
The Adhesome Network: Key Components Shaping the Tumour Stroma



Abstract
:Simple Summary
Abstract
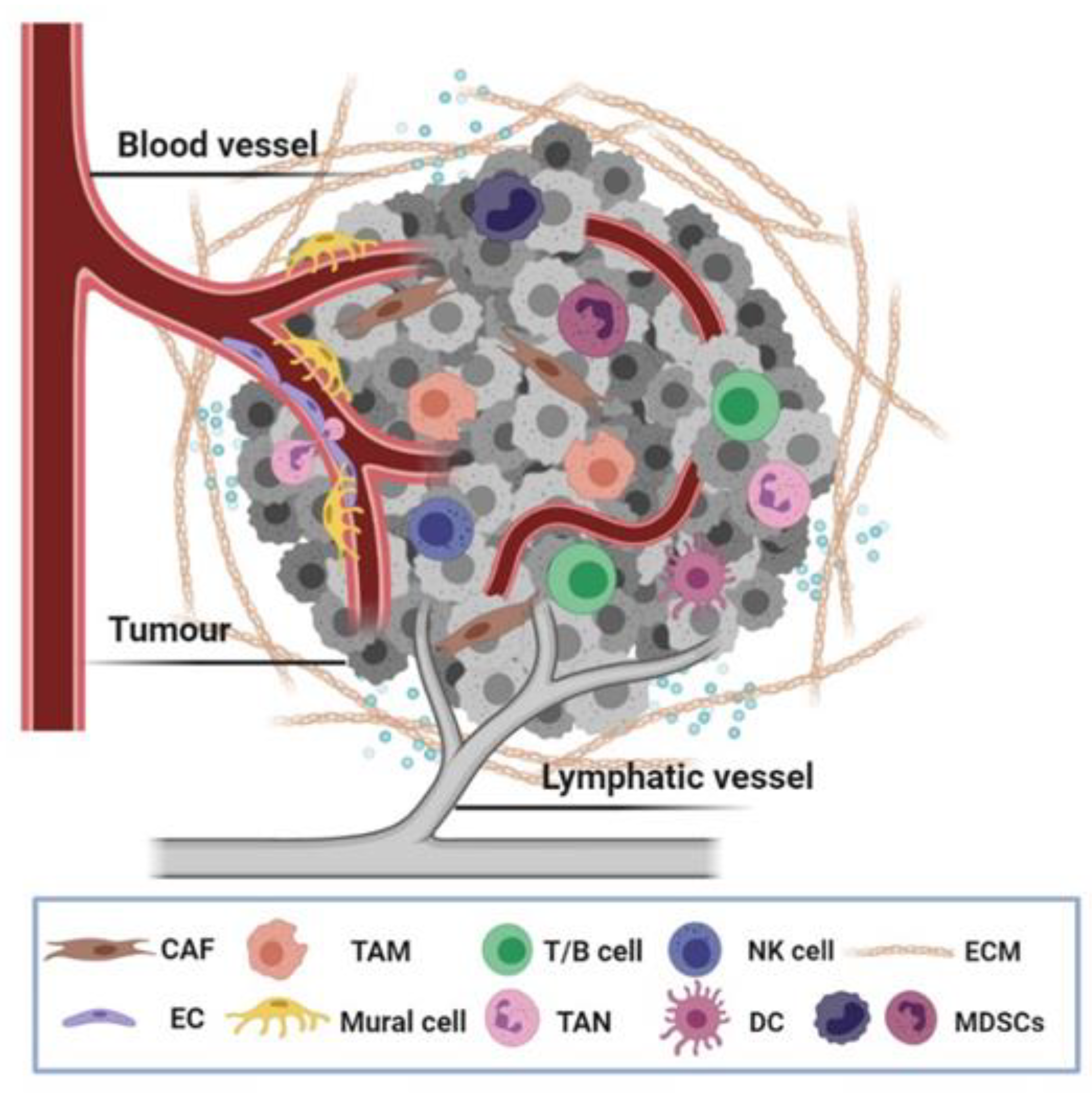
1. Introduction
2. Adhesome Function in Vasculature
2.1. Endothelial Cell Adhesome
2.1.1. Adhesome Signalling
2.1.2. Adhesion Organisation
2.1.3. Actin Regulatory Layer
2.1.4. Emerging Adhesome Players in Tumour Endothelium
2.2. Mural Adhesome
2.2.1. Adhesome Signalling
2.2.2. Adhesion Organisation
3. Adhesome Function in CAFs
3.1. Adhesome Signalling
3.2. Adhesion Organisation
3.3. Actin Regulatory Layer
4. Adhesome Function in Immune Cells
4.1. Innate Immune Cell Adhesome
4.1.1. Adhesome Signalling
4.1.2. Adhesion Organisation
4.1.3. Actin Regulatory Layer
4.2. Adaptive Immune Cell Adhesome
4.2.1. Adhesome Signalling
4.2.2. Adhesion Organisation
4.2.3. Actin Regulatory Layer
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Coussens, L.M. Accessories to the Crime: Functions of Cells Recruited to the Tumor Microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The Tumor Microenvironment at a Glance. J. Cell. Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, H.; Remark, R.; Gnjatic, S.; Merad, M. Host Tissue Determinants of Tumour Immunity. Nat. Rev. Cancer 2019, 19, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Yamada, K.M. Molecular Architecture and Function of Matrix Adhesions. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, C.A.; Bergeron, K.-F.; Whittle, J.; Brandhorst, B.P.; Burke, R.D.; Hynes, R.O. The Echinoderm Adhesome. Dev. Biol. 2006, 300, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma’ayan, A.; Iyengar, R.; Geiger, B. Functional Atlas of the Integrin Adhesome. Nat. Cell Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell Adhesion: Integrating Cytoskeletal Dynamics and Cellular Tension. Nat. Rev. Mol. Cell Biol. 2010, 11, 633–643. [Google Scholar] [CrossRef]
- Byron, A.; Frame, M.C. Adhesion Protein Networks Reveal Functions Proximal and Distal to Cell-Matrix Contacts. Curr. Opin. Cell Biol. 2016, 39, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Byron, A.; Morgan, M.R.; Humphries, M.J. Adhesion Signalling Complexes. Curr. Biol. 2010, 20, R1063–R1067. [Google Scholar] [CrossRef] [Green Version]
- Horton, E.R.; Byron, A.; Askari, J.A.; Ng, D.H.J.; Millon-Frémillon, A.; Robertson, J.; Koper, E.J.; Paul, N.R.; Warwood, S.; Knight, D.; et al. Definition of a Consensus Integrin Adhesome and Its Dynamics during Adhesion Complex Assembly and Disassembly. Nat. Cell Biol. 2015, 17, 1577–1587. [Google Scholar] [CrossRef] [Green Version]
- Horton, E.R.; Astudillo, P.; Humphries, M.J.; Humphries, J.D. Mechanosensitivity of Integrin Adhesion Complexes: Role of the Consensus Adhesome. Exp. Cell Res. 2016, 343, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, E.R.; Humphries, J.D.; James, J.; Jones, M.C.; Askari, J.A.; Humphries, M.J. The Integrin Adhesome Network at a Glance. J. Cell Sci. 2016, 129, 4159–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavora, B.; Batista, S.; Reynolds, L.E.; Jadeja, S.; Robinson, S.; Kostourou, V.; Hart, I.; Fruttiger, M.; Parsons, M.; Hodivala-Dilke, K.M. Endothelial FAK Is Required for Tumour Angiogenesis. EMBO Mol. Med. 2010, 2, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Borboa, A.K.; Chun, H.B.; Baird, A.; Eliceiri, B.P. Conditional Deletion of the Focal Adhesion Kinase FAK Alters Remodeling of the Blood-Brain Barrier in Glioma. Cancer Res. 2010, 70, 10131–10140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostourou, V.; Lechertier, T.; Reynolds, L.E.; Lees, D.M.; Baker, M.; Jones, D.T.; Tavora, B.; Ramjaun, A.R.; Birdsey, G.M.; Robinson, S.D.; et al. FAK-Heterozygous Mice Display Enhanced Tumour Angiogenesis. Nat. Commun. 2013, 4, 2020. [Google Scholar] [CrossRef] [Green Version]
- Jean, C.; Chen, X.L.; Nam, J.-O.; Tancioni, I.; Uryu, S.; Lawson, C.; Ward, K.K.; Walsh, C.T.; Miller, N.L.G.; Ghassemian, M.; et al. Inhibition of Endothelial FAK Activity Prevents Tumor Metastasis by Enhancing Barrier Function. J. Cell Biol. 2014, 204, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, A.N.; Lees, D.M.; Bodrug, N.; Lechertier, T.; Fernandez, I.; D’Amico, G.; Dukinfield, M.; Batista, S.; Tavora, B.; Serrels, B.; et al. Focal Adhesion Kinase (FAK) Tyrosine 397E Mutation Restores the Vascular Leakage Defect in Endothelium-Specific FAK-Kinase Dead Mice: Endothelial FAKY397E Mutation and Tumour Vascular Leakage. J. Pathol. 2017, 242, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, A.-R.; Bodrug, N.; Gomez-Escudero, J.; Carter, E.P.; Reynolds, L.E.; Georgiou, P.N.; Fernandez, I.; Lees, D.M.; Kostourou, V.; Alexopoulou, A.N.; et al. Tumor Angiogenesis Is Differentially Regulated by Phosphorylation of Endothelial Cell Focal Adhesion Kinase Tyrosines-397 and -861. Cancer Res. 2019, 79, 4371–4386. [Google Scholar] [CrossRef] [Green Version]
- Lechertier, T.; Reynolds, L.E.; Kim, H.; Pedrosa, A.R.; Gómez-Escudero, J.; Muñoz-Félix, J.M.; Batista, S.; Dukinfield, M.; Demircioglu, F.; Wong, P.P.; et al. Pericyte FAK Negatively Regulates Gas6/Axl Signalling to Suppress Tumour Angiogenesis and Tumour Growth. Nat. Commun. 2020, 11, 2810. [Google Scholar] [CrossRef] [PubMed]
- Lees, D.M.; Reynolds, L.E.; Pedrosa, A.R.; Roy-Luzarraga, M.; Hodivala-Dilke, K.M. Phosphorylation of Pericyte FAK-Y861 Affects Tumour Cell Apoptosis and Tumour Blood Vessel Regression. bioRxiv 2020. [Google Scholar] [CrossRef]
- Stokes, J.B.; Adair, S.J.; Slack-Davis, J.K.; Walters, D.M.; Tilghman, R.W.; Hershey, E.D.; Lowrey, B.; Thomas, K.S.; Bouton, A.H.; Hwang, R.F.; et al. Inhibition of Focal Adhesion Kinase by PF-562,271 Inhibits the Growth and Metastasis of Pancreatic Cancer Concomitant with Altering the Tumor Microenvironment. Mol. Cancer Ther. 2011, 10, 2135–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Hegde, S.; Knolhoff, B.L.; Zhu, Y.; Herndon, J.M.; Meyer, M.A.; Nywening, T.M.; Hawkins, W.G.; Shapiro, I.M.; Weaver, D.T.; et al. Targeting Focal Adhesion Kinase Renders Pancreatic Cancers Responsive to Checkpoint Immunotherapy. Nat. Med. 2016, 22, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Q.; Tu, K.; Wang, Y.; Wang, X.; Liu, D.; Chen, C.; Liu, D.; Yang, R.; Qiu, W.; et al. Focal Adhesion Kinase Promotes Hepatic Stellate Cell Activation by Regulating Plasma Membrane Localization of TGFβ Receptor 2. Hepatol. Commun. 2020, 4, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Demircioglu, F.; Wang, J.; Candido, J.; Costa, A.S.H.; Casado, P.; de Luxan Delgado, B.; Reynolds, L.E.; Gomez-Escudero, J.; Newport, E.; Rajeeve, V.; et al. Cancer Associated Fibroblast FAK Regulates Malignant Cell Metabolism. Nat. Commun. 2020, 11, 1290. [Google Scholar] [CrossRef] [Green Version]
- Zaghdoudi, S.; Decaup, E.; Belhabib, I.; Samain, R.; Cassant-Sourdy, S.; Rochotte, J.; Brunel, A.; Schlaepfer, D.; Cros, J.; Neuzillet, C.; et al. FAK Activity in Cancer-Associated Fibroblasts Is a Prognostic Marker and a Druggable Key Metastatic Player in Pancreatic Cancer. EMBO Mol. Med. 2020, 12, e12010. [Google Scholar] [CrossRef]
- Wu, H.-J.; Hao, M.; Yeo, S.K.; Guan, J.-L. FAK Signaling in Cancer-Associated Fibroblasts Promotes Breast Cancer Cell Migration and Metastasis by Exosomal MiRNAs-Mediated Intercellular Communication. Oncogene 2020, 39, 2539–2549. [Google Scholar] [CrossRef]
- Walsh, C.; Tanjoni, I.; Uryu, S.; Tomar, A.; Nam, J.-O.; Luo, H.; Phillips, A.; Patel, N.; Kwok, C.; McMahon, G.; et al. Oral Delivery of PND-1186 FAK Inhibitor Decreases Tumor Growth and Spontaneous Breast to Lung Metastasis in Pre-Clinical Models. Cancer Biol. Ther. 2010, 9, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Wendt, M.K.; Schiemann, W.P. Therapeutic Targeting of the Focal Adhesion Complex Prevents Oncogenic TGF-Beta Signaling and Metastasis. Breast Cancer Res. 2009, 11, R68. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, R.A.; Gutknecht, M.F.; Thomas, K.S.; Conaway, M.R.; Bouton, A.H. Focal Adhesion Kinase (FAK) Deficiency in Mononuclear Phagocytes Alters Murine Breast Tumor Progression. Am. J. Cancer Res. 2018, 8, 675–687. [Google Scholar]
- Canel, M.; Taggart, D.; Sims, A.H.; Lonergan, D.W.; Waizenegger, I.C.; Serrels, A. T-Cell Co-Stimulation in Combination with Targeting FAK Drives Enhanced Anti-Tumor Immunity. eLife 2020, 9, e48092. [Google Scholar] [CrossRef]
- Serrels, A.; Lund, T.; Serrels, B.; Byron, A.; McPherson, R.C.; von Kriegsheim, A.; Gómez-Cuadrado, L.; Canel, M.; Muir, M.; Ring, J.E.; et al. Nuclear FAK Controls Chemokine Transcription, Tregs, and Evasion of Anti-Tumor Immunity. Cell 2015, 163, 160–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- German, A.E.; Mammoto, T.; Jiang, E.; Ingber, D.E.; Mammoto, A. Paxillin Controls Endothelial Cell Migration and Tumor Angiogenesis by Altering Neuropilin 2 Expression. J. Cell Sci. 2014, 127, 1672–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Zhan, Y.; Huynh, N.; Dumesny, C.; Wang, X.; Asadi, K.; Herrmann, D.; Timpson, P.; Yang, Y.; Walsh, K.; et al. Inhibition of PAK1 Suppresses Pancreatic Cancer by Stimulation of Anti-Tumour Immunity through down-Regulation of PD-L1. Cancer Lett. 2020, 472, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Huynh, N.; Wang, K.; Yim, M.; Dumesny, C.J.; Sandrin, M.S.; Baldwin, G.S.; Nikfarjam, M.; He, H. Depletion of P21-Activated Kinase 1 up-Regulates the Immune System of APC∆14/+ Mice and Inhibits Intestinal Tumorigenesis. BMC Cancer 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pedros, C.; Canonigo-Balancio, A.J.; Kong, K.-F.; Altman, A. Requirement of Treg-Intrinsic CTLA4/PKCη Signaling Pathway for Suppressing Tumor Immunity. JCI Insight 2017, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluskota, E.; Dowling, J.J.; Gordon, N.; Golden, J.A.; Szpak, D.; West, X.Z.; Nestor, C.; Ma, Y.-Q.; Bialkowska, K.; Byzova, T.; et al. The Integrin Coactivator Kindlin-2 Plays a Critical Role in Angiogenesis in Mice and Zebrafish. Blood 2011, 117, 4978–4987. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Masamune, A.; Hamada, S.; Kikuta, K.; Takikawa, T.; Motoi, F.; Unno, M.; Shimosegawa, T. Kindlin-2 in Pancreatic Stellate Cells Promotes the Progression of Pancreatic Cancer. Cancer Lett. 2017, 390, 103–114. [Google Scholar] [CrossRef]
- Marcovecchio, P.M.; Zhu, Y.P.; Hanna, R.N.; Dinh, H.Q.; Tacke, R.; Wu, R.; McArdle, S.; Reynolds, S.; Araujo, D.J.; Ley, K.; et al. Frontline Science: Kindlin-3 Is Essential for Patrolling and Phagocytosis Functions of Nonclassical Monocytes during Metastatic Cancer Surveillance. J. Leukoc. Biol. 2020, 107, 883–892. [Google Scholar] [CrossRef]
- Pulous, F.E.; Carnevale, J.C.; Al-Yafeai, Z.; Pearson, B.H.; Hamilton, J.A.G.; Henry, C.J.; Wayne Orr, A.; Petrich, B.G. Talin-Dependent Integrin Activation Is Required for Endothelial Proliferation and Postnatal Angiogenesis. Angiogenesis 2020, 1–14. [Google Scholar] [CrossRef]
- Younes, M.N. Integrin-Linked Kinase Is a Potential Therapeutic Target for Anaplastic Thyroid Cancer. Mol. Cancer Ther. 2005, 4, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Yau, C.Y.F.; Wheeler, J.J.; Sutton, K.L.; Hedley, D.W. Inhibition of Integrin-Linked Kinase by a Selective Small Molecule Inhibitor, QLT0254, Inhibits the PI3K/PKB/MTOR, Stat3, and FKHR Pathways and Tumor Growth, and Enhances Gemcitabine-Induced Apoptosis in Human Orthotopic Primary Pancreatic Cancer Xenografts. Cancer Res. 2005, 65, 1497–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, L.A.; Woo, J.; Huxham, L.A.; Verreault, M.; Dragowska, W.H.; Chiu, G.; Rajput, A.; Kyle, A.H.; Kalra, J.; Yapp, D.; et al. Suppression of VEGF Secretion and Changes in Glioblastoma Multiforme Microenvironment by Inhibition of Integrin-Linked Kinase (ILK). Mol. Cancer Ther. 2008, 7, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.; Palmby, T.R.; Gavard, J.; Amornphimoltham, P.; Zheng, Y.; Gui, J.S. An Essential Role for Rac1 in Endothelial Cell Function and Vascular Development. FASEB J. 2008, 22, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, S.; García-Mata, R.; Staub, J.; Valdivia, A.; Sharek, L.; McCulloch, C.; Hwang, R.; Urrutia, R.; Yeh, J.; Kim, H.; et al. Palladin Promotes Invasion of Pancreatic Cancer Cells by Enhancing Invadopodia Formation in Cancer-Associated Fibroblasts. Oncogene 2014, 33, 1265–1273. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.; Yaqoob, U.; Geng, Z.; Bloch, K.; Liu, C.; Gomez, T.; Billadeau, D.; Shah, V. Focal Adhesion Assembly in Myofibroblasts Fosters a Microenvironment That Promotes Tumor Growth. Am. J. Pathol. 2010, 177, 1888–1900. [Google Scholar] [CrossRef]
- Tu, K.; Li, J.; Verma, V.K.; Liu, C.; Billadeau, D.D.; Lamprecht, G.; Xiang, X.; Guo, L.; Dhanasekaran, R.; Roberts, L.R.; et al. VASP Promotes TGF-β Activation of Hepatic Stellate Cells by Regulating Rab11 Dependent Plasma Membrane Targeting of TGF-β Receptors. Hepatology 2015, 61, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Nallapalli, R.K.; Ibrahim, M.X.; Zhou, A.X.; Bandaru, S.; Sunkara, S.N.; Redfors, B.; Pazooki, D.; Zhang, Y.; Borén, J.; Cao, Y.; et al. Targeting Filamin A Reduces K-RAS–Induced Lung Adenocarcinomas and Endothelial Response to Tumor Growth in Mice. Mol. Cancer 2012, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, G.; Robinson, S.D.; Germain, M.; Reynolds, L.E.; Thomas, G.J.; Elia, G.; Saunders, G.; Fruttiger, M.; Tybulewicz, V.; Mavria, G.; et al. Endothelial-Rac1 Is Not Required for Tumor Angiogenesis Unless Avβ3-Integrin Is Absent. PLoS ONE 2010, 5, e9766. [Google Scholar] [CrossRef] [Green Version]
- Vader, P.; van der Meel, R.; Symons, M.H.; Fens, M.H.A.M.; Pieters, E.; Wilschut, K.J.; Storm, G.; Jarzabek, M.; Gallagher, W.M.; Schiffelers, R.M.; et al. Examining the Role of Rac1 in Tumor Angiogenesis and Growth: A Clinically Relevant RNAi-Mediated Approach. Angiogenesis 2011, 14, 457–466. [Google Scholar] [CrossRef]
- Wilson, E.; Leszczynska, K.; Poulter, N.S.; Edelmann, F.; Salisbury, V.A.; Noy, P.J.; Bacon, A.; Rappoport, J.Z.; Heath, J.K.; Bicknell, R.; et al. RhoJ Interacts with the GIT–PIX Complex and Regulates Focal Adhesion Disassembly. J. Cell Sci. 2014, 127, 3039–3051. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Yang, H.; Fukushima, Y.; Saw, P.E.; Lee, J.; Park, J.-S.; Park, I.; Jung, J.; Kataoka, H.; Lee, D.; et al. Vascular RhoJ Is an Effective and Selective Target for Tumor Angiogenesis and Vascular Disruption. Cancer Cell 2014, 25, 102–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghban, R.; Roshangar, L.; Jahanban-Esfahlan, R.; Seidi, K.; Ebrahimi-Kalan, A.; Jaymand, M.; Kolahian, S.; Javaheri, T.; Zare, P. Tumor Microenvironment Complexity and Therapeutic Implications at a Glance. Cell Commun. Signal. 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brassart-Pasco, S.; Brézillon, S.; Brassart, B.; Ramont, L.; Oudart, J.-B.; Monboisse, J.C. Tumor Microenvironment: Extracellular Matrix Alterations Influence Tumor Progression. Front. Oncol. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.-Z.; Jin, W.-L. The Updated Landscape of Tumor Microenvironment and Drug Repurposing. Signal. Transduct. Target. Ther. 2020, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Avraamides, C.J.; Garmy-Susini, B.; Varner, J.A. Integrins in Angiogenesis and Lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidi, H.; Ivaska, J. Every Step of the Way: Integrins in Cancer Progression and Metastasis. Nat. Rev. Cancer 2018, 18, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Braren, R.; Hu, H.; Kim, Y.H.; Beggs, H.E.; Reichardt, L.F.; Wang, R. Endothelial FAK Is Essential for Vascular Network Stability, Cell Survival, and Lamellipodial Formation. J. Cell Biol. 2006, 172, 151–162. [Google Scholar] [CrossRef]
- Shen, T.-L.; Park, A.Y.-J.; Alcaraz, A.; Peng, X.; Jang, I.; Koni, P.; Flavell, R.A.; Gu, H.; Guan, J.-L. Conditional Knockout of Focal Adhesion Kinase in Endothelial Cells Reveals Its Role in Angiogenesis and Vascular Development in Late Embryogenesis. J. Cell Biol. 2005, 169, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Peng, X.; Sun, S.; Park, A.Y.J.; Guan, J.-L. Role of Kinase-Independent and -Dependent Functions of FAK in Endothelial Cell Survival and Barrier Function during Embryonic Development. J. Cell Biol. 2010, 189, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Weis, S.M.; Lim, S.-T.; Lutu-Fuga, K.M.; Barnes, L.A.; Chen, X.L.; Göthert, J.R.; Shen, T.-L.; Guan, J.-L.; Schlaepfer, D.D.; Cheresh, D.A. Compensatory Role for Pyk2 during Angiogenesis in Adult Mice Lacking Endothelial Cell FAK. J. Cell Biol. 2008, 181, 43–50. [Google Scholar] [CrossRef]
- Owen, K.A.; Pixley, F.J.; Thomas, K.S.; Vicente-Manzanares, M.; Ray, B.J.; Horwitz, A.F.; Parsons, J.T.; Beggs, H.E.; Stanley, E.R.; Bouton, A.H. Regulation of Lamellipodial Persistence, Adhesion Turnover, and Motility in Macrophages by Focal Adhesion Kinase. J. Cell Biol. 2007, 179, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasorn, A.; Alcaide, P.; Jia, Y.; Subramanian, K.K.; Sarraj, B.; Li, Y.; Loison, F.; Hattori, H.; Silberstein, L.E.; Luscinskas, W.F.; et al. Focal Adhesion Kinase Regulates Pathogen-Killing Capability and Life Span of Neutrophils via Mediating Both Adhesion-Dependent and -Independent Cellular Signals. J. Immunol. 2009, 183, 1032–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanazawa, S.; Ilic, D.; Hashiyama, M.; Okada, M.; Noumura, T.; Aizawa, S.; Suda, T. Impaired Development of CD4+ CD8+ Thymocytes by Csk-’knock-in’ into Fyn Locus. Oncogene 1996, 13, 199–204. [Google Scholar] [PubMed]
- Wiemer, A.J.; Wernimont, S.A.; Cung, T.-D.; Bennin, D.A.; Beggs, H.E.; Huttenlocher, A. The Focal Adhesion Kinase Inhibitor PF-562,271 Impairs Primary CD4+ T Cell Activation. Biochem. Pharmacol. 2013, 86, 770–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Wolfram, P.; Canty, K.; Harley, B.; Nombela-Arrieta, C.; Pivarnik, G.; Manis, J.; Beggs, H.E.; Silberstein, L.E. Focal Adhesion Kinase Regulates the Localization and Retention of Pro-B Cells in Bone Marrow Microenvironments. J. Immunol. 2013, 190, 1094–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okigaki, M.; Davis, C.; Falasca, M.; Harroch, S.; Felsenfeld, D.P.; Sheetz, M.P.; Schlessinger, J. Pyk2 Regulates Multiple Signaling Events Crucial for Macrophage Morphology and Migration. PNAS 2003, 100, 10740–10745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamen, L.A.; Schlessinger, J.; Lowell, C.A. Pyk2 Is Required for Neutrophil Degranulation and Host Defense Responses to Bacterial Infection. J. Immunol. 2011, 186, 1656–1665. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, R.A.; Thomas, K.S.; Gutknecht, M.F.; Bouton, A.H. The Nonreceptor Protein Tyrosine Kinase Pyk2 Promotes the Turnover of Monocytes at Steady State. J. Leukoc. Biol. 2017, 102, 1069–1080. [Google Scholar] [CrossRef]
- Gismondi, A.; Jacobelli, J.; Strippoli, R.; Mainiero, F.; Soriani, A.; Cifaldi, L.; Piccoli, M.; Frati, L.; Santoni, A. Proline-Rich Tyrosine Kinase 2 and Rac Activation by Chemokine and Integrin Receptors Controls NK Cell Transendothelial Migration. J. Immunol. 2003, 170, 3065–3073. [Google Scholar] [CrossRef] [Green Version]
- Guinamard, R.; Okigaki, M.; Schlessinger, J.; Ravetch, J.V. Absence of Marginal Zone B Cells in Pyk-2-Deficient Mice Defines Their Role in the Humoral Response. Nat. Immunol. 2000, 1, 31–36. [Google Scholar] [CrossRef]
- Missy, K.; Hu, B.; Schilling, K.; Harenberg, A.; Sakk, V.; Kuchenbecker, K.; Kutsche, K.; Fischer, K.-D. APIX Rho GTPase Guanine Nucleotide Exchange Factor Regulates Lymphocyte Functions and Antigen Receptor Signaling. Mol. Cell. Biol. 2008, 28, 3776–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korthals, M.; Schilling, K.; Reichardt, P.; Mamula, D.; Schlüter, T.; Steiner, M.; Langnäse, K.; Thomas, U.; Gundelfinger, E.; Premont, R.T.; et al. APIX RhoGEF Supports Positive Selection by Restraining Migration and Promoting Arrest of Thymocytes. J. Immunol. 2014, 192, 3228–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, C.T.; Rossaint, J.; Moser, M.; Petrich, B.G.; Zarbock, A.; Monkley, S.J.; Critchley, D.R.; Ginsberg, M.H.; Fässler, R.; Ley, K. Distinct Roles for Talin-1 and Kindlin-3 in LFA-1 Extension and Affinity Regulation. Blood 2012, 119, 4275–4282. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.; Bauer, M.; Schmid, S.; Ruppert, R.; Schmidt, S.; Sixt, M.; Wang, H.-V.; Sperandio, M.; Fässler, R. Kindlin-3 Is Required for Beta2 Integrin-Mediated Leukocyte Adhesion to Endothelial Cells. Nat. Med. 2009, 15, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Klapproth, S.; Moretti, F.A.; Zeiler, M.; Ruppert, R.; Breithaupt, U.; Mueller, S.; Haas, R.; Mann, M.; Sperandio, M.; Fässler, R.; et al. Minimal Amounts of Kindlin-3 Suffice for Basal Platelet and Leukocyte Functions in Mice. Blood 2015, 126, 2592–2600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretti, F.A.; Moser, M.; Lyck, R.; Abadier, M.; Ruppert, R.; Engelhardt, B.; Fässler, R. Kindlin-3 Regulates Integrin Activation and Adhesion Reinforcement of Effector T Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 17005–17010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monkley, S.J.; Kostourou, V.; Spence, L.; Petrich, B.; Coleman, S.; Ginsberg, M.H.; Pritchard, C.A.; Critchley, D.R. Endothelial Cell Talin1 Is Essential for Embryonic Angiogenesis. Dev. Biol. 2011, 349, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulous, F.E.; Grimsley-Myers, C.M.; Kansal, S.; Kowalczyk, A.P.; Petrich, B.G. Talin-Dependent Integrin Activation Regulates VE-Cadherin Localization and Endothelial Cell Barrier Function. Circ. Res. 2019, 124, 891–903. [Google Scholar] [CrossRef]
- Bromberger, T.; Klapproth, S.; Rohwedder, I.; Zhu, L.; Mittmann, L.; Reichel, C.A.; Sperandio, M.; Qin, J.; Moser, M. Direct Rap1/Talin1 Interaction Regulates Platelet and Neutrophil Integrin Activity in Mice. Blood 2018, 132, 2754–2762. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wu, H.; An, J.; Ballantyne, C.M.; Cyster, J.G. Critical Role of Integrin CD11c in Splenic Dendritic Cell Capture of Missing-Self CD47 Cells to Induce Adaptive Immunity. PNAS 2018, 115, 6786–6791. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.J.F.; Bunjamin, M.; Ruedl, C.; Su, I.-H. Talin1 Controls Dendritic Cell Activation by Regulating TLR Complex Assembly and Signaling. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Mace, E.M.; Monkley, S.J.; Critchley, D.R.; Takei, F. A Dual Role for Talin in NK Cell Cytotoxicity: Activation of LFA-1-Mediated Cell Adhesion and Polarization of NK Cells. J. Immunol. 2009, 182, 948–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wernimont, S.A.; Wiemer, A.J.; Bennin, D.A.; Monkley, S.J.; Ludwig, T.; Critchley, D.R.; Huttenlocher, A. Contact-Dependent T Cell Activation and T Cell Stopping Require Talin1. J. Immunol. 2011, 187, 6256–6267. [Google Scholar] [CrossRef] [PubMed]
- Klann, J.E.; Remedios, K.A.; Kim, S.H.; Metz, P.J.; Lopez, J.; Mack, L.A.; Zheng, Y.; Ginsberg, M.H.; Petrich, B.G.; Chang, J.T. Talin Plays a Critical Role in the Maintenance of the Regulatory T Cell Pool. J. Immunol. 2017, 198, 4639–4651. [Google Scholar] [CrossRef] [Green Version]
- Manevich-Mendelson, E.; Grabovsky, V.; Feigelson, S.W.; Cinamon, G.; Gore, Y.; Goverse, G.; Monkley, S.J.; Margalit, R.; Melamed, D.; Mebius, R.E.; et al. Talin1 Is Required for Integrin-Dependent B Lymphocyte Homing to Lymph Nodes and the Bone Marrow but Not for Follicular B-Cell Maturation in the Spleen. Blood 2010, 116, 5907–5918. [Google Scholar] [CrossRef] [Green Version]
- Wilson, Z.S.; Witt, H.; Hazlett, L.; Harman, M.; Neumann, B.M.; Whitman, A.; Patel, M.; Ross, R.S.; Franck, C.; Reichner, J.S.; et al. Context-Dependent Role of Vinculin in Neutrophil Adhesion, Motility and Trafficking. Sci. Rep. 2020, 10, 2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, E.B.; Liu, E.; Sinha, S.; Cook, S.; Milstone, D.S.; MacRae, C.A.; Mariotti, M.; Kuhlencordt, P.J.; Force, T.; Rosenzweig, A.; et al. Integrin-Linked Kinase Regulates Endothelial Cell Survival and Vascular Development. Mol. Cell. Biol. 2004, 24, 8134–8144. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Yamamoto, H.; Mohn, L.; Ambühl, L.; Kanai, K.; Schmidt, I.; Kim, K.-P.; Fraccaroli, A.; Feil, S.; Junge, H.J.; et al. Integrin-Linked Kinase Controls Retinal Angiogenesis and Is Linked to Wnt Signaling and Exudative Vitreoretinopathy. Nat. Commun. 2019, 10, 5243. [Google Scholar] [CrossRef]
- Kogata, N.; Tribe, R.M.; Fässler, R.; Way, M.; Adams, R.H. Integrin-Linked Kinase Controls Vascular Wall Formation by Negatively Regulating Rho/ROCK-Mediated Vascular Smooth Muscle Cell Contraction. Genes Dev. 2009, 23, 2278–2283. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Sinha, S.; Williams, C.; Cyrille, M.; Heller, E.; Snapper, S.B.; Georgopoulos, K.; St-Arnaud, R.; Force, T.; Dedhar, S.; et al. Targeted Deletion of Integrin-Linked Kinase Reveals a Role in T-Cell Chemotaxis and Survival. Mol. Cell Biol. 2005, 25, 11145–11155. [Google Scholar] [CrossRef] [Green Version]
- Fraccaroli, A.; Pitter, B.; Taha, A.A.; Seebach, J.; Huveneers, S.; Kirsch, J.; Casaroli-Marano, R.P.; Zahler, S.; Pohl, U.; Gerhardt, H.; et al. Endothelial Alpha-Parvin Controls Integrity of Developing Vasculature and Is Required for Maintenance of Cell-Cell Junctions. Circ. Res. 2015, 117, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Pitter, B.; Werner, A.-C.; Montanez, E. Parvins Are Required for Endothelial Cell-Cell Junctions and Cell Polarity During Embryonic Blood Vessel Formation. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1147–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montanez, E.; Wickström, S.A.; Altstätter, J.; Chu, H.; Fässler, R. α-Parvin Controls Vascular Mural Cell Recruitment to Vessel Wall by Regulating RhoA/ROCK Signalling. EMBO J. 2009, 28, 3132–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.; Thievessen, I.; Sixt, M.; Lämmermann, T.; Waisman, A.; Braun, A.; Noegel, A.A.; Fässler, R. γ-Parvin Is Dispensable for Hematopoiesis, Leukocyte Trafficking, and T-Cell-Dependent Antibody Response. Mol. Cell Biol. 2006, 26, 1817–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, K.; Diehl, L.; Rommerscheidt-Fuss, U.; Golletz, C.; Quast, T.; Kahl, P.; Kolanus, W.; Knolle, P.; Buettner, R.; Heukamp, L.C. Four-and-a-Half LIM Domain Protein 2 Is a Novel Regulator of Sphingosine 1-Phosphate Receptor 1 in CCL19-Induced Dendritic Cell Migration. J. Immunol. 2010, 185, 1466–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, Y.-P.; Sun, P.; Wang, A.; Lo, S.H. Tensin1 Positively Regulates RhoA Activity through Its Interaction with DLC1. Biochim. Biophys. Acta Mol. Cell Res. 2015, 1853, 3258–3265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, C.; Sieminski, A.L.; Kwiatkowski, A.V.; Rubinson, D.A.; Vasile, E.; Bronson, R.T.; Fässler, R.; Gertler, F.B. Ena/VASP Is Required for Endothelial Barrier Function in Vivo. J. Cell Biol. 2007, 179, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Estin, M.L.; Thompson, S.B.; Traxinger, B.; Fisher, M.H.; Friedman, R.S.; Jacobelli, J. Ena/VASP Proteins Regulate Activated T-Cell Trafficking by Promoting Diapedesis during Transendothelial Migration. PNAS 2017, 114, E2901–E2910. [Google Scholar] [CrossRef] [Green Version]
- Deevi, R.K.; Koney-Dash, M.; Kissenpfennig, A.; Johnston, J.A.; Schuh, K.; Walter, U.; Dib, K. Vasodilator-Stimulated Phosphoprotein Regulates Inside-Out Signaling of Β2 Integrins in Neutrophils. J. Immunol. 2010, 184, 6575–6584. [Google Scholar] [CrossRef] [Green Version]
- Savinko, T.; Guenther, C.; Uotila, L.M.; Llort Asens, M.; Yao, S.; Tojkander, S.; Fagerholm, S.C. Filamin A Is Required for Optimal T Cell Integrin-Mediated Force Transmission, Flow Adhesion, and T Cell Trafficking. J. Immunol. 2018, 200, 3109–3116. [Google Scholar] [CrossRef] [Green Version]
- Uotila, L.M.; Guenther, C.; Savinko, T.; Lehti, T.A.; Fagerholm, S.C. Filamin A Regulates Neutrophil Adhesion, Production of Reactive Oxygen Species, and Neutrophil Extracellular Trap Release. J. Immunol. 2017, 199, 3644–3653. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Forster, C.; Nakamura, F.; Glogauer, M. Filamin-A Regulates Neutrophil Uropod Retraction through RhoA during Chemotaxis. PLoS ONE 2013, 8, e79009. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Tian, F.; Sandzen, J.; Cao, R.; Flaberg, E.; Szekely, L.; Cao, Y.; Ohlsson, C.; Bergo, M.O.; Boren, J.; et al. Filamin B Deficiency in Mice Results in Skeletal Malformations and Impaired Microvascular Development. Proc. Natl. Acad. Sci. USA 2007, 104, 3919–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, D.M.; Xu, K.; Meadows, S.M.; Zheng, Y.; Norden, P.R.; Davis, G.E.; Cleaver, O. Cdc42 Is Required for Cytoskeletal Support of Endothelial Cell Adhesion during Blood Vessel Formation in Mice. Development 2015, 142, 3058–3070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Liu, Y.; Lin, Q.; Li, J.; Druso, J.E.; Antonyak, M.A.; Meininger, C.J.; Zhang, S.L.; Dostal, D.E.; Guan, J.-L.; et al. Deletion of Cdc42 Enhances ADAM17-Mediated Vascular Endothelial Growth Factor Receptor 2 Shedding and Impairs Vascular Endothelial Cell Survival and Vasculogenesis. Mol. Cell. Biol. 2013, 33, 4181–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Zhang, S.; Tripathi, P.; Mattner, J.; Phelan, J.; Sproles, A.; Mo, J.; Wills-Karp, M.; Grimes, H.L.; Hildeman, D.; et al. Distinct Roles of Cdc42 in Thymopoiesis and Effector and Memory T Cell Differentiation. PLoS ONE 2011, 6, e18002. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Hildeman, D.; Tripathi, P.; Velu, C.S.; Grimes, H.L.; Zheng, Y. Coordination of IL-7 Receptor and T-Cell Receptor Signaling by Cell-Division Cycle 42 in T-Cell Homeostasis. Proc. Natl. Acad. Sci. USA 2010, 107, 18505–18510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nohata, N.; Uchida, Y.; Stratman, A.; Adams, R.; Zheng, Y.; Weinstein, B.; Mukouyama, Y.; Gutkind, J.S. Temporal-Specific Roles of Rac1 during Vascular Development and Retinal Angiogenesis. Dev. Biol. 2016, 411, 183–194. [Google Scholar] [CrossRef]
- Wells, C.M.; Walmsley, M.; Ooi, S.; Tybulewicz, V.; Ridley, A.J. Rac1-Deficient Macrophages Exhibit Defects in Cell Spreading and Membrane Ruffling but Not Migration. J. Cell Sci. 2004, 117, 1259–1268. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, A.P.; Wells, C.M.; Smith, S.D.; Vega, F.M.; Henderson, R.B.; Tybulewicz, V.L.; Ridley, A.J. Rac1 and Rac2 Regulate Macrophage Morphology but Are Not Essential for Migration. J. Cell Sci. 2006, 119, 2749–2757. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Cancelas, J.A.; Hildeman, D.; Williams, D.A.; Zheng, Y. Rac GTPase Isoforms Rac1 and Rac2 Play a Redundant and Crucial Role in T-Cell Development. Blood 2008, 112, 1767–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, C.; Corsoni-Tadrzak, A.; Ruf, S.; de Boer, J.; Williams, A.; Turner, M.; Kioussis, D.; Tybulewicz, V.L.J. Rac GTPases Play Critical Roles in Early T-Cell Development. Blood 2009, 113, 3990–3998. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, M.J.; Ooi, S.K.T.; Reynolds, L.F.; Smith, S.H.; Ruf, S.; Mathiot, A.; Vanes, L.; Williams, D.A.; Cancro, M.P.; Tybulewicz, V.L.J. Critical Roles for Rac1 and Rac2 GTPases in B Cell Development and Signaling. Science 2003, 302, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Croker, B.A.; Tarlinton, D.M.; Cluse, L.A.; Tuxen, A.J.; Light, A.; Yang, F.-C.; Williams, D.A.; Roberts, A.W. The Rac2 Guanosine Triphosphatase Regulates B Lymphocyte Antigen Receptor Responses and Chemotaxis and Is Required for Establishment of B-1a and Marginal Zone B Lymphocytes. J. Immunol. 2002, 168, 3376–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faroudi, M.; Hons, M.; Zachacz, A.; Dumont, C.; Lyck, R.; Stein, J.V.; Tybulewicz, V.L.J. Critical Roles for Rac GTPases in T-Cell Migration to and within Lymph Nodes. Blood 2010, 116, 5536–5547. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.B.; Grys, K.; Vehlow, A.; de Bettignies, C.; Zachacz, A.; Henley, T.; Turner, M.; Batista, F.; Tybulewicz, V.L.J. A Novel Rac-Dependent Checkpoint in B Cell Development Controls Entry into the Splenic White Pulp and Cell Survival. J. Exp. Med. 2010, 207, 837–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahra, F.T.; Sajib, M.S.; Ichiyama, Y.; Akwii, R.G.; Tullar, P.E.; Cobos, C.; Minchew, S.A.; Doçi, C.L.; Zheng, Y.; Kubota, Y.; et al. Endothelial RhoA GTPase Is Essential for in Vitro Endothelial Functions but Dispensable for Physiological in Vivo Angiogenesis. Sci. Rep. 2019, 9, 11666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manresa-Arraut, A.; Johansen, F.F.; Brakebusch, C.; Issazadeh-Navikas, S.; Hasseldam, H. RhoA Drives T-Cell Activation and Encephalitogenic Potential in an Animal Model of Multiple Sclerosis. Front. Immunol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhou, X.; Lang, R.A.; Guo, F. RhoA of the Rho Family Small GTPases Is Essential for B Lymphocyte Development. PLoS ONE 2012, 7, e33773. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, Y.; Nishiyama, K.; Kataoka, H.; Fruttiger, M.; Fukuhara, S.; Nishida, K.; Mochizuki, N.; Kurihara, H.; Nishikawa, S.-I.; Uemura, A. RhoJ Integrates Attractive and Repulsive Cues in Directional Migration of Endothelial Cells. EMBO J. 2020, 39, 1–18. [Google Scholar] [CrossRef]
- Hakem, A. RhoC Is Dispensable for Embryogenesis and Tumor Initiation but Essential for Metastasis. Genes Dev. 2005, 19, 1974–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigorito, E.; Bell, S.; Hebeis, B.J.; Reynolds, H.; McAdam, S.; Emson, P.C.; McKenzie, A.; Turner, M. Immunological Function in Mice Lacking the Rac-Related GTPase RhoG. Mol. Cell Biol. 2004, 24, 719–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, A.S.; Lee, J.; Ferrara, N. Targeting the Tumour Vasculature: Insights from Physiological Angiogenesis. Nat. Rev. Cancer 2010, 10, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Jain, R.K. Molecular Mechanisms and Clinical Applications of Angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the Angiogenic Switch. Nat. Rev. Cancer 2003, 3, 401–410. [Google Scholar] [CrossRef]
- Jain, R.K. Normalization of Tumor Vasculature: An Emerging Concept in Antiangiogenic Therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef]
- Nagy, J.A.; Chang, S.-H.; Dvorak, A.M.; Dvorak, H.F. Why Are Tumour Blood Vessels Abnormal and Why Is It Important to Know? Br. J. Cancer 2009, 100, 865–869. [Google Scholar] [CrossRef]
- Polte, T.R.; Hanks, S.K.; Naftilan, A.J. Focal Adhesion Kinase Is Abundant in Developing Blood Vessels and Elevation of Its Phosphotyrosine Content in Vascular Smooth Muscle Cells Is a Rapid Response to Angiotensin II. J. Cell. Biochem. 1994, 55, 106–119. [Google Scholar] [CrossRef]
- Peng, X.; Ueda, H.; Zhou, H.; Stokol, T.; Shen, T.; Alcaraz, A.; Nagy, T.; Vassalli, J.; Guan, J. Overexpression of Focal Adhesion Kinase in Vascular Endothelial Cells Promotes Angiogenesis in Transgenic Mice. Cardiovasc. Res. 2004, 64, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Corsi, J.-M.; Houbron, C.; Billuart, P.; Brunet, I.; Bouvrée, K.; Eichmann, A.; Girault, J.-A.; Enslen, H. Autophosphorylation-Independent and -Dependent Functions of Focal Adhesion Kinase during Development. J. Biol. Chem. 2009, 284, 34769–34776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.-T.; Chen, X.L.; Tomar, A.; Miller, N.L.G.; Yoo, J.; Schlaepfer, D.D. Knock-in Mutation Reveals an Essential Role for Focal Adhesion Kinase Activity in Blood Vessel Morphogenesis and Cell Motility-Polarity but Not Cell Proliferation. J. Biol. Chem. 2010, 285, 21526–21536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavora, B.; Reynolds, L.E.; Batista, S.; Demircioglu, F.; Fernandez, I.; Lechertier, T.; Lees, D.M.; Wong, P.; Alexopoulou, A.; Elia, G.; et al. Endothelial–Cell FAK Targeting Sensitizes Tumours to DNA–Damaging Therapy. Nature 2014, 514, 112–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.Y.; Timpson, P.; Horvath, L.G.; Daly, R.J. FAK Signaling in Human Cancer as a Target for Therapeutics. Pharmacol. Ther. 2015, 146, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Schwock, J.; Dhani, N.; Hedley, D.W. Targeting Focal Adhesion Kinase Signaling in Tumor Growth and Metastasis. Expert Opin. Ther. Targets 2010, 14, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in Cancer: Mechanistic Findings and Clinical Applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yi, Q.; Tang, L. The Roles of Nuclear Focal Adhesion Kinase (FAK) on Cancer: A Focused Review. J. Exp. Clin. Cancer Res. 2019, 38, 250. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Nam, J.-O.; Jean, C.; Lawson, C.; Walsh, C.T.; Goka, E.; Lim, S.-T.; Tomar, A.; Tancioni, I.; Uryu, S.; et al. VEGF-Induced Vascular Permeability Is Mediated by FAK. Dev. Cell 2012, 22, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Cruet-Hennequart, S.; Troussard, A.; Fazli, L.; Costello, P.; Sutton, K.; Wheeler, J.; Gleave, M.; Sanghera, J.; Dedhar, S. Regulation of Tumor Angiogenesis by Integrin-Linked Kinase (ILK). Cancer Cell 2004, 5, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Pluskota, E.; Bledzka, K.M.; Bialkowska, K.; Szpak, D.; Soloviev, D.A.; Jones, S.V.; Verbovetskiy, D.; Plow, E.F. Kindlin-2 Interacts with Endothelial Adherens Junctions to Support Vascular Barrier Integrity: Kindlin-2 Regulates Endothelial Barier Function. J. Physiol. 2017, 595, 6443–6462. [Google Scholar] [CrossRef] [Green Version]
- Legate, K.R.; Montañez, E.; Kudlacek, O.; Fässler, R. ILK, PINCH and Parvin: The TIPP of Integrin Signalling. Nat. Rev. Mol. Cell Biol. 2006, 7, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Koul, D.; Shen, R.; Bergh, S.; Lu, Y.; de Groot, J.F.; Liu, T.J.; Mills, G.B.; Yung, W.K.A. Targeting Integrin-Linked Kinase Inhibits Akt Signaling Pathways and Decreases Tumor Progression of Human Glioblastoma. Mol. Cancer Ther. 2005, 4, 1681–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, A.A.; Jafarnejad, S.M.; Zhou, J.; Li, G. Integrin-Linked Kinase Regulates Melanoma Angiogenesis by Activating NF-ΚB/Interleukin-6 Signaling Pathway. Oncogene 2011, 30, 2778–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malan, D.; Elischer, A.; Hesse, M.; Wickström, S.A.; Fleischmann, B.K.; Bloch, W. Deletion of Integrin Linked Kinase in Endothelial Cells Results in Defective RTK Signaling Caused by Caveolin 1 Mislocalization. Development 2013, 140, 987–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vouret-Craviari, V.; Boulter, E.; Grall, D.; Matthews, C.; Obberghen-Schilling, E.V. ILK Is Required for the Assembly of Matrix-Forming Adhesions and Capillary Morphogenesis in Endothelial Cells. J. Cell Sci. 2004, 117, 4559–4569. [Google Scholar] [CrossRef] [Green Version]
- Huveneers, S.; Oldenburg, J.; Spanjaard, E.; van der Krogt, G.; Grigoriev, I.; Akhmanova, A.; Rehmann, H.; de Rooij, J. Vinculin Associates with Endothelial VE-Cadherin Junctions to Control Force-Dependent Remodeling. J. Cell Biol. 2012, 196, 641–652. [Google Scholar] [CrossRef] [Green Version]
- Birukova, A.A.; Shah, A.S.; Tian, Y.; Moldobaeva, N.; Birukov, K.G. Dual Role of Vinculin in Barrier-Disruptive and Barrier-Enhancing Endothelial Cell Responses. Cell. Signal. 2016, 28, 541–551. [Google Scholar] [CrossRef]
- Nobes, C.D.; Hall, A. Rho, Rac, and Cdc42 GTPases Regulate the Assembly of Multimolecular Focal Complexes Associated with Actin Stress Fibers, Lamellipodia, and Filopodia. Cell 1995, 81, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Wojciak-Stothard, B.; Ridley, A. Rho GTPases and the Regulation of Endothelial Permeability. Vasc. Pharmacol. 2002, 39, 187–199. [Google Scholar] [CrossRef]
- Haga, R.B.; Ridley, A.J. Rho GTPases: Regulation and Roles in Cancer Cell Biology. Small GTPases 2016, 7, 207–221. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, G.; Jones, D.T.; Nye, E.; Sapienza, K.; Ramjuan, A.R.; Reynolds, L.E.; Robinson, S.D.; Kostourou, V.; Martinez, D.; Aubyn, D.; et al. Regulation of Lymphatic-Blood Vessel Separation by Endothelial Rac1. Development 2009, 136, 4043–4053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leszczynska, K.; Kaur, S.; Wilson, E.; Bicknell, R.; Heath, V. The Role of RhoJ in Endothelial Cell Biology and Angiogenesis. Biochem. Soc. Trans. 2011, 39, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.-T.; Li, G.; Xiao, H.-T. The Role of RhoJ in Endothelial Cell Biology and Tumor Pathology. Biomed. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Leszczynska, K.; Abraham, S.; Scarcia, M.; Hiltbrunner, S.; Marshall, C.; Mavria, G.; Bicknell, R.; Heath, V. RhoJ/TCL Regulates Endothelial Motility and Tube Formation and Modulates Actomyosin Contractility and Focal Adhesion Numbers. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Sacharidou, A.; Stratman, A.N.; Le Bras, A.; Zwiers, P.J.; Spokes, K.; Bhasin, M.; Shih, S.-C.; Nagy, J.A.; Molema, G.; et al. RhoJ Is an Endothelial Cell-Restricted Rho GTPase That Mediates Vascular Morphogenesis and Is Regulated by the Transcription Factor ERG. Blood 2011, 118, 1145–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Valle-Pérez, B.; Martínez, V.G.; Lacasa-Salavert, C.; Figueras, A.; Shapiro, S.S.; Takafuta, T.; Casanovas, O.; Capellà, G.; Ventura, F.; Viñals, F. Filamin B Plays a Key Role in Vascular Endothelial Growth Factor-Induced Endothelial Cell Motility through Its Interaction with Rac-1 and Vav-2. J. Biol. Chem. 2010, 285, 10748–10760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, S.H.; Yu, Q.-C.; Degenstein, L.; Chen, L.B.; Fuchs, E. Progressive Kidney Degeneration in Mice Lacking Tensin. J. Cell Biol. 1997, 136, 1349–1361. [Google Scholar] [CrossRef] [Green Version]
- Yoshigi, M.; Hoffman, L.M.; Jensen, C.C.; Yost, H.J.; Beckerle, M.C. Mechanical Force Mobilizes Zyxin from Focal Adhesions to Actin Filaments and Regulates Cytoskeletal Reinforcement. J. Cell Biol. 2005, 171, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Li, P.; Yang, Z.; Huang, X.; Wei, G.; Sun, Y.; Kang, X.; Hu, X.; Deng, Q.; Chen, L.; et al. Zyxin Regulates Endothelial von Willebrand Factor Secretion by Reorganizing Actin Filaments around Exocytic Granules. Nat. Commun. 2017, 8, 14639. [Google Scholar] [CrossRef]
- Wójtowicz, A.; Babu, S.S.; Li, L.; Gretz, N.; Hecker, M.; Cattaruzza, M. Zyxin Mediation of Stretch-Induced Gene Expression in Human Endothelial Cells. Circ. Res. 2010, 107, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Hubbi, M.E.; Gilkes, D.M.; Baek, J.H.; Semenza, G.L. Four-and-a-Half LIM Domain Proteins Inhibit Transactivation by Hypoxia-Inducible Factor 1. J. Biol. Chem. 2012, 287, 6139–6149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Qin, X.; Zhu, Z.; Mu, J.; Zhu, L.; Wu, K.; Jiao, H.; Xu, X.; Ye, Q. FHL Family Members Suppress Vascular Endothelial Growth Factor Expression through Blockade of Dimerization of HIF1α and HIF1β. IUBMB Life 2012, 64, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimian, T.; Arfa, O.; Simeone, S.; Lemarié, C.A.; Lehoux, S.; Wassmann, S. Inhibition of Four-and-a-Half LIM Domain Protein-2 Increases Survival, Migratory Capacity, and Paracrine Function of Human Early Outgrowth Cells Through Activation of the Sphingosine Kinase-1 Pathway: Implications for Endothelial Regeneration. Circ. Res. 2014, 114, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Nakagami, H.; Takami, Y.; Koriyama, H.; Mori, M.; Tamai, K.; Sun, J.; Nagao, K.; Morishita, R.; Kaneda, Y. FHL-2 Suppresses VEGF-Induced Phosphatidylinositol 3-Kinase/Akt Activation via Interaction With Sphingosine Kinase-1. ATVB 2009, 29, 909–914. [Google Scholar] [CrossRef] [Green Version]
- Chu, P.-H.; Yeh, L.-K.; Lin, H.-C.; Jung, S.-M.; Ma, D.H.-K.; Wang, I.-J.; Wu, H.-H.; Shiu, T.-F.; Chen, J. Deletion of the FHL2 Gene Attenuating Neovascularization after Corneal Injury. Invest. Ophthalmol. Vis. Sci. 2008, 49, 5314. [Google Scholar] [CrossRef]
- Huang, P.-H.; Chen, C.-Y.; Lin, C.-P.; Wang, C.-H.; Tsai, H.-Y.; Lo, W.-Y.; Leu, H.-B.; Chen, J.-W.; Lin, S.-J.; Chu, P.-H. Deletion of FHL2 Gene Impaired Ischemia-Induced Blood Flow Recovery by Modulating Circulating Proangiogenic Cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Deakin, N.O.; Turner, C.E. Paxillin Comes of Age. J. Cell Sci. 2008, 121, 2435–2444. [Google Scholar] [CrossRef] [Green Version]
- López-Colomé, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; López, E. Paxillin: A Crossroad in Pathological Cell Migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.E. Paxillin and Focal Adhesion Signalling. Nat. Cell Biol. 2000, 2, E231–E236. [Google Scholar] [CrossRef]
- Nayal, A.; Webb, D.J.; Brown, C.M.; Schaefer, E.M.; Vicente-Manzanares, M.; Horwitz, A.R. Paxillin Phosphorylation at Ser273 Localizes a GIT1-PIX-PAK Complex and Regulates Adhesion and Protrusion Dynamics. J. Cell Biol. 2006, 173, 587–589. [Google Scholar] [CrossRef]
- Nishiya, N.; Kiosses, W.B.; Han, J.; Ginsberg, M.H. An A4 Integrin–Paxillin–Arf-GAP Complex Restricts Rac Activation to the Leading Edge of Migrating Cells. Nat. Cell Biol. 2005, 7, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.A.; Sharma, R.; Roccamatisi, D.L.; Zhang, H.; Petri, B.; Kubes, P.; Colarusso, P.; Patel, K.D. Endothelial Paxillin and Focal Adhesion Kinase (FAK) Play a Critical Role in Neutrophil Transmigration: Innate Immunity. Eur. J. Immunol. 2012, 42, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Usatyuk, P.V.; Lele, A.; Harijith, A.; Gregorio, C.C.; Garcia, J.G.N.; Salgia, R.; Natarajan, V. C-Abl Mediated Tyrosine Phosphorylation of Paxillin Regulates LPS-Induced Endothelial Dysfunction and Lung Injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L1025–L1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, P.; Usatyuk, P.V.; Jacobson, J.; Cress, A.E.; Garcia, J.G.N.; Salgia, R.; Natarajan, V. Role Played by Paxillin and Paxillin Tyrosine Phosphorylation in Hepatocyte Growth Factor/Sphingosine-1-Phosphate-Mediated Reactive Oxygen Species Generation, Lamellipodia Formation, and Endothelial Barrier Function. Pulm. Circ. 2015, 5, 619–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, R.; Bell, S.E.; Davis, G.E. Coordinate Induction of the Actin Cytoskeletal Regulatory Proteins Gelsolin, Vasodilator-Stimulated Phosphoprotein, and Profilin during Capillary Morphogenesisin Vitro. Exp. Cell Res. 1999, 249, 22–32. [Google Scholar] [CrossRef]
- Price, C.J.; Brindle, N.P.J. Vasodilator-Stimulated Phosphoprotein Is Involved in Stress-Fiber and Membrane Ruffle Formation in Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2051–2056. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, N.; Burger, S.; Golenhofen, N.; Walter, U.; Drenckhahn, D.; Waschke, J. The Role of VASP in Regulation of CAMP- and Rac 1-Mediated Endothelial Barrier Stabilization. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, C178–C188. [Google Scholar] [CrossRef]
- Schmit, M.A.; Mirakaj, V.; Stangassinger, M.; König, K.; Köhler, D.; Rosenberger, P. Vasodilator Phosphostimulated Protein (VASP) Protects Endothelial Barrier Function During Hypoxia. Inflammation 2012, 35, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Sporbert, A.; Mertsch, K.; Smolenski, A.; Haseloff, R.F.; Schönfelder, G.; Paul, M.; Ruth, P.; Walter, U.; Blasig, I.E. Phosphorylation of Vasodilator-Stimulated Phosphoprotein: A Consequence of Nitric Oxide- and CGMP-Mediated Signal Transduction in Brain Capillary Endothelial Cells and Astrocytes. Mol. Brain Res. 1999, 67, 258–266. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, S.; Baluk, P.; Kaidoh, T.; Haskell, A.; Jain, R.K.; McDonald, D.M. Abnormalities in Pericytes on Blood Vessels and Endothelial Sprouts in Tumors. Am. J. Pathol. 2002, 160, 985–1000. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.-P.; Muñoz-Félix, J.M.; Hijazi, M.; Kim, H.; Robinson, S.D.; De Luxán-Delgado, B.; Rodríguez-Hernández, I.; Maiques, O.; Meng, Y.-M.; Meng, Q.; et al. Cancer Burden Is Controlled by Mural Cell-Β3-Integrin Regulated Crosstalk with Tumor Cells. Cell 2020, 181, 1346–1363.e21. [Google Scholar] [CrossRef]
- Er, E.E.; Valiente, M.; Ganesh, K.; Zou, Y.; Agrawal, S.; Hu, J.; Griscom, B.; Rosenblum, M.; Boire, A.; Brogi, E.; et al. Pericyte-like Spreading by Disseminated Cancer Cells Activates YAP and MRTF for Metastatic Colonization. Nat. Cell Biol. 2018, 20, 966–978. [Google Scholar] [CrossRef]
- Kotecki, M.; Zeiger, A.; Van Vliet, K.; Herman, I. Calpain- and Talin-Dependent Control of Microvascular Pericyte Contractility and Cellular Stiffness. Microvasc. Res. 2010, 80, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Kollar, B.; Nahar, T.; Suresh Babu, S.; Wojtowicz, A.; Sticht, C.; Gretz, N.; Wagner, A.H.; Korff, T.; Hecker, M. Loss of the Mechanotransducer Zyxin Promotes a Synthetic Phenotype of Vascular Smooth Muscle Cells. J. Am. Heart Assoc. 2015, 4, e001712. [Google Scholar] [CrossRef] [Green Version]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A Framework for Advancing Our Understanding of Cancer-Associated Fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Houthuijzen, J.M.; Jonkers, J. Cancer-Associated Fibroblasts as Key Regulators of the Breast Cancer Tumor Microenvironment. Cancer Metastasis Rev. 2018, 37, 577–597. [Google Scholar] [CrossRef]
- Yin, Z.; Dong, C.; Jiang, K.; Xu, Z.; Li, R.; Guo, K.; Shao, S.; Wang, L. Heterogeneity of Cancer-Associated Fibroblasts and Roles in the Progression, Prognosis, and Therapy of Hepatocellular Carcinoma. J. Hematol. Oncol. 2019, 12, 1–9. [Google Scholar] [CrossRef]
- Kobayashi, H.; Enomoto, A.; Woods, S.L.; Burt, A.D.; Takahashi, M.; Worthley, D.L. Cancer-Associated Fibroblasts in Gastrointestinal Cancer. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 282–295. [Google Scholar] [CrossRef]
- Pereira, B.A.; Vennin, C.; Papanicolaou, M.; Chambers, C.R.; Herrmann, D.; Morton, J.P.; Cox, T.R.; Timpson, P. CAF Subpopulations: A New Reservoir of Stromal Targets in Pancreatic Cancer. Trends Cancer 2019, 5, 724–741. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R. The Biology and Function of Fibroblasts in Cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef]
- Kalluri, R.; Zeisberg, M. Fibroblasts in Cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Chen, X.; Song, E. Turning Foes to Friends: Targeting Cancer-Associated Fibroblasts. Nature Rev. Drug Discov. 2019, 18, 99–115. [Google Scholar] [CrossRef]
- Gieniec, K.A.; Butler, L.M.; Worthley, D.L.; Woods, S.L. Cancer-Associated Fibroblasts—Heroes or Villains? Br. J. Cancer 2019, 121, 293–302. [Google Scholar] [CrossRef]
- Shiga, K.; Hara, M.; Nagasaki, T.; Sato, T.; Takahashi, H.; Takeyama, H. Cancer-Associated Fibroblasts: Their Characteristics and Their Roles in Tumor Growth. Cancers Basel 2015, 7, 2443–2458. [Google Scholar] [CrossRef]
- LeBleu, V.S.; Kalluri, R. A Peek into Cancer-Associated Fibroblasts: Origins, Functions and Translational Impact. Dis. Models Mech. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Navab, R.; Strumpf, D.; Bandarchi, B.; Zhu, C.-Q.; Pintilie, M.; Ramnarine, V.R.; Ibrahimov, E.; Radulovich, N.; Leung, L.; Barczyk, M.; et al. Prognostic Gene-Expression Signature of Carcinoma-Associated Fibroblasts in Non-Small Cell Lung Cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 7160–7165. [Google Scholar] [CrossRef] [Green Version]
- Navab, R.; Strumpf, D.; To, C.; Pasko, E.; Kim, K.S.; Park, C.J.; Hai, J.; Liu, J.; Jonkman, J.; Barczyk, M.; et al. Integrin A11β1 Regulates Cancer Stromal Stiffness and Promotes Tumorigenicity and Metastasis in Non-Small Cell Lung Cancer. Oncogene 2016, 35, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Puig, M.; Lugo, R.; Gabasa, M.; Giménez, A.; Velásquez, A.; Galgoczy, R.; Ramírez, J.; Gómez-Caro, A.; Busnadiego, Ó.; Rodríguez-Pascual, F.; et al. Matrix Stiffening and Β1 Integrin Drive Subtype-Specific Fibroblast Accumulation in Lung Cancer. Mol. Cancer Res. 2015, 13, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Min, A.; Zhu, C.; Wang, J.; Peng, S.; Shuai, C.; Gao, S.; Tang, Z.; Su, T. Focal Adhesion Kinase Knockdown in Carcinoma-Associated Fibroblasts Inhibits Oral Squamous Cell Carcinoma Metastasis via Downregulating MCP-1/CCL2 Expression. J. Biochem. Mol. Toxicol. 2015, 29, 70–76. [Google Scholar] [CrossRef]
- Wu, J.; Yu, C.; Cai, L.; Lu, Y.; Jiang, L.; Liu, C.; Li, Y.; Feng, F.; Gao, Z.; Zhu, Z.; et al. Effects of Increased Kindlin-2 Expression in Bladder Cancer Stromal Fibroblasts. Oncotarget 2017, 8, 50692–50703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagiannis, G.S.; Poutahidis, T.; Erdman, S.E.; Kirsch, R.; Riddell, R.H.; Diamandis, E.P. Cancer-Associated Fibroblasts Drive the Progression of Metastasis through Both Paracrine and Mechanical Pressure on Cancer Tissue. Mol. Cancer Res. 2012, 10, 1403–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdogan, B.; Ao, M.; White, L.M.; Means, A.L.; Brewer, B.M.; Yang, L.; Washington, M.K.; Shi, C.; Franco, O.E.; Weaver, A.M.; et al. Cancer-Associated Fibroblasts Promote Directional Cancer Cell Migration by Aligning Fibronectin. J. Cell Biol. 2017, 216, 3799–3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, S.V.; Grither, W.R.; Brenot, A.; Hwang, P.Y.; Barcus, C.E.; Ernst, M.; Pence, P.; Walter, C.; Pathak, A.; Longmore, G.D. DDR2 Controls Breast Tumor Stiffness and Metastasis by Regulating Integrin Mediated Mechanotransduction in CAFs. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Tchou, J.; Kossenkov, A.V.; Chang, L.; Satija, C.; Herlyn, M.; Showe, L.C.; Puré, E. Human Breast Cancer Associated Fibroblasts Exhibit Subtype Specific Gene Expression Profiles. BMC Med. Genom. 2012, 5, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewell-Loftin, M.K.; Bayer, S.V.H.; Crist, E.; Hughes, T.; Joison, S.M.; Longmore, G.D.; George, S.C. Cancer-Associated Fibroblasts Support Vascular Growth through Mechanical Force. Sci. Rep. 2017, 7, 12574. [Google Scholar] [CrossRef]
- Goicoechea, S.M.; Bednarski, B.; Stack, C.; Cowan, D.W.; Volmar, K.; Thorne, L.; Cukierman, E.; Rustgi, A.K.; Brentnall, T.; Hwang, R.F.; et al. Isoform-Specific Upregulation of Palladin in Human and Murine Pancreas Tumors. PLoS ONE 2010, 5, e10347. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Bassi, D.E.; Simons, J.D.; Devarajan, K.; Al-Saleem, T.; Uzzo, R.G.; Cukierman, E. Elevated Expression of Stromal Palladin Predicts Poor Clinical Outcome in Renal Cell Carcinoma. PLoS ONE 2011, 6, e21494. [Google Scholar] [CrossRef]
- Salaria, S.N.; Illei, P.; Sharma, R.; Walter, K.M.; Klein, A.P.; Eshleman, J.R.; Maitra, A.; Schulick, R.; Winter, J.; Ouellette, M.M.; et al. Palladin Is Overexpressed in the Non-Neoplastic Stroma of Infiltrating Ductal Adenocarcinomas of the Pancreas, but Is Only Rarely Overexpressed in Neoplastic Cells. Cancer Biol. Ther. 2007, 6, 324–328. [Google Scholar] [CrossRef] [Green Version]
- Sato, D.; Tsuchikawa, T.; Mitsuhashi, T.; Hatanaka, Y.; Marukawa, K.; Morooka, A.; Nakamura, T.; Shichinohe, T.; Matsuno, Y.; Hirano, S. Stromal Palladin Expression Is an Independent Prognostic Factor in Pancreatic Ductal Adenocarcinoma. PLoS ONE 2016, 11, e0152523. [Google Scholar] [CrossRef] [Green Version]
- Azatov, M.; Goicoechea, S.M.; Otey, C.A.; Upadhyaya, A. The Actin Crosslinking Protein Palladin Modulates Force Generation and Mechanosensitivity of Tumor Associated Fibroblasts. Sci. Rep. 2016, 6, 28805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLane, J.S.; Ligon, L.A. Palladin Mediates Stiffness-Induced Fibroblast Activation in the Tumor Microenvironment. Biophys. J. 2015, 109, 249–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brentnall, T.A.; Lai, L.A.; Coleman, J.; Bronner, M.P.; Pan, S.; Chen, R. Arousal of Cancer-Associated Stroma: Overexpression of Palladin Activates Fibroblasts to Promote Tumor Invasion. PLoS ONE 2012, 7, e30219. [Google Scholar] [CrossRef] [Green Version]
- Barnes, T.A.; Amir, E. HYPE or HOPE: The Prognostic Value of Infiltrating Immune Cells in Cancer. Br. J. Cancer 2017, 117, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielbassa, K.; Vegna, S.; Ramirez, C.; Akkari, L. Understanding the Origin and Diversity of Macrophages to Tailor Their Targeting in Solid Cancers. Front. Immunol. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Qian, B.-Z.; Pollard, J.W. Macrophage Diversity Enhances Tumor Progression and Metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; McKay, D.; Pollard, J.W.; Lewis, C.E. Diverse Functions of Macrophages in Different Tumor Microenvironments. Cancer Res. 2018, 78, 5492–5503. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.M.; Ridley, A.J. Lipopolysaccharide Induces Actin Reorganization and Tyrosine Phosphorylation of Pyk2 and Paxillin in Monocytes and Macrophages. J. Immunol. 2000, 164, 2028–2036. [Google Scholar] [CrossRef]
- Xi, C.-X.; Xiong, F.; Zhou, Z.; Mei, L.; Xiong, W.-C. PYK2 Interacts with MyD88 and Regulates MyD88-Mediated NF-ΚB Activation in Macrophages. J. Leukoc. Biol. 2010, 87, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Kume, A.; Nishiura, H.; Suda, J.; Suda, T. Focal Adhesion Kinase Upregulated by Granulocyte-Macrophage Colony-Stimulating Factor but Not by Interleukin-3 in Differentiating Myeloid Cells. Blood 1997, 89, 3434–3442. [Google Scholar] [CrossRef]
- He, X.; Chen, X.; Li, B.; Ji, J.; Chen, S. FAK Inhibitors Induce Cell Multinucleation and Dramatically Increase Pro-Tumoral Cytokine Expression in RAW 264.7 Macrophages. FEBS Lett. 2017, 591, 3861–3871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisel, M.B.; Druet, V.A.; Sibilia, J.; Klein, J.-P.; Quesniaux, V.; Wachsmann, D. Cross Talk between MyD88 and Focal Adhesion Kinase Pathways. J. Immunol. 2005, 174, 7393–7397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duong, L.T.; Rodan, G.A. PYK2 Is an Adhesion Kinase in Macrophages, Localized in Podosomes and Activated by Beta(2)-Integrin Ligation. Cell Motil. Cytoskel. 2000, 47, 174–188. [Google Scholar] [CrossRef]
- Kedzierska, K.; Vardaxis, N.J.; Jaworowski, A.; Crowe, S.M. FcγR-Mediated Phagocytosis by Human Macrophages Involves Hck, Syk, and Pyk2 and Is Augmented by GM-CSF. J. Leukoc. Biol. 2001, 70, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.A.; Thomas, K.S.; Bouton, A.H. The Differential Expression of Yersinia Pseudotuberculosis Adhesins Determines the Requirement for FAK and/or Pyk2 during Bacterial Phagocytosis by Macrophages. Cell. Microbiol. 2007, 9, 596–609. [Google Scholar] [CrossRef]
- Allen, L.A.; Aderem, A. Molecular Definition of Distinct Cytoskeletal Structures Involved in Complement- and Fc Receptor-Mediated Phagocytosis in Macrophages. J. Exp. Med. 1996, 184, 627–637. [Google Scholar] [CrossRef]
- Fuortes, M.; Jin, W.W.; Nathan, C. Beta 2 Integrin-Dependent Tyrosine Phosphorylation of Paxillin in Human Neutrophils Treated with Tumor Necrosis Factor. J. Cell Biol. 1994, 127, 1477–1483. [Google Scholar] [CrossRef]
- Greenberg, S.; Chang, P.; Silverstein, S.C. Tyrosine Phosphorylation of the Gamma Subunit of Fc Gamma Receptors, P72syk, and Paxillin during Fc Receptor-Mediated Phagocytosis in Macrophages. J. Biol. Chem. 1994, 269, 3897–3902. [Google Scholar] [CrossRef]
- Abshire, M.Y.; Thomas, K.S.; Owen, K.A.; Bouton, A.H. Macrophage Motility Requires Distinct A5β1/FAK and A4β1/Paxillin Signaling Events. J. Leukoc. Biol. 2011, 89, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Rhee, I.; Zhong, M.-C.; Reizis, B.; Cheong, C.; Veillette, A. Control of Dendritic Cell Migration, T Cell-Dependent Immunity, and Autoimmunity by Protein Tyrosine Phosphatase PTPN12 Expressed in Dendritic Cells. Mol. Cell Biol. 2014, 34, 888–899. [Google Scholar] [CrossRef] [Green Version]
- Riol-Blanco, L.; Sánchez-Sánchez, N.; Torres, A.; Tejedor, A.; Narumiya, S.; Corbí, A.L.; Sánchez-Mateos, P.; Rodríguez-Fernández, J.L. The Chemokine Receptor CCR7 Activates in Dendritic Cells Two Signaling Modules That Independently Regulate Chemotaxis and Migratory Speed. J. Immunol. 2005, 174, 4070–4080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.; Fang, B.; Zhang, Y.; Wang, C.; Zhou, J.; Niu, C.; Gao, Y.; Zhao, D.; He, J.; Wang, J.; et al. Mechanical Stretch Promotes Tumoricidal M1 Polarization via the FAK/NF-ΚB Signaling Pathway. FASEB J. 2019, 33, 13254–13266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiossone, L.; Dumas, P.-Y.; Vienne, M.; Vivier, E. Natural Killer Cells and Other Innate Lymphoid Cells in Cancer. Nat. Rev. Immunol. 2018, 18, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Gismondi, A.; Jacobelli, J.; Mainiero, F.; Paolini, R.; Piccoli, M.; Frati, L.; Santoni, A. Cutting Edge: Functional Role for Proline-Rich Tyrosine Kinase 2 in NK Cell-Mediated Natural Cytotoxicity. J. Immunol. 2000, 164, 2272–2276. [Google Scholar] [CrossRef] [PubMed]
- March, M.E.; Long, E.O. Β2 Integrin Induces TCR ζ–Syk–PLC-γ Phosphorylation and Paxillin-Dependent Granule Polarization in Human NK Cells. J. Immunol. 2011, 186. [Google Scholar] [CrossRef] [Green Version]
- Sancho, D.; Nieto, M.; Llano, M.; Rodríguez-Fernández, J.L.; Tejedor, R.; Avraham, S.; Cabañas, C.; López-Botet, M.; Sánchez-Madrid, F. The Tyrosine Kinase PYK-2/RAFTK Regulates Natural Killer (NK) Cell Cytotoxic Response, and Is Translocated and Activated upon Specific Target Cell Recognition and Killing. J. Cell Biol. 2000, 149, 1249–1262. [Google Scholar] [CrossRef]
- Steblyanko, M.; Anikeeva, N.; Campbell, K.S.; Keen, J.H.; Sykulev, Y. Integrins Influence the Size and Dynamics of Signaling Microclusters in a Pyk2-Dependent Manner. J. Biol. Chem. 2015, 290, 11833–11842. [Google Scholar] [CrossRef] [Green Version]
- Coca, S.; Perez-Piqueras, J.; Martinez, D.; Colmenarejo, A.; Saez, M.A.; Vallejo, C.; Martos, J.A.; Moreno, M. The Prognostic Significance of Intratumoral Natural Killer Cells in Patients with Colorectal Carcinoma. Cancer 1997, 79, 2320–2328. [Google Scholar] [CrossRef]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic Value of Intratumoral Natural Killer Cells in Gastric Carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Villegas, F.R.; Coca, S.; Villarrubia, V.G.; Jiménez, R.; Chillón, M.J.; Jareño, J.; Zuil, M.; Callol, L. Prognostic Significance of Tumor Infiltrating Natural Killer Cells Subset CD57 in Patients with Squamous Cell Lung Cancer. Lung Cancer 2002, 35, 23–28. [Google Scholar] [CrossRef]
- Bassani, B.; Baci, D.; Gallazzi, M.; Poggi, A.; Bruno, A.; Mortara, L. Natural Killer Cells as Key Players of Tumor Progression and Angiogenesis: Old and Novel Tools to Divert Their Pro-Tumor Activities into Potent Anti-Tumor Effects. Cancers Basel 2019, 11, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, S.; Burridge, K.; Silverstein, S.C. Colocalization of F-Actin and Talin during Fc Receptor-Mediated Phagocytosis in Mouse Macrophages. J. Exp. Med. 1990, 172, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Jaumouillé, V.; Cartagena-Rivera, A.X.; Waterman, C.M. Coupling of β 2 Integrins to Actin by a Mechanosensitive Molecular Clutch Drives Complement Receptor-Mediated Phagocytosis. Nat. Cell Biol. 2019, 21, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Wiedemann, A.; Tzircotis, G.; Monkley, S.J.; Critchley, D.R.; Caron, E. An Essential Role for Talin during AMβ2-Mediated Phagocytosis. Mol. Biol. Cell 2007, 18, 976–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaidano, G.; Bergui, L.; Schena, M.; Gaboli, M.; Cremona, O.; Marchisio, P.C.; Caligaris-Cappio, F. Integrin Distribution and Cytoskeleton Organization in Normal and Malignant Monocytes. Leukemia 1990, 4, 682–687. [Google Scholar] [PubMed]
- Joosten, B.; Willemse, M.; Fransen, J.; Cambi, A.; van den Dries, K. Super-Resolution Correlative Light and Electron Microscopy (SR-CLEM) Reveals Novel Ultrastructural Insights Into Dendritic Cell Podosomes. Front. Immunol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Linder, S.; Kopp, P. Podosomes at a Glance. J. Cell Sci. 2005, 118, 2079–2082. [Google Scholar] [CrossRef] [Green Version]
- Baranov, M.; Ter Beest, M.; Reinieren-Beeren, I.; Cambi, A.; Figdor, C.G.; van den Bogaart, G. Podosomes of Dendritic Cells Facilitate Antigen Sampling. J. Cell. Sci. 2014, 127, 1052–1064. [Google Scholar] [CrossRef] [Green Version]
- Lämmermann, T.; Bader, B.L.; Monkley, S.J.; Worbs, T.; Wedlich-Söldner, R.; Hirsch, K.; Keller, M.; Förster, R.; Critchley, D.R.; Fässler, R.; et al. Rapid Leukocyte Migration by Integrin-Independent Flowing and Squeezing. Nature 2008, 453, 51–55. [Google Scholar] [CrossRef]
- Gunawan, M.; Venkatesan, N.; Loh, J.T.; Wong, J.F.; Berger, H.; Neo, W.H.; Li, L.Y.J.; La Win, M.K.; Yau, Y.H.; Guo, T.; et al. The Methyltransferase Ezh2 Controls Cell Adhesion and Migration through Direct Methylation of the Extranuclear Regulatory Protein Talin. Nat. Immunol. 2015, 16, 505–516. [Google Scholar] [CrossRef]
- Lim, T.J.F.; Su, I.-H. Talin1 Methylation Is Required for Neutrophil Infiltration and Lipopolysaccharide-Induced Lethality. J. Immunol. 2018, 201, 3651–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Wang, Z.; Zhou, J.; Huang, J.; Zhou, L.; Luo, J.; Wan, Y.Y.; Long, H.; Zhu, B. EZH2 Inhibitor GSK126 Suppresses Antitumor Immunity by Driving Production of Myeloid-Derived Suppressor Cells. Cancer Res. 2019, 79, 2009–2020. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, E.B.; Sinha, S.; Li, L.; Dedhar, S.; Force, T.; Rosenzweig, A.; Gerszten, R.E. Role of Integrin-Linked Kinase in Leukocyte Recruitment. J. Biol. Chem. 2002, 277, 16371–16375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troussard, A.A.; Mawji, N.M.; Ong, C.; Mui, A.; St -Arnaud, R.; Dedhar, S. Conditional Knock-out of Integrin-Linked Kinase Demonstrates an Essential Role in Protein Kinase B/Akt Activation. J. Biol. Chem. 2003, 278, 22374–22378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimi, R.; Yamaji, S.; Suzuki, A.; Mishima, W.; Okamura, M.; Obana, T.; Matsuda, C.; Miwa, Y.; Ohno, S.; Ishigatsubo, Y. The γ-Parvin-Integrin-Linked Kinase Complex Is Critically Involved in Leukocyte-Substrate Interaction. J. Immunol. 2006, 176, 3611–3624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, L.; Howarth, K.; McDowall, A.; Patzak, I.; Evans, R.; Ussar, S.; Moser, M.; Metin, A.; Fried, M.; Tomlinson, I.; et al. Leukocyte Adhesion Deficiency-III Is Caused by Mutations in KINDLIN3 Affecting Integrin Activation. Nat. Med. 2009, 15, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, W.E.; Zicha, D.; Ridley, A.J.; Jones, G.E. A Role for Cdc42 in Macrophage Chemotaxis. J. Cell Biol. 1998, 141, 1147–1157. [Google Scholar] [CrossRef] [Green Version]
- Burns, S.; Thrasher, A.J.; Blundell, M.P.; Machesky, L.; Jones, G.E. Configuration of Human Dendritic Cell Cytoskeleton by Rho GTPases, the WAS Protein, and Differentiation. Blood 2001, 98, 1142–1149. [Google Scholar] [CrossRef]
- Zhao, T.; Bokoch, G.M. Critical Role of Proline-Rich Tyrosine Kinase 2 in Reversion of the Adhesion-Mediated Suppression of Reactive Oxygen Species Generation by Human Neutrophils. J. Immunol. 2005, 174, 8049–8055. [Google Scholar] [CrossRef] [Green Version]
- Coppolino, M.G.; Krause, M.; Hagendorff, P.; Monner, D.A.; Trimble, W.; Grinstein, S.; Wehland, J.; Sechi, A.S. Evidence for a Molecular Complex Consisting of Fyb/SLAP, SLP-76, Nck, VASP and WASP That Links the Actin Cytoskeleton to Fcgamma Receptor Signalling during Phagocytosis. J. Cell. Sci. 2001, 114, 4307–4318. [Google Scholar]
- Wilton, K.M.; Billadeau, D.D. VASP Regulates NK Cell Lytic Granule Convergence. J. Immunol. 2018, 201, 2899–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, S.G.; Rabinovich, P.M. T Cell Reprogramming Against Cancer. Methods Mol. Biol. 2020, 2097, 3–44. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lostao, L.; Anel, A.; Pardo, J. How Do Cytotoxic Lymphocytes Kill Cancer Cells? Clin. Cancer Res. 2015, 21, 5047–5056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-J.; Cantor, H. CD4 T-Cell Subsets and Tumor Immunity: The Helpful and the Not-so-Helpful. Cancer Immunol. Res. 2014, 2, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farhood, B.; Najafi, M.; Mortezaee, K. CD8+ Cytotoxic T Lymphocytes in Cancer Immunotherapy: A Review. J. Cell. Physiol. 2019, 234, 8509–8521. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Zhang, Y.; Xu, J.; Yin, T.; Lu, X.-J. T Cell Dysfunction in Cancer Immunity and Immunotherapy. Front. Immunol. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Liu, S.; Zhang, B.; Qiao, L.; Zhang, Y.; Zhang, Y. T Cell Dysfunction and Exhaustion in Cancer. Front. Cell Dev. Biol. 2020, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yuen, G.J.; Demissie, E.; Pillai, S. B Lymphocytes and Cancer: A Love-Hate Relationship. Trends Cancer 2016, 2, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Tsou, P.; Katayama, H.; Ostrin, E.J.; Hanash, S.M. The Emerging Role of B Cells in Tumor Immunity. Cancer Res. 2016, 76, 5597–5601. [Google Scholar] [CrossRef] [Green Version]
- Chapman, N.M.; Houtman, J.C.D. Functions of the FAK Family Kinases in T Cells: Beyond Actin Cytoskeletal Rearrangement. Immunol. Res. 2014, 59, 23–34. [Google Scholar] [CrossRef]
- Cheung, S.M.S.; Ostergaard, H.L. Pyk2 Controls Integrin-Dependent CTL Migration through Regulation of De-Adhesion. J. Immunol. 2016, 197, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.M.; Conolly, S.F.; Reinl, E.L.; Houtman, J.C. Focal Adhesion Kinase Negatively Regulates Lck Function Downstream of the T Cell Antigen Receptor. J. Immunol. 2013, 191, 6208–6221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, N.M.; Yoder, A.N.; Barbón, K.M.; Bilal, M.Y.; Connolly, S.F.; Houtman, J.C.D. Proline-Rich Tyrosine Kinase 2 Controls PI3-Kinase Activation Downstream of the T Cell Antigen Receptor in Human T Cells. J. Leukoc. Biol. 2015, 97, 285–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beinke, S.; Phee, H.; Clingan, J.M.; Schlessinger, J.; Matloubian, M.; Weiss, A. Proline-Rich Tyrosine Kinase-2 Is Critical for CD8 T-Cell Short-Lived Effector Fate. PNAS 2010, 107, 16234–16239. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto-Tane, A.; Sakuma, M.; Ike, H.; Yokosuka, T.; Kimura, Y.; Ohara, O.; Saito, T. Micro–Adhesion Rings Surrounding TCR Microclusters Are Essential for T Cell Activation. J. Exp. Med. 2016, 213, 1609–1625. [Google Scholar] [CrossRef]
- Van Seventer, G.A.; Salmen, H.J.; Law, S.F.; O’Neill, G.M.; Mullen, M.M.; Franz, A.M.; Kanner, S.B.; Golemis, E.A.; van Seventer, J.M. Focal Adhesion Kinase Regulates Beta1 Integrin-Dependent T Cell Migration through an HEF1 Effector Pathway. Eur. J. Immunol. 2001, 31, 1417–1427. [Google Scholar] [CrossRef]
- Astier, A.; Avraham, H.; Manie, S.N.; Groopman, J.; Canty, T.; Avraham, S.; Freedman, A.S. The Related Adhesion Focal Tyrosine Kinase Is Tyrosine-Phosphorylated after Beta1-Integrin Stimulation in B Cells and Binds to P130cas. J. Biol. Chem. 1997, 272, 228–232. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.; Zhu, B.-M.; Harley, B.; Park, S.-Y.; Kobayashi, T.; Manis, J.P.; Luo, H.R.; Yoshimura, A.; Hennighausen, L.; Silberstein, L.E. SOCS3 Protein Developmentally Regulates the Chemokine Receptor CXCR4-FAK Signaling Pathway during B Lymphopoiesis. Immunity 2007, 27, 811–823. [Google Scholar] [CrossRef] [Green Version]
- Tse, K.W.K.; Dang-Lawson, M.; Lee, R.L.; Vong, D.; Bulic, A.; Buckbinder, L.; Gold, M.R. B Cell Receptor-Induced Phosphorylation of Pyk2 and Focal Adhesion Kinase Involves Integrins and the Rap GTPases and Is Required for B Cell Spreading. J. Biol. Chem. 2009, 284, 22865–22877. [Google Scholar] [CrossRef] [Green Version]
- Serrels, B.; McGivern, N.; Canel, M.; Byron, A.; Johnson, S.C.; McSorley, H.J.; Quinn, N.; Taggart, D.; Von Kreigsheim, A.; Anderton, S.M.; et al. IL-33 and ST2 Mediate FAK-Dependent Antitumor Immune Evasion through Transcriptional Networks. Sci. Signal. 2017, 10, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Symeonides, S.N.; Anderton, S.M.; Serrels, A. FAK-Inhibition Opens the Door to Checkpoint Immunotherapy in Pancreatic Cancer. J. Immunother. Cancer 2017, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, L.; Corgnac, S.; Boutet, M.; Gros, G.; Validire, P.; Bismuth, G.; Mami-Chouaib, F. Paxillin Binding to the Cytoplasmic Domain of CD103 Promotes Cell Adhesion and Effector Functions for CD8+ Resident Memory T Cells in Tumors. Cancer Res. 2017, 77, 7072–7082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ando, M.; Ito, M.; Srirat, T.; Kondo, T.; Yoshimura, A. Memory T Cell, Exhaustion, and Tumor Immunity. Immunol. Med. 2020, 43, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corgnac, S.; Boutet, M.; Kfoury, M.; Naltet, C.; Mami-Chouaib, F. The Emerging Role of CD8+ Tissue Resident Memory T (TRM) Cells in Antitumor Immunity: A Unique Functional Contribution of the CD103 Integrin. Front. Immunol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, M.; Arrigoni, A.; Rossetti, G.; Gruarin, P.; Ranzani, V.; Politano, C.; Bonnal, R.J.P.; Provasi, E.; Sarnicola, M.L.; Panzeri, I.; et al. Transcriptional Landscape of Human Tissue Lymphocytes Unveils Uniqueness of Tumor-Infiltrating T Regulatory Cells. Immunity 2016, 45, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Guinoa, J.S.; Barrio, L.; Carrasco, Y.R. Vinculin Arrests Motile B Cells by Stabilizing Integrin Clustering at the Immune Synapse. J. Immunol. 2013, 191, 2742–2751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: New Insights into Their Functions from in Vivo Studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Saoudi, A.; Kassem, S.; Dejean, A.; Gaud, G. Rho-GTPases as Key Regulators of T Lymphocyte Biology. Small GTPases 2014, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tybulewicz, V.L.J.; Henderson, R.B. Rho Family GTPases and Their Regulators in Lymphocytes. Nat. Rev. Immunol. 2009, 9, 630–644. [Google Scholar] [CrossRef]
- Guo, F.; Velu, C.S.; Grimes, H.L.; Zheng, Y. Rho GTPase Cdc42 Is Essential for B-Lymphocyte Development and Activation. Blood 2009, 114, 2909–2916. [Google Scholar] [CrossRef] [Green Version]
- Faure, S.; Salazar-Fontana, L.I.; Semichon, M.; Tybulewicz, V.L.J.; Bismuth, G.; Trautmann, A.; Germain, R.N.; Delon, J. ERM Proteins Regulate Cytoskeleton Relaxation Promoting T Cell-APC Conjugation. Nat. Immunol. 2004, 5, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Roumier, A.; Olivo-Marin, J.C.; Arpin, M.; Michel, F.; Martin, M.; Mangeat, P.; Acuto, O.; Dautry-Varsat, A.; Alcover, A. The Membrane-Microfilament Linker Ezrin Is Involved in the Formation of the Immunological Synapse and in T Cell Activation. Immunity 2001, 15, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Derry, J.M.; Ochs, H.D.; Francke, U. Isolation of a Novel Gene Mutated in Wiskott-Aldrich Syndrome. Cell 1994, 78, 635–644. [Google Scholar] [CrossRef]
- Krause, M.; Sechi, A.S.; Konradt, M.; Monner, D.; Gertler, F.B.; Wehland, J. Fyn-Binding Protein (Fyb)/SLP-76-Associated Protein (SLAP), Ena/Vasodilator-Stimulated Phosphoprotein (VASP) Proteins and the Arp2/3 Complex Link T Cell Receptor (TCR) Signaling to the Actin Cytoskeleton. J. Cell Biol. 2000, 149, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Molecule | Cell Type | Murine Tumour Model | Intervention | Effect | References |
|---|---|---|---|---|---|
| Adhesome signalling | |||||
| FAK/Pyk2 | ECs | Subcutaneous tumours (B16F0 or CMT19T) | Pdgfb iCreERT2; FAKfl/fl | anti-tumourigenic: vessel density (↓), no Pyk2 compensation | [13] |
| Orthotopic patient-derived human glioma (DBTRG) | Tie2 CreERT2; FAKfl/fl | anti-tumourigenic: vascular normalisation, no Pyk2 compensation | [14] | ||
| Subcutaneous tumours (B16F0 or CMT19T) | FAKhet | pro-tumourigenic: vessel density (↑) | [15] | ||
| Subcutaneous tumours (B16F10), Metastasis models: I.V or spontaneous metastasis after B16F10 injection | SCL CreERT; FAKfl/KD | metastasis (↓) tumour growth (−) | [16] | ||
| Subcutaneous tumours (B16F0) | PDGFb iCreERT; FAKfl/fl; R26FAKKD/KD (FAK kinase-dead mice) | anti-tumourigenic: vessel density (↓), vascular permeability (↓), VE−CAD pY658 levels (↓) | [17] | ||
| Subcutaneous tumours (B16F0) | PDGFb iCreERT; FAKfl/fl; R26FAKDM/DM (FAK DM mice: KD with a putatively phosphomimetic Y397E mutation) | anti-tumourigenic: vessel density (↓), vascular permeability (−), VE−CAD pY658 levels (−) | [17] | ||
| Subcutaneous tumours (B16F0 or CMT19T) | PDGFb iCreERT; FAKfl/fl; R26FAKY397F/Y397F | anti-tumourigenic: vessel density (↓) | [18] | ||
| Subcutaneous tumours (B16F0 or CMT19T) | PDGFb iCreERT; FAKfl/fl; R26FAKY861F/Y8617F | no effect: tumour growth (−), vessel density (−) | [18] | ||
| Pericytes | Subcutaneous tumours (B16F0 or LLC) | PDGFRβ Cre; FAKfl/fl | pro-tumourigenic: vessel density (↑) | [19] | |
| Subcutaneous tumours (LLC) | PDGFRβ Cre; FAKY397F/Y397F | no effect: tumour growth (−), vessel density (−) | [20] | ||
| Subcutaneous tumours (B16F0 or LLC) | PDGFRβ Cre; FAKY861F/Y861F | anti-tumourigenic: vessel density (↓) | [20] | ||
| CAFs | Orthotopic xenografts of pancreatic cancer (MPanc-96 or MAD08-608) | Inhibition (FAK/Pyk2 inhibitor: PF-562,271) | anti-tumourigenic: CAFs migration and infiltration (↓) | [21] | |
| Genetically engineered mice of pancreatic cancer (KPC and KPPC) | Inhibition (FAK inhibitor: VS-4718) | tumour stasis: intratumoural aSMA+ and FAP+ cells (↓), FAP+ cell proliferation (↓) | [22] | ||
| Subcutaneous tumours (HT29) | Inhibition (FAK inhibitor: PF-573,228) or HT29 expressing FAKY397F construct | anti-tumourigenic | [23] | ||
| Syngeneic orthotopic breast (E0771) and pancreatic (TB32048) tumours/MMTV-PyMT mouse model of breast cancer | FSP Cre+; FAKfl/fl mice/MMTV+; FSP Cre+; FAKfl/fl | pro-tumourigenic: CAF−mediated changes in malignant metabolism (↑) | [24] | ||
| Pancreatic cancer cell and fibroblast syngeneic and orthotopic cografting | FAKKD(kinase−dead) fibroblasts, inhibition (FAK inhibitor: PF562,271) | anti-tumourigenic: tumour growth (−), lung metastasis (↓), fibrosis (↓), M2 macrophage polarisation and migration (↓), | [25] | ||
| MMTV-PyMT mouse model of spontaneous breast cancer | MMTV-PyMT; Col1a2 CreER; FAKfl/fl | tumour development and growth (−), breast cancer metastasis (↓), exosome amount and functions (↓) | [26] | ||
| TAMs/monocytes | Orthotopic pancreatic tumours (MPanc-96 or MAD08-608) | Inhibition (FAK/PYK2 inhibitor: PF-562,271) | anti-tumourigenic: TAMs (F4/80+) migration and infiltration (↓) | [21] | |
| Genetically engineered mice of pancreatic cancer(KPC and KPPC) | Inhibition (FAK inhibitor: VS-4718) | tumour stasis: intratumoural F4/80+ and CD206+ macrophages (↓) | [22] | ||
| Syngeneic orthotopic breast tumours (4T1 or MDA-MB-231) | Inhibition (FAK inhibitor: PND-1186) | anti-tumourigenic: metastasis (↓), leukocyte infiltration (↓) | [27] | ||
| Orthotopic breast tumours (4T1) | Inhibition (FAK inhibitors: PF-562271 and PF-573,228) | anti-tumourigenic: intratumoural F4/80+ macrophages (↓) | [28] | ||
| MMTV-PyMT mouse model of spontaneous breast cancer | PyVmT+/−;LysMwt/cre; FAKΔmyeloid | pro−and anti-tumorigenic: depending on the stage of malignancy, carcinoma stage: NK cells (↓) | [29] | ||
| Subcutaneous tumours (SCC) | Inhibition (FAK inhibitor: BI 853,520) | anti−tumorigenic: PD−L2 surface expression on TAMs (↓) | [30] | ||
| MDSCs/TANs | Genetically engineered mice of pancreatic cancer (KPC and KPPC) | Inhibition (FAK inhibitor: VS-4718) | tumour stasis: recruitment of Gr1+ granulocytes (↓) | [22] | |
| Subcutaneous tumours (SCC cells) | Inhibition (FAK inhibitor: BI 853,520) | anti-tumorigenic: PD−L2 surface expression on M−MDSCs (↓) | [30] | ||
| CD3+/CD4+/CD8+ T cells | Subcutaneous tumours (SCC cells) | SSC FAK−/− cells, inhibition (FAK inhibitor: VS-4718) | anti-tumourigenic: CD4+ and CD8+ cell recruitment (↑) | [31] | |
| Genetically engineered mice of pancreatic cancer (KPC and KPPC) | Inhibition (FAK inhibitor: VS-4718) | tumour stasis: CD4+ and CD8+ cell recruitment (↑) | [22] | ||
| Subcutaneous tumours (SCC cells) | Inhibition (FAK inhibitor: BI 853,520) | anti-tumorigenic: ICOS surface expression in CD8+ cells (↑) | [30] | ||
| Tregs | Genetically engineered mice of pancreatic cancer (KPC and KPPC) | SSC FAK−/− cells, inhibition (FAK inhibitor: VS-4718) | anti-tumourigenic: infiltration of CD4+FoxP3+CD25+ Tregs in the tumour niche (↓) | [31] | |
| Genetically engineered mice of pancreatic cancer (KPC and KPPC) | Inhibition (FAK inhibitor: VS-4718) | anti-tumorigenic: infiltration of CD4+FOXP3+ TREGS in the tumour niche (↓) | [22] | ||
| Subcutaneous tumours (SCC) | Inhibition (FAK inhibitor: BI 853,520) | anti-tumorigenic: Tregs (↓) | [30] | ||
| Paxillin | ECs | Matrigel plugs implanted subcutaneously (LLC) | siRNA | matrigel plugs: angiogenesis (↑) | [32] |
| GIT/PIX/PAK | CAFs | Genetically engineered mice of pancreatic cancer (KPC) | PAK1−/− | anti-tumourigenic: intratumoural aSMA+ and desmin+ cells (↓) | [33] |
| CD3+/CD4+/CD8+ T cells | Genetically engineered mice of pancreatic cancer (KPC) | PAK1−/− | anti-tumourigenic: CD3+, CD4+ and CD8+ cell recruitment (↑) | [33] | |
| Genetically engineered mice of intestinal-colorectal cancer APCΔ14/+ | PAK1−/−, inhibition (PAK inhibitor PF-3758,309) | anti-tumourigenic: CD3+, CD4+ and CD8+ cell recruitment (↑) | [34] | ||
| Tregs | Subcutaneous tumours (B16F10 or TRAMP-C1) | GIT2−/− | anti-tumourigenic | [35] | |
| Adhesion organisation | |||||
| Kindlin2 | ECs | Subcutaneous tumours (RM1) | Kindlin2+/− | anti-tumourigenic: vessel density (↓) | [36] |
| CAFs | Subcutaneous coengraftments (SUIT-2 and PSCs) | siRNA | anti-tumourigenic | [37] | |
| Kindlin3 | TAMs/monocytes | Intravenous injections of B16F10 cells | Mx1 Cre+/− (Poly(I:C)); CX3CR1gfp/+; kindlin3fl/fl | pro-tumorigenic: monocyte patrolling (↓), NK cells (↓), | [38] |
| Talin | ECs | Subcutaneous tumours (B16F0) | Cdh5 CreERT2; talin1f/L325R | anti-tumourigenic: vessel density (↓) | [39] |
| ILK | ECs | Thyroid (DRO) tumour xenografts | Inhibition (ILK inhibitor: QLT0267) | anti-tumourigenic: vessel density (↓) | [40] |
| Orthotopic pancreatic tumour xenografts | Inhibition (ILK inhibitor: QLT0254) | anti-tumourigenic: vessel density (−) | [41] | ||
| Glioblastoma (U87MG) tumour xenografts | Inhibition (ILK inhibitor: QLT0267) | anti-tumourigenic: vessel density (↓) | [42] | ||
| Subcutaneous tumours (PC3) | Inhibition (ILK inhibitor: KP-307-2) | anti-tumourigenic: vessel density (↓) | [43] | ||
| Actin regulatory layer | |||||
| Palladin | CAFs | Orthotopic xenografts of pancreatic cancer (AsPC-1 coimplantation with palladin KD CAFs) | siRNA | anti-tumourigenic | [44] |
| VASP | CAFs | Subcutaneous tumours (LLC coimplantation with VASP KD MEFs) | siRNA and EVH2 VASP mutant | anti-tumourigenic: intratumoural aSMA+ and desmin+ cells (↓) | [45] |
| Subcutaneous tumours (HT-29 coimplantation with VASP KD HSCs) | siRNA | anti-tumourigenic: intratumoural aSMA+ cells (↓) | [46] | ||
| Filamin | ECs | Subcutaneous tumours (T241 or B16F0) | VE-Cad Cre; flnAfl/fl | anti-tumourigenic: vessel density (↓) | [47] |
| Rac1 | ECs | Subcutaneous tumours (B16F0) | PDGFb iCreERT; Rac1fl/fl | no effect: tumour growth (−), vessel density (−) | [48] |
| Subcutaneous tumours (Neuro2a cells) | siRNA | anti-tumourigenic: vessel density (↓) | [49] | ||
| RhoJ | ECs | Subcutaneous tumours (LLC) | RhoJKO | anti-tumourigenic: vessel density (↓) | [50] |
| Subcutaneous tumours (B16F0 or LLC) | Cad5 CreERT2; RhoJfl/fl | anti-tumourigenic: vessel density (↓) | [51] | ||
| Molecule | Cell Type | Mouse Model | Effect | References |
|---|---|---|---|---|
| Adhesome signalling | ||||
| Paxillin | ECs | siRNA | Retinal angiogenesis (↑), vessel sprouting (↑) | [32] |
| FAK | ECs | Tie2 Cre; FAKfl/fl | EC migration (↓), EC proliferation (↓), EC retraction and death (↑), haemorrhage, vessel growth (↓), vessel regression (↑), embryonic lethality (E10.5–E11.5) | [57,58] |
| Tie2 Cre; FAKflox/KD | Vascular permeability (↑), VE-cadherin Y658 phosphorylation (↓), embryonic lethality (E13.5) | [59] | ||
| End-SCL CreERT; FAKfl/fl | No effect-Pyk2 compensation (adult mice) | [60] | ||
| Myeloid lineages → macrophages | LysM cre; FAKfl/fl | Macrophages: adhesion (↓), migration and invasion (↓), recruitment (↓) | [61] | |
| Myeloid lineages→ neutrophils | LysM cre; FAKfl/fl | Adhesion to fibronectin or ICAM-1 (↓), life span (↓), pathogen-killing capability (↓) | [62] | |
| T cells | FAK+/− | Normal T cell development | [63] | |
| CD4 Cre; FAKfl/fl | Normal T cell development | [64] | ||
| B cells | CD19 Cre; Fak fl/fl | Progenitor B and immature B cells (↓), homing to the BM (↓), retention in BM (↓) | [65] | |
| Pyk2 | Macrophages | Pyk2–/– | Macrophages: cell polarisation (↓), cell contractility (↓), size of lamellipodia (↑), migration (↓), | [66] |
| Neutrophils | Pyk2–/– | Neutrophil migration (↓), degranulation (↓) | [67] | |
| Monocytes | Pyk2–/– | Number of BM monocyte-lineage cells (↓), Pyk2 promotes the turnover of monocytes at steady state | [68] | |
| T cells | Pyk2–/– | Normal T cell development | [66] | |
| Pyk2–/– | CD8 T cell activation (↓), LFA-1-dependent CD8 T cell adhesion and migration (↓), CD8+ TEFF (↓) | [69] | ||
| B cells | Pyk2–/– | B cell migration into the marginal zone of spleen (↓), humoral responses to T dependent antigen (↓) | [70] | |
| a-PIX | Immune cells→ T and B cells | a-PIX−/− | Mature lymphocytes (↓), T cell proliferation (↓), B cell proliferation (↓), B or T motility (↑), disrupted immune synapse | [71] |
| T cells | a-PIX−/− | Thymocytes motility (↑), T cell positive selection (↓) | [72] | |
| Adhesion organisation | ||||
| Kindlin3 | Neutrophils | kindlin3−/− | Neutrophil: firm adhesion (↓), vascular arrest (↓), recruitment (↓), bleeding, death | [73,74] |
| HSCs→ neutrophils | Mx1 Cre (Poly(I:C)); kindlin3fl/fl | Leukocyte adhesion (↓), bleeding, death, 5% protein expression viable | [75] | |
| T cells | kindlin3−/−, Mx1 Cre (Poly(I:C)); kindlin3fl/fl, CD4 Cre; kindlin3fl/fl | Thumocyte proliferation (↓), T cell homing to thymus (↓),T cell adhesion at low vascular shear stress (↓) | [76] | |
| Talin1 | ECs | Tie2 Cre; talin1fl/fl | Haemorrhaging, vessel density (↓), small and round ECs, embryonic lethality (E10.5) | [77] |
| Cdh5 CreERT2; talin1 | Haemorrhaging of intestine vasculature, death (adult mice) | [78] | ||
| Cdh5 CreERT2; talin1 | Retinal angiogenesis (↓), haemorrhaging, EC proliferation (↓), embryonic lethality | [39] | ||
| Cdh5 CreERT2; talin1fl/L325R | Retinal angiogenesis (↓) | [39] | ||
| Neutrophils, platelets | Rap1 binding deficient talin1 knock-in | Neutrophil adhesion (↓), leukocyte adhesion and extravasation (↓) | [79] | |
| HSCs → neutrophils | Mx1 Cre (Poly(I:C)); talin1fl/fl | Neutrophil slow rolling and vascular arrest (↓), neutrophil recruitment (↓) | [73] | |
| Dendritic cells | CD11c Cre; talin1fl/fl | DC activation (↓), T cell and B cell priming responses (↓) | [80] | |
| CD11c Cre; talin1fl/fl | DC migration and activation (↓) | [81] | ||
| NK | Talin1−/− ES -> differentiate into NK | NK LFA-1-mediated-adhesion (↓), NK cytotoxicity (↓): only retained for selective target cells lacking ICAM-1 | [82] | |
| T cells | CD4 Cre; talin1fl/fl | T cell activation and proliferation (↓), T cell–APC interactions (transient) | [83] | |
| CD4 Cre; talin1fl/fl | T regs (↓), T cell–DCs interactions (↓) | [84] | ||
| B cells | CD19 Cre; talin1−/fl | Normal B cell development and maturation, humoral responses to T-dependent antigens (↓), B cells homing to lymph nodes and BM (bone marrow) (↓) | [85] | |
| Vinculin | HSCs → neutrophils | Mx1 Cre (Poly(I:C)); Vclfl/fl | Normal neutrophil recruitment, neutrophil spreading (↓) | [86] |
| ILK | ECs | Tie2 Cre; ILKfl/fl | Labyrinthine vascularisation (↓), EC apoptosis (↑), embryonic lethality (E9.5–E10) | [87] |
| PDGFb iCreERT; ILKfl/fl | Vessel sprouting (↓), EC proliferation (↓), disruption of the blood–retina barrier | [88] | ||
| vSMCs | Pdgfrb Cre; ILK fl/fl | vSMCs: contraction (↑), migration (↓), focal adhesion assembly (↓) | [89] | |
| T cells | Lck Cre+/ILKfl/fl | T cell chemotaxis (↓), T cell survival (↓) | [90] | |
| Parvin | ECs | Tie2 Cre; α-pvfl/fl | Haemorrhaging, vessel density (↓), embryonic lethality (E13.5) | [91] |
| Cdh5 CreERT2; α-pvfl/fl | Vessel density (↓), vessel sprouting (↓), EC proliferation (↓), vessel regression (↑) | [91] | ||
| Tie2 Cre; α-pvfl/fl; β-pv−/− | Haemorrhaging, vessel branching (↓), vessel diameter (↑), vessel density (↓), embryonic lethality (E10.5–E12.5) | [92] | ||
| vSMCs | a-pv−/− | vSMCs: contraction (↑), directed migration (↓) | [93] | |
| Hematopoietic cells | γ-pv−/− | Normal haematopoiesis | [94] | |
| FHL2 | Dendritic cells | FHL2−/− | DC migration (↑) | [95] |
| Tensin | ECs | TNS1−/− | EC migration (↓), EC proliferation (↓) | [96] |
| Actin regulatory layer | ||||
| Ena/VASP family | ECs | Ena/VASP triple null | Oedema, haemorrhaging, stress fibre formation (↓), vascular integrity (↓), embryonic lethality (E18.5) | [97] |
| EVL/VASP | T cells | EVL−/−; VASP−/− | Normal T cell development, activated T cell trafficking (↓), T cell trans endothelial migration (↓) | [98] |
| VASP | Polymorphonuclear leukocytes or neutrophils (PMNs) | VASP−/− | Neutrophil/PMNs adhesion (↓) | [99] |
| Filamin A | T cells | CD4 Cre; flnAfl/fl | Normal T cell development, Teff flow adhesion (↓), T cell trafficking (↓) | [100] |
| Myeloid lineages → neutrophils | LysM Cre; flnAfl/fl | Neutrophil adhesion (↑), normal adhesion underflow | [101] | |
| Myeloid lineages → neutrophils | LysM Cre; flnAfl/fl | Neutrophil recruitment due to inflammatory response (↓) | [102] | |
| Filamin B | ECs | flnb−/− | Microvascular development (↓) | [103] |
| Cdc42 | ECs | Tie2 Cre; Cdc42fl/fl | Haemorrhaging, vascular integrity (↓), EC migration (↓), EC proliferation (↓), embryonic lethality (E9–10) | [104,105] |
| Cad5 CreERT2; Cdc42fl/fl | Retinal angiogenesis (↓), vessel lumen formation (↓), EC adhesion (↓), embryonic lethality | [104] | ||
| T cells | Lck Cre; Cdc42fl/fl | T cell development (↓), CD4+CD8+ double-positive (DP) (↑), CD4+ and CD8+ single-positive (↓), T cell migration (↓), T cell survival (↓), CD8+ Teff (↑), CD8+ T memory (↑) | [106] | |
| Lck Cre; Cdc42fl/fl | T cell survival (↓), mature CD4+ and CD8+ T cells in thymus, spleen and lymph nodes (↓), T cell effector/memory (↑) | [107] | ||
| Rac1 | ECs | Tie2 Cre; Rac1fl/fl | Vessel density (↓), EC adhesion (↓), EC migration (↓), embryonic lethality (E9.5) | [43] |
| Cad5 CreERT2; Rac1fl/fl | Haemorrhaging, vessel density (↓), retinal angiogenesis (↓), embryonic lethality | [108] | ||
| HSCs→ macrophages | Mx1 Cre (Poly(I:C)) ;Rac1fl/fl | Macrophages: elongated morphology, normal migration speed and chemotaxis | [109] | |
| Mx1 Cre (Poly(I:C)) ;Rac1fl/fl | Macrophages: trans endothelial migration (↓) | [110] | ||
| HSCs → T cells | Mx1 Cre (Poly(I:C)) ;Rac1fl/fl | T cells (↓), B cells (↑), bone marrow lymphopoiesis (↓), CD4+CD8+ T cells (↓) | [111] | |
| T cells | Lck Cre;Rac1fl/fl | Normal T cell development | [111] | |
| CD2 Cre;Rac1fl/fl | Normal T cell development | [112] | ||
| B cells | CD19 Cre;Rac1 fl/fl | Normal B cell development | [113] | |
| Rac2 | Macrophages | Rac2−/− | Macrophages: normal trans endothelial migration, elongated morphology, adhesion (↓) | [110] |
| T cells | Rac2−/− | Normal T cell development, T lymphocyte migration (↓), chemotaxis (↓), CD4+ and CD8+ T lymphocytes in spleen, CD8+ T lymphocytes in lymph nodes (↑) | [114] | |
| Rac2−/− | Normal T cell development | [112] | ||
| Rac1 and Rac1 | HSCs→ macrophages | Mx1 Cre (Poly(I:C)); Rac1fl/fl; Rac2−/− | Trans endothelial migration (↓), normal migration speed and chemotaxis, stellate or elongated morphology | [110] |
| HSCs→ T cells | Mx1 Cre (Poly(I:C)); Rac1fl/fl; Rac2−/− | Common lymphoid progenitor (CLP, Lin/IL-7Ra) (↓) | [111] | |
| T cells | Lck Cre; Rac1fl/fl; Rac2−/− | T cell proliferation (↓), T cell survival (↓), adhesion (↓), migration (↓), immature CD4+CD8+ and mature CD4+ in thymus (↓), CD4+ and CD8+ in spleen (↓) | [111] | |
| CD2 Cre; Rac1fl/fl; Rac2−/− | T cell development (↓), CD4+CD8+ double-positive (DP) (↓), CD4+CD8− single-positive (4SP) (↓), CD4−CD8+ single-positive (8SP) (↓) | [112] | ||
| dLck iCre; Rac1fl/fl; Rac2−/− | T cells (↓), T cell chemotaxis (↓), T cell adhesion (↓), homing to secondary lymphoid organs (↓) | [115] | ||
| B cells | CD19 Cre; Rac1 fl/fl; Rac2−/− | Normal B cell development in bone marrow, B cell development in spleen (↓), B cell proliferation (↓), B cell survival (↓) | [113] | |
| CD19 Cre; Rac1 fl/fl; Rac2−/− | B cell development in bone marrow (↓), B cell development in spleen (↓), B cell chemotaxis (↓) | [116] | ||
| RhoA | ECs | Tie2 Cre; RhoAfl/fl | Normal retinal angiogenesis | [117] |
| Cad5 CreERT2; RhoAfl/fl | Normal retinal angiogenesis | [117] | ||
| T cells | Lck Cre; RhoAfl/fl | T cell proliferation (↓), T cell survival n (↓), T cell differentiation n (↓), mature CD4 CD8 T cells in thymus and spleen (↓), trans endothelial migration (↓), Th1 inflammation responses (↓) | [118] | |
| B cells | CD19 Cre; RhoA fl/fl | B cell development (↓) | [119] | |
| HSCs→ B cells | Mx1 Cre (Poly(I:C)); RhoAfl/fl | B cell development (↓) | [119] | |
| Rhoj | ECs | Pdgfb iCreERT2:Rhojfl/fl | Delayed retinal angiogenesis, EC motility (↓) | [120] |
| RhoG, RhoC | B, T cells | RhoG−/−, RhoC−/− | Normal T and B cell development | [121,122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolopoulou, P.A.; Koufaki, M.A.; Kostourou, V. The Adhesome Network: Key Components Shaping the Tumour Stroma. Cancers 2021, 13, 525. https://doi.org/10.3390/cancers13030525
Nikolopoulou PA, Koufaki MA, Kostourou V. The Adhesome Network: Key Components Shaping the Tumour Stroma. Cancers. 2021; 13(3):525. https://doi.org/10.3390/cancers13030525
Chicago/Turabian StyleNikolopoulou, Pinelopi A., Maria A. Koufaki, and Vassiliki Kostourou. 2021. "The Adhesome Network: Key Components Shaping the Tumour Stroma" Cancers 13, no. 3: 525. https://doi.org/10.3390/cancers13030525
APA StyleNikolopoulou, P. A., Koufaki, M. A., & Kostourou, V. (2021). The Adhesome Network: Key Components Shaping the Tumour Stroma. Cancers, 13(3), 525. https://doi.org/10.3390/cancers13030525