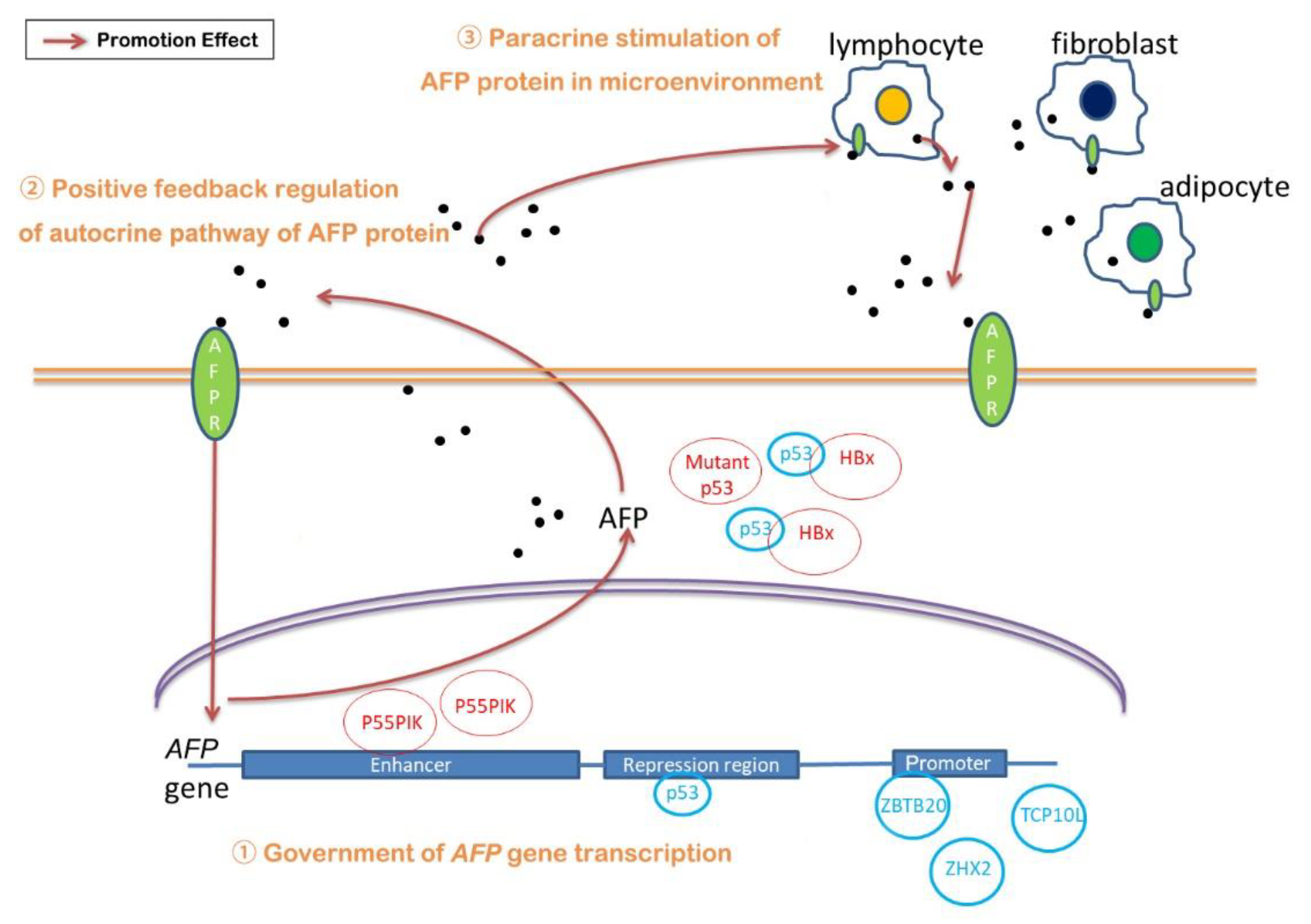
3. Regulatory Mechanisms of AFP Expression
3.1. The Government of AFP Gene Transcription
3.2. Regulation of AFP/Receptor Autocrine and Paracrine Pathway
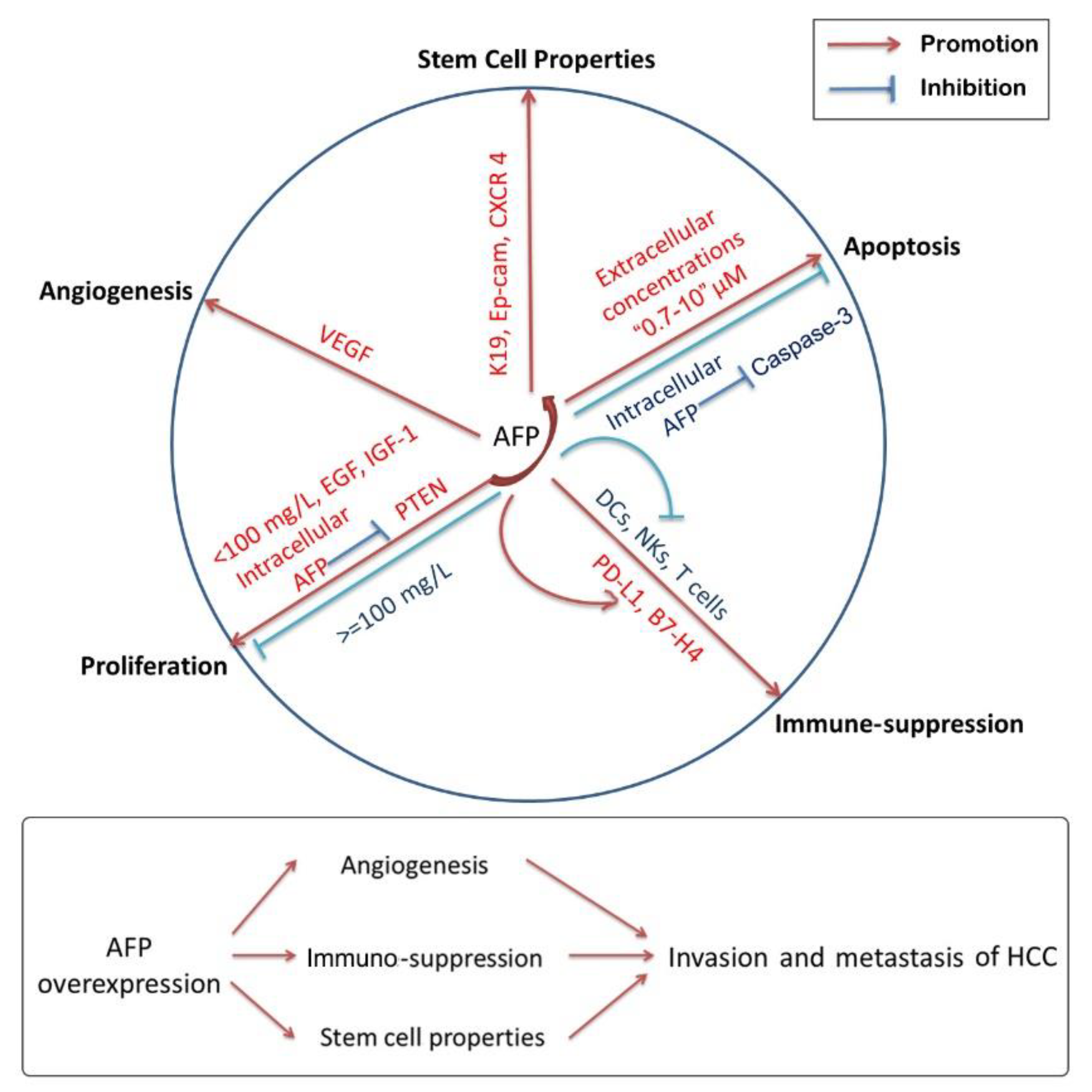
4. Effects of AFP on the Tumorigenesis and Progression of HCC
4.1. AFP and Cell Proliferation of HCC
4.2. AFP and Cancer Cell Apoptosis
4.3. Immuno-Suppressive Activity of AFP
4.4. AFP and Invasion, Metastasis and Angiogenesis of HCC
5. Summary
6. Prospect of AFP-Targeted Therapies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, K.; Jiang, Y.; Cai, N.; Fan, J.; Mao, X.; Suo, C.; Jin, L.; Zhang, T.; Chen, X. Global trend of aetiology-based primary liver cancer incidence from 1990 to 2030: A modelling study. Int. J. Epidemiol. 2021, 50, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Choo, S.P.; Tan, W.L.; Goh, B.; Tai, W.M.; Zhu, A.X. Comparison of hepatocellular carcinoma in Eastern versus Western populations. Cancer 2016, 122, 3430–3446. [Google Scholar] [CrossRef]
- Guideline for stratified screening and surveillance of primary liver cancer(2020 Edition). Zhonghua Zhong Liu Za Zhi 2021, 43, 60–77. [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Marengo, A.; Rosso, C.; Bugianesi, E. Liver Cancer: Connections with Obesity, Fatty Liver, and Cirrhosis. Annu. Rev. Med. 2016, 67, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Vanni, E.; Bugianesi, E. Obesity and liver cancer. Clin. Liver Dis. 2014, 18, 191–203. [Google Scholar] [CrossRef]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1343–1351. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, H.; Takaki, A. Alcohol and hepatocellular carcinoma. BMJ Open Gastroenterol. 2019, 6, e000260. [Google Scholar] [CrossRef]
- Taniai, M. Alcohol and hepatocarcinogenesis. Clin. Mol. Hepatol. 2020, 26, 736–741. [Google Scholar] [CrossRef]
- Tsochatzis, E.; Meyer, T.; O’Beirne, J.; Burroughs, A.K. Transarterial chemoembolisation is not superior to embolisation alone: The recent European Association for the Study of the Liver (EASL) European Organisation for Research and Treatment of Cancer (EORTC) guidelines. Eur. J. Cancer 2013, 49, 1509–1510. [Google Scholar] [CrossRef]
- Bruix, J.; Sherman, M.; American Association for the Study of Liver Diseases. Management of hepatocellular carcinoma: An update. Hepatology 2011, 53, 1020–1022. [Google Scholar] [CrossRef]
- Shiina, S.; Tateishi, R.; Arano, T.; Uchino, K.; Enooku, K.; Nakagawa, H.; Asaoka, Y.; Sato, T.; Masuzaki, R.; Kondo, Y.; et al. Radiofrequency ablation for hepatocellular carcinoma: 10-year outcome and prognostic factors. Am. J. Gastroenterol. 2012, 107, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Shiina, S.; Tateishi, R.; Imamura, M.; Teratani, T.; Koike, Y.; Sato, S.; Obi, S.; Kanai, F.; Kato, N.; Yoshida, H.; et al. Percutaneous ethanol injection for hepatocellular carcinoma: 20-year outcome and prognostic factors. Liver Int. 2012, 32, 1434–1442. [Google Scholar] [CrossRef] [Green Version]
- Vogel, A.; Cervantes, A.; Chau, I.; Daniele, B.; Llovet, J.M.; Meyer, T.; Nault, J.C.; Neumann, U.; Ricke, J.; Sangro, B.; et al. Hepatocellular carcinoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2018, 29, iv238–iv255. [Google Scholar] [CrossRef]
- El-Serag, H.B.; Marrero, J.A.; Rudolph, L.; Reddy, K.R. Diagnosis and treatment of hepatocellular carcinoma. Gastroenterology 2008, 134, 1752–1763. [Google Scholar] [CrossRef] [Green Version]
- Anwanwan, D.; Singh, S.K.; Singh, S.; Saikam, V.; Singh, R. Challenges in liver cancer and possible treatment approaches. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188314. [Google Scholar] [CrossRef] [PubMed]
- Heimbach, J.K.; Kulik, L.M.; Finn, R.S.; Sirlin, C.B.; Abecassis, M.M.; Roberts, L.R.; Zhu, A.X.; Murad, M.H.; Marrero, J.A. AASLD guidelines for the treatment of hepatocellular carcinoma. Hepatology 2018, 67, 358–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumada, T.; Nakano, S.; Takeda, I.; Kiriyama, S.; Sone, Y.; Hayashi, K.; Katoh, H.; Endoh, T.; Sassa, T.; Satomura, S. Clinical utility of Lens culinaris agglutinin-reactive alpha-fetoprotein in small hepatocellular carcinoma: Special reference to imaging diagnosis. J. Hepatol. 1999, 30, 125–130. [Google Scholar] [CrossRef]
- Butterfield, L.H.; Economou, J.S.; Gamblin, T.C.; Geller, D.A. Alpha fetoprotein DNA prime and adenovirus boost immunization of two hepatocellular cancer patients. J. Transl. Med. 2014, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Bai, D.S.; Zhang, C.; Chen, P.; Jin, S.J.; Jiang, G.Q. The prognostic correlation of AFP level at diagnosis with pathological grade, progression, and survival of patients with hepatocellular carcinoma. Sci. Rep. 2017, 7, 12870. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Liu, Y.; Shou, D.; Sun, Q.; Shi, J.; Chen, L.; Liang, T.; Gong, W. AFP (alpha fetoprotein): Who are you in gastrology? Cancer Lett. 2015, 357, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Mizejewski, G.J. The phylogeny of alpha-fetoprotein in vertebrates: Survey of biochemical and physiological data. Crit. Rev. Eukaryot Gene Expr. 1995, 5, 281–316. [Google Scholar] [CrossRef] [PubMed]
- Bergstrand, C.G.; Czar, B. Demonstration of a new protein fraction in serum from the human fetus. Scand. J. Clin. Lab. Investig. 1956, 8, 174. [Google Scholar] [CrossRef]
- Obiekwe, B.C.; Malek, N.; Kitau, M.J.; Chard, T. Maternal and fetal alphafetoprotein (AFP) levels at term. Relation to sex, weight and gestation of the infant. Acta Obstet. Gynecol. Scand. 1985, 64, 251–253. [Google Scholar] [CrossRef]
- Waller, D.K.; Lustig, L.S.; Cunningham, G.C.; Feuchtbaum, L.B.; Hook, E.B. The association between maternal serum alpha-fetoprotein and preterm birth, small for gestational age infants, preeclampsia, and placental complications. Obstet. Gynecol. 1996, 88, 816–822. [Google Scholar]
- Bader, D.; Riskin, A.; Vafsi, O.; Tamir, A.; Peskin, B.; Israel, N.; Merksamer, R.; Dar, H.; David, M. Alpha-fetoprotein in the early neonatal period-a large study and review of the literature. Clin. Chim. Acta 2004, 349, 15–23. [Google Scholar] [CrossRef]
- Ballas, M. Yolk sac carcinoma of the ovary with alpha fetoprotein in serum and ascitic fluid demonstrated by immunoosmophoresis. Am. J. Clin. Pathol. 1972, 57, 511–516. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, J.; Endo, Y.; Oda, T. Lectin affinities of alpha-fetoprotein in liver cirrhosis, hepatocellular carcinoma and metastatic liver tumor. Kanzo 1981, 22, 1559–1568. [Google Scholar] [CrossRef]
- Abelev, G.I.; Perova, S.D.; Khramkova, N.I.; Postnikova, Z.A.; Irlin, I.S. Production of embryonal alpha-globulin by transplantable mouse hepatomas. Transplantation 1963, 1, 174–180. [Google Scholar] [CrossRef]
- Tatarinov, Y.S. Content of embryo-specific alpha-globulin in fetal and neonatal sera and sera from adult humans with primary carcinoma of the liver. Fed. Proceedings. Transl. Suppl. Sel. Transl. Med.-Relat. Sci. 1966, 25, 344–346. [Google Scholar]
- Bellissimo, F.; Pinzone, M.R.; Cacopardo, B.; Nunnari, G. Diagnostic and therapeutic management of hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 12003–12021. [Google Scholar] [CrossRef]
- Tsuchiya, N.; Sawada, Y.; Endo, I.; Saito, K.; Uemura, Y.; Nakatsura, T. Biomarkers for the early diagnosis of hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 10573–10583. [Google Scholar] [CrossRef]
- Zong, J.; Fan, Z.; Zhang, Y. Serum Tumor Markers for Early Diagnosis of Primary Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2020, 7, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Amin, D.H.; Abdelkhalek, Y.I. Efficiency of whole-body 18F-FDG PET CT in detecting the cause of rising serum AFP level in post-therapeutic follow-up for HCC patients. Jpn. J. Radiol. 2020, 38, 472–479. [Google Scholar] [CrossRef]
- Qin, L.; Huang, H.; Huang, J.; Wang, G.; Huang, J.; Wu, X.; Li, J.; Yi, W.; Liu, L.; Huang, D. Biological characteristics of heat shock protein 90 in human liver cancer cells. Am. J. Transl. Res. 2019, 11, 2477–2483. [Google Scholar] [PubMed]
- Tang, Y.; Li, K.; Cai, Z.; Xie, Y.; Tan, X.; Su, C.; Li, J. HSP90α combined with AFP and TK1 improved the diagnostic value for hepatocellular carcinoma. Biomark. Med. 2020, 14, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, J.; Ju, L.; Chen, L.; Cai, W.; Yang, J. Diagnostic and prognostic potential of serum miR-132/212 cluster in patients with hepatocellular carcinoma. Ann. Clin. Biochem. 2018, 55, 576–582. [Google Scholar] [CrossRef]
- Yang, X.P.; Zhou, L.X.; Yang, Q.J.; Liu, L.; Cai, Y.; Ma, S.L. Diagnostic and prognostic roles of serum vitronectin in hepatitis B-related hepatocellular carcinoma. Cancer Biomark. 2016, 17, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Dong, X.; Chen, Y.; Wang, X. Serum exosomal hnRNPH1 mRNA as a novel marker for hepatocellular carcinoma. Clin. Chem. Lab. Med. 2018, 56, 479–484. [Google Scholar] [CrossRef]
- Ming, X.L.; Feng, Y.L.; He, D.D.; Luo, C.L.; Rong, J.L.; Zhang, W.W.; Ye, P.; Chai, H.Y.; Liang, C.Z.; Tu, J.C. Role of BCYRN1 in hepatocellular carcinoma pathogenesis by lncRNA-miRNA-mRNA network analysis and its diagnostic and prognostic value. Epigenomics 2019, 11, 1209–1231. [Google Scholar] [CrossRef]
- Bakr, N.M.; Awad, A.; Moustafa, E.A. Association of genetic variants in the interleukin-18 gene promoter with risk of hepatocellular carcinoma and metastasis in patients with hepatitis C virus infection. IUBMB Life 2018, 70, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Fan, Y.C.; Gao, S.; Zhao, J.; Chen, L.Y.; Li, F.; Wang, K. Methylation of SOX1 and VIM promoters in serum as potential biomarkers for hepatocellular carcinoma. Neoplasma 2017, 64, 745–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.Y.; Jiang, J.H.; Jin, R.M.; Jin, G.Z.; Jia, N.Y. The clinical significance of lipopolysaccharide binding protein in hepatocellular carcinoma. Oncol. Lett. 2020, 19, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.P.; Fan, X.P.; Fan, Y.C.; Zhao, J.; Gao, S.; Wang, K. Combined detection of insulin-like growth factor-binding protein 7 promoter methylation improves the diagnostic efficacy of AFP in hepatitis B virus-associated hepatocellular carcinoma. Pathol. Res. Pract. 2018, 214, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Fujiyama, S.; Morishita, T.; Hashiguchi, O.; Sato, T. Plasma abnormal prothrombin (des-gamma-carboxy prothrombin) as a marker of hepatocellular carcinoma. Cancer 1988, 61, 1621–1628. [Google Scholar] [CrossRef]
- Ikoma, J.; Kaito, M.; Ishihara, T.; Nakagawa, N.; Kamei, A.; Fujita, N.; Iwasa, M.; Tamaki, S.; Watanabe, S.; Adachi, Y. Early diagnosis of hepatocellular carcinoma using a sensitive assay for serum des-gamma-carboxy prothrombin: A prospective study. Hepatogastroenterology 2002, 49, 235–238. [Google Scholar] [PubMed]
- Inagaki, Y.; Tang, W.; Makuuchi, M.; Hasegawa, K.; Sugawara, Y.; Kokudo, N. Clinical and molecular insights into the hepatocellular carcinoma tumour marker des-γ-carboxyprothrombin. Liver. Int. 2011, 31, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Ishii, M.; Gama, H.; Chida, N.; Ueno, Y.; Shinzawa, H.; Takagi, T.; Toyota, T.; Takahashi, T.; Kasukawa, R. Simultaneous measurements of serum alpha-fetoprotein and protein induced by vitamin K absence for detecting hepatocellular carcinoma. South. Tohoku District Study Group. Am. J. Gastroenterol. 2000, 95, 1036–1040. [Google Scholar] [PubMed]
- Shimauchi, Y.; Tanaka, M.; Kuromatsu, R.; Ogata, R.; Tateishi, Y.; Itano, S.; Ono, N.; Yutani, S.; Nagamatsu, H.; Matsugaki, S.; et al. A simultaneous monitoring of Lens culinaris agglutinin A-reactive alpha-fetoprotein and des-gamma-carboxy prothrombin as an early diagnosis of hepatocellular carcinoma in the follow-up of cirrhotic patients. Oncol. Rep. 2000, 7, 249–256. [Google Scholar] [CrossRef]
- Durazo, F.A.; Blatt, L.M.; Corey, W.G.; Lin, J.H.; Han, S.; Saab, S.; Busuttil, R.W.; Tong, M.J. Des.-gamma-carboxyprothrombin, alpha-fetoprotein and AFP-L3 in patients with chronic hepatitis, cirrhosis and hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2008, 23, 1541–1548. [Google Scholar] [CrossRef]
- Johnson, P.; Berhane, S.; Kagebayashi, C.; Satomura, S.; Teng, M.; Fox, R.; Yeo, W.; Mo, F.; Lai, P.; Chan, S.L.; et al. Impact of disease stage and aetiology on survival in hepatocellular carcinoma: Implications for surveillance. Br. J. Cancer 2017, 116, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Singal, A.G.; El-Serag, H.B. Hepatocellular Carcinoma From Epidemiology to Prevention: Translating Knowledge into Practice. Clin. Gastroenterol. Hepatol. 2015, 13, 2140–2151. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Li, H.Y.; Zhang, L.; Zhou, Y.; Wu, J. Hedgehog Signaling, a Critical Pathway Governing the Development and Progression of Hepatocellular Carcinoma. Cells 2021, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Jiang, M.; Zhao, Y.; Wang, B.L.; Shen, M.N.; Zhou, Y.; Zhang, C.Y.; Sun, Y.F.; Chen, J.W.; Hu, B.; et al. Application of Serum Annexin A3 in Diagnosis, Outcome Prediction and Therapeutic Response Evaluation for Patients with Hepatocellular Carcinoma. Ann. Surg. Oncol. 2018, 25, 1686–1694. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.M.; Berhane, S.; de Groot, C.A.E.; Toyoda, H.; Tada, T.; Kumada, T.; Satomura, S.; Nishida, N.; Kudo, M.; Kimura, T.; et al. Serum Levels of α-Fetoprotein Increased More Than 10 Years Before Detection of Hepatocellular Carcinoma. Clin. Gastroenterol. Hepatol. 2021, 19, 162.e4–170.e4. [Google Scholar] [CrossRef] [PubMed]
- Vacher, J.; Tilghman, S.M. Dominant negative regulation of the mouse alpha-fetoprotein gene in adult liver. Science 1990, 250, 1732–1735. [Google Scholar] [CrossRef]
- Lazarevich, N.L. Molecular mechanisms of alpha-fetoprotein gene expression. Biochemistry 2000, 65, 117–133. [Google Scholar]
- Jeon, Y.; Choi, Y.S.; Jang, E.S.; Kim, J.W.; Jeong, S.H. Persistent α-Fetoprotein Elevation in Healthy Adults and Mutational Analysis of α-Fetoprotein Promoter, Enhancer, and Silencer Regions. Gut. Liver. 2017, 11, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata-Tsubouchi, Y.; Ido, A.; Uto, H.; Numata, M.; Moriuchi, A.; Kim, I.; Hasuike, S.; Nagata, K.; Sekiya, T.; Hayashi, K.; et al. Molecular mechanisms of hereditary persistence of alpha-fetoprotein (AFP) in two Japanese families A hepatocyte nuclear factor-1 site mutation leads to induction of the AFP gene expression in adult livers. Hepatol. Res. 2005, 31, 79–87. [Google Scholar] [CrossRef]
- Montal, R.; Andreu-Oller, C.; Bassaganyas, L.; Esteban-Fabró, R.; Moran, S.; Montironi, C.; Moeini, A.; Pinyol, R.; Peix, J.; Cabellos, L.; et al. Molecular portrait of high alpha-fetoprotein in hepatocellular carcinoma: Implications for biomarker-driven clinical trials. Br. J. Cancer 2019, 121, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Cao, D.; Zhou, L.; Zhang, Y.; Guo, X.; Li, H.; Chen, Y.; Spear, B.T.; Wu, J.W.; Xie, Z.; et al. ZBTB20 is a sequence-specific transcriptional repressor of alpha-fetoprotein gene. Sci. Rep. 2015, 5, 11979. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Zhang, H.; Tsai, W.; Zhang, Y.; Du, Y.; Zhong, J.; Szpirer, C.; Zhu, M.; Cao, X.; Barton, M.C.; et al. Zinc finger protein ZBTB20 is a key repressor of alpha-fetoprotein gene transcription in liver. Proc. Natl. Acad. Sci. USA 2008, 105, 10859–10864. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Shi, J.H.; Jiang, H.; Wang, K.; Lu, J.Y.; Jiang, X.; Ma, X.; Chen, Y.X.; Ren, A.J.; Zheng, J.; et al. ZBTB20 regulates EGFR expression and hepatocyte proliferation in mouse liver regeneration. Cell Death Dis. 2018, 9, 462. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tan, Y.X.; Ren, Y.B.; Dong, L.W.; Xie, Z.F.; Tang, L.; Cao, D.; Zhang, W.P.; Hu, H.P.; Wang, H.Y. Zinc finger protein ZBTB20 expression is increased in hepatocellular carcinoma and associated with poor prognosis. BMC Cancer 2011, 11, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, H.; Huang, Y.; Li, X.; Liu, D.; Chen, J.; Shu, M. Zinc finger protein ZBTB20 is an independent prognostic marker and promotes tumor growth of human hepatocellular carcinoma by repressing FoxO1. Oncotarget 2016, 7, 14336–14349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Luan, F.; Liu, H.; Gao, L.; Liang, X.; Zhang, L.; Sun, W.; Ma, C. ZHX2 is a repressor of alpha-fetoprotein expression in human hepatoma cell lines. J. Cell Mol. Med. 2008, 12, 2772–2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchio, A.; Bertani, S.; Rojas Rojas, T.; Doimi, F.; Terris, B.; Deharo, E.; Dejean, A.; Ruiz, E.; Pineau, P. A peculiar mutation spectrum emerging from young peruvian patients with hepatocellular carcinoma. PLoS ONE 2014, 9, e114912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Feng, H.; Liu, L.; Su, W.; Yu, L.; Wu, J. TCP10L negatively regulates alpha-fetoprotein expression in hepatocellular carcinoma. BMB Rep. 2020, 53, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Ogden, S.K.; Lee, K.C.; Wernke-Dollries, K.; Stratton, S.A.; Aronow, B.; Barton, M.C. p53 targets chromatin structure alteration to repress alpha-fetoprotein gene expression. J. Biol. Chem. 2001, 276, 42057–42062. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Cho, K.; Stratton, S.A.; Barton, M.C. Transcription factor interactions and chromatin modifications associated with p53-mediated, developmental repression of the alpha-fetoprotein gene. Mol. Cell Biol. 2005, 25, 2147–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.Y.; Chen, W.J.; Lai, P.L.; Jeng, Y.M.; Sheu, J.C.; Hsu, H.C. High alpha-fetoprotein level correlates with high stage, early recurrence and poor prognosis of hepatocellular carcinoma: Significance of hepatitis virus infection, age, p53 and beta-catenin mutations. Int. J. Cancer 2004, 112, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Ogden, S.K.; Lee, K.C.; Barton, M.C. Hepatitis B viral transactivator HBx alleviates p53-mediated repression of alpha-fetoprotein gene expression. J. Biol. Chem. 2000, 275, 27806–27814. [Google Scholar] [CrossRef] [Green Version]
- Ye, G.; Sun, G.; Cheng, Z.; Zhang, L.; Hu, K.; Xia, X.; Zhou, Y. p55PIK regulates alpha-fetoprotein expression through the NF-κB signaling pathway. Life Sci. 2017, 191, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Li, M.S.; Li, P.F.; He, S.P.; Du, G.G.; Li, G. The promoting molecular mechanism of alpha-fetoprotein on the growth of human hepatoma Bel7402 cell line. World J. Gastroenterol. 2002, 8, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhu, M.; Li, W.; Lu, Y.; Xie, X.; Wu, Y.; Zheng, S. Alpha-fetoprotein receptor as an early indicator of HBx-driven hepatocarcinogenesis and its applications in tracing cancer cell metastasis. Cancer Lett. 2013, 330, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Esteban, C.; Terrier, P.; Frayssinet, C.; Uriel, J. Expression of the alpha-fetoprotein gene in human breast cancer. Tumour. Biol. 1996, 17, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Ogunwobi, O.O.; Harricharran, T.; Huaman, J.; Galuza, A.; Odumuwagun, O.; Tan, Y.; Ma, G.X.; Nguyen, M.T. Mechanisms of hepatocellular carcinoma progression. World J. Gastroenterol. 2019, 25, 2279–2293. [Google Scholar] [CrossRef]
- Toder, V.; Blank, M.; Gold-Gefter, L.; Nebel, L. The effect of alpha-fetoprotein on the growth of placental cells in vitro. Placenta 1983, 4, 79–86. [Google Scholar] [CrossRef]
- Semenkova, L.N.; Dudich, E.I.; Dudich, I.V. Induction of apoptosis in human hepatoma cells by alpha-fetoprotein. Tumour. Biol. 1997, 18, 261–273. [Google Scholar] [CrossRef]
- Li, M.S.; Li, P.F.; Chen, Q.; Du, G.G.; Li, G. Alpha-fetoprotein stimulated the expression of some oncogenes in human hepatocellular carcinoma Bel 7402 cells. World J. Gastroenterol. 2004, 10, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Sporn, M.B.; Roberts, A.B. Peptide growth factors are multifunctional. Nature 1988, 332, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Dudich, E.; Semenkova, L.; Gorbatova, E.; Dudich, I.; Khromykh, L.; Tatulov, E.; Grechko, G.; Sukhikh, G. Growth-regulative activity of human alpha-fetoprotein for different types of tumor and normal cells. Tumour. Biol. 1998, 19, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Keel, B.A.; Eddy, K.B.; Cho, S.; May, J.V. Synergistic action of purified alpha-fetoprotein and growth factors on the proliferation of porcine granulosa cells in monolayer culture. Endocrinology 1991, 129, 217–225. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Li, C.; Wang, S.; Jiang, W.; Liu, Z.; Zhou, S.; Liu, X.; McNutt, M.A.; Li, G. Alpha-fetoprotein: A new member of intracellular signal molecules in regulation of the PI3K/AKT signaling in human hepatoma cell lines. Int. J. Cancer 2011, 128, 524–532. [Google Scholar] [CrossRef]
- Zhu, M.; Lin, B.; Zhou, P.; Li, M. Molecular Analysis of AFP and HSA Interactions with PTEN Protein. Biomed. Res. Int. 2015, 2015, 256916. [Google Scholar] [CrossRef] [PubMed]
- Dudich, E.; Semenkova, L.; Dudich, I.; Gorbatova, E.; Tochtamisheva, N.; Tatulov, E.; Nikolaeva, M.; Sukhikh, G. alpha-fetoprotein causes apoptosis in tumor cells via a pathway independent of CD95, TNFR1 and TNFR2 through activation of caspase-3-like proteases. Eur. J. Biochem. 1999, 266, 750–761. [Google Scholar] [CrossRef]
- Semenkova, L.N.; Dudich, E.I.; Dudich, I.V.; Shingarova, L.N.; Korobko, V.G. Alpha-fetoprotein as a TNF resistance factor for the human hepatocarcinoma cell line HepG2. Tumour. Biol. 1997, 18, 30–40. [Google Scholar] [CrossRef]
- Semenkova, L.; Dudich, E.; Dudich, I.; Tokhtamisheva, N.; Tatulov, E.; Okruzhnov, Y.; Garcia-Foncillas, J.; Palop-Cubillo, J.A.; Korpela, T. Alpha-fetoprotein positively regulates cytochrome c-mediated caspase activation and apoptosome complex formation. Eur. J. Biochem. 2003, 270, 4388–4399. [Google Scholar] [CrossRef]
- Dudich, E.; Semenkova, L.; Dudich, I.; Denesyuk, A.; Tatulov, E.; Korpela, T. Alpha-fetoprotein antagonizes X-linked inhibitor of apoptosis protein anticaspase activity and disrupts XIAP-caspase interaction. FEBS J. 2006, 273, 3837–3849. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Y.; Zhang, L.; Zhang, L.; Mao, J. Silencing alpha-fetoprotein expression induces growth arrest and apoptosis in human hepatocellular cancer cell. Cancer Lett. 2008, 271, 281–293. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Li, C.; Zhou, S.; Guo, L.; Liu, H.; Jiang, W.; Liu, X.; Li, P.; McNutt, M.A.; et al. Alpha fetoprotein is a novel protein-binding partner for caspase-3 and blocks the apoptotic signaling pathway in human hepatoma cells. Int. J. Cancer 2009, 124, 2845–2854. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Zhu, M.; Wang, W.; Li, W.; Dong, X.; Chen, Y.; Lu, Y.; Guo, J.; Li, M. Structural basis for alpha fetoprotein-mediated inhibition of caspase-3 activity in hepatocellular carcinoma cells. Int. J. Cancer 2017, 141, 1413–1421. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, S.; Liu, X.; Li, P.; McNutt, M.A.; Li, G. alpha-Fetoprotein shields hepatocellular carcinoma cells from apoptosis induced by tumor necrosis factor-related apoptosis-inducing ligand. Cancer Lett. 2007, 249, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Crainie, M.; Semeluk, A.; Lee, K.C.; Wegmann, T. Regulation of constitutive and lymphokine-induced Ia expression by murine alpha-fetoprotein. Cell. Immunol. 1989, 118, 41–52. [Google Scholar] [CrossRef]
- Nicholas, N.S.; Panayi, G.S. Immunosuppressive properties of pregnancy serum on the mixed lymphocyte reaction. Br. J. Obstet. Gynaecol. 1986, 93, 1251–1255. [Google Scholar] [CrossRef]
- Um, S.H.; Mulhall, C.; Alisa, A.; Ives, A.R.; Karani, J.; Williams, R.; Bertoletti, A.; Behboudi, S. Alpha-fetoprotein impairs APC function and induces their apoptosis. J. Immunol. 2004, 173, 1772–1778. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; Zhou, S.; Li, P.; Li, G. Effects of alpha fetoprotein on escape of Bel 7402 cells from attack of lymphocytes. BMC Cancer 2005, 5, 96. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.T.; Qiu, M.J.; Yang, S.L.; Fang, X.; He, X.X.; Wang, M.M.; Li, Y.N.; Xiong, Z.F.; Huang, S. Alpha-Fetoprotein Regulates the Expression of Immune-Related Proteins through the NF-κB (P65) Pathway in Hepatocellular Carcinoma Cells. J. Oncol. 2020, 2020, 9327512. [Google Scholar] [CrossRef]
- Zahran, A.M.; Nafady-Hego, H.; Mansor, S.G.; Abbas, W.A.; Abdel-Malek, M.O.; Mekky, M.A.; Hetta, H.F. Increased frequency and FOXP3 expression of human CD8(+)CD25(High.+) T lymphocytes and its relation to CD4 regulatory T cells in patients with hepatocellular carcinoma. Hum. Immunol. 2019, 80, 510–516. [Google Scholar] [CrossRef]
- Esteban, C.; Trojan, J.; Macho, A.; Mishal, Z.; Lafarge-Frayssinet, C.; Uriel, J. Activation of an alpha-fetoprotein/receptor pathway in human normal and malignant peripheral blood mononuclear cells. Leukemia 1993, 7, 1807–1816. [Google Scholar] [PubMed]
- Zhou, S.; Venkatramani, R.; Gupta, S.; Wang, K.; Stein, J.E.; Wang, L.; Mascarenhas, L. Hepatocellular malignant neoplasm, NOS: A clinicopathological study of 11 cases from a single institution. Histopathology 2017, 71, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Uchino, K.; Tateishi, R.; Shiina, S.; Kanda, M.; Masuzaki, R.; Kondo, Y.; Goto, T.; Omata, M.; Yoshida, H.; Koike, K. Hepatocellular carcinoma with extrahepatic metastasis: Clinical features and prognostic factors. Cancer 2011, 117, 4475–4483. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.J.; Kim, K.M.; Jin, Y.J.; Shim, J.H.; Ko, G.Y.; Yoon, H.K.; Sung, K.B.; Lee, J.L.; Kang, Y.K.; Lim, Y.S.; et al. Clinical outcome of 251 patients with extrahepatic metastasis at initial diagnosis of hepatocellular carcinoma: Does transarterial chemoembolization improve survival in these patients? J. Gastroenterol. Hepatol. 2011, 26, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Katyal, S.; Oliver, J.H., 3rd; Peterson, M.S.; Ferris, J.V.; Carr, B.S.; Baron, R.L. Extrahepatic metastases of hepatocellular carcinoma. Radiology 2000, 216, 698–703. [Google Scholar] [CrossRef]
- Yokoo, T.; Patel, A.D.; Lev-Cohain, N.; Singal, A.G.; Yopp, A.C.; Pedrosa, I. Extrahepatic metastasis risk of hepatocellular carcinoma based on α-fetoprotein and tumor staging parameters at cross-sectional imaging. Cancer Manag. Res. 2017, 9, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S. Metastatic Recurrent Hepatocellular Carcinoma Post Liver Transplant. With Marked Pretransplant Elevation of Alpha Fetoprotein and No Evidence of Primary Neoplasm. Exp. Clin. Transplant. 2018, 16, 99–102. [Google Scholar]
- Hameed, B.; Mehta, N.; Sapisochin, G.; Roberts, J.P.; Yao, F.Y. Alpha-fetoprotein level >1000 ng/mL as an exclusion criterion for liver transplantation in patients with hepatocellular carcinoma meeting the Milan criteria. Liver Transpl. 2014, 20, 945–951. [Google Scholar] [CrossRef]
- Jin, J.; Niu, X.; Zou, L.; Li, L.; Li, S.; Han, J.; Zhang, P.; Song, J.; Xiao, F. AFP mRNA level in enriched circulating tumor cells from hepatocellular carcinoma patient blood samples is a pivotal predictive marker for metastasis. Cancer Lett. 2016, 378, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zhu, M.; Li, W.; Lin, B.; Dong, X.; Chen, Y.; Xie, X.; Guo, J.; Li, M. Alpha fetoprotein plays a critical role in promoting metastasis of hepatocellular carcinoma cells. J. Cell. Mol. Med. 2016, 20, 549–558. [Google Scholar] [CrossRef]
- Li, Y.; Tang, Z.Y.; Ye, S.L.; Liu, Y.K.; Chen, J.; Xue, Q.; Chen, J.; Gao, D.M.; Bao, W.H. Establishment of cell clones with different metastatic potential from the metastatic hepatocellular carcinoma cell line MHCC97. World J. Gastroenterol. 2001, 7, 630–636. [Google Scholar] [CrossRef]
- Koide, N.; Nishio, A.; Igarashi, J.; Kajikawa, S.; Adachi, W.; Amano, J. Alpha-fetoprotein-producing gastric cancer: Histochemical analysis of cell proliferation, apoptosis, and angiogenesis. Am. J. Gastroenterol. 1999, 94, 1658–1663. [Google Scholar] [CrossRef]
- Sheppard, H.W., Jr.; Sell, S.; Trefts, P.; Bahu, R. Effects of alpha-fetoprotein on murine immune responses. I. Studies on mice. J. Immunol. 1977, 119, 91–97. [Google Scholar]
- Lutsenko, S.V.; Feldman, N.B.; Finakova, G.V.; Gukasova, N.V.; Petukhov, S.P.; Posypanova, G.A.; Skryabin, K.G.; Severin, S.E. Antitumor activity of alpha fetoprotein and epidermal growth factor conjugates in vitro and in vivo. Tumour. Biol. 2000, 21, 367–374. [Google Scholar] [CrossRef]
- Sotnichenko, A.I.; Severin, S.E.; Posypanova, G.A.; Feldman, N.B.; Grigor’ev, M.I.; Severin, E.S.; Petrov, R.V. Water-soluble 2,3,7,8-tetrachlorodibenzo-p-dioxin complex with human alpha-fetoprotein: Properties, toxicity in vivo and antitumor activity in vitro. FEBS Lett. 1999, 450, 49–51. [Google Scholar] [CrossRef] [Green Version]
- Moskaleva, E.Y.; Posypanova, G.A.; Shmyrev, I.I.; Rodina, A.V.; Muizhnek, E.L.; Severin, E.S.; Katukov, V.Y.; Luzhkov, Y.M.; Severin, S.E. Alpha-fetoprotein-mediated targeting-a new strategy to overcome multidrug resistance of tumour cells in vitro. Cell Biol. Int. 1997, 21, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Ido, A.; Nakata, K.; Kato, Y.; Nakao, K.; Murata, K.; Fujita, M.; Ishii, N.; Tamaoki, T.; Shiku, H.; Nagataki, S. Gene therapy for hepatoma cells using a retrovirus vector carrying herpes simplex virus thymidine kinase gene under the control of human alpha-fetoprotein gene promoter. Cancer Res. 1995, 55, 3105–3109. [Google Scholar] [CrossRef]
- Zhu, W.; Peng, Y.; Wang, L.; Hong, Y.; Jiang, X.; Li, Q.; Liu, H.; Huang, L.; Wu, J.; Celis, E.; et al. Identification of α-fetoprotein-specific T-cell receptors for hepatocellular carcinoma immunotherapy. Hepatology 2018, 68, 574–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Caraballo Galva, L.D.; Peng, Y.; Luo, X.; Zhu, W.; Yao, Y.; Ji, Y.; He, Y. Preclinical Studies of the Off.-Target. Reactivity of AFP(158)-Specific TCR Engineered T Cells. Front. Immunol. 2020, 11, 607. [Google Scholar] [CrossRef]
- Liu, H.; Xu, Y.; Xiang, J.; Long, L.; Green, S.; Yang, Z.; Zimdahl, B.; Lu, J.; Cheng, N.; Horan, L.H.; et al. Targeting Alpha-Fetoprotein (AFP)-MHC Complex. with CAR T-Cell Therapy for Liver Cancer. Clin. Cancer Res. 2017, 23, 478–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Potential Diagnostic Biomarker | Molecular Functions | Diagnostic Application for HCC | Reference |
|---|---|---|---|
| HSP-90α | A molecular chaperone; is associated with the folding and unfolding of diverse proteins, such as signaling protein kinases, transcription factors. | Plasma HSP-90α levels in HCC group were significantly higher than that in the healthy control group and the benign liver disorder group, which did not depend on the origin of the patients with HCC. Combined with AFP and thymidine kinase 1(TK1), the detection of HSP90-α increased the sensitivity for the diagnosis of HCC. | [36,37] |
| miR-132/212 | Dysregulated in many human malignancies and plays important roles in tumor progression. | miR-132 and miR-212 are downregulated in HCC and cell lines, as tumor-suppressive roles. Combination of serum miR-132/212 cluster and AFP markedly improved sensitivity of diagnosis of HCC, compared with AFP or miR-132/212 alone. | [38] |
| Serum VN | A cell-adhesive glycoprotein in serum and plasma; is associated with inflammation, cell adhesion, cell necrosis, angiogenesis, etc. | Serum VN levels were significantly elevated in HBV-related HCC patients. Serum VN levels had similar diagnostic and prognostic values compared with serum AFP levels in HBV-positive HCC patients. | [39] |
| hnRNPH1 mRNA | As a kind of RNA binding protein, hnRNPH1 is associated with pre-mRNAs and appears to influence pre-mRNA processing and other aspects of mRNA metabolism and transport. It is highly expressed in many kinds of cancers. | The mRNA levels of hnRNPH1 in serum of HCC patients were remarkably higher than those in normal individuals, which were positively associated with Child–Pugh classification, portal vein tumor emboli, lymph node metastasis and tumor node metastasis (TNM) stage. It may be a novel biomarker for diagnosis of HCC in high-HBV-prevalence areas. | [40] |
| BCYRN1 | A type of long noncoding RNA; acts as a competing endogenous RNA, regulating cell migration, proliferation and survival. | BCYRN1 expression was elevated in HCC patients. It suggested that the diagnosis of HCC was remarkably improved with the combination of plasma BCYRN1 and AFP. | [41] |
| LBP | A serum protein, synthesized in the liver; is involved in the recognition, binding and transport of the bacterial cell wall compound lipopolysaccharide/endotoxin. | LBP is overexpressed in HCC tissues, especially in poorly differentiated tumors. | [44] |
| DCP | An abnormal prothrombin precursor produced in HCC; lacks the ability to interact with other coagulation factors. | In 63% of patients with HCC, serum DCP levels were found to be elevated. Combination of AFP and 2 additional biomarkers, AFP-L3 and DCP, results in improvement of overall survival of HCC patients. | [52,53] |
| SHh ligand | A unique signaling molecule in hedgehog signaling; controls embryonic development and tissue repair under physiological conditions. | The upregulation of SHh ligand was frequently observed in HCC and was greatly associated with drug resistance and metastasis of the malignancy through epithelial-to-mesenchymal transition (EMT). It could also be compensatory for detection of AFP-negative HCC patients. | [54] |
| Serum ANXA3 | A member of calcium-dependent phospholipid-binding protein family; participates in a diverse range of physiological activities that include anticoagulation, anti-inflammatory activities, endocytosis and exocytosis, signal transduction, cell proliferation, cell differentiation and cell apoptosis. | ANXA3 expression is significantly unregulated in patients with HCC. Clinically, ANXA3 expression in HCC patients sera and tissues is closely associated with aggressive clinical features. Serum ANXA3 provides greater diagnostic performance than AFP, especially in early diagnosis and discriminating HCC from patients at risk. | [55] |
| Regulations | Effects | Circulating (Extracellular) AFP | Cytoplasmic (Intracellular) AFP |
|---|---|---|---|
| Proliferation | Promotion | High doses (more than 100 mg/L) of purified human AFP induced dose-dependent growth inhibition. | Intracellular AFP was involved in the regulation of PI3K/AKT via the direct interaction with PTEN which resulted in suppression of the PTEN gene and enhancement of cell growth. |
| Inhibition | Low concentration of AFP (less than 100 mg/L) showed a weak stimulative growth effect in HepG2 cells. | None. | |
| Apoptosis | Induction | The induction of apoptosis was dose-dependent in various types of tumor cells, such as Raji, Jurkat and MCF-7 cells, at AFP concentrations of 0.7–3.0 μM and, more significantly, at 5.0–10.0 μM. | AFP interacted with IAP-2 and XIAP, abolished IAP caspase binding and rescued caspase-3 from inhibition, triggering cytochrome c-mediated apoptosis. |
| Inhibition | Extracellular and membrane-bound AFP prevented TNF-induced cytotoxicity and apoptosis of HepG2 cells. | Since its interaction with caspase-3, AFP eliminated apoptotic activity of TRAIL or TNF-induced in Bel 7402 cells and HepG2 cells, respectively. | |
| Immuno-suppression | For tumor cells | Extracellular AFP promotes tumor cells to secrete ligands such as Fas-L. | Intracellular AFP promotes the expression of surface antigens such as PD-L1 and B7-H4, so as to achieve the role of immune escape. |
| For immune cells | Extracellular AFP induces the apoptosis of immune cells and attenuates their antitumor function. | None. | |
| Stem cell properties | Promotion | None. | Transient changes of AFP expression in Bel 7402 or HLE cells could regulate the expression of K19, EpCAM and CXCR4, which are markers of stem cells. |
| Angiogenesis | Positive relation | Whole-exome sequencing data and DNA methylome profiling of 520 HCC patients demonstrated that AFP-high (serum concentration > 400 mg/L) tumors displayed significant activation of VEGF signaling. | None. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Guo, Q.; Wei, L. The Emerging Influences of Alpha-Fetoprotein in the Tumorigenesis and Progression of Hepatocellular Carcinoma. Cancers 2021, 13, 5096. https://doi.org/10.3390/cancers13205096
Xu Y, Guo Q, Wei L. The Emerging Influences of Alpha-Fetoprotein in the Tumorigenesis and Progression of Hepatocellular Carcinoma. Cancers. 2021; 13(20):5096. https://doi.org/10.3390/cancers13205096
Chicago/Turabian StyleXu, Ye, Qinglong Guo, and Libin Wei. 2021. "The Emerging Influences of Alpha-Fetoprotein in the Tumorigenesis and Progression of Hepatocellular Carcinoma" Cancers 13, no. 20: 5096. https://doi.org/10.3390/cancers13205096
APA StyleXu, Y., Guo, Q., & Wei, L. (2021). The Emerging Influences of Alpha-Fetoprotein in the Tumorigenesis and Progression of Hepatocellular Carcinoma. Cancers, 13(20), 5096. https://doi.org/10.3390/cancers13205096