
The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients

 , ,
, ,  ,
,  , , , ,
, , , , 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Overall Cohort
2.2. Histological Evaluation
2.3. Genotyping
2.4. UK Biobank Cohort
2.5. Transcriptomic Analysis
2.6. The Cancer Genome Atlas-Liver Hepatocellular Carcinoma (TCGA-LIHC) Data Description
2.7. Statistical Analysis
3. Results
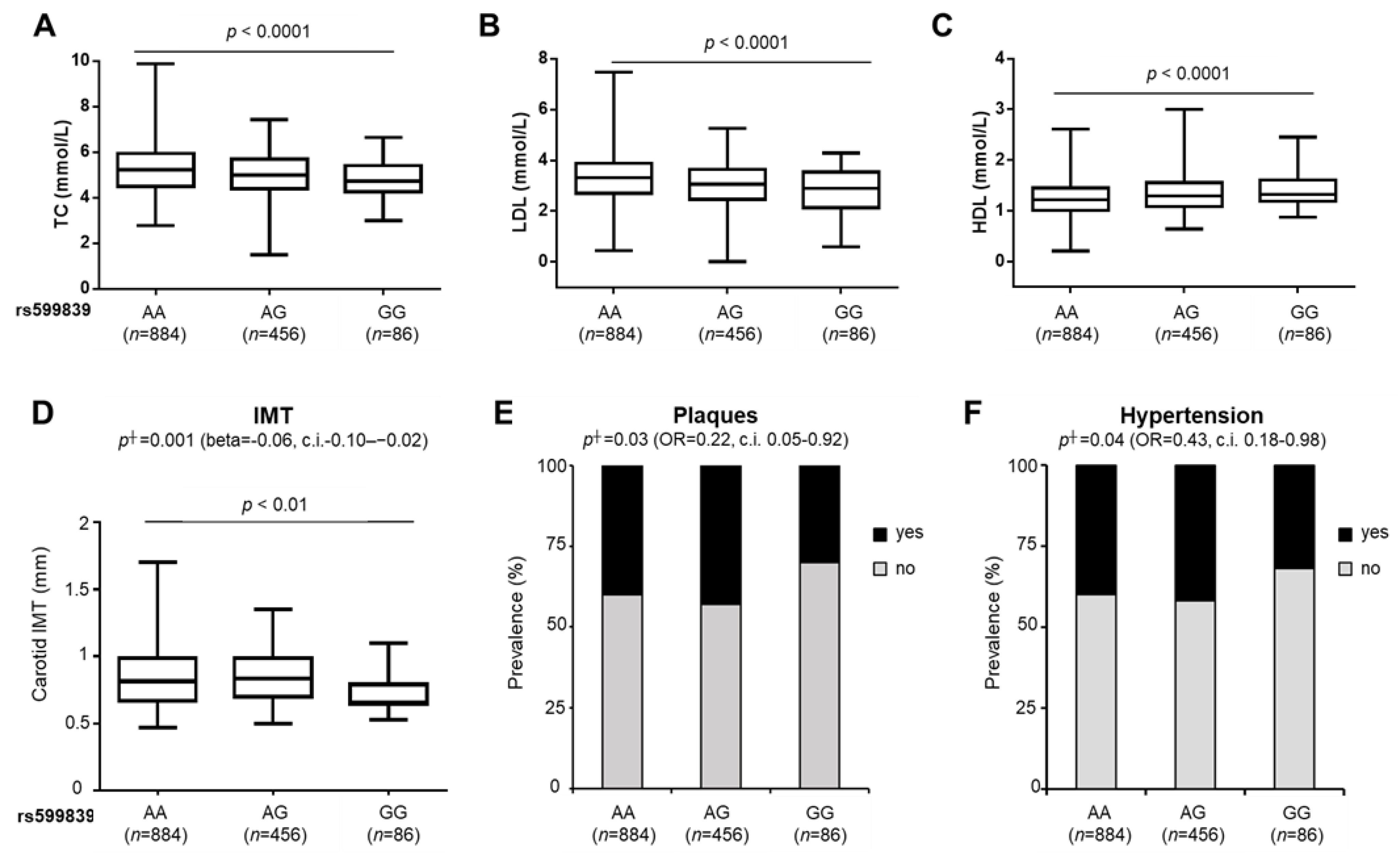
3.1. The rs599839 A>G Gene Variant Affects Circulating Lipids and Cardiovascular Risk in NAFLD Patients
3.2. The rs599839 A>G Variant Is Not Associated with Histological NAFLD
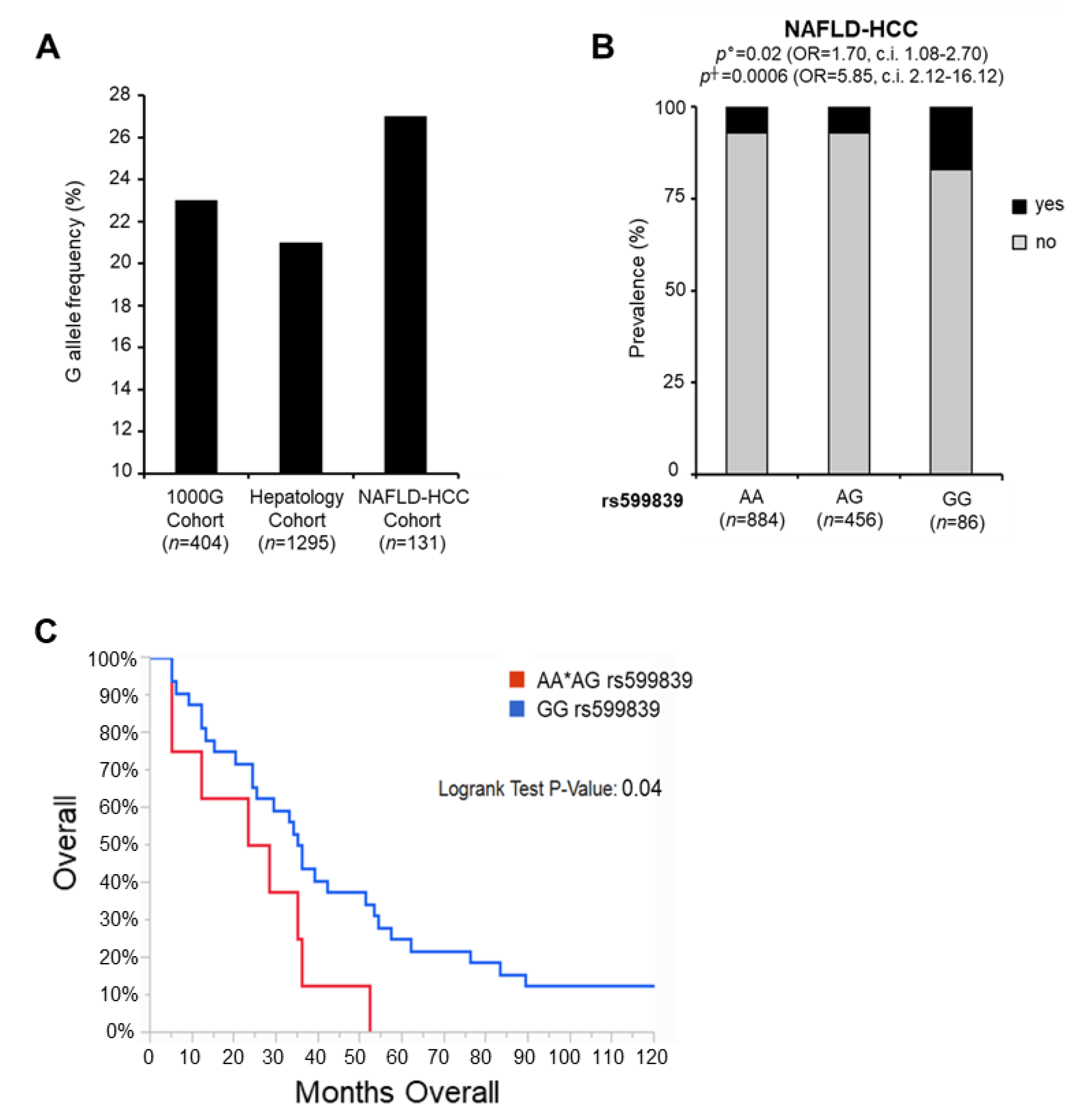
3.3. The rs599839 Variation Is Associated with Increased Risk of HCC in NAFLD Patients
3.4. The rs599839 A>G Gene Variant Affects Circulating Cholesterol and CAD Risk but Not Liver Damage in UKBBC
3.5. Association between rs599839 and Other Genetic Variants at the 1p13.3 Locus
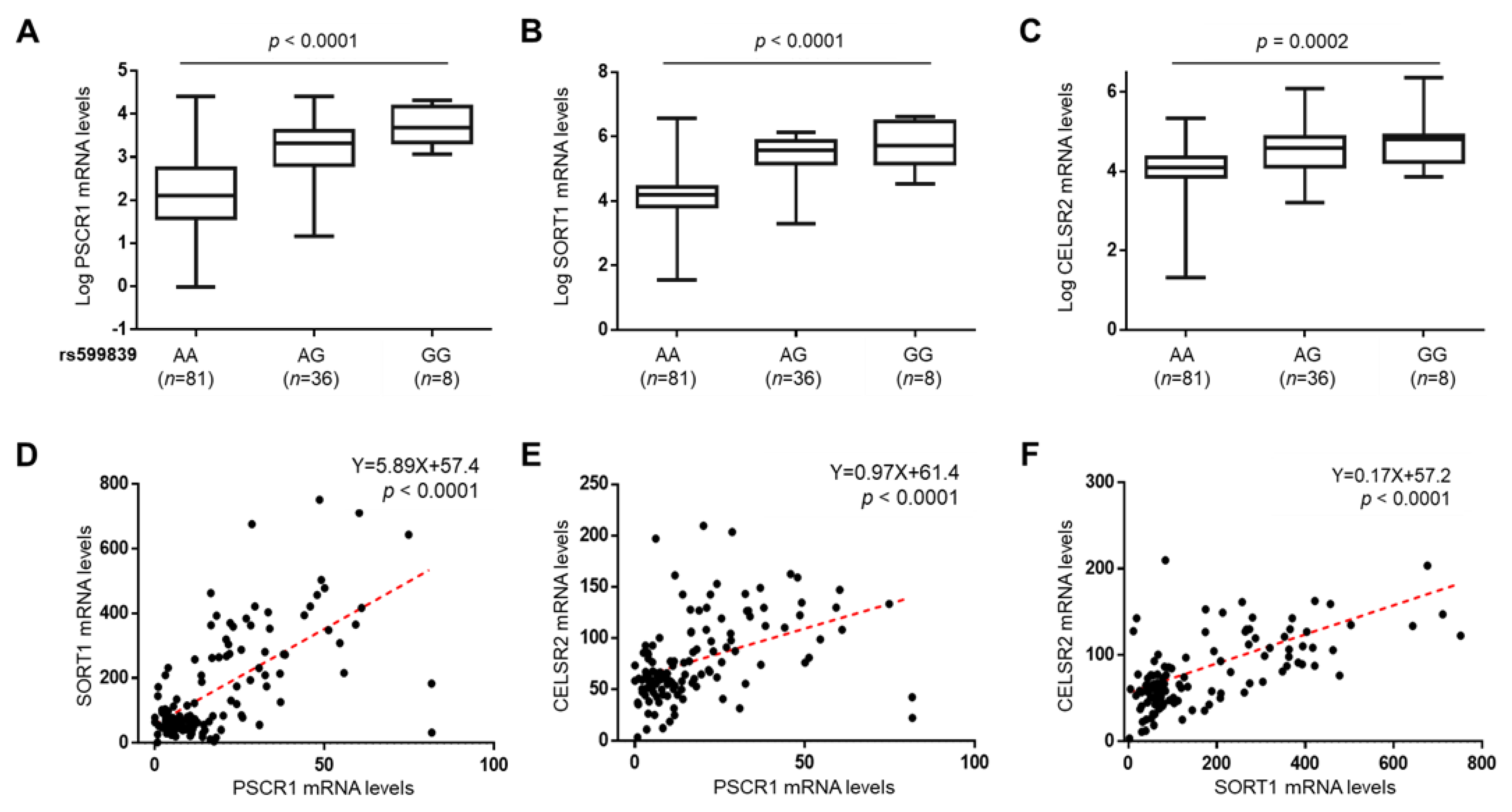
3.6. The rs599839 A>G Impacts on PSRC1, SORT1 and CELSR2 Expression
3.7. PSRC1 Overexpression Aggravates HCC Features and Prognosis in TCGA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
References
- Younossi, Z.; Henry, L. Contribution of Alcoholic and Nonalcoholic Fatty Liver Disease to the Burden of Liver-Related Morbidity and Mortality. Gastroenterology 2016, 150, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Sanyal, A.J.; George, J. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.J.; Aguilar, M.; Cheung, R.; Perumpail, R.B.; Harrison, S.A.; Younossi, Z.M.; Ahmed, A. Nonalcoholic steatohepatitis is the second leading etiology of liver disease among adults awaiting liver transplantation in the United States. Gastroenterology 2015, 148, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, P.; Meroni, M. miRNA Signature in NAFLD: A Turning Point for a Non-Invasive Diagnosis. Int. J. Mol. Sci. 2018, 19, 3966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meroni, M.; Dongiovanni, P.; Longo, M.; Carli, F.; Baselli, G.; Rametta, R.; Pelusi, S.; Badiali, S.; Maggioni, M.; Gaggini, M.; et al. Mboat7 down-regulation by hyper-insulinemia induces fat accumulation in hepatocytes. EBioMedicine 2020, 52, 102658. [Google Scholar] [CrossRef] [Green Version]
- Meroni, M.; Longo, M.; Rustichelli, A.; Dongiovanni, P. Nutrition and Genetics in NAFLD: The Perfect Binomium. Int. J. Mol. Sci. 2020, 21, 2986. [Google Scholar] [CrossRef] [Green Version]
- Meroni, M.; Longo, M. mir-101-3p Downregulation Promotes Fibrogenesis by Facilitating Hepatic Stellate Cell Transdifferentiation during Insulin Resistance. Nutrients 2019, 11, 2597. [Google Scholar] [CrossRef] [Green Version]
- Dongiovanni, P.; Meroni, M.; Baselli, G.; Mancina, R.M.; Ruscica, M.; Longo, M.; Rametta, R.; Cespiati, A.; Pelusi, S.; Ferri, N.; et al. PCSK7 gene variation bridges atherogenic dyslipidemia with hepatic inflammation in NAFLD patients. J. Lipid Res. 2019, 60, 1144–1153. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Petta, S.; Maglio, C.; Fracanzani, A.L.; Pipitone, R.; Mozzi, E.; Motta, B.M.; Kaminska, D.; Rametta, R.; Grimaudo, S.; et al. Transmembrane 6 superfamily member 2 gene variant disentangles nonalcoholic steatohepatitis from cardiovascular disease. Hepatology 2015, 61, 506–514. [Google Scholar] [CrossRef]
- Samani, N.J.; Braund, P.S.; Erdmann, J.; Gotz, A.; Tomaszewski, M.; Linsel-Nitschke, P.; Hajat, C.; Mangino, M.; Hengstenberg, C.; Stark, K.; et al. The novel genetic variant predisposing to coronary artery disease in the region of the PSRC1 and CELSR2 genes on chromosome 1 associates with serum cholesterol. J. Mol. Med. 2008, 86, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Musunuru, K.; Strong, A.; Frank-Kamenetsky, M.; Lee, N.E.; Ahfeldt, T.; Sachs, K.V.; Li, X.; Li, H.; Kuperwasser, N.; Ruda, V.M.; et al. From noncoding variant to phenotype via SORT1 at the 1p13 cholesterol locus. Nature 2010, 466, 714–719. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Hong, S.C.; Yang, Q.; Yin, R.X.; Cao, X.L.; Chen, W.X. Association of variants in CELSR2-PSRC1-SORT1 with risk of serum lipid traits, coronary artery disease and ischemic stroke. Int. J. Clin. Exp. Pathol. 2015, 8, 9543–9551. [Google Scholar] [PubMed]
- Arvind, P.; Nair, J.; Jambunathan, S.; Kakkar, V.V.; Shanker, J. CELSR2-PSRC1-SORT1 gene expression and association with coronary artery disease and plasma lipid levels in an Asian Indian cohort. J. Cardiol. 2014, 64, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.B.; Skaug, J.; Scherer, S.W. The human homologue of flamingo, EGFL2, encodes a brain-expressed large cadherin-like protein with epidermal growth factor-like domains, and maps to chromosome 1p13.3–p21.1. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2000, 7, 233–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, P.C.; Chang, J.C.; Sun, W.T.; Hsieh, S.C.; Wang, M.C.; Wang, F.F. p53 downstream target DDA3 is a novel microtubule-associated protein that interacts with end-binding protein EB3 and activates beta-catenin pathway. Oncogene 2007, 26, 4928–4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, W.J.; Hsieh, S.C.; Chen, C.C.; Wang, F.F. Human DDA3 is an oncoprotein down-regulated by p53 and DNA damage. Biochem. Biophys. Res. Commun. 2008, 369, 567–572. [Google Scholar] [CrossRef]
- Kjolby, M.; Andersen, O.M.; Breiderhoff, T.; Fjorback, A.W.; Pedersen, K.M.; Madsen, P.; Jansen, P.; Heeren, J.; Willnow, T.E.; Nykjaer, A. Sort1, encoded by the cardiovascular risk locus 1p13.3, is a regulator of hepatic lipoprotein export. Cell Metab. 2010, 12, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Mancina, R.M.; Dongiovanni, P.; Petta, S.; Pingitore, P.; Meroni, M.; Rametta, R.; Borén, J.; Montalcini, T.; Pujia, A.; Wiklund, O.; et al. The MBOAT7-TMC4 Variant rs641738 Increases Risk of Nonalcoholic Fatty Liver Disease in Individuals of European Descent. Gastroenterology 2016, 150, 1219–1230.e6. [Google Scholar] [CrossRef] [Green Version]
- Donati, B.; Dongiovanni, P.; Romeo, S.; Meroni, M.; McCain, M.; Miele, L.; Petta, S.; Maier, S.; Rosso, C.; De Luca, L.; et al. MBOAT7 rs641738 variant and hepatocellular carcinoma in non-cirrhotic individuals. Sci. Rep. 2017, 7, 4492. [Google Scholar] [CrossRef] [Green Version]
- Donati, B.; Pietrelli, A.; Pingitore, P.; Dongiovanni, P.; Caddeo, A.; Walker, L.; Baselli, G.; Pelusi, S.; Rosso, C.; Vanni, E.; et al. Telomerase reverse transcriptase germline mutations and hepatocellular carcinoma in patients with nonalcoholic fatty liver disease. Cancer Med. 2017, 6, 1930–1940. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver; European Organisation for Research and Treatment of Cancer. EASL-EORTC clinical practice guidelines: Management of hepatocellular carcinoma. J. Hepatol. 2012, 56, 908–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Dongiovanni, P.; Petta, S.; Mannisto, V.; Mancina, R.M.; Pipitone, R.; Karja, V.; Maggioni, M.; Kakela, P.; Wiklund, O.; Mozzi, E.; et al. Statin use and non-alcoholic steatohepatitis in at risk individuals. J. Hepatol. 2015, 63, 705–712. [Google Scholar] [CrossRef]
- Abecasis, G.R.; Auton, A.; Brooks, L.D.; DePristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1,092 human genomes. Nature 2012, 491, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Baselli, G.A.; Dongiovanni, P. Liver transcriptomics highlights interleukin-32 as novel NAFLD-related cytokine and candidate biomarker. Gut 2020, 69, 1855–1866. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Shima, T.; Mizuno, M.; Mitsumoto, Y.; Umemura, A.; Kanbara, Y.; Tanaka, S.; Sumida, Y.; Yasui, K.; Takahashi, M.; et al. Risk estimation model for nonalcoholic fatty liver disease in the Japanese using multiple genetic markers. PLoS ONE 2018, 13, e0185490. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kogiso, T. Differences in the genetic backgrounds of patients with alcoholic liver disease and non-alcoholic fatty liver disease. JGH Open 2019, 3, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, S.; Voight, B.F.; Purcell, S.; Musunuru, K.; Ardissino, D.; Mannucci, P.M.; Anand, S.; Engert, J.C.; Samani, N.J.; Schunkert, H.; et al. Genome-wide association of early-onset myocardial infarction with single nucleotide polymorphisms and copy number variants. Nat. Genet. 2009, 41, 334–341. [Google Scholar] [CrossRef] [Green Version]
- Kleber, M.E.; Renner, W.; Grammer, T.B.; Linsel-Nitschke, P.; Boehm, B.O.; Winkelmann, B.R.; Bugert, P.; Hoffmann, M.M.; Marz, W. Association of the single nucleotide polymorphism rs599839 in the vicinity of the sortilin 1 gene with LDL and triglyceride metabolism, coronary heart disease and myocardial infarction. The Ludwigshafen Risk and Cardiovascular Health Study. Atherosclerosis 2010, 209, 492–497. [Google Scholar] [CrossRef]
- Saade, S.; Cazier, J.B.; Ghassibe-Sabbagh, M.; Youhanna, S.; Badro, D.A.; Kamatani, Y.; Hager, J.; Yeretzian, J.S.; El-Khazen, G.; Haber, M.; et al. Large scale association analysis identifies three susceptibility loci for coronary artery disease. PLoS ONE 2011, 6, e29427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, G.T.; Bown, M.J.; Gretarsdottir, S.; Romaine, S.P.; Helgadottir, A.; Yu, G.; Tromp, G.; Norman, P.E.; Jin, C.; Baas, A.F.; et al. A sequence variant associated with sortilin-1 (SORT1) on 1p13.3 is independently associated with abdominal aortic aneurysm. Hum. Mol. Genet. 2013, 22, 2941–2947. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, R.; Abe, S.; Tokoro, F.; Arai, M.; Noda, T.; Watanabe, S.; Horibe, H.; Fujimaki, T.; Oguri, M.; Kato, K.; et al. Association of six genetic variants with myocardial infarction. Int. J. Mol. Med. 2015, 35, 1451–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Ding, H.; Zhang, X.; He, M.; Huang, S.; Xu, Y.; Shi, Y.; Cui, G.; Cheng, L.; Wang, Q.K.; et al. Genetic variants at newly identified lipid loci are associated with coronary heart disease in a Chinese Han population. PLoS ONE 2011, 6, e27481. [Google Scholar] [CrossRef]
- Braun, T.R.; Been, L.F.; Singhal, A.; Worsham, J.; Ralhan, S.; Wander, G.S.; Chambers, J.C.; Kooner, J.S.; Aston, C.E.; Sanghera, D.K. A replication study of GWAS-derived lipid genes in Asian Indians: The chromosomal region 11q23.3 harbors loci contributing to triglycerides. PLoS ONE 2012, 7, e37056. [Google Scholar] [CrossRef] [Green Version]
- Fujimaki, T.; Oguri, M.; Horibe, H.; Kato, K.; Matsuoka, R.; Abe, S.; Tokoro, F.; Arai, M.; Noda, T.; Watanabe, S.; et al. Association of a transcription factor 21 gene polymorphism with hypertension. Biomed. Rep. 2015, 3, 118–122. [Google Scholar] [CrossRef]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef]
- Innocenti, F.; Cooper, G.M.; Stanaway, I.B.; Gamazon, E.R.; Smith, J.D.; Mirkov, S.; Ramirez, J.; Liu, W.; Lin, Y.S.; Moloney, C.; et al. Identification, replication, and functional fine-mapping of expression quantitative trait loci in primary human liver tissue. PLoS Genet. 2011, 7, e1002078. [Google Scholar] [CrossRef]
- Conlon, D.M. Role of sortilin in lipid metabolism. Curr. Opin. Lipidol. 2019, 30, 198–204. [Google Scholar] [CrossRef]
- Calandra, S.; Tarugi, P.; Speedy, H.E.; Dean, A.F.; Bertolini, S.; Shoulders, C.C. Mechanisms and genetic determinants regulating sterol absorption, circulating LDL levels, and sterol elimination: Implications for classification and disease risk. J. Lipid Res. 2011, 52, 1885–1926. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.Y.; Cayabyab, F.S.; Tang, C.K.; Zheng, X.L.; Peng, T.H.; Lv, Y.C. Sortilin: A novel regulator in lipid metabolism and atherogenesis. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 460, 11–17. [Google Scholar] [CrossRef]
- Tauris, J.; Ellgaard, L.; Jacobsen, C.; Nielsen, M.S.; Madsen, P.; Thogersen, H.C.; Gliemann, J.; Petersen, C.M.; Moestrup, S.K. The carboxy-terminal domain of the receptor-associated protein binds to the Vps10p domain of sortilin. FEBS Lett. 1998, 429, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Linsel-Nitschke, P.; Heeren, J.; Aherrahrou, Z.; Bruse, P.; Gieger, C.; Illig, T.; Prokisch, H.; Heim, K.; Doering, A.; Peters, A.; et al. Genetic variation at chromosome 1p13.3 affects sortilin mRNA expression, cellular LDL-uptake and serum LDL levels which translates to the risk of coronary artery disease. Atherosclerosis 2010, 208, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Matye, D.J.; Li, T. Insulin resistance induces posttranslational hepatic sortilin 1 degradation in mice. J. Biol. Chem. 2015, 290, 11526–11536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigante, B.; Leander, K.; Vikstrom, M.; Ye, S.; de Faire, U. Chromosome 1p13 genetic variants antagonize the risk of myocardial infarction associated with high ApoB serum levels. BMC Cardiovasc. Disord. 2012, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.M.; Strong, A.; Tohyama, J.; Jin, X.; Morales, C.R.; Billheimer, J.; Millar, J.; Kruth, H.; Rader, D.J. Macrophage sortilin promotes LDL uptake, foam cell formation, and atherosclerosis. Circ. Res. 2015, 116, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.T.; Hsieh, P.C.; Chiang, M.L.; Wang, M.C.; Wang, F.F. p53 target DDA3 binds ASPP2 and inhibits its stimulation on p53-mediated BAX activation. Biochem. Biophys. Res. Commun. 2008, 376, 395–398. [Google Scholar] [CrossRef]
- Hsieh, S.C.; Lo, P.K.; Wang, F.F. Mouse DDA3 gene is a direct transcriptional target of p53 and p73. Oncogene 2002, 21, 3050–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, B.H.; Chao, C.F.; Huang, H.C.; Lee, H.Y.; Kannagi, R. Roles of p53 Family Structure and Function in Non-Canonical Response Element Binding and Activation. Int. J. Mol. Sci. 2019, 20, 3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, P.K.; Chen, J.Y.; Lo, W.C.; Chen, B.F.; Hsin, J.P.; Tang, P.P.; Wang, F.F. Identification of a novel mouse p53 target gene DDA3. Oncogene 1999, 18, 7765–7774. [Google Scholar] [CrossRef] [Green Version]
- Bianco, C.; Jamialahmadi, O.; Pelusi, S.; Baselli, G.; Dongiovanni, P.; Zanoni, I.; Santoro, L.; Maier, S.; Liguori, A.; Meroni, M.; et al. Non-invasive stratification of hepatocellular carcinoma risk in non-alcoholic fatty liver using polygenic risk scores. J. Hepatol. 2021, 74, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, G.; Citro, V.; Capone, D. Nonalcoholic Fatty Liver Disease: A Challenge from Mechanisms to Therapy. J. Clin. Med. 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall Cohort (n = 1426) | Hepatology Service Cohort (n = 1295) | NAFLD-HCC (n = 131) | p-Value † | |
|---|---|---|---|---|
| Sex, M | 770 (54) | 669 (52) | 101 (78) | <0.0001 |
| Age, years | 49.6 ± 13.6 | 48 ± 12.6 | 68 ± 10 | <0.0001 |
| BMI, kg/m2 | 34.2 ± 8.7 | 34.7 ± 8.76 | 28.7 ± 5.12 | <0.0001 |
| T2D, yes | 385 (27) | 310 (24) | 75 (57) | <0.0001 |
| Total cholesterol, mmol/L | 5.1 ± 1.07 | 5.2 ± 1.05 | 4.2 ± 1.17 | <0.0001 |
| LDL cholesterol, mmol/L | 3.1 ± 0.97 | 3.2 ± 0.95 | 2.37 ± 0.96 | <0.0001 |
| HDL cholesterol, mmoL/L | 1.3 ± 0.39 | 1.3 ± 0.4 | 1.28 ± 0.51 | 0.73 |
| Triglycerides, mmoL/L | 1.37 {0.99–1.98} | 1.4 {1.0–2.0} | 1.18 {0.8–1.5} | 0.11 |
| ALT, IU/l | 34 {20–57} | 33 {20–57} | 42 {27–55} | 0.07 |
| AST, IU/l | 25 {19–38} | 25 {18–37} | 39 {26–59} | <0.0001 |
| PNPLA3, I148M | <0.0001 | |||
| I/I | 557 (39) | 524 (41) | 33 (25) | |
| I/M | 627 (44) | 567 (43) | 60 (46) | |
| M/M | 242 (17) | 204 (16) | 38 (29) | |
| TM6SF2, E167K | 0.25 | |||
| E/E | 1240 (87) | 1131 (87.3) | 109 (83.5) | |
| E/K | 168 (11.8) | 150 (11.6) | 18 (14) | |
| K/K | 18 (1.2) | 14 (1.1) | 4 (2.5) | |
| MBOAT7, rs641738 T allele | 0.11 | |||
| C/C | 456 (32) | 423 (33) | 33 (25) | |
| C/T | 642 (45) | 581 (45) | 61 (47) | |
| T/T | 328 (23) | 291 (22) | 37 (28) | |
| rs599839 A>G | 0.01 | |||
| A/A | 884 (62) | 808 (63) | 76 (58) | |
| A/G | 456 (32) | 417 (32) | 39 (30) | |
| G/G | 86 (6) | 70 (5) | 16 (12) |
| AA (n = 884) | AG (n = 456) | GG (n = 86) | p-Value ° | p-Value † | |
|---|---|---|---|---|---|
| Sex, M | 487 (55) | 234 (51) | 49 (56) | 0.33 | 0. 72 |
| Age, years | 49.18 ± 13.5 | 49.7 ± 13.5 | 50.0 ± 14.1 | 0.74 | 0.67 |
| BMI, kg/m2 | 33.9 ± 8.50 | 34.4 ± 8.78 | 35.8 ± 10.4 | 0.16 | 0.10 |
| IFG/T2D, yes (%) | 234 (26) | 1123 (26) | 28 (32) | 0.49 | 0.23 |
| Glucose mg/dL | 102 ± 30 | 103 ± 30 | 107 ± 39 | 0.56 * | 0.18 * |
| HOMA-IR | 5.3 ± 6.4 | 5.5 ± 11 | 4.56 ± 3.87 | 0.91 * | 0.65 * |
| Insulin, IU/mL | 20.7 ± 18.4 | 22.7 ± 34.5 | 15.9 ± 7.88 | 0.97 * | 0.28 * |
| Total cholesterol, mmol/L | 5.2 ± 1.1 | 5.0 ± 1.0 | 4.9 ± 1.0 | 0.005 * | 0. 15 * |
| LDL cholesterol, mmol/L | 3.25 ± 0.97 | 3.03 ± 0.95 | 2.8 ± 0.98 | <0.0001 * | 0.03 * |
| HDL cholesterol, mmol/L | 1.26 ± 0.3 | 1.34 ± 0.43 | 1.36 ± 0.37 | 0.0003 * | 0.12 * |
| Triglycerides, mmol/L | 1.41 {1.0–2.0} | 1.32 {0.92–1.97} | 1.36 {1.02–1.97} | 0.28 * | 0.83 * |
| Dyslipidemia, yes (%) | 300 (34) | 127 (28) | 15 (18) | 0.01 * | 0.007 * |
| ALT, IU/L | 35 {21–56} | 33 {20–62} | 28 {19–52} | 0.26 * | 0.78 * |
| AST, IU/L | 26 {19–38} | 25 {19–39} | 24{18–36} | 0.76 * | 0.72 * |
| Iron ug/dL | 97.8 ± 44.4 | 96.8 ± 40.8 | 9.48 ± 33.6 | 0.23 ** | 0.67 ** |
| Transferrin mg/dL | 270.4 ± 60.8 | 264.6 ± 47.2 | 258.6 ± 47.0 | 0.13 ** | 0.48 ** |
| Transferrin saturation (%) | 30.1 ± 24.9 | 29.1 ± 15.2 | 34.9 ± 43.9 | 0.79 ** | 0.06 ** |
| Ferritin ng/mL | 324.4 ± 411.6 | 386.2 ± 508.2 | 459.2 ± 526.9 | 0.28 ** | 0.31 ** |
| IMT | Plaques, Yes | Hypertension, Yes | |||||||
|---|---|---|---|---|---|---|---|---|---|
| β | 95% CI | p-Value | OR | 95% CI | p-Value | OR | 95% CI | p-Value | |
| Sex, M | 0.05 | 0.02–0.06 | <0.0001 | 1.78 | 1.04–3.06 | 0.03 | 1.32 | 0.95–1.84 | 0.09 |
| Age, years | 0.008 | 0.006–0.009 | <0.0001 | 1.09 | 1.07–1.12 | <0.0001 | 1.06 | 1.05–1.07 | <0.0001 |
| BMI, kg/m2 | 0.001 | −0.003–0.005 | 0.54 | 1.02 | 0.97–1.07 | 0.41 | 1.06 | 1.04–1.08 | <0.0001 |
| IFG/T2D, yes (%) | 0.002 | −0.02–0.02 | 0.86 | 1.54 | 0.91–2.61 | 0.11 | 1.68 | 1.18–2.38 | 0.003 |
| TM6SF2, E167K allele | −0.02 | −0.06–0.01 | 0.22 | 0.78 | 0.46–1.34 | 0.37 | 1.71 | 1.16–2.51 | 0.006 |
| Statin use, yes | −0.01 | −0.04–0.01 | 0.38 | 0.43 | 0.22–0.83 | 0.01 | 0.97 | 0.56–1.66 | 0.91 |
| Active smoking | 0.01 | −0.01–0.03 | 0.36 | 1.840 | 1.08–3.14 | 0.02 | 1.30 | 0.92–1.84 | 0.12 |
| rs599839 GG yes | −0.06 | −0.10–0.02 | 0.001 | 0.22 | 0.05–0.92 | 0.03 | 0.43 | 0.18–0.98 | 0.04 |
| Steatosis | Lobular Inflammation | Ballooning | Fibrosis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | 95% CI | p-Value | β | 95% CI | p-Value | β | 95% CI | p-Value | β | 95% CI | p-Value | |
| Sex, M | 0.29 | 0.17–0.41 | <0.0001 | 0.25 | 0.13–0.37 | <0.0001 | 0.29 | 0.14–0.44 | <0.0001 | 0.32 | 0.20–0.44 | <0.0001 |
| Age, years | 0.002 | −0.007–0.01 | 0.67 | 0.02 | 0.01–0.03 | <0.0001 | 0.03 | 0.02–0.04 | <0.0001 | 0.07 | 0.04–0.06 | <0.0001 |
| BMI, kg/m2 | 0.04 | 0.02–0.05 | <0.0001 | 0.03 | 0.01–0.04 | <0.0001 | −0.003 | −0.02–0.01 | 0.67 | 0.002 | −0.01–0.015 | 0.81 |
| IFG/T2D, yes (%) | 0.33 | 0.19–0.47 | <0.0001 | 0.32 | 0.18–0.47 | <0.0001 | 0.32 | 0.17–0.48 | <0.0001 | 0.60 | 0.47–0.74 | <0.0001 |
| PNPLA3, I148M allele | 0.43 | 0.28–0.57 | <0.0001 | 0.35 | 0.19–0.50 | <0.0001 | 0.28 | 0.10–0.47 | 0.002 | 0.49 | 0.34–0.64 | <0.0001 |
| TM6SF2, E167K allele | 0.75 | 0.46–1.04 | <0.0001 | 0.57 | 0.28–0.86 | 0.0001 | 0.41 | 0.08–0.75 | 0.01 | 0.57 | 0.29–0.84 | <0.0001 |
| MBOAT7, rs641738 T allele | 0.11 | −0.04–0.25 | 0.14 | 0.07 | −0.08–0.22 | 0.33 | 0.02 | −0.15–0.20 | 0.76 | 0.16 | 0.01–0.31 | 0.02 |
| rs599839 G allele | −0.02 | −0.19–0.16 | 0.83 | −0.02 | −0.21–0.16 | 0.80 | 0.20 | −0.02–0.42 | 0.07 | 0.02 | −0.16–0.19 | 0.82 |
| HCC ° | HCC † | HCC ° Adjusted for f > 2 | HCC † Adjusted for f > 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OR | 95% CI | p-Value | OR | 95% CI | p-Value | OR | 95% CI | p-Value | OR | 95% CI | p-Value | |
| Sex, M | 1.82 | 0.98–3.36 | 0.05 | 1.64 | 0.89–3.01 | 0.10 | 2.23 | 1.14–4.34 | 0.01 | 2.08 | 1.07–4.06 | 0.03 |
| Age, years | 1.16 | 1.12–1.20 | <0.0001 | 1.16 | 1.12–1.20 | <0.0001 | 1.13 | 1.09–1.17 | <0.0001 | 1.13 | 1.09–1.17 | <0.0001 |
| BMI, kg/m2 | 0.87 | 0.82–0.93 | <0.0001 | 0.88 | 0.83–0.93 | <0.0001 | 0.90 | 0.84–0.96 | 0.0009 | 0.90 | 0.84–0.96 | 0.001 |
| IFG/T2D, yes (%) | 3.22 | 1.79–5.78 | <0.0001 | 3.25 | 1.80–5.86 | <0.0001 | 1.86 | 0.96–3.57 | 0.06 | 1.90 | 0.98–3.67 | 0.05 |
| PNPLA3, I148M allele | 1.35 | 0.93–1.95 | 0.11 | 1.34 | 0.92–1.94 | 0.12 | 1.02 | 0.67–1.53 | 0.93 | 1.01 | 0.67–1.52 | 0.95 |
| TM6SF2, E167K allele | 1.51 | 0.84–2.72 | 0.16 | 1.47 | 0.82–2.65 | 0.19 | 1.20 | 0.64–2.25 | 0.57 | 1.17 | 0.61–2.17 | 0.63 |
| MBOAT7, rs641738 T allele | 1.33 | 0.90–1.96 | 0.14 | 1.39 | 0.94–2.06 | 0.09 | 1.37 | 0.90–2.09 | 0.14 | 1.42 | 0.93–2.18 | 0.10 |
| rs599839 G allele | 1.70 | 1.08–2.70 | 0.02 | 5.85 | 2.12–16.12 | 0.0006 | 1.70 | 1.03–2.80 | 0.03 | 5.62 | 1.77–17.84 | 0.003 |
| Stage (S) > 1 | Tumor size (T) > 1 | Child-Pugh > A6 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| OR | 95% CI | p-Value | OR | 95% CI | p-Value | OR | 95% CI | p-Value | |
| Sex, M | 1.10 | 0.33–3.65 | 0.87 | 0.82 | 0.234–2.83 | 0.01 | 0.47 | 0.08–2.90 | 0.42 |
| Age, years | 0.99 | 0.94–1.04 | 0.67 | 0.98 | 0.93–1.04 | 0.82 | 0.97 | 0.89–1.05 | 0.51 |
| PNPLA3, I148M allele | 0.61 | 0.30–1.23 | 0.16 | 0.41 | 0.19–0.85 | 0.93 | 0.43 | 0.13–1.45 | 0.17 |
| TM6SF2, E167K allele | 1.08 | 0.42–2.81 | 0.87 | 1.27 | 0.47–3.41 | 0.57 | 0.21 | 0.22–1.89 | 0.16 |
| MBOAT7, rs641738 T allele | 0.73 | 0.34–1.55 | 0.41 | 0.66 | 0.30–1.44 | 0.29 | 0.43 | 0.11–1.63 | 0.21 |
| rs599839 G allele | 3.27 | 1.36–7.85 | 0.008 | 2.86 | 1.17–6.96 | 0.02 | 4.38 | 1.35–14.22 | 0.01 |
| Phenotype | Cases | β | OR | p-Value |
|---|---|---|---|---|
| K70: Alcoholic liver disease | 808 | −1.99 × 10−5 | 1.01 | 0.86 |
| K74: Fibrosis and cirrhosis of liver | 805 | 3.31 × 10−6 | 0.99 | 0.97 |
| K75: Other inflammatory liver diseases | 662 | −9.95 × 10−5 | 1.06 | 0.32 |
| K76: Other diseases of liver | 3351 | −5.32 × 10−6 | 1 | 0.98 |
| K70-K77: Diseases of liver | 4894 | −0.0002 | 1.02 | 0.39 |
| High cholesterol | 55,265 | −0.017 | 1.17 | 7.82 × 10−112 |
| E78 Disorders of lipoprotein metabolism and other lipidaemias | 39,308 | −0.010 | 1.14 | 4.3 × 10−55 |
| E70-E90 Metabolic disorders | 47,969 | −0.010 | 1.11 | 3.09 × 10−44 |
| I20-I25 Ischaemic heart diseases | 33,387 | −0.006 | 1.1 | 6.72 × 10−25 |
| I25 Chronic ischaemic heart disease | 27,772 | −0.006 | 1.11 | 9.76 × 10−25 |
| I20 Angina pectoris | 19,935 | −0.004 | 1.1 | 3.85 × 10−17 |
| Heart/cardiac problem | 32,474 | −0.005 | 1.08 | 2.29 × 10−16 |
| Angina | 14,399 | −0.003 | 1.12 | 2.88 × 10−16 |
| Heart attack/myocardial infarction | 10,356 | −0.002 | 1.13 | 3.84 × 10−14 |
| I21 Acute myocardial infarction | 8764 | −0.002 | 1.11 | 4.05 × 10−9 |
| I30-I52 Other forms of heart disease | 31,135 | −0.002 | 1.04 | 0.0002 |
| I50 Heart failure | 5901 | −0.001 | 1.08 | 0.0003 |
| I71 Aortic aneurysm and dissection | 1470 | −0.0004 | 1.14 | 0.003 |
| Pace-maker | 1355 | −0.0003 | 1.12 | 0.013 |
| I10-I15 Hypertensive diseases | 84,910 | −0.002 | 1.01 | 0.018 |
| G45 Transient cerebral ischaemic attacks and related syndromes | 2765 | −0.0004 | 1.07 | 0.03 |
| I70 Atherosclerosis | 1371 | −0.0003 | 1.1 | 0.037 |
| PSRC1 Expression | |||
|---|---|---|---|
| β | 95% CI | p-Value | |
| Sex, M | 0.04 | −0.08–0.31 | 0.50 |
| Ethnicity; Hispanic | 0.08 | −0.18–0.35 | 0.55 |
| Diagnosis Age | −0.0003 | −0.01–0.006 | 0.50 |
| Stage (S) >1 | 0.7 | 0.05–0.27 | 0.006 |
| Sex, M | 0.06 | −0.06–0.18 | 0.31 |
| Ethnicity; Hispanic | 0.10 | −0.16–0.36 | 0.45 |
| Diagnosis Age | −0.001 | −0.01–0.07 | 0.67 |
| Grade (G) >2 | 0.17 | 0.05–0.29 | 0.005 |
| Sex, M | 0.05 | −0.07–0.17 | 0.41 |
| Ethnicity; Hispanic | 0.08 | −0.18–0.34 | 0.54 |
| Diagnosis Age | −0.002 | −0.01–0.006 | 0.53 |
| Tumor size (T) >1 | 0.17 | 0.06–0.28 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meroni, M.; Longo, M.; Paolini, E.; Alisi, A.; Miele, L.; De Caro, E.R.; Pisano, G.; Maggioni, M.; Soardo, G.; Valenti, L.V.; et al. The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients. Cancers 2021, 13, 1783. https://doi.org/10.3390/cancers13081783
Meroni M, Longo M, Paolini E, Alisi A, Miele L, De Caro ER, Pisano G, Maggioni M, Soardo G, Valenti LV, et al. The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients. Cancers. 2021; 13(8):1783. https://doi.org/10.3390/cancers13081783
Chicago/Turabian StyleMeroni, Marica, Miriam Longo, Erika Paolini, Anna Alisi, Luca Miele, Emilia Rita De Caro, Giuseppina Pisano, Marco Maggioni, Giorgio Soardo, Luca Vittorio Valenti, and et al. 2021. "The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients" Cancers 13, no. 8: 1783. https://doi.org/10.3390/cancers13081783
APA StyleMeroni, M., Longo, M., Paolini, E., Alisi, A., Miele, L., De Caro, E. R., Pisano, G., Maggioni, M., Soardo, G., Valenti, L. V., Fracanzani, A. L., & Dongiovanni, P. (2021). The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients. Cancers, 13(8), 1783. https://doi.org/10.3390/cancers13081783