
Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
2. The Molecular Characteristic and Function of GPX1
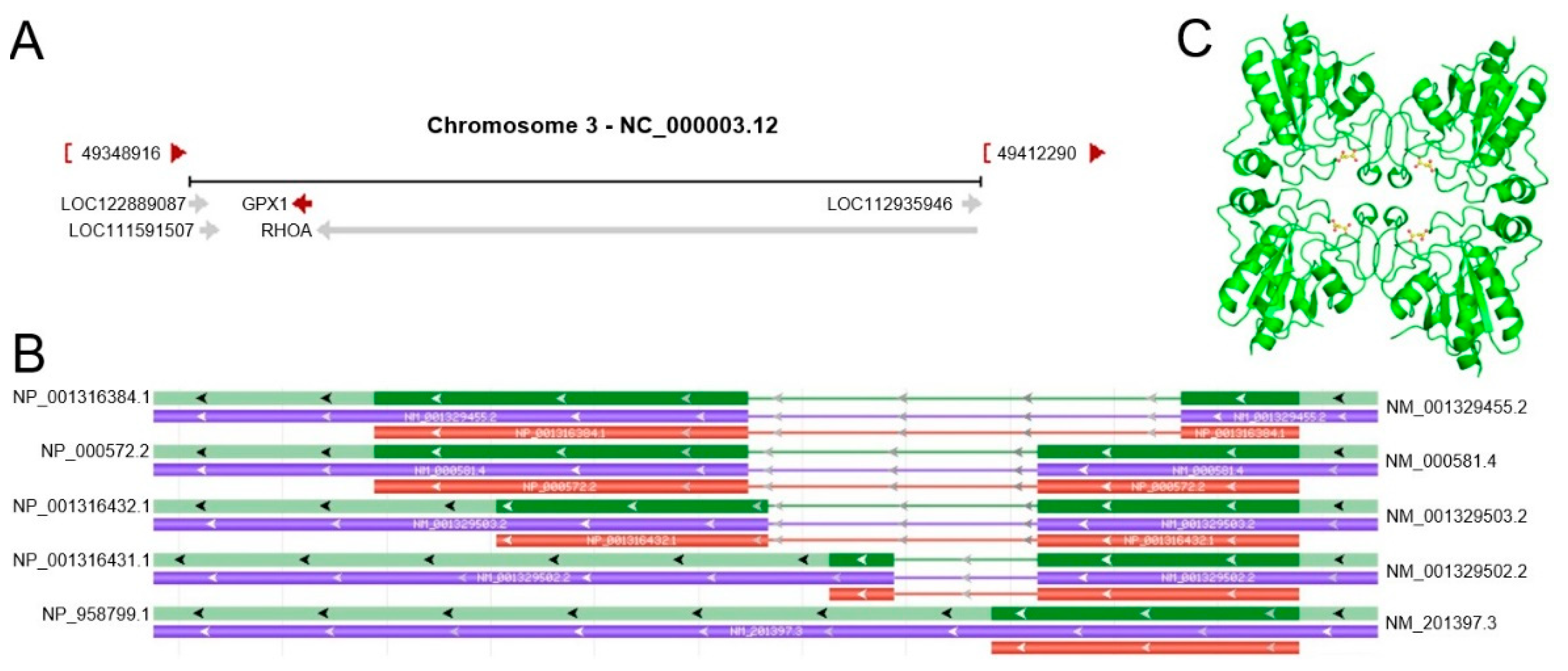
2.1. GPX1 Molecular Structure
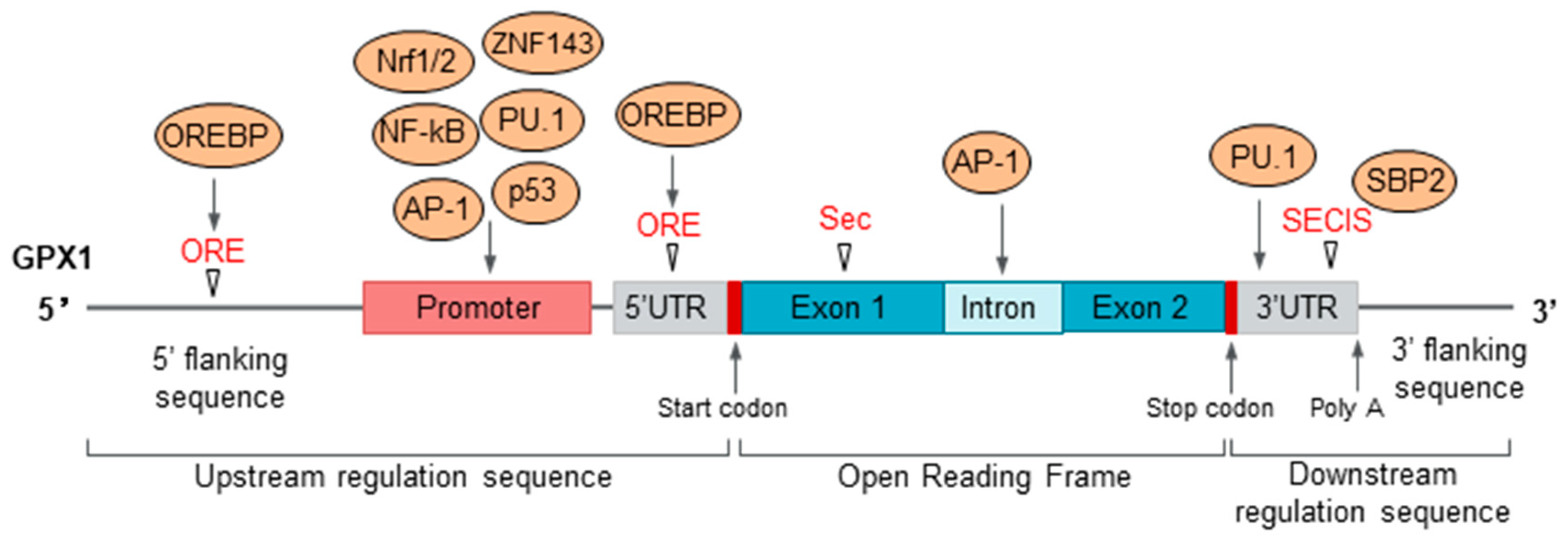
2.2. Regulation of GPX1 Expression and Activity
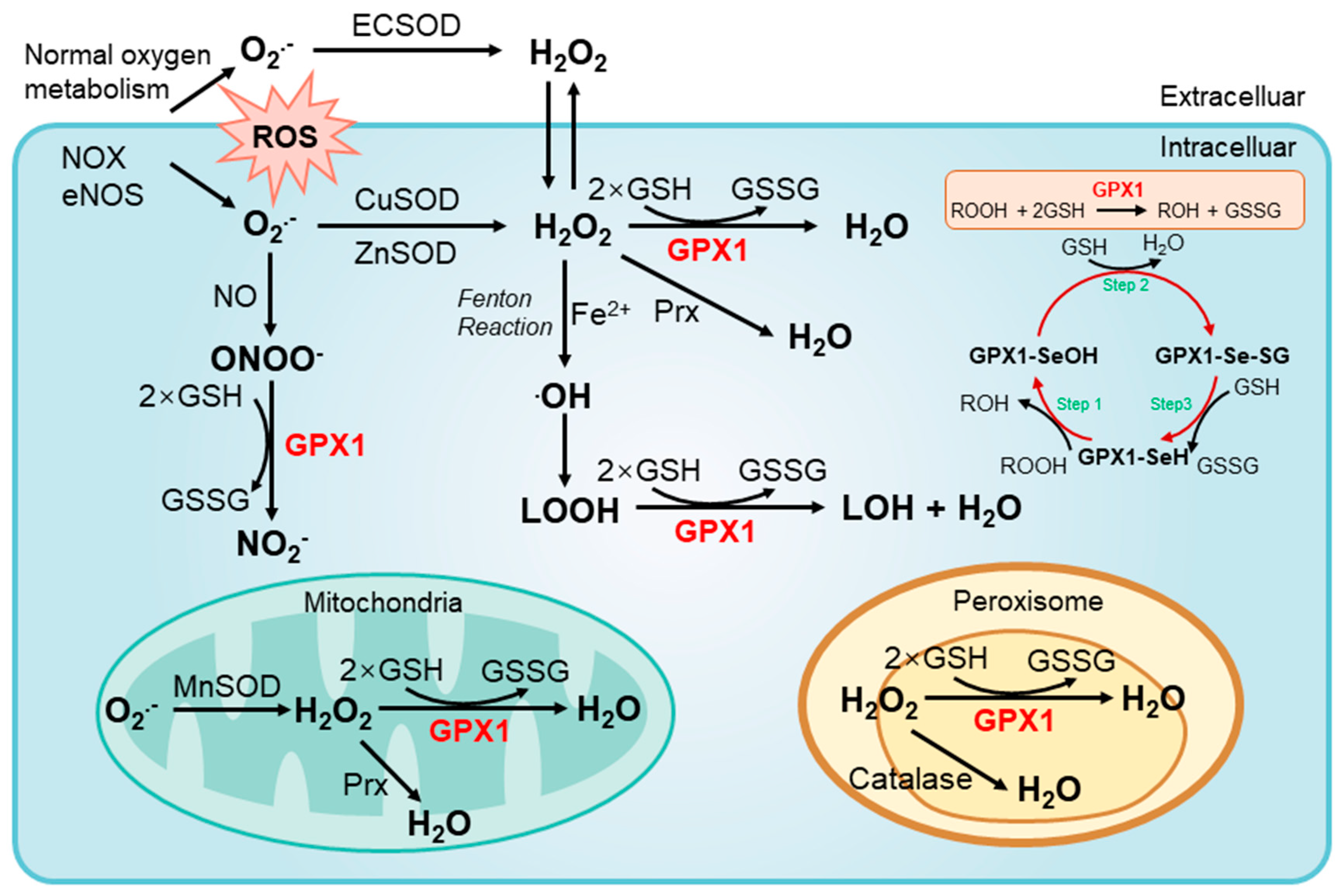
2.3. Enzymatic Mechanisms of GPX1
3. GPX1 and Cancer
3.1. GPX1 Gene Polymorphisms and Cancer Susceptibility
3.1.1. GPX1 Polymorphism and Breast Cancer Susceptibility
3.1.2. GPX1 Polymorphism and Bladder Cancer Susceptibility
3.1.3. GPX1 Polymorphism and Prostate Cancer Susceptibility
3.1.4. GPX1 Polymorphism and Other Types of Cancer Susceptibility

{kind=link}
{kind=link}
{kind=link}
| Tumor Type | Study Cohort (Race/Country) | GPX1 Polymorphism | Sample Type | Associations with Cancer risk | Refs. |
|---|---|---|---|---|---|
| Breast cancer | Caucasian, Rwanda | Pro198Leu -1040 G/A, Pro198Leu | Unknown, Peripheral blood | No association | [43,44] |
| Chicago/America, Denmark | Pro198Leu | Peripheral blood | Variant Leu-allele is associated with higher cancer risk and lower enzyme activity | [45,46] | |
| Meta-analysis | Pro198Leu | — | No association in Caucasians; Variant Leu-allele increases breast cancer risk in African population | [35] | |
| Altai krai/Russia, Poland | Pro198Leu | Unknown, Peripheral blood | Variant Leu-allele decreases sporadic breast cancer risk | [31,47] | |
| Bladder cancer | Egypt, Morocco, Serbia | Pro198Leu | Unknown, Peripheral blood | No association | [50,51,52] |
| Japan, Turkey, Ecuador | Pro198Leu | Peripheral blood, Tumor tissues in paraffin | Variant Leu-allele increases bladder cancer risk | [53,54,55] | |
| Meta-analysis | Pro198Leu | — | Variant Leu-allele increases bladder cancer risk | [34,36,56] | |
| America | Pro198Leu | Peripheral blood | Variant Leu-allele decreases bladder cancer risk | [57] | |
| Prostate cancer | Turkey | Pro198Leu | Peripheral blood | No association | [60,61] |
| Meta-analysis | Pro198Leu | — | No association | [36,62] | |
| Caucasian/England | GCG repeat | Peripheral blood | Ala6/Ala6 genotype increases in prostate cancer, but has no association with cancer risk | [59] | |
| Macedonia | Pro198Leu | Peripheral blood | Variant Leu-allele decreases prostate cancer risk | [37] | |
| Lung cancer | Caucasians and Chinese | Pro198Leu | Unknown | Variant Leu-allele increases bladder cancer risk in Caucasians but not Chinese | [33] |
| Denmark | Pro198Leu | Frozen lymphocytes | Variant Leu-allele decreases lung cancer risk | [38] | |
| Head and neck cancer | Meta-analysis | Pro198Leu | — | Variant Leu-allele increases head and neck cancer risk | [34] |
| England | GCG repeat | Peripheral blood | Ala6 and Ala7 genotypes increase in head and neck cancer risk | [63] | |
| Brain cancer | Meta-analysis | Pro198Leu | — | Variant Leu-allele increases bladder cancer risk | [34] |
| Colorectal cancer | Norway, Denmark | Pro198Leu | Peripheral blood | No association | [40,64] |
| Endometrial cancer | Poland | Pro198Leu | Peripheral blood | No association | [65] |
| Urothelial tumors | Balkan | Pro198Leu | Peripheral blood | No association | [66] |
| Chronic myeloid leukemia | Romania | Pro198Leu | Peripheral blood | No association | [39] |
| Papillary thyroid carcinoma | America | Pro198Leu | Peripheral blood | No association | [67] |
3.2. GPX1 Altered Expression and Activity in Cancer
3.2.1. GPX1 as Tumor Suppressor
- Pancreatic cancer
- Other cancers
3.2.2. GPX1 as Tumor Promoter
- Breast cancer
- Other cancers
3.3. Prognostic Values of GPX1
3.4. GPX1 Inhibitors and Therapeutic Potential
4. Discussion and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Herb, M.; Gluschko, A.; Schramm, M. Reactive oxygen species: Not omnipresent but important in many locations. Front. Cell Dev. Biol. 2021, 9, 716406. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Zhu, J.; Cheng, W.; Bao, Y.; Ho, Y.; Reddi, A.R.; Holmgren, A.; Arnér, E.S.J. Paradoxical roles of antioxidant enzymes: Basic mechanisms and health implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brigelius-Flohé, R.; Flohé, L. Regulatory phenomena in the glutathione peroxidase superfamily. Antioxid. Redox Sign. 2020, 33, 498–516. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Guo, S.; Wang, G. Glutathione peroxidases as oncotargets. Oncotarget 2017, 8, 80093–80102. [Google Scholar] [CrossRef] [Green Version]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Nirgude, S.; Choudhary, B. Insights into the role of GPX3, a highly efficient plasma antioxidant, in cancer. Biochem. Pharmacol. 2021, 184, 114365. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, J.; Wu, Y.; Ren, F.; Lei, X.G. Role of glutathione peroxidase 1 in glucose and lipid metabolism-related diseases. Free Radic. Bio. Med. 2018, 127, 108–115. [Google Scholar] [CrossRef]
- Sharma, G.; Shin, E.J.; Sharma, N.; Nah, S.Y.; Mai, H.N.; Nguyen, B.T.; Jeong, J.H.; Lei, X.G.; Kim, H.C. Glutathione peroxidase-1 and neuromodulation: Novel potentials of an old enzyme. Food Chem. Toxicol. 2021, 148, 111945. [Google Scholar] [CrossRef]
- Vlahos, R.; Bozinovski, S. Glutathione peroxidase-1 as a novel therapeutic target for COPD. Redox Rep. 2013, 18, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekoue, D.N.; He, C.; Diamond, A.M.; Bonini, M.G. Manganese superoxide dismutase and glutathione peroxidase-1 contribute to the rise and fall of mitochondrial reactive oxygen species which drive oncogenesis. Biochim. Biophys. Acta (BBA)-Bioenerg. 2017, 1858, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Sign. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipp, A.P. Selenium-dependent glutathione peroxidases during tumor development. Adv. Cancer Res. 2017, 136, 109–138. [Google Scholar]
- Miwa, T.; Adachi, T.; Ito, Y.; Hirano, K.; Sugiura, M. Purification and properties of glutathione peroxidase from human liver. Chem. Pharm. Bull. 1983, 31, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Hubert, N.; Walczak, R.; Sturchler, C.; Myslinski, E.; Schuster, C.; Westhof, E.; Carbon, P.; Krol, A. RNAs mediating cotranslational insertion of selenocysteine in eukaryotic selenoproteins. Biochimie 1996, 78, 590–596. [Google Scholar] [CrossRef]
- Berry, M.J.; Banu, L.; Harney, J.W.; Larsen, P.R. Functional characterization of the eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons. Embo J. 1993, 12, 3315–3322. [Google Scholar] [CrossRef]
- Throm, S.L.; Klemsz, M.J. PU.1 regulates glutathione peroxidase expression in neutrophils. J. Leukoc. Biol. 2003, 74, 111–117. [Google Scholar] [CrossRef]
- Hussain, S.P.; Amstad, P.; He, P.; Robles, A.; Lupold, S.; Kaneko, I.; Ichimiya, M.; Sengupta, S.; Mechanic, L.; Okamura, S.; et al. P53-induced up-regulation of MnSOD and GPx but not catalase increases oxidative stress and apoptosis. Cancer Res. 2004, 64, 2350–2356. [Google Scholar] [CrossRef] [Green Version]
- Budanov, A.V. The role of tumor suppressor p53 in the antioxidant defense and metabolism. Subcell. Biochem. 2014, 85, 337–358. [Google Scholar]
- Wu, Y.; Lee, S.; Bobadilla, S.; Duan, S.Z.; Liu, X. High glucose-induced p53 phosphorylation contributes to impairment of endothelial antioxidant system. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 2355–2362. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.V.; Savini, I.; Duranti, G.; Caporossi, D.; Ceci, R.; Sabatini, S.; Avigliano, L. Nuclear factor κB and activating protein 1 are involved in differentiation-related resistance to oxidative stress in skeletal muscle cells. Free Radic. Bio. Med. 2004, 37, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Chen, Z.; Zhang, H.; Wang, Y.; Luo, Y.; Huang, P. ZNF143 transcription factor mediates cell survival through upregulation of the GPX1 activity in the mitochondrial respiratory dysfunction. Cell Death Dis. 2012, 3, e422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowan, D.B.; Weisel, R.D.; Williams, W.G.; Mickle, D.A. Identification of oxygen responsive elements in the 5′-flanking region of the human glutathione peroxidase gene. J. Biol. Chem. 1993, 268, 26904–26910. [Google Scholar] [CrossRef]
- Stoytcheva, Z.R.; Berry, M.J. Transcriptional regulation of mammalian selenoprotein expression. Biochim. Biophys. Acta 2009, 1790, 1429–1440. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Flores, J.N.; Shetty, S.P.; Dubey, A.; Copeland, P.R. The molecular biology of selenocysteine. BioMol. Concepts 2013, 4, 349–365. [Google Scholar] [CrossRef] [Green Version]
- Low, S.C.; Grundner-Culemann, E.; Harney, J.W.; Berry, M.J. SECIS-SBP2 interactions dictate selenocysteine incorporation efficiency and selenoprotein hierarchy. Embo J. 2000, 19, 6882–6890. [Google Scholar] [CrossRef] [Green Version]
- Handy, D.E.; Zhang, Y.; Loscalzo, J. Homocysteine down-regulates cellular glutathione peroxidase (GPx1) by decreasing translation. J. Biol. Chem. 2005, 280, 15518–15525. [Google Scholar] [CrossRef] [Green Version]
- de Haan, J.B.; Cooper, M.E. Targeted antioxidant therapies in hyperglycemia-mediated endothelial dysfunction. Front. Biosci. 2011, 3, 709–729. [Google Scholar] [CrossRef] [Green Version]
- Imyanitov, E.N.; Togo, A.V.; Hanson, K.P. Searching for cancer-associated gene polymorphisms: Promises and obstacles. Cancer Lett. 2004, 204, 3–14. [Google Scholar] [CrossRef]
- Jablonska, E.; Gromadzinska, J.; Peplonska, B.; Fendler, W.; Reszka, E.; Krol, M.B.; Wieczorek, E.; Bukowska, A.; Gresner, P.; Galicki, M.; et al. Lipid peroxidation and glutathione peroxidase activity relationship in breast cancer depends on functional polymorphism of GPX1. BMC Cancer 2015, 15, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonska, E.; Gromadzinska, J.; Reszka, E.; Wasowicz, W.; Sobala, W.; Szeszenia-Dabrowska, N.; Boffetta, P. Association between GPx1 Pro198Leu polymorphism, GPx1 activity and plasma selenium concentration in humans. Eur. J. Nutr. 2009, 48, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Ratnasinghe, D.; Tangrea, J.A.; Andersen, M.R.; Barrett, M.J.; Virtamo, J.; Taylor, P.R.; Albanes, D. Glutathione peroxidase codon 198 polymorphism variant increases lung cancer risk. Cancer Res. 2000, 60, 6381–6383. [Google Scholar]
- Wang, C.; Zhang, R.; Chen, N.; Yang, L.; Wang, Y.; Sun, Y.; Huang, L.; Zhu, M.; Ji, Y.; Li, W. Association between glutathione peroxidase-1 (GPX1) Rs1050450 polymorphisms and cancer risk. Int. J. Clin. Exp. Pathol. 2017, 10, 9527–9540. [Google Scholar] [PubMed]
- Hu, J.; Zhou, G.; Wang, N.; Wang, Y. GPX1 Pro198Leu polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat. 2010, 124, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Men, T.; Zhang, X.; Yang, J.; Shen, B.; Li, X.; Chen, D.; Wang, J. The rs1050450 C > T polymorphism of GPX1 is associated with the risk of bladder but not prostate cancer: Evidence from a meta-analysis. Tumor Biol. 2014, 35, 269–275. [Google Scholar] [CrossRef]
- Arsova-Sarafinovska, Z.; Matevska, N.; Eken, A.; Petrovski, D.; Banev, S.; Dzikova, S.; Georgiev, V.; Sikole, A.; Erdem, O.; Sayal, A.; et al. Glutathione peroxidase 1 (GPX1) genetic polymorphism, erythrocyte GPX activity, and prostate cancer risk. Int. Urol. Nephrol. 2009, 41, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Raaschou-Nielsen, O.; Sørensen, M.; Hansen, R.D.; Frederiksen, K.; Tjønneland, A.; Overvad, K.; Vogel, U. GPX1 Pro198Leu polymorphism, interactions with smoking and alcohol consumption, and risk for lung cancer. Cancer Lett. 2007, 247, 293–300. [Google Scholar] [CrossRef]
- Bănescu, C.; Trifa, A.P.; Voidăzan, S.; Moldovan, V.G.; Macarie, I.; Benedek Lazar, E.; Dima, D.; Duicu, C.; Dobreanu, M. CAT, GPX1, MnSOD, GSTM1, GSTT1, and GSTP1 genetic polymorphisms in chronic myeloid leukemia: A case-control study. Oxid. Med. Cell. Longev. 2014, 2014, 875861. [Google Scholar] [CrossRef] [Green Version]
- Hansen, R.; Sæbø, M.; Skjelbred, C.F.; Nexø, B.A.; Hagen, P.C.; Bock, G.; Bowitz Lothe, I.M.; Johnson, E.; Aase, S.; Hansteen, I.; et al. GPX Pro198Leu and OGG1 Ser326Cys polymorphisms and risk of development of colorectal adenomas and colorectal cancer. Cancer Lett. 2005, 229, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Cao, Q.; Qin, C.; Shao, P.; Wu, Y.; Wang, M.; Zhang, Z.; Yin, C. GPx-1 polymorphism (rs1050450) contributes to tumor susceptibility: Evidence from meta-analysis. J. Cancer Res. Clin. 2011, 137, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Tian, C.; Zhang, X. GPX1 gene Pro200Leu polymorphism, erythrocyte GPX activity, and cancer risk. Mol. Biol. Rep. 2013, 40, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.G.; Hankinson, S.E.; Kraft, P.; Hunter, D.J. No association between GPX1 Pro198Leu and breast cancer risk. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1821–1822. [Google Scholar]
- Habyarimana, T.; Bakri, Y.; Mugenzi, P.; Mazarati, J.B.; Attaleb, M.; El Mzibri, M. Association between glutathione peroxidase 1 codon 198 variant and the occurrence of breast cancer in Rwanda. Mol. Genet. Genom. Med. 2018, 6, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.J.; Diamond, A.M. Role of glutathione peroxidase 1 in breast cancer: Loss of heterozygosity and allelic differences in the response to selenium. Cancer Res. 2003, 63, 3347–3351. [Google Scholar]
- Ravn-Haren, G.; Olsen, A.; Tjønneland, A.; Dragsted, L.O.; Nexø, B.A.; Wallin, H.; Overvad, K.; Raaschou-Nielsen, O.; Vogel, U. Associations between GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis 2006, 27, 820–825. [Google Scholar] [CrossRef] [Green Version]
- Ermolenko, N.A.; Boiarskikh, U.A.; Sushko, A.G.; Voronina, E.N.; Selezneva, I.A.; Sinkina, T.V.; Lazarev, A.F.; Petrova, V.D.; Filipenko, M.L. Effect of point substitutions in the MnSOD, GPX1, and GSTP1 genes on the risk of familial and sporadic breast cancers in residents of the Altai region of the Russian Federation. Genetika 2010, 46, 1685–1691. [Google Scholar]
- Bera, S.; Weinberg, F.; Ekoue, D.N.; Ansenberger-Fricano, K.; Mao, M.; Bonini, M.G.; Diamond, A.M. Natural allelic variations in glutathione peroxidase-1 affect its subcellular localization and function. Cancer Res. 2014, 74, 5118–5126. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, P.; Goldberg, M.; Herman, L.; Lee, B.; Wang, H.; Brown, R.L.; Foster, C.B.; Peters, U.; Diamond, A.M. Molecular consequences of genetic variations in the glutathione peroxidase 1 selenoenzyme. Cancer Res. 2009, 69, 8183–8190. [Google Scholar] [CrossRef] [Green Version]
- Goerlitz, D.; El Daly, M.; Abdel-Hamid, M.; Saleh, D.A.; Goldman, L.; El Kafrawy, S.; Hifnawy, T.; Ezzat, S.; Abdel-Aziz, M.A.; Zaghloul, M.S.; et al. GSTM1, GSTT1 null variants, and GPX1 single nucleotide polymorphism are not associated with bladder cancer risk in Egypt. Cancer Epidemiol. Biomark. 2011, 20, 1552–1554. [Google Scholar] [CrossRef] [Green Version]
- Hadami, K.; Ameziane, E.H.R.; Ameur, A.; Dakka, N.; Abbar, M.; Al, B.A.; Attaleb, M.; El, M.M. Association between GPX1 Pro189Leu polymorphism and the occurrence of bladder cancer in Morocco. Cell. Mol. Biol. 2016, 62, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Nikic, P.; Dragicevic, D.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Coric, V.; Jovanovic, D.; Bumbasirevic, U.; Pekmezovic, T.; Simic, T.; Dzamic, Z.; et al. Association between GPX1 and SOD2 genetic polymorphisms and overall survival in patients with metastatic urothelial bladder cancer: A single-center study in Serbia. J. Buon 2018, 23, 1130–1135. [Google Scholar] [PubMed]
- Ichimura, Y.; Habuchi, T.; Tsuchiya, N.; Wang, L.; Oyama, C.; Sato, K.; Nishiyama, H.; Ogawa, O.; Kato, T. Increased risk of bladder cancer associated with a glutathione peroxidase 1 codon 198 variant. J. Urol. 2004, 172, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Paz-Y-Miño, C.; Muñoz, M.J.; López-Cortés, A.; Cabrera, A.; Palacios, A.; Castro, B.; Paz-Y-Miño, N.; Sánchez, M.E. Frequency of polymorphisms pro198leu in GPX-1 gene and ile58thr in MnSOD gene in the altitude Ecuadorian population with bladder cancer. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2009, 18, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Kucukgergin, C.; Sanli, O.; Amasyalı, A.S.; Tefik, T.; Seckin, S. Genetic variants of MnSOD and GPX1 and susceptibility to bladder cancer in a Turkish population. Med. Oncol. 2012, 29, 1928–1934. [Google Scholar] [CrossRef]
- Cao, M.; Mu, X.; Jiang, C.; Yang, G.; Chen, H.; Xue, W. Single-nucleotide polymorphisms of GPX1 and MnSOD and susceptibility to bladder cancer: A systematic review and meta-analysis. Tumor Biol. 2014, 35, 759–764. [Google Scholar] [CrossRef]
- Zhao, H.; Liang, D.; Grossman, H.B.; Wu, X. Glutathione peroxidase 1 gene polymorphism and risk of recurrence in patients with superficial bladder cancer. Urology 2005, 66, 769–774. [Google Scholar] [CrossRef]
- Moscow, J.A.; Schmidt, L.; Ingram, D.T.; Gnarra, J.; Johnson, B.; Cowan, K.H. Loss of heterozygosity of the human cytosolic glutathione peroxidase I gene in lung cancer. Carcinogenesis 1994, 15, 2769–2773. [Google Scholar] [CrossRef] [Green Version]
- Kote-Jarai, Z.; Durocher, F.; Edwards, S.M.; Hamoudi, R.; Jackson, R.A.; Ardern-Jones, A.; Murkin, A.; Dearnaley, D.P.; Kirby, R.; Houlston, R.; et al. Association between the GCG polymorphism of the selenium dependent GPX1 gene and the risk of young onset prostate cancer. Prostate Cancer Prostatic Dis. 2002, 5, 189–192. [Google Scholar] [CrossRef] [Green Version]
- Erdem, O.; Eken, A.; Akay, C.; Arsova-Sarafinovska, Z.; Matevska, N.; Suturkova, L.; Erten, K.; Özgök, Y.; Dimovski, A.; Sayal, A.; et al. Association of GPX1 polymorphism, GPX activity and prostate cancer risk. Hum. Exp. Toxicol. 2011, 31, 24–31. [Google Scholar] [CrossRef]
- Parlaktas, B.S.; Atilgan, D.; Gencten, Y.; Benli, I.; Ozyurt, H.; Uluocak, N.; Erdemir, F. A pilot study of the association of manganese superoxide dismutase and glutathione peroxidase 1 single gene polymorphisms with prostate cancer and serum prostate specific antigen levels. Arch. Med. Sci. 2015, 11, 994–1000. [Google Scholar] [PubMed]
- Liwei, L.; Wei, Z.; Ruifa, H.; Chunyu, L. Association between genetic variants in glutathione peroxidase 1 gene and risk of prostate cancer: A meta-analysis. Mol. Biol. Rep. 2012, 39, 8615–8619. [Google Scholar] [CrossRef]
- Jefferies, S.; Kotejarai, Z.; Goldgar, D.; Houlston, R.; Frazerwilliams, M.; Ahern, R.; Henk, J.; Gore, M.; Rhysevans, P.; Archer, D. Association between polymorphisms of the gene and second primary tumours after index squamous cell cancer of the head and neck. Oral Oncol. 2005, 41, 455–461. [Google Scholar] [CrossRef]
- Hansen, R.D.; Krath, B.N.; Frederiksen, K.; Tjønneland, A.; Overvad, K.; Roswall, N.; Loft, S.; Dragsted, L.O.; Vogel, U.; Raaschou-Nielsen, O. GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, interaction with alcohol consumption and smoking, and risk of colorectal cancer. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2009, 664, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Janowska, M.; Potocka, N.; Paszek, S.; Skrzypa, M.; Żulewicz, K.; Kluz, M.; Januszek, S.; Baszuk, P.; Gronwald, J.; Lubiński, J.; et al. An assessment of GPX1 (rs1050450), DIO2 (rs225014) and SEPP1 (rs7579) gene polymorphisms in women with endometrial cancer. Genes 2022, 13, 188. [Google Scholar] [CrossRef]
- Dragicevic, B.; Suvakov, S.; Jerotic, D.; Reljic, Z.; Djukanovic, L.; Zelen, I.; Pljesa-Ercegovac, M.; Savic-Radojevic, A.; Simic, T.; Dragicevic, D.; et al. Association of SOD2 (rs4880) and GPX1 (rs1050450) gene polymorphisms with risk of Balkan endemic nephropathy and its related tumors. Medicina 2019, 55, 435. [Google Scholar] [CrossRef] [Green Version]
- Salimi, S.; Harati Sadegh, M.; Eskandari, M.; Heidari, Z. The effects of the genetic polymorphisms of antioxidant enzymes on susceptibility to papillary thyroid carcinoma. Iubmb Life 2020, 72, 1045–1053. [Google Scholar] [CrossRef]
- Wei, R.; Qiu, H.; Xu, J.; Mo, J.; Liu, Y.; Gui, Y.; Huang, G.; Zhang, S.; Yao, H.; Huang, X.; et al. Expression and prognostic potential of GPX1 in human cancers based on data mining. Ann. Transl. Med. 2020, 8, 124. [Google Scholar] [CrossRef]
- Cullen, J.J.; Mitros, F.A.; Oberley, L.W. Expression of antioxidant enzymes in diseases of the human pancreas: Another link between chronic pancreatitis and pancreatic cancer. Pancreas 2003, 26, 23–27. [Google Scholar] [CrossRef]
- Cullen, J.J.; Weydert, C.; Hinkhouse, M.M.; Ritchie, J.; Domann, F.E.; Spitz, D.; Oberley, L.W. The role of manganese superoxide dismutase in the growth of pancreatic adenocarcinoma. Cancer Res. 2003, 63, 1297–1303. [Google Scholar]
- Liu, J.; Hinkhouse, M.M.; Sun, W.; Weydert, C.J.; Ritchie, J.M.; Oberley, L.W.; Cullen, J.J. Redox regulation of pancreatic cancer cell growth: Role of glutathione peroxidase in the suppression of the malignant phenotype. Hum. Gene Ther. 2004, 15, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Shi, S.; Liang, C.; Liang, D.; Hua, J.; Zhang, B.; Xu, J.; Yu, X. Abrogation of glutathione peroxidase-1 drives EMT and chemoresistance in pancreatic cancer by activating ROS-mediated Akt/GSK3β/Snail signaling. Oncogene 2018, 37, 5843–5857. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Xu, J.; Liang, C.; Liu, J.; Hua, J.; Zhang, Y.; Ni, Q.; Shi, S.; Yu, X. GPx1 is involved in the induction of protective autophagy in pancreatic cancer cells in response to glucose deprivation. Cell Death Dis. 2018, 9, 1187. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.Y.; Hou, Y.Y.; Chu, S.T.; Liu, C.F.; Huang, C.H.; Chen, H.C.; Hsiao, M.; Lu, P.J.; Wang, J.S.; Ger, L.P. Manganese superoxide dismutase and glutathione peroxidase as prognostic markers in patients with buccal mucosal squamous cell carcinomas. Head Neck 2011, 33, 1606–1615. [Google Scholar] [CrossRef]
- Lee, J.R.; Roh, J.; Lee, S.M.; Park, Y.; Cho, K.; Choi, S.; Nam, S.Y.; Kim, S.Y. Overexpression of glutathione peroxidase 1 predicts poor prognosis in oral squamous cell carcinoma. J. Cancer Res. Clin. 2017, 143, 2257–2265. [Google Scholar] [CrossRef]
- Min, S.Y.; Kim, H.S.; Jung, E.J.; Jung, E.J.; Jee, C.D.; Kim, W.H. Prognostic significance of glutathione peroxidase 1 (GPX1) down-regulation and correlation with aberrant promoter methylation in human gastric cancer. Anticancer Res. 2012, 32, 3169–3175. [Google Scholar]
- Metere, A.; Frezzotti, F.; Graves, C.E.; Vergine, M.; De Luca, A.; Pietraforte, D.; Giacomelli, L. A possible role for selenoprotein glutathione peroxidase (GPx1) and thioredoxin reductases (TrxR1) in thyroid cancer: Our experience in thyroid surgery. Cancer Cell Int. 2018, 18, 7. [Google Scholar] [CrossRef]
- Chen, S.; Su, X.; Mi, H.; Dai, X.; Li, S.; Chen, S.; Zhang, S. Comprehensive analysis of glutathione peroxidase-1 (GPX1) expression and prognostic value in three different types of renal cell carcinoma. Transl. Androl. Urol. 2020, 9, 2737–2750. [Google Scholar] [CrossRef]
- Cheng, Y.; Xu, T.; Li, S.; Ruan, H. GPX1, a biomarker for the diagnosis and prognosis of kidney cancer, promotes the progression of kidney cancer. Aging 2019, 11, 12165–12176. [Google Scholar] [CrossRef]
- Gouaze, V.; Mirault, M.E.; Carpentier, S.; Salvayre, R.; Levade, T.; Andrieu-Abadie, N. Glutathione peroxidase-1 overexpression prevents ceramide production and partially inhibits apoptosis in doxorubicin-treated human breast carcinoma cells. Mol. Pharmacol. 2001, 60, 488–496. [Google Scholar]
- Vibet, S.; Goupille, C.; Bougnoux, P.; Steghens, J.; Goré, J.; Mahéo, K. Sensitization by docosahexaenoic acid (DHA) of breast cancer cells to anthracyclines through loss of glutathione peroxidase (GPx1) response. Free. Radic. Biol. Med. 2008, 44, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Choi, A.; Jun, Y.; Kim, N.; Yook, J.I.; Kim, S.Y.; Lee, S.; Kang, S.W. Glutathione peroxidase-1 regulates adhesion and metastasis of triple-negative breast cancer cells via FAK signaling. Redox Biol. 2020, 29, 101391. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, E.K.; Kang, D.H.; Lee, J.; Hong, S.H.; Jeong, W.; Kang, S.W. Glutathione peroxidase-1 regulates ASK1-dependent apoptosis via interaction with TRAF2 in RIPK3-negative cancer cells. Exp. Mol. Med. 2021, 53, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, D.; Alesi, G.N.; Fan, J.; Kang, H.; Lu, Z.; Boggon, T.J.; Jin, P.; Yi, H.; Wright, E.R.; et al. Glutamate dehydrogenase 1 signals through antioxidant glutathione peroxidase 1 to regulate redox homeostasis and tumor growth. Cancer Cell 2015, 27, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Shen, Y.; Wei, J.; Liu, F. MicroRNA-153/Nrf-2/GPx1 pathway regulates radiosensitivity and stemness of glioma stem cells via reactive oxygen species. Oncotarget 2015, 6, 22006–22027. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Ding, G.; Gu, C.; Zhou, J.; Kuang, M.; Ji, Y.; He, Y.; Kondo, T.; Fan, J. Decreased selenium-binding protein 1 enhances glutathione peroxidase 1 activity and downregulates HIF-1α to promote hepatocellular carcinoma invasiveness. Clin. Cancer Res. 2012, 18, 3042–3053. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Fang, W.; Wang, Y.; Yang, W.; Xiong, B. Transforming growth factor-β1 induces glutathione peroxidase-1 and protects from H2O2-induced cell death in colon cancer cells via the Smad2/ERK1/2/HIF-1alpha pathway. Int. J. Mol. Med. 2012, 29, 906–912. [Google Scholar]
- Gan, X.; Chen, B.; Shen, Z.; Liu, Y.; Li, H.; Xie, X.; Xu, X.; Li, H.; Huang, Z.; Chen, J. High GPX1 expression promotes esophageal squamous cell carcinoma invasion, migration, proliferation and cisplatin-resistance but can be reduced by vitamin D. Int. J. Clin. Exp. Med. 2014, 7, 2530–2540. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Liu, Y.; Huang, Z.; Li, H.; Gan, X.; Shen, Z. 1,25-Dihydroxyvitamin D3 alleviates salivary adenoid cystic carcinoma progression by suppressing GPX1 expression through the NF-kappaB pathway. Int. J. Oncol. 2016, 48, 1271–1279. [Google Scholar] [CrossRef]
- Chen, B.; Shen, Z.; Wu, D.; Xie, X.; Xu, X.; Lv, L.; Dai, H.; Chen, J.; Gan, X. Glutathione peroxidase 1 promotes NSCLC resistance to cisplatin via ROS-induced activation of PI3K/AKT pathway. BioMed Res. Int. 2019, 2019, 7640547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xu, H.; You, Y.; Zhang, J.; Chen, R. High Gpx1 expression predicts poor survival in laryngeal squamous cell carcinoma. Auris Nasus Larynx 2018, 45, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peng, Y.; He, Y.; Xiao, Y.; Wang, Q.; Zhao, Y.; Zhang, T.; Wu, C.; Xie, Y.; Zhou, J.; et al. GPX1-associated prognostic signature predicts poor survival in patients with acute myeloid leukemia and involves in immunosuppression. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2022, 1868, 166268. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Emmrich, T.; Lemmerhirt, H.; Leffler, U.; Sydow, K.; Hirt, C.; Kiefer, T.; Link, A.; Bednarski, P.J. Identification of a glutathione peroxidase inhibitor that reverses resistance to anticancer drugs in human B-cell lymphoma cell lines. Bioorg. Med. Chem. Lett. 2012, 22, 6712–6715. [Google Scholar] [CrossRef] [PubMed]
- Gharib, A.F.; Alaa Eldeen, M.; Khalifa, A.S.; Elsawy, W.H.; Eed, E.M.; Askary, A.E.; Eid, R.A.; Soltan, M.A.; Raafat, N. Assessment of glutathione peroxidase-1 (GPX1) gene expression as a specific diagnostic and prognostic biomarker in malignant pleural mesothelioma. Diagnostics 2021, 11, 2285. [Google Scholar] [CrossRef]
- Ekoue, D.; Ansong, E.; Hong, L.; Nonn, L.; Macias, V.; Deaton, R.; Rupnow, R.; Gann, P.; Kajdacsy-Balla, A.; Diamond, A. GPX1 localizes to the nucleus in prostate epithelium and its levels are not associated with prostate cancer recurrence. Antioxidants 2018, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Chaudiere, J.; Wilhelmsen, E.C.; Tappel, A.L. Mechanism of selenium-glutathione peroxidase and its inhibition by mercaptocarboxylic acids and other mercaptans. J. Biol. Chem. 1984, 259, 1043–1050. [Google Scholar] [CrossRef]
- Behnisch Cornwell, S.; Bandaru, S.S.M.; Napierkowski, M.; Wolff, L.; Zubair, M.; Urbainsky, C.; Lillig, C.; Schulzke, C.; Bednarski, P.J. Pentathiepins: A novel class of glutathione peroxidase 1 inhibitors that induce oxidative stress, loss of mitochondrial membrane potential and apoptosis in human cancer cells. ChemMedChem 2020, 15, 1515–1528. [Google Scholar] [CrossRef]
- Lange, C.; Bednarski, P.J. In vitro assessment of synergistic effects in combinations of a temoporfin-based photodynamic therapy with glutathione peroxidase 1 inhibitors. Photodiagn. Photodyn. 2021, 36, 102478. [Google Scholar] [CrossRef]



| Tumor Type | Sample Types | Detection Methods | Expression (Tumor vs. Normal) | Roles in Cancer | Functions In Vitro | Functions In Vivo | Ref. |
|---|---|---|---|---|---|---|---|
| Pancreatic cancer | Patient tissues embedded in paraffin | Immunohistochemistry | Down | Tumor suppressor | — | — | [69] |
| Cells (BxPC-3, Capan-1, MIAPaCa-2, AsPC-1) | Western blot, Enzyme activity assay | Down (protein, immunoreactivity) | Tumor suppressor | — | — | [70] | |
| Cells (MIAPaCa-2, AsPC-1), Mice | — | — | Tumor suppressor | GPX1 overexpression inhibits cell growth, plating efficiency, soft agar plating efficiency | GPX1 overexpression slows tumor growth and increases animal survival | [71] | |
| Patient frozen tissues Patient tissues embedded in paraffin Cells (PANC-1, MiaPaCa-2, BxPC-3, CFPAC-1, SW1990) | RT-qPCR, Western blot, Immunohistochemistry | Down | Tumor suppressor | GPX1 silencing increases colony formation capacity, EMT phenotype, gemcitabine resistance | Decreased GPX1 expression predicts poor OS, induces EMT phenotype and gemcitabine resistance | [72] | |
| Cells (MiaPaCa-2, SW1990, PANC-1) | — | — | Tumor suppressor | GPX1 overexpression sensitizes cells to starvation-induced cell death | — | [73] | |
| Oral squamous cell carcinoma | Patient tissues embedded in paraffin | Immunohistochemistry | — | Tumor suppressor | — | High expression of GPX1 predicts better disease-specific survival | [74] |
| Patient tissues embedded in paraffin | Immunohistochemistry | Up (Protein) | Tumor promoter | — | GPX1 overexpression predicts nodal metastasis, advanced stage, high grade, invasion and poor survival | [75] | |
| Gastric cancer | Cells (SNU1, 5, 16, 216, 484, 601, 620, 638, 668, 719) Patient tissues embedded in paraffin | RT-qPCR, Immunohistochemistry, Bisulfite sequencing | Down (mRNA, protein) Up (promoter methylation) | Tumor suppressor | — | Decreased GPX1 expression predicts aggressiveness, lymphatic invasion, and poor survival | [76] |
| Thyroid cancer | Patient fresh tissues | — | Down | Tumor suppressor | — | Decreased GPX1 expression is related to the imbalance of oxidant/antioxidant system | [77] |
| Kidney cancer | Patient frozen tissues (KIRP) | RNA-sequencing | Up (mRNA) | Tumor suppressor (KIRP) | — | High GPX1 expression predicts better OS | [68] |
| Patient frozen tissues (RCC) | RNA-sequencing | Up (mRNA) | Tumor suppressor (KIRP), Tumor promoter (KICH, KIRC) | — | High GPX1 expression predicts poor prognosis in KICH and KIRC, but better OS in KIRP | [78] | |
| Cells (A-498, ACHN, 786-O, CAKI-1), Patient frozen tissues (RCC) | Western blot, Immunohistochemistry | Up (Protein) | Tumor promoter | GPX1 knockdown inhibits proliferation and clonogenic capacity | GPX1 overexpression predicts poor overall survival, distant metastasis, lymphatic metastasis, and tumor stage | [79] | |
| Breast cancer | Cells (T47D) | — | — | Tumor promoter | GPX1 overexpression inhibits doxorubicin-induced apoptosis | — | [80] |
| Cells (MDA-MB-231), Rat mammary tumors | — | — | Tumor promoter | — | Decreased GPX1 activity sensitizes breast cancer cells to anthracyclines | [81] | |
| Cells (MDA-MB-231, MDA-MB-468, Hs578T, BT-549) Mice | RT-qPCR, Western blot, enzyme activity analysis | Up (mRNA, protein, enzyme activity, in TNBC cells) | Tumor promoter | GPX1 expression promotes migration and invasion | GXP1 silencing reduces lung metastasis of TNBC cells | [82] | |
| Cells (MDA-MB-231) | — | — | Tumor promoter | GPx1 silencing increases TNF-α-induced apoptosis | GXP1 silencing reduces tumorigenic growth | [83] | |
| Cells (MDA-MB-231) | — | — | Tumor promoter | GPX1 activated by glutamate dehydrogenase 1 to promote cancer cell proliferation | GPX1 activated by glutamate dehydrogenase 1 to promote cancer cell growth | [84] | |
| Glioma | Glioma stem cells (U87, SU-2) | RT-qPCR, Western blot, Enzyme activity assay | Up (mRNA, protein, enzyme activity) | Tumor promoter | Increased GPX1 expression decreases ROS level, and increases radioresistance and stemness in GSCs | — | [85] |
| Patient frozen tissues | RNA-sequencing | Up (mRNA) | Tumor promoter | — | High GPX1 expression predicts poor OS and DFS in LGG | [68] | |
| Hepatocellular carcinoma | Patient fresh tissues | Enzyme activity assay | Up (enzyme activity) | Tumor promoter | — | Increased GPX1 activity correlates with vascular invasion | [86] |
| Colorectal cancer | Cells (DLD-1) | — | — | Tumor promoter | Increased GPX1 activity protects cancer cells from H2O2-induced cell death | — | [87] |
| Esophageal cancer | Cells (EC9706, EC109, K150, K180) | — | — | Tumor promoter | GPX1 overexpression promotes invasion, migration, proliferation, and cisplatin resistance | — | [88] |
| Salivary adenoid cystic carcinoma | Cells (ACC-M, SACC-83, ACC-2) | — | — | Tumor promoter | GPX1 overexpression promotes proliferation, invasion, migration and cisplatin resistance | — | [89] |
| Lung cancer | Cells (H1299) | — | — | Tumor promoter | GPX1 activated by glutamate dehydrogenase 1 to promote cancer cell proliferation | GPX1 activated by glutamate dehydrogenase 1 to promote cancer cell growth | [84] |
| Cells (A549, H1975, H460, H1650, GLC-82, H1993, H2170, Spc-a1, H1299) | RT-qPCR, Western blot | Up (in cisplatin-resistant cell lines) | Tumor promoter | GPX1 overexpression inhibits ROS accumulation and leads to cisplatin resistance | — | [90] | |
| Laryngeal squamous cell carcinoma | Patient frozen tissues Patient tissues embedded in paraffin | RT-qPCR, Immunohistochemistry | Up | Tumor promoter | — | GPX1 overexpression predicts nodal lymph node metastasis, TNM stage and poor survival | [91] |
| Acute myeloid leukemia | Patient frozen tissues | RNA-sequencing | Up (mRNA) | Tumor promoter | — | High GPX1 expression predicts poor OS | [68] |
| Cells (MV4-11), Patient frozen tissues | RT-qPCR, Western blot, RNA-sequencing | Up (mRNA) | Tumor promoter | GPX1 knockdown inhibits cell viability | High GPX1 expression is associated with poor survival and immunosuppression | [92] | |
| Lymphoma | Cells (DOGUM, GUMBUS) | Western blot | Up (protein, in chemoresistant cells) | Tumor promoter | GPX1 expression increases chemoresistance | — | [93] |
| Malignant pleural mesothelioma | Patient frozen tissues | RT-qPCR | Up (mRNA) | Tumor promoter | — | High GPX1 expression predicts poor OS | [94] |
| Prostate cancer | Patient tissues embedded in paraffin | Immunohistochemistry | — | — | — | GPX1 expression has no association with cancer recurrence | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, H.; Zhou, J.; Shao, Q. Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer. Cancers 2022, 14, 2560. https://doi.org/10.3390/cancers14102560
Zhao Y, Wang H, Zhou J, Shao Q. Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer. Cancers. 2022; 14(10):2560. https://doi.org/10.3390/cancers14102560
Chicago/Turabian StyleZhao, Yangjing, Hui Wang, Jingdong Zhou, and Qixiang Shao. 2022. "Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer" Cancers 14, no. 10: 2560. https://doi.org/10.3390/cancers14102560
APA StyleZhao, Y., Wang, H., Zhou, J., & Shao, Q. (2022). Glutathione Peroxidase GPX1 and Its Dichotomous Roles in Cancer. Cancers, 14(10), 2560. https://doi.org/10.3390/cancers14102560