
High Antitumor Activity of the Dual Topoisomerase Inhibitor P8-D6 in Breast Cancer

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Preparation and Culture
2.2. Western Blot
2.3. Fluorescence Imaging
2.4. Two-Dimensional Viability and Apoptosis Assay
2.5. Three-Dimensional Cytotoxicity, Viability and Apoptosis Assay
2.6. Scanning Electron Microscopy (SEM)
2.7. Statistical Analysis
3. Results
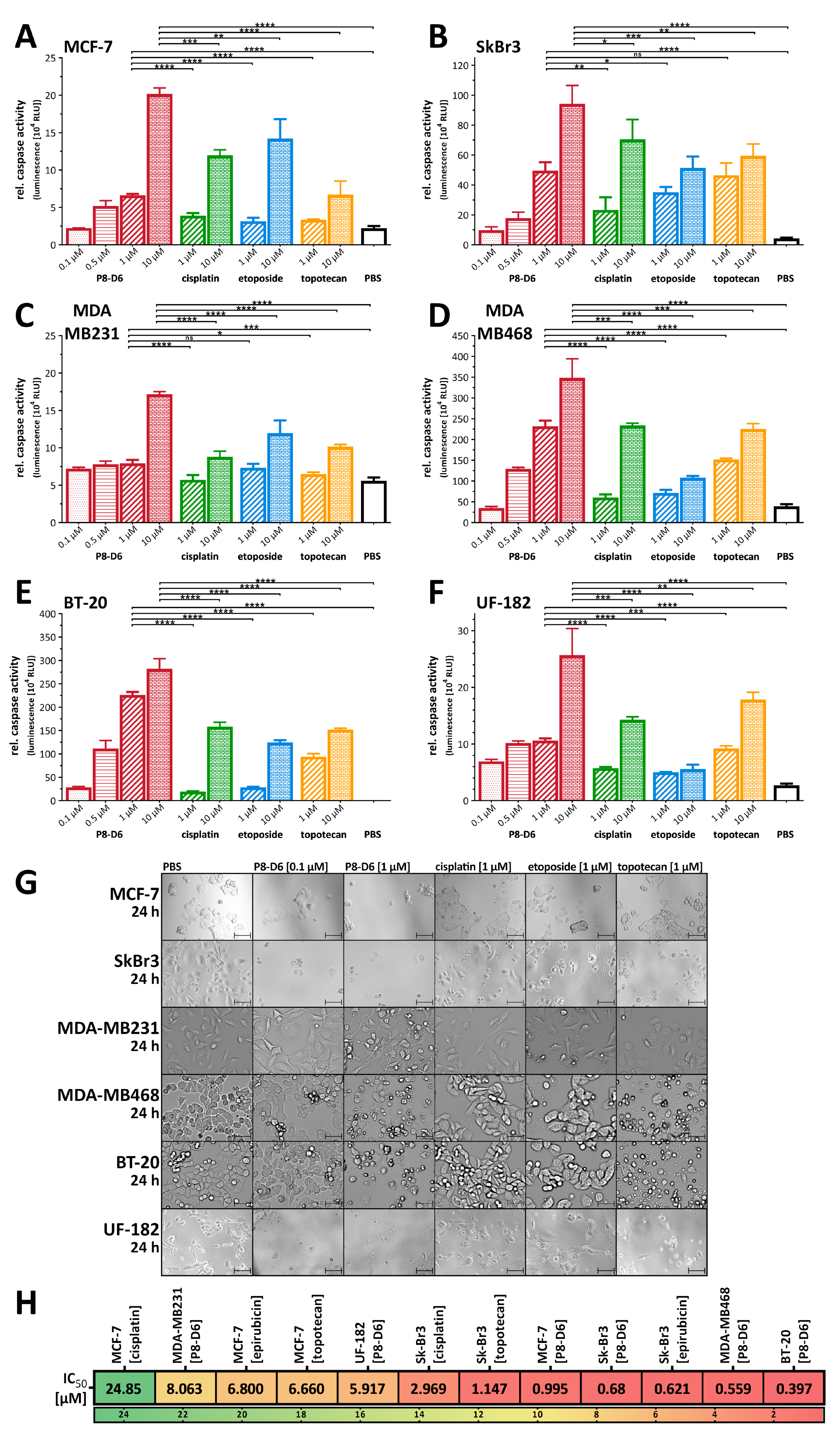
3.1. P8-D6 Is Highly Effective in BC 2D Monolayers
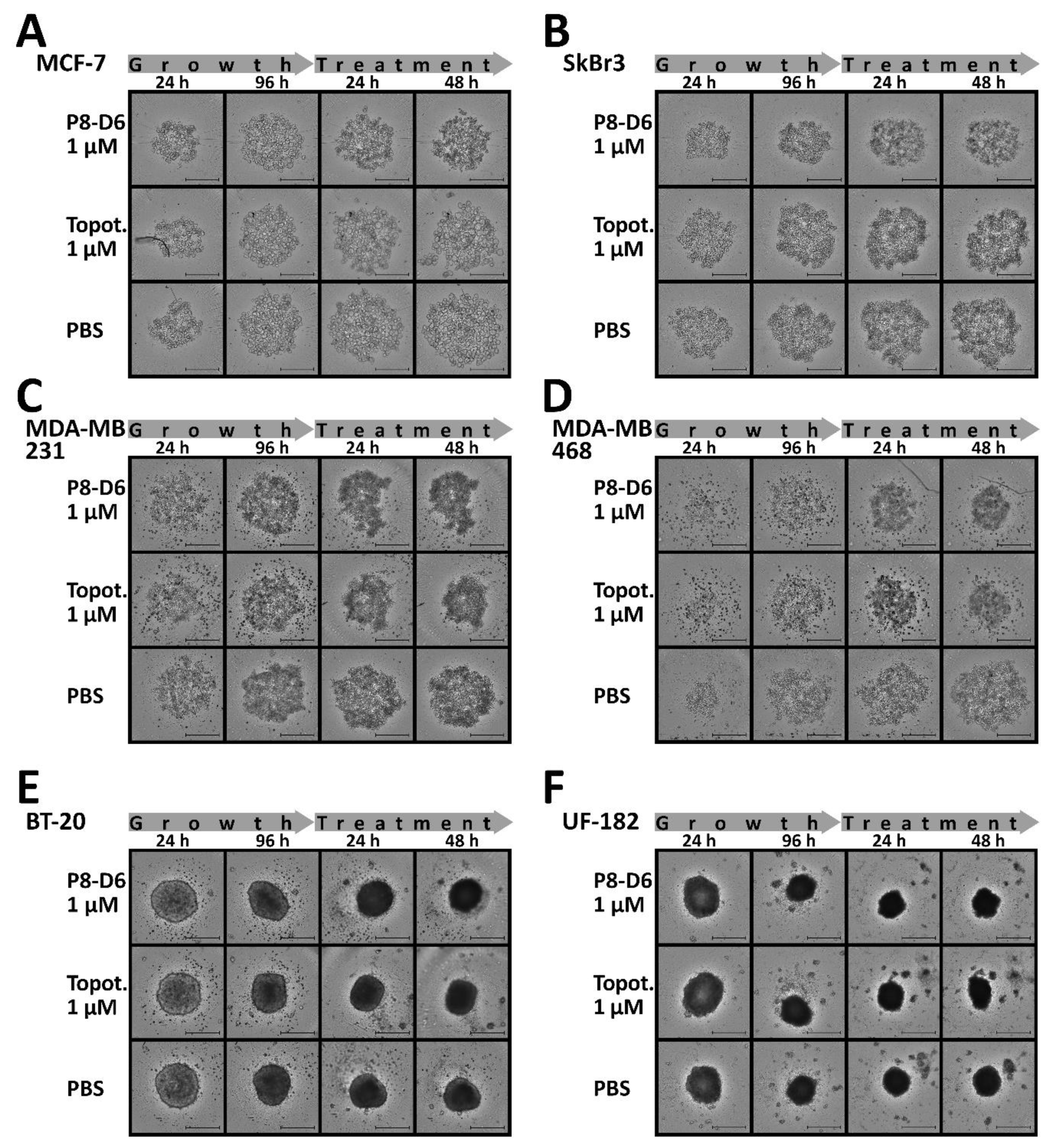
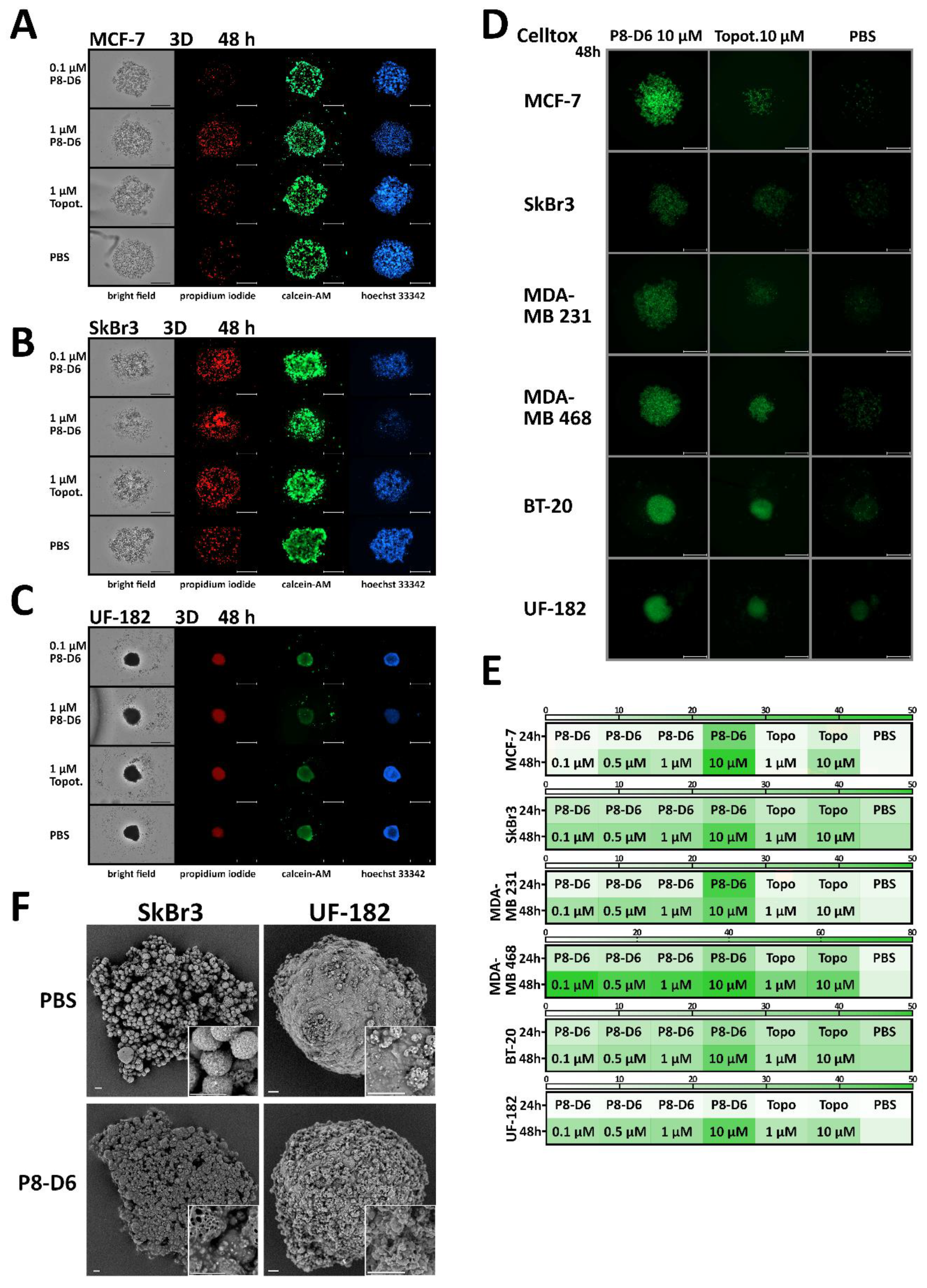
3.2. P8-D6 Induces Strong Effects in 3D Target Tumor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Ji, X.; Lu, Y.; Tian, H.; Meng, X.; Wei, M.; Cho, W.C. Chemoresistance mechanisms of breast cancer and their countermeasures. Biomed. Pharmacother. 2019, 114, 108800. [Google Scholar] [CrossRef]
- Meier, C.; Steinhauer, T.N.; Koczian, F.; Plitzko, B.; Jarolim, K.; Girreser, U.; Braig, S.; Marko, D.; Vollmar, A.M.; Clement, B. A Dual Topoisomerase Inhibitor of Intense Pro-Apoptotic and Antileukemic Nature for Cancer Treatment. ChemMedChem 2017, 12, 347–352. [Google Scholar] [CrossRef]
- Aichinger, G.; Lichtenberger, F.-B.; Steinhauer, T.N.; Flörkemeier, I.; Del Favero, G.; Clement, B.; Marko, D. The Aza-Analogous Benzo[c]phenanthridine P8-D6 Acts as a Dual Topoisomerase I and II Poison, thus Exhibiting Potent Genotoxic Properties. Molecules 2020, 25, 1524. [Google Scholar] [CrossRef] [Green Version]
- Pommier, Y.; Sun, Y.; Huang, S.-Y.N.; Nitiss, J.L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell. Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef]
- Deweese, J.E.; Osheroff, N. The DNA cleavage reaction of topoisomerase II: Wolf in sheep’s clothing. Nucleic Acids Res. 2009, 37, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Pommier, Y.; Barcelo, J.M.; Rao, V.A.; Sordet, O.; Jobson, A.G.; Thibaut, L.; Miao, Z.-H.; Seiler, J.A.; Zhang, H.; Marchand, C.; et al. Repair of topoisomerase I-mediated DNA damage. Prog. Nucleic Acid. Res. Mol. Biol. 2006, 81, 179–229. [Google Scholar] [CrossRef] [Green Version]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA topoisomerases and their poisoning by anticancer and antibacterial drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [Green Version]
- National Cancer Institute (NCI). DTP Homepage: Cancer Screen 09/2014: NCI-60 DTP Human Tumor Cell Line Screen. The National Cancer Institute (NCI) is Gratefully Acknowledged for Its Excellent Screening Service. Available online: http://dtp.nci.nih.gov (accessed on 1 November 2018).
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Lagies, S.; Schlimpert, M.; Neumann, S.; Wäldin, A.; Kammerer, B.; Borner, C.; Peintner, L. Cells grown in three-dimensional spheroids mirror in vivo metabolic response of epithelial cells. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kimlin, L.C.; Casagrande, G.; Virador, V.M. In vitro three-dimensional (3D) models in cancer research: An update. Mol. Carcinog. 2013, 52, 167–182. [Google Scholar] [CrossRef]
- Goodman, T.T.; Olive, P.L.; Pun, S.H. Increased nanoparticle penetration in collagenase-treated multicellular spheroids. Int. J. Nanomed. 2007, 2, 265–274. [Google Scholar]
- Kurbacher, C.M.; Korn, C.; Dexel, S.; Schween, U.; Kurbacher, J.A.; Reichelt, R.; Arenz, P.N. Isolation and Culture of Ovarian Cancer Cells and Cell Lines. Methods Mol. Biol. 2011, 161–180. [Google Scholar] [CrossRef]
- Huang, X.; Weimer, J.; von Wurmb-Schwark, N.; Fredrik, R.; Arnold, N.; Schem, C. Alteration of STR profiles in ovarian carcinoma cells during primary culture. Arch. Gynecol. Obstet. 2016, 294, 369–376. [Google Scholar] [CrossRef]
- Plitzko, B.; Havemeyer, A.; Kunze, T.; Clement, B. The pivotal role of the mitochondrial amidoxime reducing component 2 in protecting human cells against apoptotic effects of the base analog N6-hydroxylaminopurine. J. Biol. Chem. 2015, 290, 10126–10135. [Google Scholar] [CrossRef] [Green Version]
- Hedemann, N.; Herz, A.; Schiepanski, J.H.; Dittrich, J.; Sebens, S.; Dempfle, A.; Feuerborn, J.; Rogmans, C.; Tribian, N.; Flörkemeier, I.; et al. ADAM17 Inhibition Increases the Impact of Cisplatin Treatment in Ovarian Cancer Spheroids. Cancers 2021, 13, 2039. [Google Scholar] [CrossRef]
- Flörkemeier, I.; Steinhauer, T.N.; Hedemann, N.; Ölander, M.; Artursson, P.; Clement, B.; Bauerschlag, D.O. Newly developed dual topoisomerase inhibitor P8-D6 is highly active in ovarian cancer. Ther. Adv. Med. Oncol. 2021, 13, 175883592110598. [Google Scholar] [CrossRef] [PubMed]
- Jänicke, R.U. MCF-7 breast carcinoma cells do not express caspase-3. Breast Cancer Res. Treat. 2009, 117, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Wind, N.S.; Holen, I. Multidrug resistance in breast cancer: From in vitro models to clinical studies. Int. J. Breast Cancer 2011, 2011, 967419. [Google Scholar] [CrossRef]
- Aronson, J.K. (Ed.) Side Effects of Drugs Annual 27: Cytostatic Drugs, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 2004; ISBN 9780444513564. [Google Scholar]
- Shapiro, C.L.; Hardenbergh, P.H.; Gelman, R.; Blanks, D.; Hauptman, P.; Recht, A.; Hayes, D.F.; Harris, J.; Henderson, I.C. Cardiac effects of adjuvant doxorubicin and radiation therapy in breast cancer patients. J. Clin. Oncol. 1998, 16, 3493–3501. [Google Scholar] [CrossRef]
- Lankelma, J.; Dekker, H.; Luque, F.R.; Luykx, S.; Hoekman, K.; van der Valk, P.; van Diest, P.J.; Pinedo, H.M. Doxorubicin gradients in human breast cancer. Clin. Cancer Res. 1999, 5, 1703–1707. [Google Scholar]
- Franco, Y.L.; Vaidya, T.R.; Ait-Oudhia, S. Anticancer and cardio-protective effects of liposomal doxorubicin in the treatment of breast cancer. Breast Cancer 2018, 10, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Ansari, L.; Shiehzadeh, F.; Taherzadeh, Z.; Nikoofal-Sahlabadi, S.; Momtazi-borojeni, A.A.; Sahebkar, A.; Eslami, S. The most prevalent side effects of pegylated liposomal doxorubicin monotherapy in women with metastatic breast cancer: A systematic review of clinical trials. Cancer Gene Ther. 2017, 24, 189–193. [Google Scholar] [CrossRef]
- Van Gijn, R.; Lendfers, R.R.H.; Schellens, J.H.M.; Bult, A.; Beijnen, J.H. Dual topoisomerase I/II inhibitors. J. Oncol. Pharm. Pract. 2000, 6, 92–108. [Google Scholar] [CrossRef]
- Robert, J.; Rivory, L. Pharmacology of irinotecan. Drugs Today 1998, 34, 777–803. [Google Scholar] [CrossRef]
- Sachdev, J.; Munster, P.; Northfelt, D.; Han, H.S.; MA, C.; Maxwell, F.; Wang, T.; Belanger, B.; Zhang, B.; Moore, Y.; et al. Abstract CT048: Phase I expansion study of irinotecan liposome injection (nal-IRI) in patients with metastatic breast cancer (mBC). Clin. Trials 2019, 79, CT048. [Google Scholar] [CrossRef]
- Park, I.H.; Im, S.-A.; Jung, K.H.; Sohn, J.H.; Park, Y.H.; Lee, K.S.; Sim, S.H.; Park, K.-H.; Kim, J.H.; Nam, B.H.; et al. Randomized Open Label Phase III Trial of Irinotecan Plus Capecitabine versus Capecitabine Monotherapy in Patients with Metastatic Breast Cancer Previously Treated with Anthracycline and Taxane: PROCEED Trial (KCSG BR 11-01). Cancer Res. Treat. 2019, 51, 43–52. [Google Scholar] [CrossRef]
- Segar, J.M.; Reed, D.; Stopeck, A.; Livingston, R.B.; Chalasani, P. A Phase II Study of Irinotecan and Etoposide as Treatment for Refractory Metastatic Breast Cancer. Oncologist 2019, 24, 1512. [Google Scholar] [CrossRef] [Green Version]
- Ewesuedo, R.B.; Iyer, L.; Das, S.; Koenig, A.; Mani, S.; Vogelzang, N.J.; Schilsky, R.L.; Brenckman, W.; Ratain, M.J. Phase I clinical and pharmacogenetic study of weekly TAS-103 in patients with advanced cancer. J. Clin. Oncol. 2001, 19, 2084–2090. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flörkemeier, I.; Steinhauer, T.N.; Hedemann, N.; Weimer, J.P.; Rogmans, C.; van Mackelenbergh, M.T.; Maass, N.; Clement, B.; Bauerschlag, D.O. High Antitumor Activity of the Dual Topoisomerase Inhibitor P8-D6 in Breast Cancer. Cancers 2022, 14, 2. https://doi.org/10.3390/cancers14010002
Flörkemeier I, Steinhauer TN, Hedemann N, Weimer JP, Rogmans C, van Mackelenbergh MT, Maass N, Clement B, Bauerschlag DO. High Antitumor Activity of the Dual Topoisomerase Inhibitor P8-D6 in Breast Cancer. Cancers. 2022; 14(1):2. https://doi.org/10.3390/cancers14010002
Chicago/Turabian StyleFlörkemeier, Inken, Tamara N. Steinhauer, Nina Hedemann, Jörg Paul Weimer, Christoph Rogmans, Marion T. van Mackelenbergh, Nicolai Maass, Bernd Clement, and Dirk O. Bauerschlag. 2022. "High Antitumor Activity of the Dual Topoisomerase Inhibitor P8-D6 in Breast Cancer" Cancers 14, no. 1: 2. https://doi.org/10.3390/cancers14010002
APA StyleFlörkemeier, I., Steinhauer, T. N., Hedemann, N., Weimer, J. P., Rogmans, C., van Mackelenbergh, M. T., Maass, N., Clement, B., & Bauerschlag, D. O. (2022). High Antitumor Activity of the Dual Topoisomerase Inhibitor P8-D6 in Breast Cancer. Cancers, 14(1), 2. https://doi.org/10.3390/cancers14010002