
Characterization of Cell-Bound CA125 on Immune Cell Subtypes of Ovarian Cancer Patients Using a Novel Imaging Platform



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Sample Staining
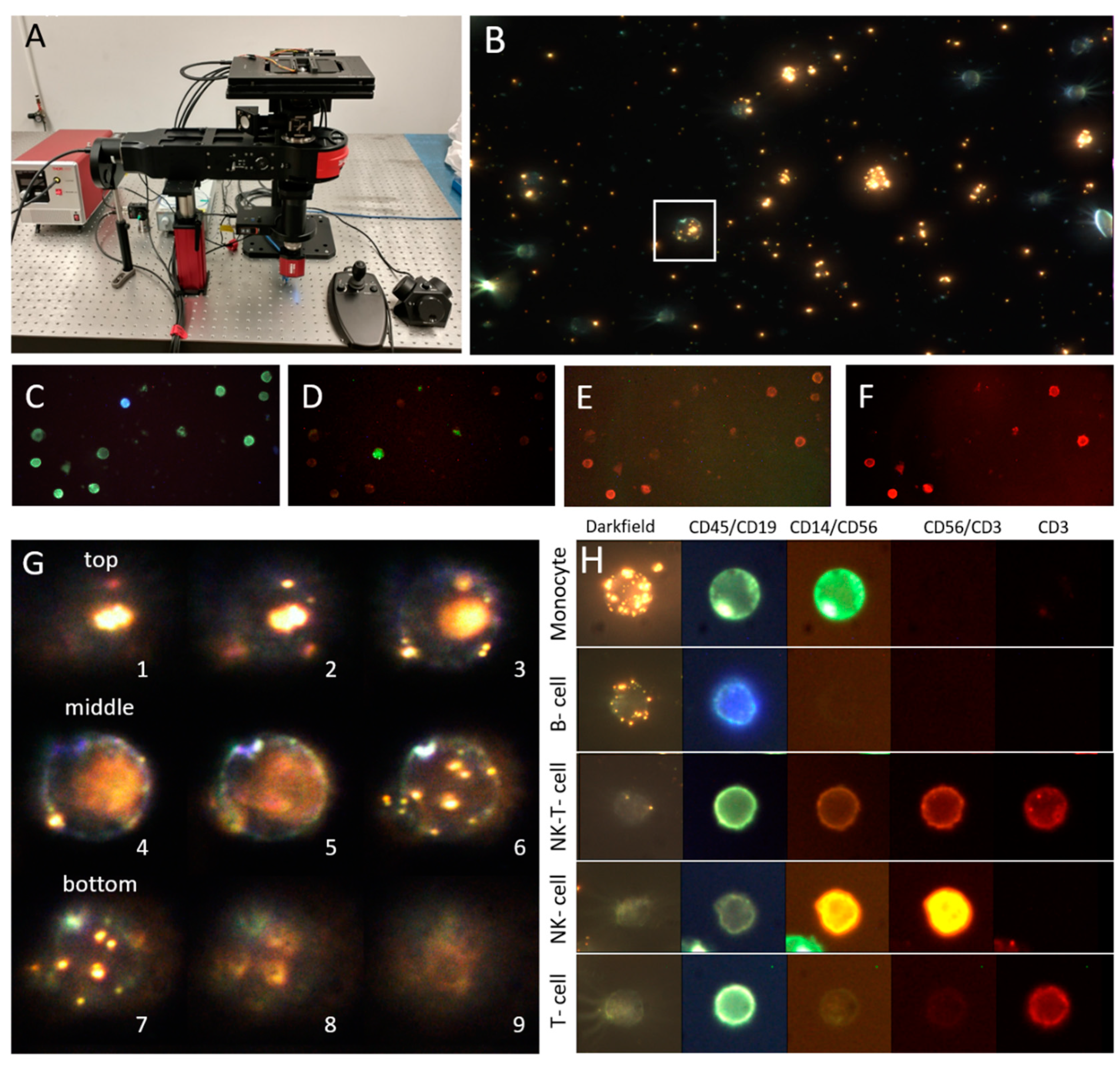
2.3. Microscope and Image Acquisition
2.4. Image Processing and Data Analysis
2.4.1. Fluorescent Imaging and Color Compensation
2.4.2. Cell Finding on CD45 Images
2.4.3. Cell Contour Delineation
2.4.4. Cell Lineage Determination
2.4.5. PNP Quantification
2.4.6. Data Aggregation
2.5. Statistical Analysis
3. Results
3.1. Cohort
3.2. Technical Validation
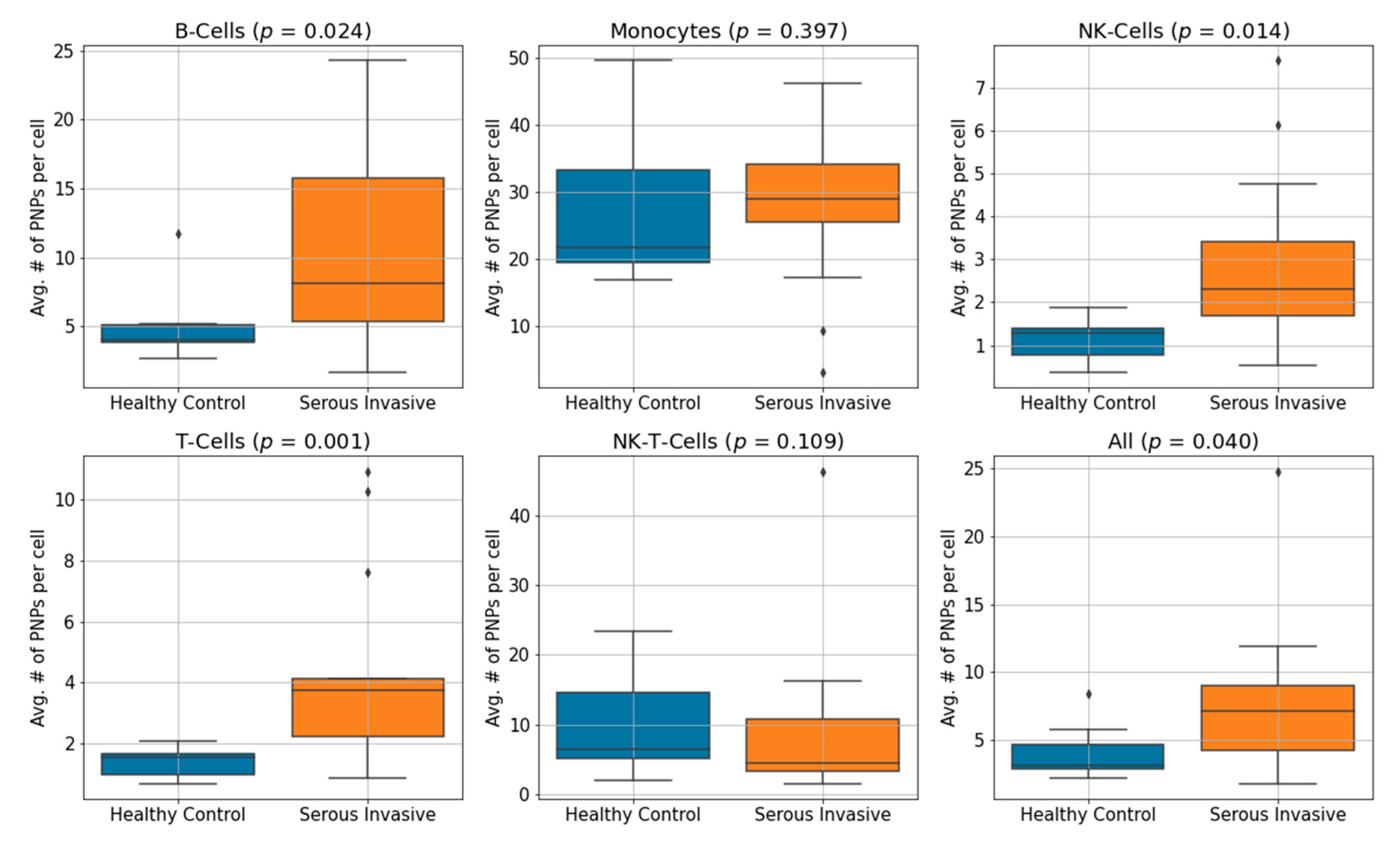
3.3. Serous Invasive Ovarian Cancer Patients Show an Elevation of CA125 Bound to Immune Cell Subsets Compared to Healthy Controls
3.4. Immune Cell-Bound CA125 Does Not Correlate to Serum CA125 Levels in Serous Invasive Ovarian Cancer Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Brien, T.J.; Beard, J.B.; Underwood, L.J.; Dennis, R.A.; Santin, A.D.; York, L. The CA 125 Gene: An Extracellular Superstructure Dominated by Repeat Sequences. Tumor Biol. 2001, 22, 348–366. [Google Scholar] [CrossRef]
- Felder, M.; Kapur, A.; Gonzalez-Bosquet, J.; Horibata, S.; Heintz, J.; Albrecht, R.; Fass, L.; Kaur, J.; Hu, K.; Shojaei, H.; et al. MUC16 (CA125): Tumor biomarker to cancer therapy, a work in progress. Mol. Cancer 2014, 13, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, K.O.; Yin, B.W.T.; Kudryashov, V. Isolation and characterization of ovarian cancer antigen CA 125 using a new monoclonal antibody (VK-8): Identification as a mucin-type molecule. Int. J. Cancer 1997, 71, 842–850. [Google Scholar] [CrossRef]
- Medeiros, L.R.; Rosa, D.D.; da Rosa, M.I.; Bozzetti, M.C. Accuracy of CA 125 in the diagnosis of ovarian tumors: A quantitative systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 142, 99–105. [Google Scholar] [CrossRef]
- Bast, R.C.; Feeney, M.; Lazarus, H.; Nadler, L.M.; Colvin, R.B.; Knapp, R.C. Reactivity of a monoclonal antibody with human ovarian carcinoma. J. Clin. Investig. 1981, 68, 1331–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuxen, M.K.; Sölétormos, G.; Dombernowsky, P. Serum tumour marker CA 125 in monitoring of ovarian cancer during first-line chemotherapy. Br. J. Cancer 2001, 84, 1301–1307. [Google Scholar] [CrossRef] [Green Version]
- Rustin, G.J.S.; Nelstrop, A.E.; Tuxen, M.K.; Lambert, H.E. Defining progression of ovarian carcinoma during follow-up according to CA 125: A North Thames Ovary Group study. Ann. Oncol. 1996, 7, 361–364. [Google Scholar] [CrossRef]
- Geisler, J.P.; Miller, A.G.; Lee, T.H.; Harwood, R.M.; Wiemann, M.C.; Geisler, E.H. Relationship of preoperative serum CA-125 to survival in epithelial ovarian carcinoma. J. Reprod. Med. 1996, 41, 140–142. [Google Scholar] [PubMed]
- Im, S.S.; Gordon, A.N.; Buttin, B.M.; Leath, C.A.; Gostout, B.S.; Shah, C.; Hatch, K.D.; Wang, J.; Berman, M.L. Validation of Referral Guidelines for Women with Pelvic Masses. Obstet. Gynecol. 2005, 105, 35–41. [Google Scholar] [CrossRef]
- Charkhchi, P.; Cybulski, C.; Gronwald, J.; Wong, F.O.; Narod, S.A.; Akbari, M.R. CA125 and Ovarian Cancer: A Comprehensive Review. Cancers 2020, 12, 3730. [Google Scholar] [CrossRef]
- Belisle, J.A.; Horibata, S.; Gubbels, J.A.; Petrie, S.; Kapur, A.; Andre, S.; Gabius, H.-J.; Rancourt, C.; Connor, J.; Paulson, J.C.; et al. Identification of Siglec-9 as the receptor for MUC16 on human NK cells, B cells, and monocytes. Mol. Cancer 2010, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Patankar, M.S.; Jing, Y.; Morrison, J.C.; Belisle, J.A.; Lattanzio, F.A.; Deng, Y.; Wong, N.K.; Morris, H.R.; Dell, A.; Clark, G.F. Potent suppression of natural killer cell response mediated by the ovarian tumor marker CA125. Gynecol. Oncol. 2005, 99, 704–713. [Google Scholar] [CrossRef]
- Gubbels, A.A.J.; Felder, M.; Horibata, S.; Belisle, A.J.; Kapur, A.; Holden, H.; Petrie, S.; Migneault, M.; Rancourt, C.; Connor, J.P.; et al. MUC16 provides immune protection by inhibiting synapse formation between NK and ovarian tumor cells. Mol. Cancer 2010, 9, 11–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felder, M.; Kapur, A.; Rakhmilevich, A.L.; Qu, X.; Sondel, P.M.; Gillies, S.D.; Connor, J.; Patankar, M.S. MUC16 suppresses human and murine innate immune responses. Gynecol. Oncol. 2019, 152, 618–628. [Google Scholar] [CrossRef]
- Yguerabide, J.; Yguerabide, E.E.; Bee, G.; Yamout, K.; Korb, L.; Beck, J.; Peterson, T. Resonance light-scattering particles for ultra-sensitive detection of nucleic acids on microarrays. Nat. Genet. 1999, 23, 67–68. [Google Scholar] [CrossRef]
- Yguerabide, J.; Yguerabide, E.E. Resonance light scattering particles as ultrasensitive labels for detection of analytes in a wide range of applications. J. Cell. Biochem. 2001, 84, 71–81. [Google Scholar] [CrossRef]
- Peng, F.; Jeong, S.; Gonzalez, G.; Marks, H.; Ho, A.; Roussakis, E.; Krauledat, P.B.; Hansen, P.; Evans, C.L. Assessment of Glial Fibrillary Acidic Protein Binding to the Surface of Leukocytes with Dark-Field Imaging and Computational Analysis. Adv. Funct. Mater. 2021, 31, 1–9. [Google Scholar] [CrossRef]
- Jeong, S.; González, G.; Ho, A.; Nowell, N.; Austin, L.A.; Hoballah, J.; Mubarak, F.; Kapur, A.; Patankar, M.S.; Cramer, D.W.; et al. Plasmonic Nanoparticle-Based Digital Cytometry to Quantify MUC16 Binding on the Surface of Leukocytes in Ovarian Cancer. ACS Sens. 2020, 5, 2772–2782. [Google Scholar] [CrossRef]
- Smereka, M.; Dulęba, I. Circular Object Detection Using a Modified Hough Transform. Int. J. Appl. Math. Comput. Sci. 2008, 18, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. In Proceedings of the International Conference on Medical Image Computing and Computer-Assisted Intervention, Munich, Germany, 5–9 October 2015; Springer: Berlin/Heidelberg, Germany, 2015; pp. 234–241. [Google Scholar]
- Falk, T.; Mai, D.; Bensch, R.; Çiçek, Ö.; Abdulkadir, A.; Marrakchi, Y.; Böhm, A.; Deubner, J.; Jäckel, Z.; Seiwald, K.; et al. U-Net: Deep learning for cell counting, detection, and morphometry. Nat. Methods 2019, 16, 67–70. [Google Scholar] [CrossRef]
- Sudre, C.H.; Li, W.; Vercauteren, T.; Ourselin, S.; Cardoso, M.J. Generalised Dice Overlap as a Deep Learning Loss Function for Highly Unbalanced Segmentations. In Transactions on Petri Nets and Other Models of Concurrency XV; Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany, 2017; Volume 10553, pp. 240–248. [Google Scholar] [CrossRef] [Green Version]
- González, G.; Evans, C.L. Biomedical Image Processing with Containers and Deep Learning: An Automated Analysis Pipeline. BioEssays 2019, 41, e1900004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, E.; Sander, J.; Ester, M.; Kriegel, H.P.; Xu, X. DBSCAN Revisited, Revisited. ACM Trans. Database Syst. 2017, 42, 1–21. [Google Scholar] [CrossRef]
- Yin, B.W.; Lloyd, K.O. Molecular Cloning of the CA125 Ovarian Cancer Antigen. J. Biol. Chem. 2001, 276, 27371–27375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, R.; Marcos-Silva, L.; Ricardo, C.; Ponte, F.; Costa, A.; Lopes, J.M.; David, L. Peritoneal dissemination of ovarian cancer: Role of MUC16-mesothelin interaction and implications for treatment. Expert Rev. Anticancer. Ther. 2018, 18, 177–186. [Google Scholar] [CrossRef]
- Chhour, P.; Kim, J.; Benardo, B.; Tovar, A.; Mian, S.; Litt, H.I.; Ferrari, V.A.; Cormode, D.P. Effect of Gold Nanoparticle Size and Coating on Labeling Monocytes for CT Tracking. Bioconjugate Chem. 2017, 28, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, N.K.; Easton, R.L.; Panico, M.; Sutton-Smith, M.; Morrison, J.C.; Lattanzio, F.A.; Morris, H.R.; Clark, G.F.; Dell, A.; Patankar, M.S. Characterization of the Oligosaccharides Associated with the Human Ovarian Tumor Marker CA125. J. Biol. Chem. 2003, 278, 28619–28634. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.K.; Lockhart, D.C.; Von Bernstorff, W.; Spanjaard, R.A.; JOO, H.G.; Eberlein, T.J.; Goedegebuure, P.S. Soluble MUC1 secreted by human epithelial cancer cells mediates immune suppression by blocking T-cell activation. Int. J. Cancer 1999, 82, 721–726. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Siddiqui, S.S.; Trefny, M.P.; Thommen, D.S.; Boligan, K.F.; Von Gunten, S.; Tzankov, A.; Tietze, L.; Lardinois, D.; Heinzelmann-Schwarz, V.; et al. Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells. J. Clin. Investig. 2018, 128, 4912–4923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedermann, B.; Gil, D.; Bowen, D.T.; Crocker, P.R. Analysis of the CD33-related siglec family reveals that Siglec-9 is an endocytic receptor expressed on subsets of acute myeloid leukemia cells and absent from normal hematopoietic progenitors. Leuk. Res. 2007, 31, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Skates, S.J.; Mai, P.; Horick, N.K.; Piedmonte, M.; Drescher, C.W.; Isaacs, C.; Armstrong, D.K.; Buys, S.S.; Rodriguez, G.C.; Horowitz, I.R.; et al. Large Prospective Study of Ovarian Cancer Screening in High-Risk Women: CA125 Cut-Point Defined by Menopausal Status. Cancer Prev. Res. 2011, 4, 1401–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| T Cells | B Cells | NK-T Cells | Monocytes | NK Cells | |
|---|---|---|---|---|---|
| Flow cytometry | 74.8% | 11.9% | 7.14% | 6.27% | 4.52% |
| New platform (std) | 77.2% (6.7) | 7.7% (4.9) | 6.2% (1.9) | 6.4% (0.7) | 1.2% (0.2) |
| All PBMC | NK Cells | T Cells | Monocytes | B Cells | NK-T Cells | |
|---|---|---|---|---|---|---|
| Controls | 4.11 | 1.11 | 1.38 | 27.58 | 5.19 | 10.03 |
| Patients | 7.78 | 2.86 | 4.36 | 27.71 | 10.45 | 9.10 |
| p-value | 0.04 | 0.014 | 0.001 | 0.400 | 0.024 | 0.109 |
| NK CA125 | T CA125 | Monocyte CA125 | NK-T-CA125 | Serum CA125 * | ||
|---|---|---|---|---|---|---|
| Patients | B CA125 | 0.67 (<0.01) | 0.68 (<0.01) | 0.32 (0.03) | 0.22 (0.09) | 0.07 (0.42) |
| NK CA125 | 0.80 (<0.01) | 0.18 (0.12) | 0.14 (0.19) | 0.08 (0.39) | ||
| T CA125 | 0.23 (0.08) | 0.49 (<0.01) | 0.03 (0.62) | |||
| Monocyte CA125 | 0.26 (0.06) | 0.06 (0.44) | ||||
| NK-T-CA125 | 0.08 (0.36) | |||||
| All PBMCs CA125 | 0.01 (0.81) | |||||
| Healthy Controls | B CA125 | 0.17 (0.35) | 0.45 (0.10) | 0.71 (0.02) | 0.12 (0.50) | |
| NK CA125 | 0.31 (0.19) | 0.03 (0.71) | 0.18 (0.39) | |||
| T CA125 | 0.48 (0.09) | 0.06 (0.64) | ||||
| Monocyte CA125 | 0.01 (0.84) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, G.; Lakatos, K.; Hoballah, J.; Fritz-Klaus, R.; Al-Johani, L.; Brooker, J.; Jeong, S.; Evans, C.L.; Krauledat, P.; Cramer, D.W.; et al. Characterization of Cell-Bound CA125 on Immune Cell Subtypes of Ovarian Cancer Patients Using a Novel Imaging Platform. Cancers 2021, 13, 2072. https://doi.org/10.3390/cancers13092072
González G, Lakatos K, Hoballah J, Fritz-Klaus R, Al-Johani L, Brooker J, Jeong S, Evans CL, Krauledat P, Cramer DW, et al. Characterization of Cell-Bound CA125 on Immune Cell Subtypes of Ovarian Cancer Patients Using a Novel Imaging Platform. Cancers. 2021; 13(9):2072. https://doi.org/10.3390/cancers13092072
Chicago/Turabian StyleGonzález, Germán, Kornél Lakatos, Jawad Hoballah, Roberta Fritz-Klaus, Lojain Al-Johani, Jeff Brooker, Sinyoung Jeong, Conor L. Evans, Petra Krauledat, Daniel W. Cramer, and et al. 2021. "Characterization of Cell-Bound CA125 on Immune Cell Subtypes of Ovarian Cancer Patients Using a Novel Imaging Platform" Cancers 13, no. 9: 2072. https://doi.org/10.3390/cancers13092072
APA StyleGonzález, G., Lakatos, K., Hoballah, J., Fritz-Klaus, R., Al-Johani, L., Brooker, J., Jeong, S., Evans, C. L., Krauledat, P., Cramer, D. W., Hoffman, R. A., Hansen, W. P., & Patankar, M. S. (2021). Characterization of Cell-Bound CA125 on Immune Cell Subtypes of Ovarian Cancer Patients Using a Novel Imaging Platform. Cancers, 13(9), 2072. https://doi.org/10.3390/cancers13092072