
SGK1 in Cancer: Biomarker and Drug Target

 ,
, 
Abstract
:1. Introduction
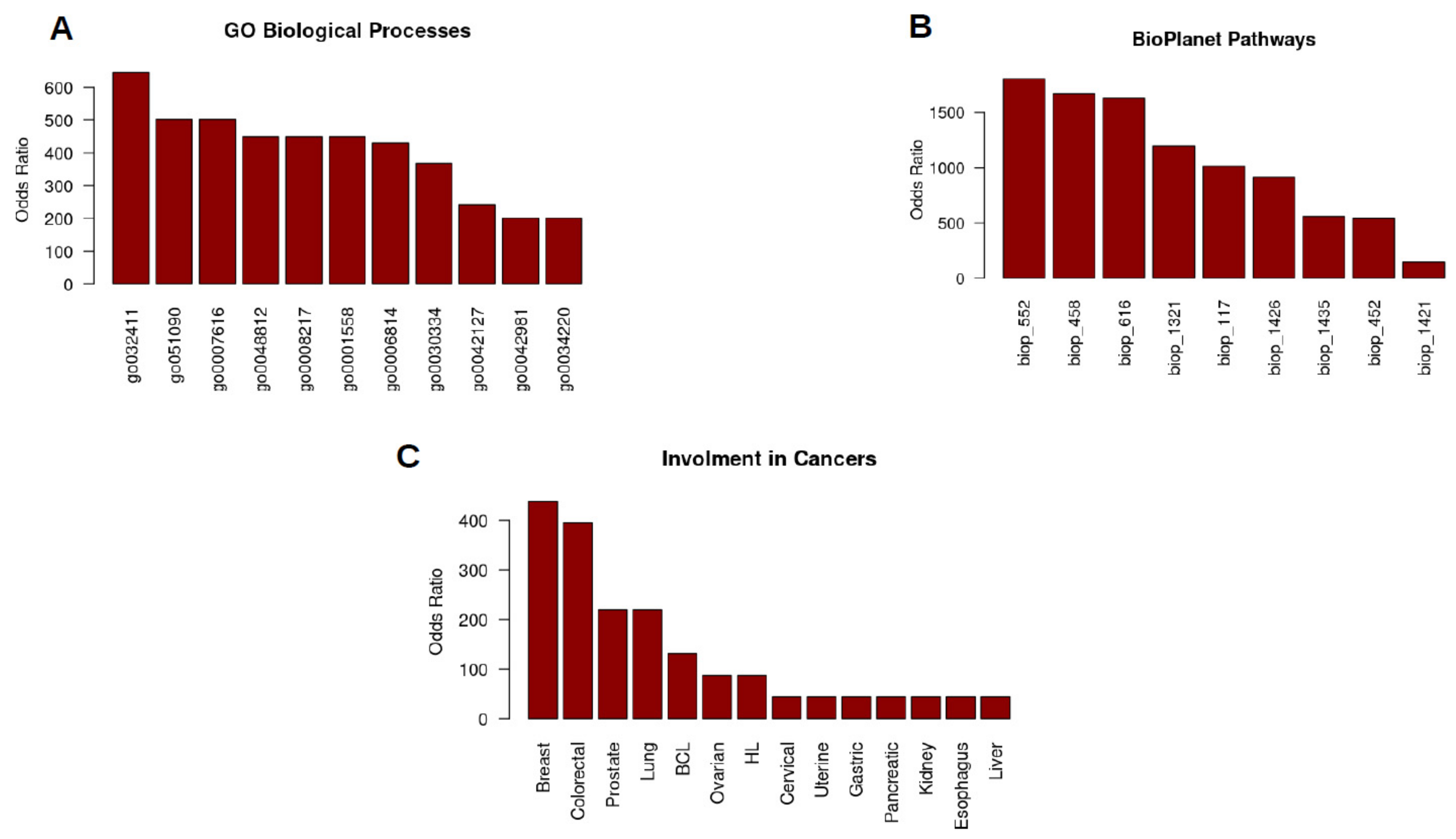
2. SGK1 as Biomarker
3. SGK1 as Drug Target
4. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- Firestone, G.L.; Giampaolo, J.R.; O’Keeffe, B.A. Stimulus-dependent regulation of serum and glucocorticoid inducible protein kinase (SGK) transcription, subcellular localization and enzymatic activity. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2003, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Waldegger, S.; Barth, P.; Raber, G.; Lang, F. Cloning and characterization of a putative human serine/threonine protein kinase transcriptionally modified during anisotonic and isotonic alterations of cell volume. Proc. Natl. Acad. Sci. USA 1997, 94, 4440–4445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, F.; Gorlach, A.; Vallon, V. Targeting SGK1 in diabetes. Expert Opin. Ther. Targets 2009, 13, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Notch, E.G.; Chapline, C.; Flynn, E.; Lameyer, T.; Lowell, A.; Sato, D.; Shaw, J.R.; Stanton, B.A. Mitogen activated protein kinase 14-1 regulates serum glucocorticoid kinase 1 during seawater acclimation in Atlantic killifish, Fundulus heteroclitus. Comparative biochemistry and physiology. Part A Mol. Integr. Physiol. 2012, 162, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talarico, C.; Dattilo, V.; D’Antona, L.; Barone, A.; Amodio, N.; Belviso, S.; Musumeci, F.; Abbruzzese, C.; Bianco, C.; Trapasso, F.; et al. SI113, a SGK1 inhibitor, potentiates the effects of radiotherapy, modulates the response to oxidative stress and induces cytotoxic autophagy in human glioblastoma multiforme cells. Oncotarget 2016, 7, 15868–15884. [Google Scholar] [CrossRef] [Green Version]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef] [Green Version]
- Lang, F.; Shumilina, E. Regulation of ion channels by the serum- and glucocorticoid-inducible kinase SGK1. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 3–12. [Google Scholar] [CrossRef]
- Faletti, C.J.; Perrotti, N.; Taylor, S.I.; Blazer-Yost, B.L. sgk: An essential convergence point for peptide and steroid hormone regulation of ENaC-mediated Na+ transport. Am. J. Physiol.-Cell Physiol. 2002, 282, C494–C500. [Google Scholar] [CrossRef] [Green Version]
- Amato, R.; D’Antona, L.; Porciatti, G.; Agosti, V.; Menniti, M.; Rinaldo, C.; Costa, N.; Bellacchio, E.; Mattarocci, S.; Fuiano, G.; et al. Sgk1 activates MDM2-dependent p53 degradation and affects cell proliferation, survival, and differentiation. J. Mol. Med. 2009, 87, 1221–1239. [Google Scholar] [CrossRef]
- Lyo, D.; Xu, L.; Foster, D.A. Phospholipase D stabilizes HDM2 through an mTORC2/SGK1 pathway. Biochem. Biophys. Res. Commun. 2010, 396, 562–565. [Google Scholar] [CrossRef] [Green Version]
- Amato, R.; Scumaci, D.; D’Antona, L.; Iuliano, R.; Menniti, M.; Di Sanzo, M.; Faniello, M.C.; Colao, E.; Malatesta, P.; Zingone, A.; et al. Sgk1 enhances RANBP1 transcript levels and decreases taxol sensitivity in RKO colon carcinoma cells. Oncogene 2013, 32, 4572–4578. [Google Scholar] [CrossRef] [Green Version]
- Dattilo, V.; D’Antona, L.; Talarico, C.; Capula, M.; Catalogna, G.; Iuliano, R.; Schenone, S.; Roperto, S.; Bianco, C.; Perrotti, N.; et al. SGK1 affects RAN/RANBP1/RANGAP1 via SP1 to play a critical role in pre-miRNA nuclear export: A new route of epigenomic regulation. Sci. Rep. 2017, 7, 45361. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.M.; Gu, S.; Anagnostopoulou, V.; Alevizopoulos, K.; Föller, M.; Lang, F.; Stournaras, C. Serum- and glucocorticoid-dependent kinase-1-induced cell migration is dependent on vinculin and regulated by the membrane androgen receptor. FEBS J. 2012, 279, 1231–1242. [Google Scholar] [CrossRef]
- Chen, L.; Wei, T.Q.; Wang, Y.; Zhang, J.; Li, H.; Wang, K.J. Simulated bladder pressure stimulates human bladder smooth muscle cell proliferation via the PI3K/SGK1 signaling pathway. J. Urol. 2012, 188, 661–667. [Google Scholar] [CrossRef]
- Schmid, E.; Gu, S.; Yang, W.; Münzer, P.; Schaller, M.; Lang, F.; Stournaras, C.; Shumilina, E. Serum- and glucocorticoid-inducible kinase SGK1 regulates reorganization of actin cytoskeleton in mast cells upon degranulation. Am. J. Physiol.-Cell Physiol. 2012, 304, C49–C55. [Google Scholar] [CrossRef] [Green Version]
- Rotte, A.; Mack, A.F.; Bhandaru, M.; Kempe, D.S.; Beier, N.; Scholz, W.; Dicks, E.; Potzsch, S.; Kuhl, D.; Lang, F. Pioglitazone induced gastric acid secretion. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2009, 24, 193–200. [Google Scholar] [CrossRef]
- Lang, F.; Bohmer, C.; Palmada, M.; Seebohm, G.; Strutz-Seebohm, N.; Vallon, V. (Patho) physiological significance of the serum- and glucocorticoid-inducible kinase isoforms. Physiol. Rev. 2006, 86, 1151–1178. [Google Scholar] [CrossRef]
- Andres-Mateos, E.; Brinkmeier, H.; Burks, T.N.; Mejias, R.; Files, D.C.; Steinberger, M.; Soleimani, A.; Marx, R.; Simmers, J.L.; Lin, B.; et al. Activation of serum/glucocorticoid-induced kinase 1 (SGK1) is important to maintain skeletal muscle homeostasis and prevent atrophy. EMBO Mol. Med. 2013, 5, 80–91. [Google Scholar] [CrossRef]
- Lang, F.; Vallon, V. Serum- and glucocorticoid-inducible kinase 1 in the regulation of renal and extrarenal potassium transport. Clin. Exp. Nephrol. 2012, 16, 73–80. [Google Scholar] [CrossRef]
- Faresse, N.; Lagnaz, D.; Debonneville, A.; Ismailji, A.; Maillard, M.; Fejes-Toth, G.; Náray-Fejes-Tóth, A.; Staub, O. Inducible kidney-specific Sgk1 knockout mice show a salt-losing phenotype. Am. J. Physiol.-Ren. Physiol. 2012, 302, F977–F985. [Google Scholar] [CrossRef]
- Ghani, M.J. SGK1, autophagy and cancer: An overview. Mol. Biol. Rep. 2022, 49, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, Z.; Xiao, S.; Thalhamer, T.; Madi, A.; Han, T.; Kuchroo, V. SGK1 Governs the Reciprocal Development of Th17 and Regulatory T Cells. Cell Rep. 2018, 22, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norlander, A.E.; Saleh, M.A.; Pandey, A.K.; Itani, H.A.; Wu, J.; Xiao, L.; Kang, J.; Dale, B.L.; Goleva, S.B.; Laroumanie, F.; et al. A salt-sensing kinase in T lymphocytes, SGK1, drives hypertension and hypertensive end-organ damage. JCI Insight 2017, 2, e92801. [Google Scholar] [CrossRef] [PubMed]
- Kleinewietfeld, M.; Manzel, A.; Titze, J.; Kvakan, H.; Yosef, N.; Linker, R.A.; Muller, D.N.; Hafler, D.A. Sodium chloride drives autoimmune disease by the induction of pathogenic TH17 cells. Nature 2013, 496, 518–522. [Google Scholar] [CrossRef]
- Spagnuolo, R.; Dattilo, V.; D’Antona, L.; Cosco, C.; Tallerico, R.; Ventura, V.; Conforti, F.; Camastra, C.; Mancina, R.M.; Catalogna, G.; et al. Deregulation of SGK1 in Ulcerative Colitis: A Paradoxical Relationship Between Immune Cells and Colonic Epithelial Cells. Inflamm. Bowel Dis. 2018, 24, 1967–1977. [Google Scholar] [CrossRef]
- Wang, H.R.; Chen, D.L.; Zhao, M.; Shu, S.W.; Xiong, S.X.; Gan, X.D.; Chao, S.P.; Cao, J.L. C-reactive protein induces interleukin-6 and thrombospondin-1 protein and mRNA expression through activation of nuclear factor-ĸB in HK-2 cells. Kidney Blood Press. Res. 2012, 35, 211–219. [Google Scholar] [CrossRef]
- Dahlberg, J.; Smith, G.; Norrving, B.; Nilsson, P.; Hedblad, B.; Engström, G.; Lövkvist, H.; Carlson, J.; Lindgren, A.; Melander, O. Genetic variants in serum and glucocortocoid regulated kinase 1, a regulator of the epithelial sodium channel, are associated with ischaemic stroke. J. Hypertens. 2011, 29, 884–889. [Google Scholar] [CrossRef]
- Borst, O.; Schmidt, E.M.; Münzer, P.; Schönberger, T.; Towhid, S.T.; Elvers, M.; Leibrock, C.; Schmid, E.; Eylenstein, A.; Kuhl, D.; et al. The serum- and glucocorticoid-inducible kinase 1 (SGK1) influences platelet calcium signaling and function by regulation of Orai1 expression in megakaryocytes. Blood 2012, 119, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Schwab, M.; Lupescu, A.; Mota, M.; Mota, E.; Frey, A.; Simon, P.; Mertens, P.R.; Floege, J.; Luft, F.; Asante-Poku, S.; et al. Association of SGK1 gene polymorphisms with type 2 diabetes. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2008, 21, 151–160. [Google Scholar] [CrossRef]
- Cicenas, J.; Zalyte, E.; Rimkus, A.; Dapkus, D.; Noreika, R.; Urbonavicius, S. JNK, p38, ERK, and SGK1 Inhibitors in Cancer. Cancers 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Kong, P.; Zhang, S.; Zhang, L.; Cao, Y.; Duan, X.; Sun, T.; Tao, Z.; Liu, W. SGK1 in Human Cancer: Emerging Roles and Mechanisms. Front. Oncol. 2021, 19, 608722. [Google Scholar] [CrossRef]
- Lang, F.; Perrotti, N.; Stournaras, C. Colorectal carcinoma cells—Regulation of survival and growth by SGK1. Int. J. Biochem. Cell Biol. 2010, 42, 1571–1575. [Google Scholar] [CrossRef]
- Szmulewitz, R.Z.; Chung, E.; Al-Ahmadie, H.; Daniel, S.; Kocherginsky, M.; Razmaria, A.; Zagaja, G.P.; Brendler, C.B.; Stadler, W.M.; Conzen, S.D. Serum/glucocorticoid-regulated kinase 1 expression in primary human prostate cancers. Prostate 2012, 72, 157–164. [Google Scholar] [CrossRef]
- Melhem, A.; Yamada, S.D.; Fleming, G.F.; Delgado, B.; Brickley, D.R.; Wu, W.; Kocherginsky, M.; Conzen, S.D. Administration of glucocorticoids to ovarian cancer patients is associated with expression of the anti-apoptotic genes SGK1 and MKP1/DUSP1 in ovarian tissues. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2009, 15, 3196–3204. [Google Scholar] [CrossRef] [Green Version]
- Abbruzzese, C.; Mattarocci, S.; Pizzuti, L.; Mileo, A.M.; Visca, P.; Antoniani, B.; Alessandrini, G.; Facciolo, F.; Amato, R.; D’Antona, L.; et al. Determination of SGK1 mRNA in non-small cell lung cancer samples underlines high expression in squamous cell carcinomas. J. Exp. Clin. Cancer Res. CR 2012, 31, 4. [Google Scholar] [CrossRef] [Green Version]
- Chung, E.J.; Sung, Y.K.; Farooq, M.; Kim, Y.; Im, S.; Tak, W.Y.; Hwang, Y.J.; Kim, Y.I.; Han, H.S.; Kim, J.C.; et al. Gene expression profile analysis in human hepatocellular carcinoma by cDNA microarray. Mol. Cells 2002, 14, 382–387. [Google Scholar]
- Fagerli, U.M.; Ullrich, K.; Stühmer, T.; Holien, T.; Köchert, K.; Holt, R.U.; Bruland, O.; Chatterjee, M.; Nogai, H.; Lenz, G.; et al. Serum/glucocorticoid-regulated kinase 1 (SGK1) is a prominent target gene of the transcriptional response to cytokines in multiple myeloma and supports the growth of myeloma cells. Oncogene 2011, 30, 3198–3206. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.W.; Gilbertson, R.; Iannaccone, S.; Iannaccone, P.; Walterhouse, D. Defining a role for Sonic hedgehog pathway activation in desmoplastic medulloblastoma by identifying GLI1 target genes. Int. J. Cancer 2009, 124, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Rauhala, H.E.; Porkka, K.P.; Tolonen, T.T.; Martikainen, P.M.; Tammela, T.L.; Visakorpi, T. Dual-specificity phosphatase 1 and serum/glucocorticoid-regulated kinase are downregulated in prostate cancer. Int. J. Cancer 2005, 117, 738–745. [Google Scholar] [CrossRef]
- Chu, S.; Rushdi, S.; Zumpe, E.T.; Mamers, P.; Healy, D.L.; Jobling, T.; Burger, H.G.; Fuller, P.J. FSH-regulated gene expression profiles in ovarian tumours and normal ovaries. Mol. Hum. Reprod. 2002, 8, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Segditsas, S.; Sieber, O.; Deheragoda, M.; East, P.; Rowan, A.; Jeffery, R.; Nye, E.; Clark, S.; Spencer-Dene, B.; Stamp, G.; et al. Putative direct and indirect Wnt targets identified through consistent gene expression changes in APC-mutant intestinal adenomas from humans and mice. Hum. Mol. Genet. 2008, 17, 3864–3875. [Google Scholar] [CrossRef] [Green Version]
- Ger, M.; Kaupinis, A.; Petrulionis, M.; Kurlinkus, B.; Cicenas, J.; Sileikis, A.; Valius, M.; Strupas, K. Proteomic Identification of FLT3 and PCBP3 as Potential Prognostic Biomarkers for Pancreatic Cancer. Anticancer Res. 2018, 38, 759–5765. [Google Scholar] [CrossRef]
- Ronchi, C.L.; Leich, E.; Sbiera, S.; Weismann, D.; Rosenwald, A.; Allolio, B.; Fassnacht, M. Single nucleotide polymorphism microarray analysis in cortisol-secreting adrenocortical adenomas identifies new candidate genes and pathways. Neoplasia 2012, 14, 206–218. [Google Scholar] [CrossRef] [Green Version]
- Ronchi, C.L.; Sbiera, S.; Leich, E.; Tissier, F.; Steinhauer, S.; Deutschbein, T.; Fassnacht, M.; Allolio, B. Low SGK1 expression in human adrenocortical tumors is associated with ACTH-independent glucocorticoid secretion and poor prognosis. J. Clin. Endocrinol. Metab. 2012, 97, E2251–E2260. [Google Scholar] [CrossRef]
- Sommer, E.M.; Dry, H.; Cross, D.; Guichard, S.; Davies, B.R.; Alessi, D.R. Elevated SGK1 predicts resistance of breast cancer cells to Akt inhibitors. Biochem. J. 2013, 452, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Shen, Q.; Xie, H.; Zhou, Z.; Shi, G.; Zhang, C.; Mohammed, A.; Wu, Y.; Ni, S.; Zhou, X. Serum and glucocorticoid-regulated kinase 1 (SGK1) is a predictor of poor prognosis in non-small cell lung cancer, and its dynamic pattern following treatment with SGK1 inhibitor and γ-ray irradiation was elucidated. Oncol. Rep. 2018, 39, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
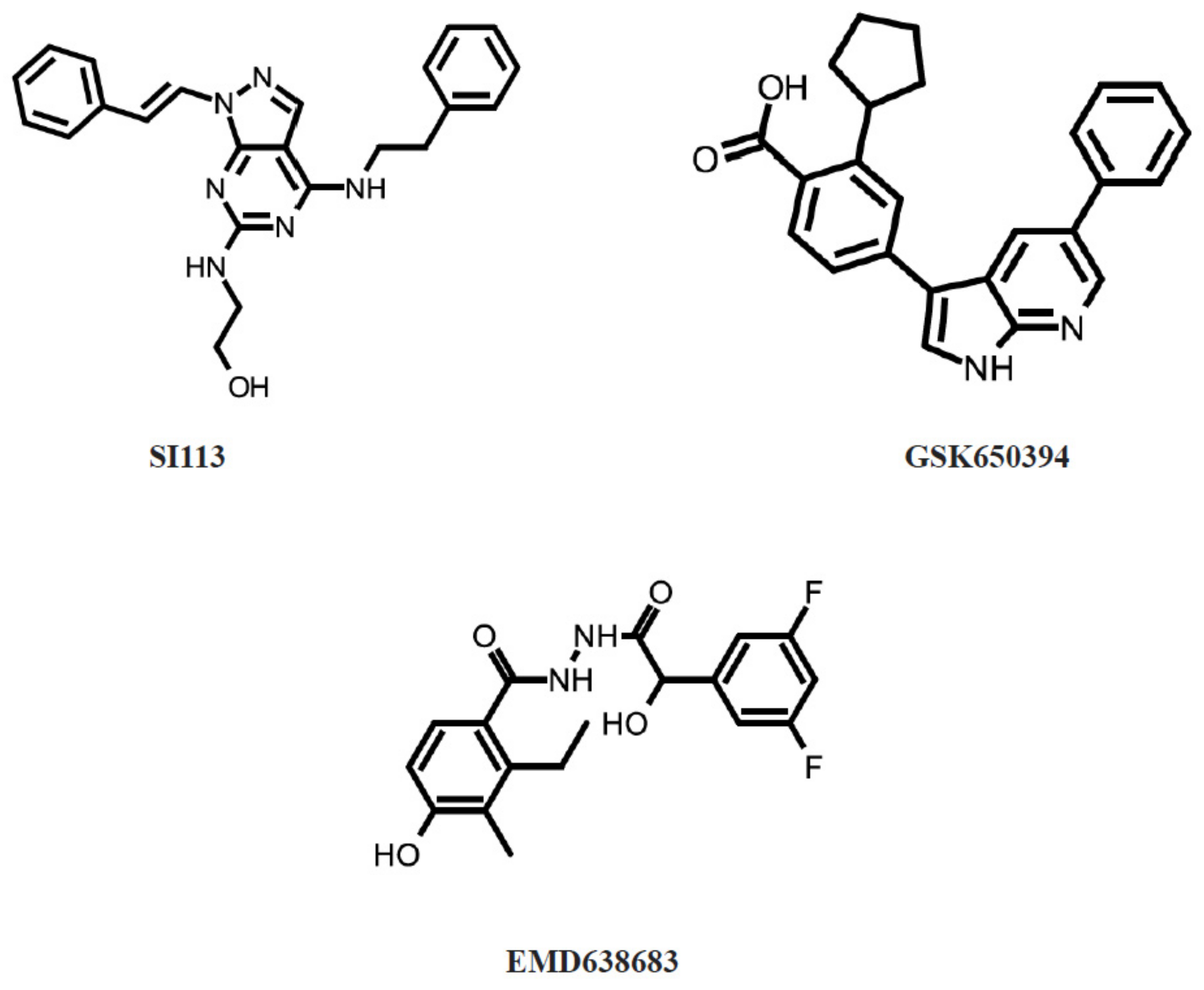
- D’Antona, L.; Amato, R.; Talarico, C.; Ortuso, F.; Menniti, M.; Dattilo, V.; Iuliano, R.; Gigliotti, F.; Artese, A.; Costa, G.; et al. SI113, a specific inhibitor of the Sgk1 kinase activity that counteracts cancer cell proliferation. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 35, 2006–2018. [Google Scholar] [CrossRef]
- Talarico, C.; D’Antona, L.; Scumaci, D.; Barone, A.; Gigliotti, F.; Fiumara, C.V.; Dattilo, V.; Gallo, E.; Visca, P.; Ortuso, F.; et al. Preclinical model in HCC: The SGK1 kinase inhibitor SI113 blocks tumor progression in vitro and in vivo and synergizes with radiotherapy. Oncotarget 2015, 6, 37511–37525. [Google Scholar] [CrossRef] [Green Version]
- Conza, D.; Mirra, P.; Calì, G.; Tortora, T.; Insabato, L.; Fiory, F.; Schenone, S.; Amato, R.; Beguinot, F.; Perrotti, N.; et al. The SGK1 inhibitor SI113 induces autophagy, apoptosis, and endoplasmic reticulum stress in endometrial cancer cells. J. Cell. Physiol. 2017, 232, 3735–3743. [Google Scholar] [CrossRef]
- Matschke, J.; Wiebeck, E.; Hurst, S.; Rudner, J.; Jendrossek, V. Role of SGK1 for fatty acid uptake, cell survival and radioresistance of NCI-H460 lung cancer cells exposed to acute or chronic cycling severe hypoxia. Radiat. Oncol. 2016, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Lan, C.; Zhou, J.; Fu, W.; Long, X.; An, Y.; Jiao, G.; Wang, K.; Li, Y.; Xu, J.; et al. Development of a new analog of SGK1 inhibitor and its evaluation as a therapeutic molecule of colorectal cancer. J. Cancer 2017, 8, 2256–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, X.; Wang, Y.; Dai, Y.; Xie, Y.; Ping, Y.; Yin, B.; Yu, P.; Liu, Z.; Duan, X.; et al. SGK1 inhibition-induced autophagy impairs prostate cancer metastasis by reversing EMT. J. Exp. Clin. Cancer Res. CR 2018, 37, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Lan, C.; Jiao, G.; Fu, W.; Long, X.; An, Y.; Wang, K.; Zhou, J.; Chen, T.; Li, Y.; et al. Therapeutic inhibition of SGK1 suppresses colorectal cancer. Exp. Mol. Med. 2017, 24, 11.e399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Xue, Y.; Shen, L.; Qin, P.; Sang, X.; Tao, Z.; Yi, J.; Wang, J.; Liu, P.; Cheng, H. Inhibition of SGK1 confers vulnerability to redox dysregulation in cervical cancer. Redox Biol. 2019, 24, 101225. [Google Scholar] [CrossRef]
- Zhou, C.; Xiao, W.; Jiang, T.; Guo, Z.; Li, M.; Chang, H.; Wu, Y.; Chen, M.; Shi, M.; Xu, W.; et al. Targeting SGK1 enhances the efficacy of radiotherapy in locally advanced rectal cancer. Biomed. Pharmacother. 2020, 125, 109954. [Google Scholar] [CrossRef]
- Towhid, S.T.; Liu, G.L.; Ackermann, T.F.; Beier, N.; Scholz, W.; Fuchß, T.; Toulany, M.; Rodemann, H.P.; Lang, F. Inhibition of Colonic Tumor Growth by the Selective SGK Inhibitor EMD638683. Cell. Physiol. Biochem. 2013, 32, 838–848. [Google Scholar] [CrossRef]
- Liu, G.; Honisch, S.; Liu, G.; Schmidt, S.; Pantelakos, S.; Alkahtani, S.; Toulany, M.; Lang, F.; Stournaras, C. Inhibition of SGK1 enhances mAR-induced apoptosis in MCF-7 breast cancer cells. Cancer Biol. Ther. 2015, 16, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Schmid, E.; Stagno, M.J.; Yan, J.; Schleicher, S.; Yu, W.; Honisch, S.; Lang, F.; Fuchs, J.; Seitz, G. Serum and Glucocorticoid Inducible Kinase 1-Sensitive Survival, Proliferation and Migration of Rhabdomyosarcoma Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 1301–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Antona, L.; Dattilo, V.; Catalogna, G.; Scumaci, D.; Fiumara, C.V.; Musumeci, F.; Perrotti, G.; Schenone, S.; Tallerico, R.; Spoleti, C.B.; et al. In Preclinical Model of Ovarian Cancer, the SGK1 Inhibitor SI113 Counteracts the Development of Paclitaxel Resistance and Restores Drug Sensitivity. Transl. Oncol. 2019, 12, 1045–1055. [Google Scholar] [CrossRef]
- Catalogna, G.; Talarico, C.; Dattilo, V.; Gangemi, V.; Calabria, F.; D’Antona, L.; Schenone, S.; Musumeci, F.; Bianco, C.; Perrotti, N.; et al. The SGK1 Kinase Inhibitor SI113 Sensitizes Theranostic Effects of the 64CuCl2 in Human Glioblastoma Multiforme Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Li, N.; Xue, Z.; Liu, L.R.; Li, J.; Huang, X.; Xie, X.; Zou, Y.; Tang, H.; Xie, X. Synergistic therapeutic effect of combined PDGFR and SGK1 inhibition in metastasis-initiating cells of breast cancer. Cell Death Differ. 2020, 27, 2066–2080. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, J.; Wu, Y.; Gao, Q.; Zhang, S. Potential Biomarkers for Liver Cancer Diagnosis Based on Multi-Omics Strategy. Front. Oncol. 2022, 12, 822449. [Google Scholar] [CrossRef] [PubMed]
- Cicenas, J.; Račienė, A. Anti-Cancer Drugs Targeting Protein Kinases Approved by FDA in 2020. Cancers 2021, 13, 947. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Experiment Performed | Significance | Reference |
|---|---|---|---|
| Adrenocortical adenoma | single nucleotide polymorphism microarrays, FISH, DNA qRT-PCR | 31 gains and 15 losses of gene detected | [43] |
| Adrenocortical tumor | immunohistochemistry, tissue microarrays. | Survival was shorter in patients with low SGK1 protein expression (HR = 2; 95% CI = 1.24–3.24; p = 0.0048). | [44] |
| Breast cancer | qRT-PCR | SGK1 mRNA is higher in AKT-inhibitor-resistant cells | [45] |
| Non-small cell lung cancer | tissue microarray immunohistochemistry, GSK650394 treatment | high expression of SGK1 (HR = 1.726; 95% CI = 1.396–2.865; p < 0.001) is an independent prognostic factor for the five year survival. GSK650394 induced apoptosis and p53 pathway. | [46] |
| Colon carcinoma | SI113 treatment | Cell cycle progression delay compared with untreated cells (p = 0.0024). | [47] |
| Hepatocellular carcinoma | SI113 treatment | Smaller tumor volume than control mice (p = 0.0009) | [48] |
| Glioblastoma multiforme | SI113 treatment | in cell lines, significant increase in caspase-mediated apoptosis were detected (p < 0.05). | [5] |
| Endometrial cancer | SI113 treatment | Apoptosis and autophagy were induced by inhibitor. | [49] |
| Lung adenocarcinoma | GSK650394 treatment | Decreased long-term survival and sensitized to the cytotoxic effects of ionizing radiation (p ≤ 0.05) | [50] |
| Colon cancer | GSK650394 treatment | inhibitor also obstructed colonic tumor growth and HCT116 cell proliferation in vivo (p < 0.05). | [51] |
| Prostate cancer | GSK650394 treatment | Induced autophagy, which led to the inhibition of cell metastasis (p < 0.05). | [52] |
| Colorectal cancer | GSK650394 treatment | Repressed tumor cell proliferation (p < 0.05) and tumor growth (p < 0.001). | [53] |
| Cervical cancer | GSK650394 treatment | GSK650394 in combination with melatonin, caused significant tumor size decrease in all cases and even complete tumor remission in 33% of mice (p ≤ 0.001). | [54] |
| Colorectal cancer | GSK650394 treatment | Combination of inhibitor with radiotherapy resulted in minimal tumor size compared with radiotherapy or inhibitor alone (p < 0.05) or control groups (p < 0.01). | [55] |
| Colon cancer | EMD638683 treatment | Number of developing tumors was significantly blunted by EMD638683 treatment. | [56] |
| Breast cancer | EMD638683 treatment | GSK650394 and testosterone albumin conjugate induced strong apoptotic response and caspase 3 activation (p < 0.01), enhanced radiation-induced cell growth control (p < 0.001). | [57] |
| Rhabdomyosarcoma | EMD638683 treatment | Statistically significant decline of cell viability and migration (p < 0.05). | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicenas, J.; Meskinyte-Kausiliene, E.; Jukna, V.; Rimkus, A.; Simkus, J.; Soderholm, D. SGK1 in Cancer: Biomarker and Drug Target. Cancers 2022, 14, 2385. https://doi.org/10.3390/cancers14102385
Cicenas J, Meskinyte-Kausiliene E, Jukna V, Rimkus A, Simkus J, Soderholm D. SGK1 in Cancer: Biomarker and Drug Target. Cancers. 2022; 14(10):2385. https://doi.org/10.3390/cancers14102385
Chicago/Turabian StyleCicenas, Jonas, Edita Meskinyte-Kausiliene, Vigilijus Jukna, Arnas Rimkus, Jokubas Simkus, and Diana Soderholm. 2022. "SGK1 in Cancer: Biomarker and Drug Target" Cancers 14, no. 10: 2385. https://doi.org/10.3390/cancers14102385
APA StyleCicenas, J., Meskinyte-Kausiliene, E., Jukna, V., Rimkus, A., Simkus, J., & Soderholm, D. (2022). SGK1 in Cancer: Biomarker and Drug Target. Cancers, 14(10), 2385. https://doi.org/10.3390/cancers14102385