
Towards Raman-Based Screening of Acute Lymphoblastic Leukemia-Type B (B-ALL) Subtypes

 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
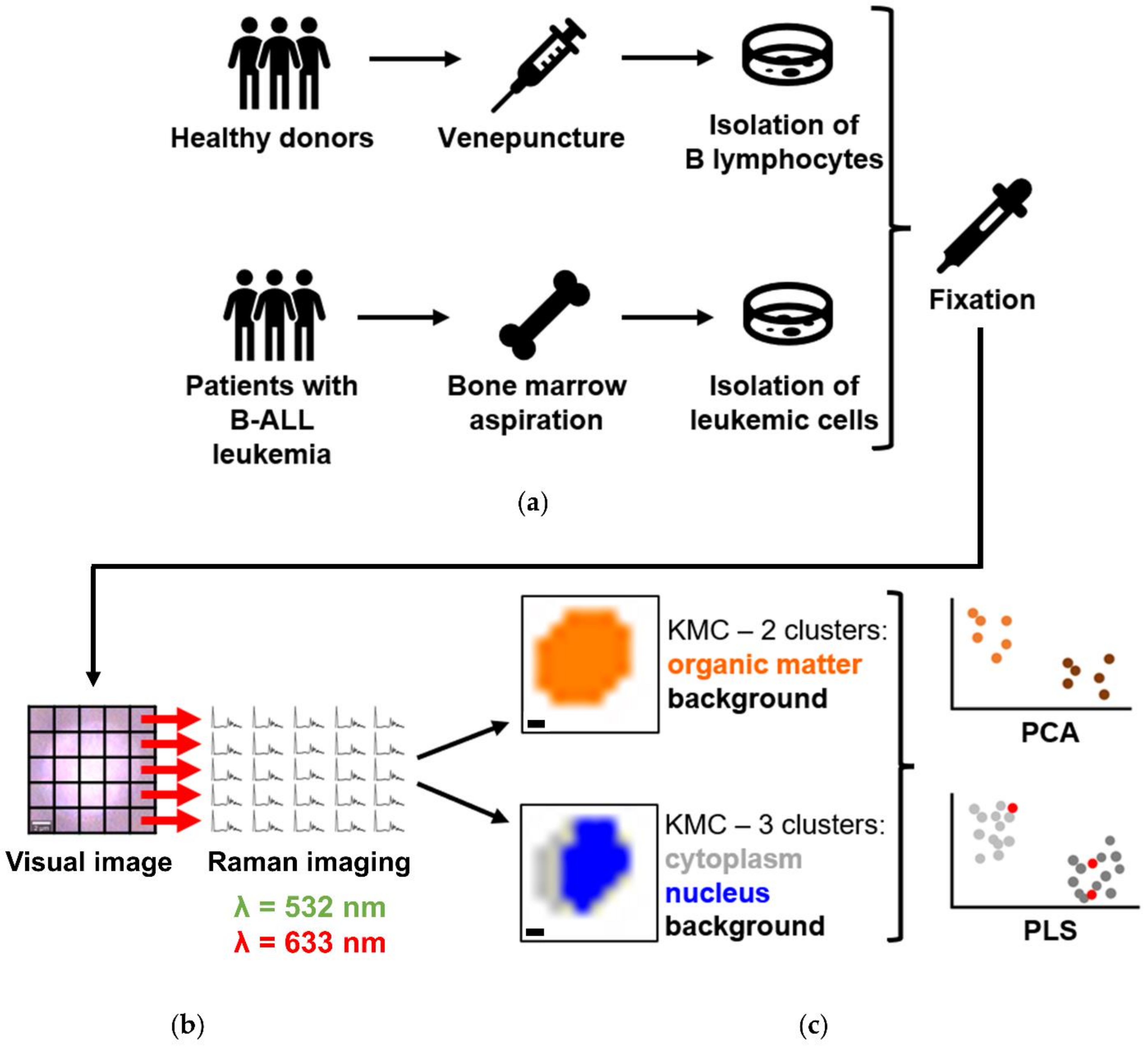
2. Materials and Methods
2.1. B Lymphocytes and Cancer Cells Isolation
2.2. Confocal Raman Imaging
2.3. Spectral Data Post-Processing and Analysis
3. Results
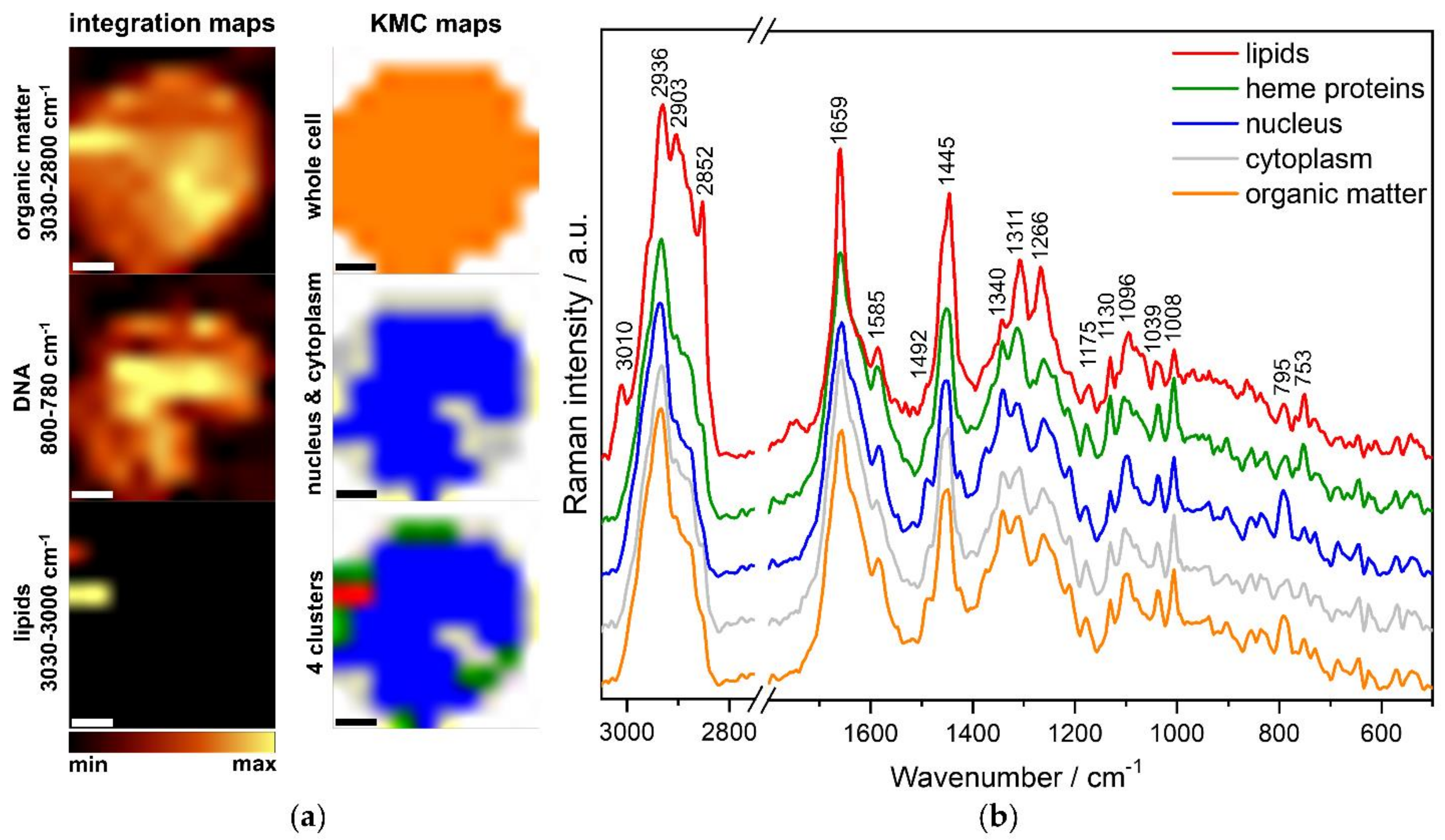
3.1. Characterization of the Molecular Composition of Normal B Lymphocytes and B-ALL Cells
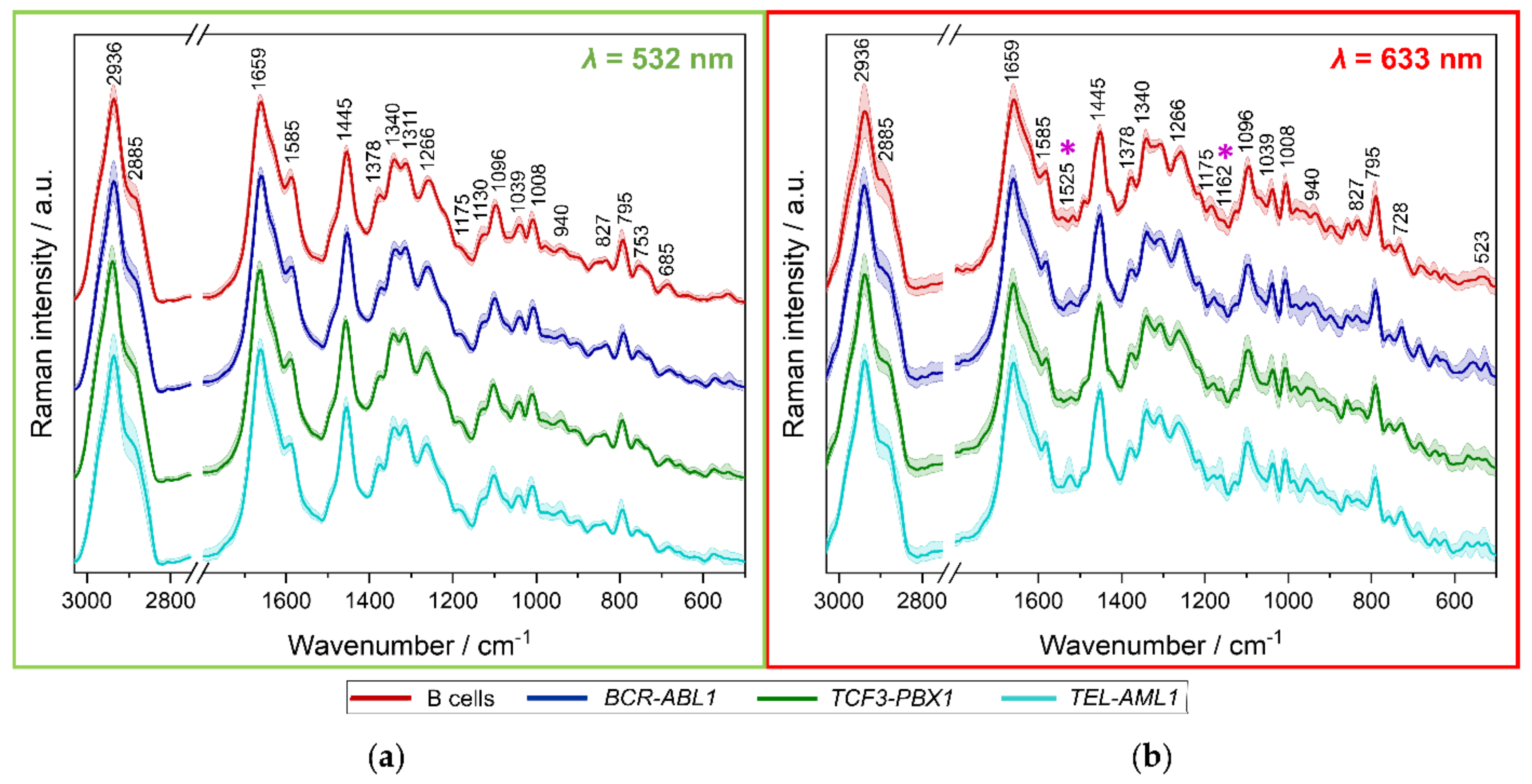
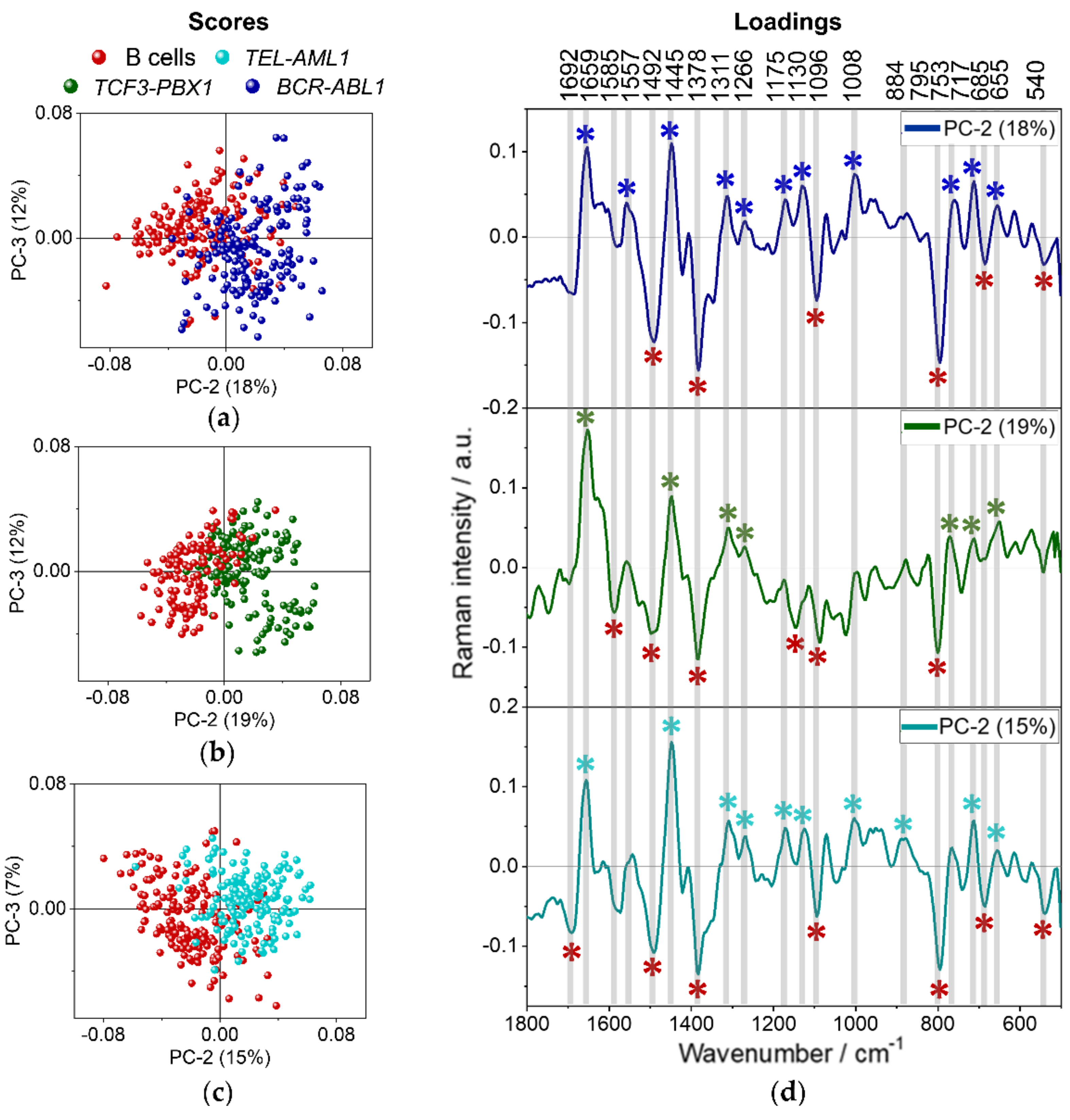
3.2. Spectroscopic Analysis of Different B-ALL Subtypes
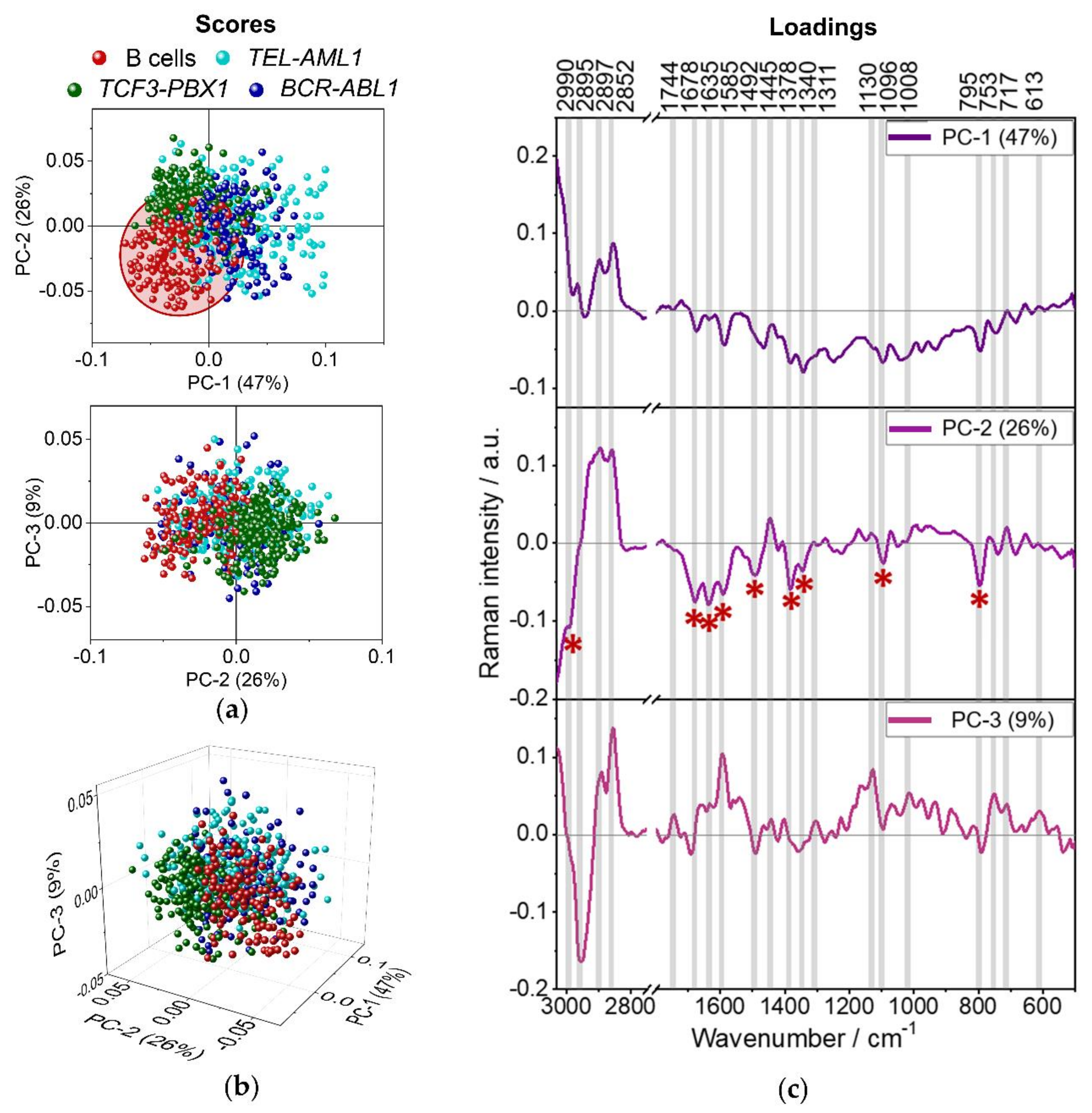
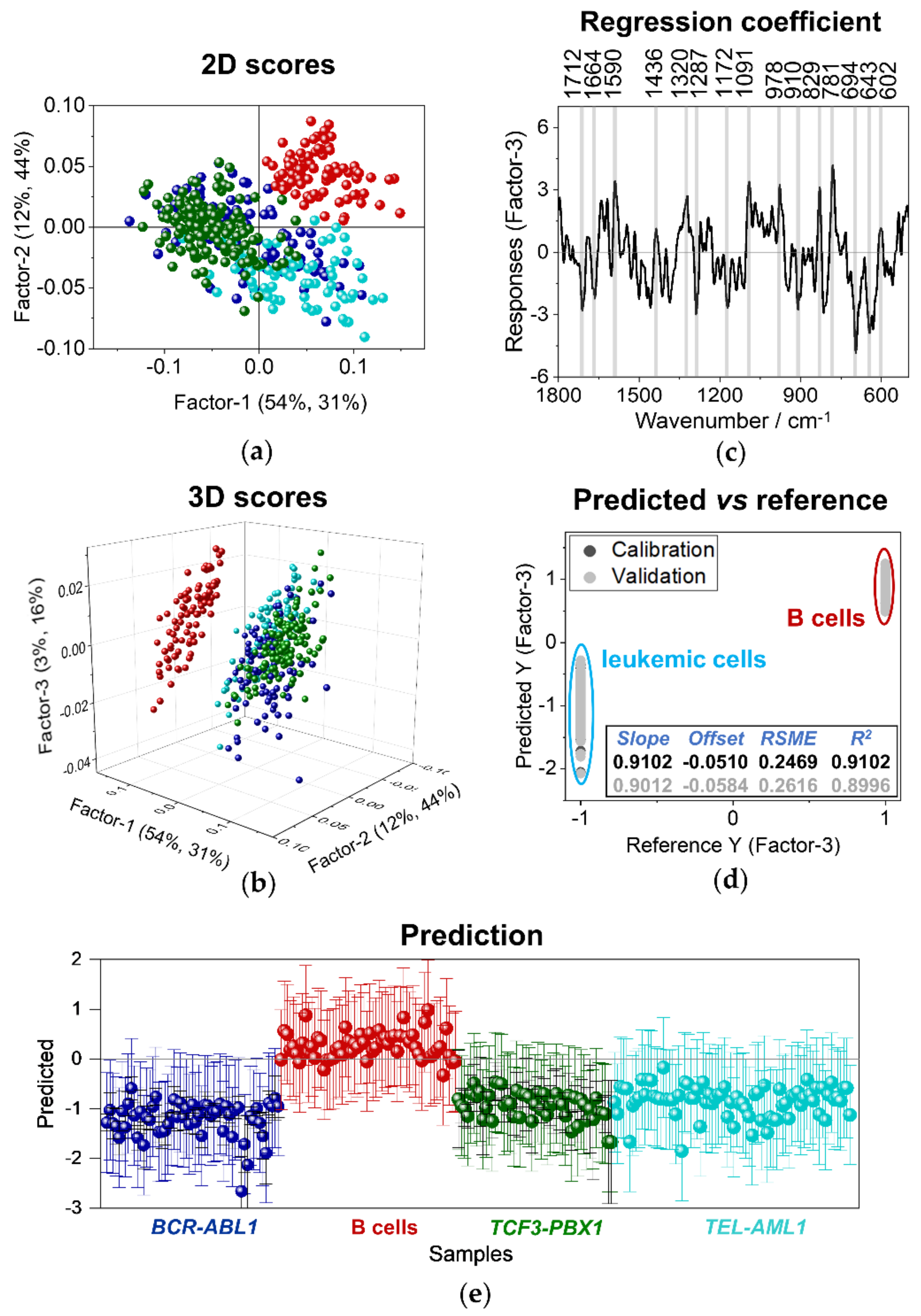
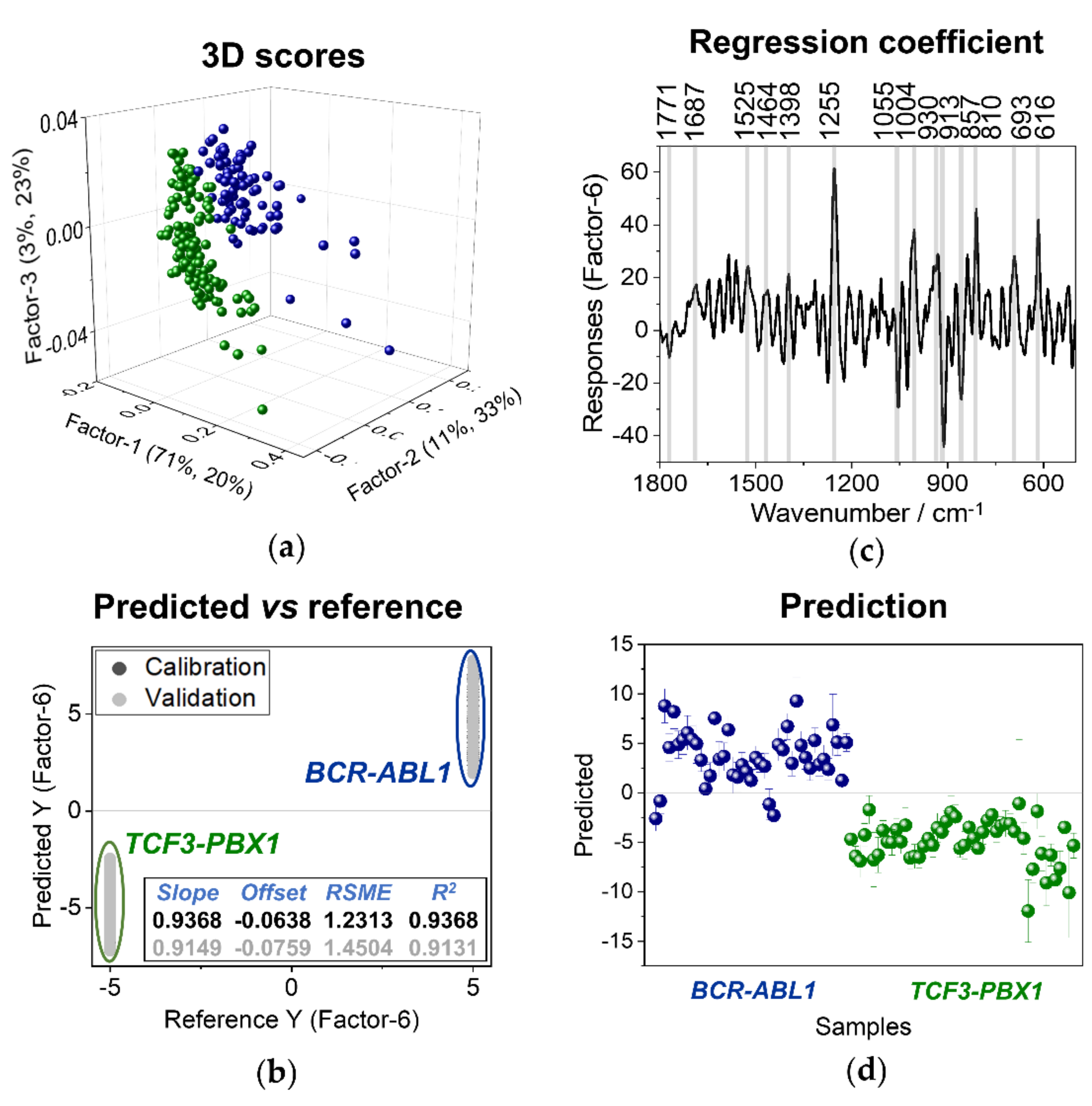
3.3. Discriminant Analysis of Normal B Lymphocytes and B-ALL Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malard, F.; Mohty, M. Acute lymphoblastic leukaemia. Lancet 2020, 395, 1146–1162. [Google Scholar] [CrossRef]
- Inaba, H.; Mullighan, C.G. Pediatric acute lymphoblastic leukemia. Haematologica 2020, 105, 2524–2539. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.G.; Mullighan, C.G. The biology of B-progenitor acute lymphoblastic leukemia. Cold Spring Harb. Perspect. Med. 2020, 10, 1–22. [Google Scholar] [CrossRef]
- Vibrational Spectroscopy. From Theory to Practice; Małek, K., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2016; ISBN 978-83-01 18885-6. [Google Scholar]
- Hedegaard, M.; Matthäus, C.; Hassing, S.; Krafft, C.; Diem, M.; Popp, J. Spectral unmixing and clustering algorithms for assessment of single cells by Raman microscopic imaging. Theor. Chem. Acc. 2011, 130, 1249–1260. [Google Scholar] [CrossRef]
- Kupis, J.; Skowron-Jaskólska, M.; Szczukocki, D.; Krawczyk, B. Metrologia i Chemometria w Analityce Środowiska; wersja ele.; Wydawnictwo Uniwersytetu Łódzkiego: Łódź, Poland, 2016; ISBN 978-83-8088-177-8. [Google Scholar]
- Miller, J.; Miller, J. Statystyka i Chemometria w Chemii Analitycznej; Wydanie, I., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2016; ISBN 978-83-01-18303-5. [Google Scholar]
- Tobias, R.D. An introduction to partial least squares regression. In Proceedings of the Twentieth Annual SAS Users Group International Conference, Orlando, FL, USA, 2–5 April 1995; pp. 1250–1257. [Google Scholar]
- Brereton, R.G.; Lloyd, G.R. Partial least squares discriminant analysis: Taking the magic away. J. Chemom. 2014, 28, 213–225. [Google Scholar] [CrossRef]
- Xu, J.; Yu, T.; Zois, C.E.; Cheng, J.X.; Tang, Y.; Harris, A.L.; Huang, W.E. Unveiling cancer metabolism through spontaneous and coherent raman spectroscopy and stable isotope probing. Cancers 2021, 13, 1718. [Google Scholar] [CrossRef]
- Contorno, S.; Darienzo, R.E.; Tannenbaum, R. Evaluation of aromatic amino acids as potential biomarkers in breast cancer by Raman spectroscopy analysis. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, Y.; Song, Y.; Xu, J. Accuracy of Raman spectroscopy for differentiating skin cancer from normal tissue. Medicine 2018, 97. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, C.J.; Gala De Pablo, J.; Tinkler-Hundal, E.; Wood, H.M.; Evans, S.D.; West, N.P. Developing a Raman spectroscopy-based tool to stratify patient response to pre-operative radiotherapy in rectal cancer. Analyst 2021, 146, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Li, J.; Yang, L.; Zheng, B.; Wang, J.; Lv, N.; Luo, J.; Martin, F.L.; Liu, D.; He, J. Raman spectroscopy as a potential diagnostic tool to analyse biochemical alterations in lung cancer. Analyst 2020, 145, 385–392. [Google Scholar] [CrossRef]
- Bai, Y.; Yu, Z.; Yi, S.; Yan, Y.; Huang, Z.; Qiu, L. Raman spectroscopy-based biomarker screening by studying the fingerprint characteristics of chronic lymphocytic leukemia and diffuse large B-cell lymphoma. J. Pharm. Biomed. Anal. 2020, 190, 113514. [Google Scholar] [CrossRef]
- Féré, M.; Gobinet, C.; Liu, L.H.; Beljebbar, A.; Untereiner, V.; Gheldof, D.; Chollat, M.; Klossa, J.; Chatelain, B.; Piot, O. Implementation of a classification strategy of Raman data collected in different clinical conditions: Application to the diagnosis of chronic lymphocytic leukemia. Anal. Bioanal. Chem. 2020, 412, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Hassoun, M.; Köse, N.; Kiselev, R.; Kirchberger-Tolstik, T.; Schie, I.W.; Krafft, C.; Popp, J. Quantitation of acute monocytic leukemia cells spiked in control monocytes using surface-enhanced Raman spectroscopy. Anal. Methods 2018, 10, 2785–2791. [Google Scholar] [CrossRef]
- Managò, S.; Valente, C.; Mirabelli, P.; De Luca, A.C. Discrimination and classification of acute lymphoblastic leukemia cells by Raman spectroscopy. Opt. Sensors 2015, 9506, 95060Z. [Google Scholar] [CrossRef]
- Managò, S.; Zito, G.; De Luca, A.C. Raman microscopy based sensing of leukemia cells: A review. Opt. Laser Technol. 2018, 108, 7–16. [Google Scholar] [CrossRef]
- Managò, S.; Valente, C.; Mirabelli, P.; Circolo, D.; Basile, F.; Corda, D.; De Luca, A.C. A reliable Raman-spectroscopy-based approach for diagnosis, classification and follow-up of B-cell acute lymphoblastic leukemia. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Walther, T.C.; Farese, R.V. Lipid droplets and cellular lipid metabolism. Annu. Rev. Biochem. 2012, 81, 687–714. [Google Scholar] [CrossRef] [Green Version]
- Schie, I.W.; Nolte, L.; Pedersen, T.L.; Smith, Z.; Wu, J.; Yahiatène, I.; Newman, J.W.; Huser, T. Direct comparison of fatty acid ratios in single cellular lipid droplets as determined by comparative Raman spectroscopy and gas chromatography. Analyst 2013, 138, 6662–6670. [Google Scholar] [CrossRef] [Green Version]
- Wart, H.E.V.A.N.; Harold, A. Agreement with the disulfide stretching frequency-conformation correlation of Sugeta, Go, and Miyazawa. Proc. Nat. Acad. Sci. USA 1986, 83, 3064–3067. [Google Scholar] [CrossRef] [Green Version]
- Hernández, B.; Pflüger, F.; Kruglik, S.G.; Ghomi, M. Characteristic Raman lines of phenylalanine analyzed by a multiconformational approach. J. Raman Spectrosc. 2013, 44, 827–833. [Google Scholar] [CrossRef]
- Krafft, C.; Knetschke, T.; Siegner, A.; Funk, R.H.W.; Salzer, R. Mapping of single cells by near infrared Raman microspectroscopy. Vib. Spectrosc. 2003, 32, 75–83. [Google Scholar] [CrossRef]
- Notingher, I.; Verrier, S.; Romanska, H.; Bishop, A.E.; Polak, J.M.; Hench, L.L. In situ characterisation of living cells by Raman spectroscopy. Spectroscopy 2002, 16, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Czamara, K.; Majzner, K.; Pacia, M.Z.; Kochan, K.; Kaczor, A. Raman spectroscopy of lipids: A review. J. Raman Spectrosc. 2015, 46, 4–20. [Google Scholar] [CrossRef]
- Wood, B.R.; Mcnaughton, D. Raman excitation wavelength investigation of single red blood cells in vivo. J. Raman. Spectrosc. 2002, 33, 517–523. [Google Scholar] [CrossRef]
- Atkins, C.G.; Buckley, K.; Blades, M.W.; Turner, R.F.B. Raman Spectroscopy of Blood and Blood Components. Appl. Spectrosc. 2017, 71, 767–793. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, C.; Severcan, F. Role of vibrational spectroscopy in stem cell research. Spectroscopy 2012, 27, 167–184. [Google Scholar] [CrossRef]
- Rygula, A.; Majzner, K.; Marzec, K.M.; Kaczor, A.; Pilarczyk, M.; Baranska, M. Raman spectroscopy of proteins: A review. J. Raman Spectrosc. 2013, 44, 1061–1076. [Google Scholar] [CrossRef]
- Lyng, F.M.; Ramos, I.R.M.; Ibrahim, O.; Byrne, H.J. Vibrational microspectroscopy for cancer screening. Appl. Sci. 2015, 5, 23–35. [Google Scholar] [CrossRef]
- Ramoji, A.; Neugebauer, U.; Bocklitz, T.; Foerster, M.; Kiehntopf, M.; Bauer, M.; Popp, J. Toward a Spectroscopic Hemogram: Raman Spectroscopic Differentiation of the Two Most Abundant Leukocytes from Peripheral Blood. Anal. Chem. 2012, 84, 5335–5342. [Google Scholar] [CrossRef]
- Perez-Guaita, D.; Kochan, K.; Martin, M.; Andrew, D.W.; Heraud, P.; Richards, J.S.; Wood, B.R. Multimodal vibrational imaging of cells. Vib. Spectrosc. 2017, 91, 46–58. [Google Scholar] [CrossRef]
- Puppels, G.J.; Garritsen, H.S.P.; Kummer, J.A.; Greve, J. Carotenoids located in human lymphocyte subpopulations and natural killer cells by Raman microspectroscopy. Cytometry 1993, 14, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Strokotov, D.I.; Yurkin, M.A.; Gilev, K.V.; van Bockstaele, D.R.; Hoekstra, A.G.; Rubtsov, N.B.; Maltsev, V.P. Is there a difference between T- and B-lymphocyte morphology? J. Biomed. Opt. 2009, 14, 064036. [Google Scholar] [CrossRef] [PubMed]
- Morrish, R.; Yim, K.H.W.; Pagliara, S.; Palombo, F.; Chahwan, R.; Stone, N. Single Cell Label-Free Probing of Chromatin Dynamics During B Lymphocyte Maturation. Front. Cell Dev. Biol. 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, D.; Jarori, G.K.; Davisson, V.J.; Ben-Amotz, D. The Raman detection of peptide tyrosine phosphorylation. Anal. Biochem. 2004, 332, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Managò, S.; Valente, C.; Mirabelli, P.; Napolitano, M.; Corda, D.; De Luca, A.C. Identification and classification of acute lymphoblastic leukemia cells from peripheral blood by using Raman spectroscopy. IET Conf. Publ. 2016, 2016. [Google Scholar] [CrossRef]
- Chan, J.W.; Taylor, D.S.; Lane, S.M.; Zwerdling, T.; Tuscano, J.; Huser, T. Nondestructive identification of individual leukemia cells by laser trapping raman spectroscopy. Anal. Chem. 2008, 80, 2180–2187. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, C.M.; Gunn, L.; Skibola, C.F.; Zhang, L.; Shiao, R.; Fu, S.; Dalmasso, E.A.; Metayer, C.; Dahl, G.V.; Buffler, P.A.; et al. Proteomic analysis of childhood leukemia. Leukemia 2005, 19, 1713–1718. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.W.; Wang, J.; He, K.; Jin, B.F.; Wang, H.X.; Li, W.; Kang, L.H.; Hu, M.R.; Li, H.Y.; Yu, M.; et al. Proteomic analysis of human acute leukemia cells: Insight into their classification. Clin. Cancer Res. 2004, 10, 6887–6896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.R.; Leikin, S.; Albo, V.; Sather, H.; Hammond, D. Prognostic Importance of Morphology (FAB Classification) in Childhood Acute Lymphoblastic Leukaemia (ALL). Br. J. Haematol. 1981, 48, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Mulas, M.F.; Abete, C.; Pulisci, D.; Pani, A.; Massidda, B.; Dessì, S.; Mandas, A. Cholesterol esters as growth regulators of lymphocytic leukaemia cells. Cell Prolif. 2011, 44, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Tucci, J.; Chen, T.; Margulis, K.; Orgel, E.; Paszkiewicz, R.L.; Cohen, M.D.; Oberley, M.J.; Wahhab, R.; Jones, A.E.; Divakaruni, A.S.; et al. Adipocytes Provide Fatty Acids to Acute Lymphoblastic Leukemia Cells. Front. Oncol. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Okada, M.; Isaac, N.; Flotildes, A.; Endo, H.; Kawata, S. Label-free Raman observation of cytochrome c dynamics during apoptosis. Proc. Nat. Acad. Sci. USA 2012, 109, 28–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.C.; Borutaite, V. Regulation of apoptosis by the redox state of cytochrome c. Biochim. Biophys. Acta-Bioenerg. 2008, 1777, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.; Johnson, D.B.F.; Tardif, H.L.; Tome, M.E.; Briehl, M.M. Increased cytochrome c correlates with poor survival in aggressive lymphoma. Oncol. Lett. 2010, 1, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Ramanauskaite, R.B.; Segers-Nolten, I.G.M.J.; De Grauw, K.J.; Sijtsema, N.M.; Van Der Maas, L.; Greve, J.; Otto, C.; Figdor, C.G. Carotenoid levels in human lymphocytes, measured by Raman microspectroscopy. Pure Appl. Chem. 1997, 69, 2131–2134. [Google Scholar] [CrossRef] [Green Version]
- Erber, W.N. Human leucocyte differentiation antigens: Review of the CD nomenclature. Pathology 1990, 22, 61–69. [Google Scholar] [CrossRef]
- Pully, V.V.; Lenferink, A.T.M.; Otto, C. Time-lapse Raman imaging of single live lymphocytes. J. Raman Spectrosc. 2011, 42, 167–173. [Google Scholar] [CrossRef]
- Ermakov, I.V.; Sharifzadeh, M.; Ermakova, M.R.; Gellermann, W. Resonance Raman detection of carotenoid antioxidants in living human tissue. J. Biomed. Opt. 2005, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Veronelli, M.; Zerbi, G.; Stradi, R. In situ resonance Raman spectra of carotenoids in bird’s feathers. J. Raman Spectrosc. 1995, 26, 683–692. [Google Scholar] [CrossRef]
- Adamczyk, A.; Matuszyk, E.; Radwan, B.; Rocchetti, S.; Chlopicki, S.; Baranska, M. Toward Raman Subcellular Imaging of Endothelial Dysfunction. J. Med. Chem. 2021, 64, 4396–4409. [Google Scholar] [CrossRef]
- Andersson, A.; Olofsson, T.; Lindgren, D.; Nilsson, B.; Ritz, C.; Edén, P.; Lassen, C.; Råde, J.; Fontes, M.; Mörse, H.; et al. Molecular signatures in childhood acute leukemia and their correlations to expression patterns in normal hematopoietic subpopulations. Proc. Natl. Acad. Sci. USA 2005, 102, 19069–19074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, A.A.; Shi, L. Mammalian cell and tissue imaging using Raman and coherent Raman microscopy. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1501. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BAND/cm−1 | Assignment | Classification |
|---|---|---|
| 540 | Disulfide bonds [23] | Proteins |
| 613 | Ring breathing of Phe [24,25,26] | Proteins |
| 655 | Ring breathing of Phe and Tyr [25,26] | Proteins |
| 685–694 | C-C twist, G [19] | Proteins, nucleic acids |
| 717 | Vibrations of the choline group [27] | Phospholipids, sphingolipids |
| 728 | Ring breathing of A [4] | Nucleic acids |
| 753 | Pyrrole breathing mode [28,29] | Heme proteins |
| 795 | Ring breathing of U, T, C; backbone O-P-O [4] | Nucleic acids |
| 827 | PO2− asymmetric stretching, Tyr [19,29,30] | Nucleic acids, proteins |
| 884 | CH2 rocking deformation mode [4] | Proteins |
| 940 | Skeletal modes [4] N–Cα–C for α-helix [31] | Polisaccharides Proteins |
| 1008 | Ring breathing of Phe [4] | Proteins |
| 1039 | C-N stretching [4] | Proteins |
| 1096 | PO2− symmetric stretching [4,19] | Nucleic acids, phospholipids |
| 1130 | C–C, C–O, C-N stretching [31,32] | Heme proteins, proteins, lipids |
| 1162 | C-C stretching [33] | Carotenoids |
| 1175 | Tyr, Phe, C-H bend | Proteins |
| 1266 | =C−H deformation, amide III; C-N stretching in α-helix [4] | Lipids, proteins |
| 1311 | CH2 twist, C-N stretching, Cα-Cβ stretching [19,32] | Lipids, proteins, heme proteins |
| 1340 | CH2/CH3 wagging, twisting and/or bending; CH deformation, G modes [4,19] | Lipids Proteins Nucleic acids |
| 1378 | COO– symmetric stretching [30,34] | Proteins, nucleic acids |
| 1449–1455 | CH2/CH3 bending in-plane [4] | Lipids, proteins |
| 1492 | Ring breathing of G, A, CH deformation mode [19] | Nucleic acids Nucleic acids, proteins |
| 1525 | C=C stretching [33] | Carotenoids |
| 1557 | Phe and Trp [30] | Proteins |
| 1585 | Pyrimidine ring; C=C bending in Phe [4] | Nucleic acids, heme proteins |
| 1635–1670 | Amide I, C=C stretching [4,19,32] | Proteins, lipids |
| 2850–2870 | CH2 symmetric stretching [4] | Lipids, fatty acids |
| 2885 | CH3 symmetric stretching, CH2 asymmetric stretching [4] | Lipids, fatty acids, proteins |
| 2933–2945 | CH3 asymmetric stretching [4] | Lipids, fatty acids, proteins |
| 3010 | =CH2 stretching [4] | Lipids, fatty acids |
| CELLS | N | λ [nm] | Number of Cells | Heme Proteins Clusters | Carotenoid Clusters | Lipid Clusters |
|---|---|---|---|---|---|---|
| B cells | 5 | 532 | 255 | 144 (56.5%) | 0 (0.0%) | 3 (1.2%) |
| 633 | 247 | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | ||
| BCR-ABL1 | 3 | 532 | 183 | 124 (67.8%) | 0 (0.0%) | 5 (2.7%) |
| 633 | 169 | 0 (0.0%) | 14 (8.3%) | 7 (4.1%) | ||
| TCF3-PBX1 | 4 | 532 | 331 | 210 (63.4%) | 8 (2.4%) | 9 (2.7%) |
| 633 | 271 | 4 (1.5%) | 32 (11.8%) | 2 (0.7%) | ||
| TEL-AML1 | 4 | 532 | 314 | 189 (60.2%) | 16 (5.1%) | 66 (21.0%) |
| 633 | 257 | 3 (1.2%) | 32 (12.5%) | 2 (1.1%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leszczenko, P.; Borek-Dorosz, A.; Nowakowska, A.M.; Adamczyk, A.; Kashyrskaya, S.; Jakubowska, J.; Ząbczyńska, M.; Pastorczak, A.; Ostrowska, K.; Baranska, M.; et al. Towards Raman-Based Screening of Acute Lymphoblastic Leukemia-Type B (B-ALL) Subtypes. Cancers 2021, 13, 5483. https://doi.org/10.3390/cancers13215483
Leszczenko P, Borek-Dorosz A, Nowakowska AM, Adamczyk A, Kashyrskaya S, Jakubowska J, Ząbczyńska M, Pastorczak A, Ostrowska K, Baranska M, et al. Towards Raman-Based Screening of Acute Lymphoblastic Leukemia-Type B (B-ALL) Subtypes. Cancers. 2021; 13(21):5483. https://doi.org/10.3390/cancers13215483
Chicago/Turabian StyleLeszczenko, Patrycja, Aleksandra Borek-Dorosz, Anna Maria Nowakowska, Adriana Adamczyk, Sviatlana Kashyrskaya, Justyna Jakubowska, Marta Ząbczyńska, Agata Pastorczak, Kinga Ostrowska, Malgorzata Baranska, and et al. 2021. "Towards Raman-Based Screening of Acute Lymphoblastic Leukemia-Type B (B-ALL) Subtypes" Cancers 13, no. 21: 5483. https://doi.org/10.3390/cancers13215483
APA StyleLeszczenko, P., Borek-Dorosz, A., Nowakowska, A. M., Adamczyk, A., Kashyrskaya, S., Jakubowska, J., Ząbczyńska, M., Pastorczak, A., Ostrowska, K., Baranska, M., Marzec, K. M., & Majzner, K. (2021). Towards Raman-Based Screening of Acute Lymphoblastic Leukemia-Type B (B-ALL) Subtypes. Cancers, 13(21), 5483. https://doi.org/10.3390/cancers13215483