Model Cell Lines and Tissues of Different HGSOC Subtypes Differ in Local Estrogen Biosynthesis



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. RNA Isolation
2.3. Quantitative PCR
2.4. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.5. Transcriptome and Proteome Analysis in HGSOC and Normal Tissues Using Publicly Available Databases
2.6. Hierarchical Clustering
2.7. Statistical Analysis
3. Results and Discussion
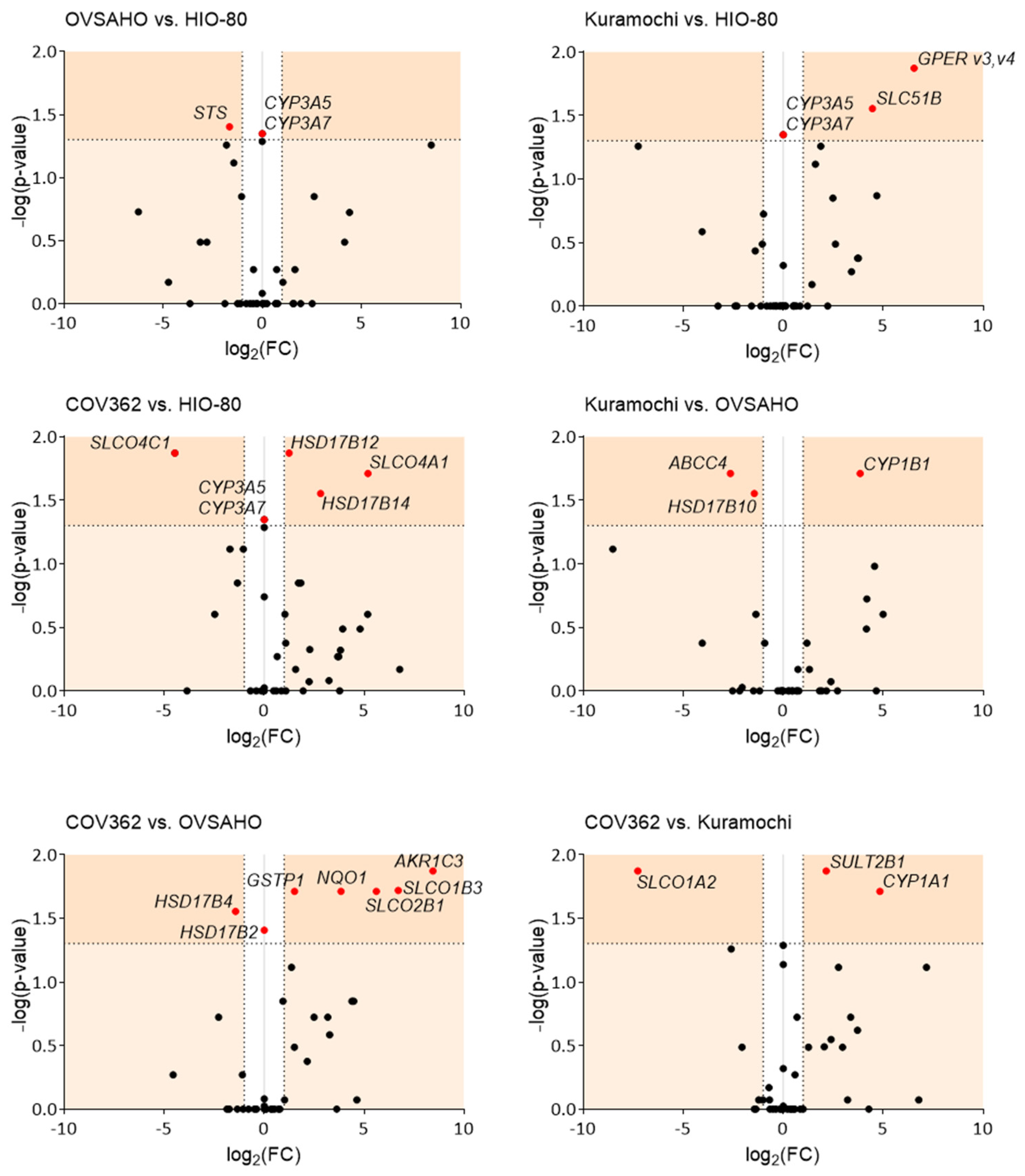
3.1. Targeted Transcriptomic Analysis Suggests That Estrogen Metabolism Differs between Different HGSOC Cells
3.2. Differential Gene Expression Correlates with Differences in Chemoresistance
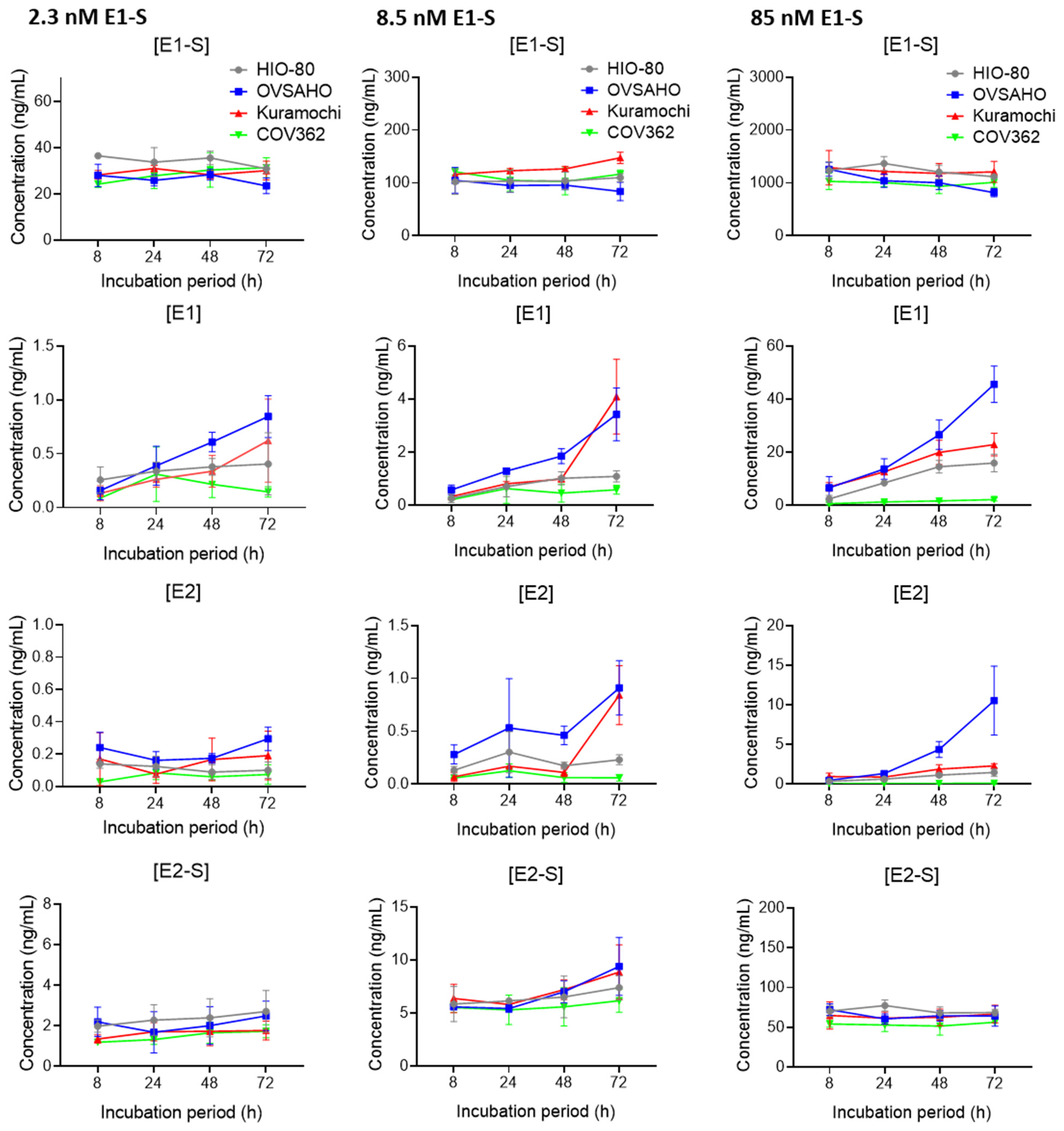
3.3. OVSAHO Cells Have the Highest Capacity for E1-S Uptake and Metabolism
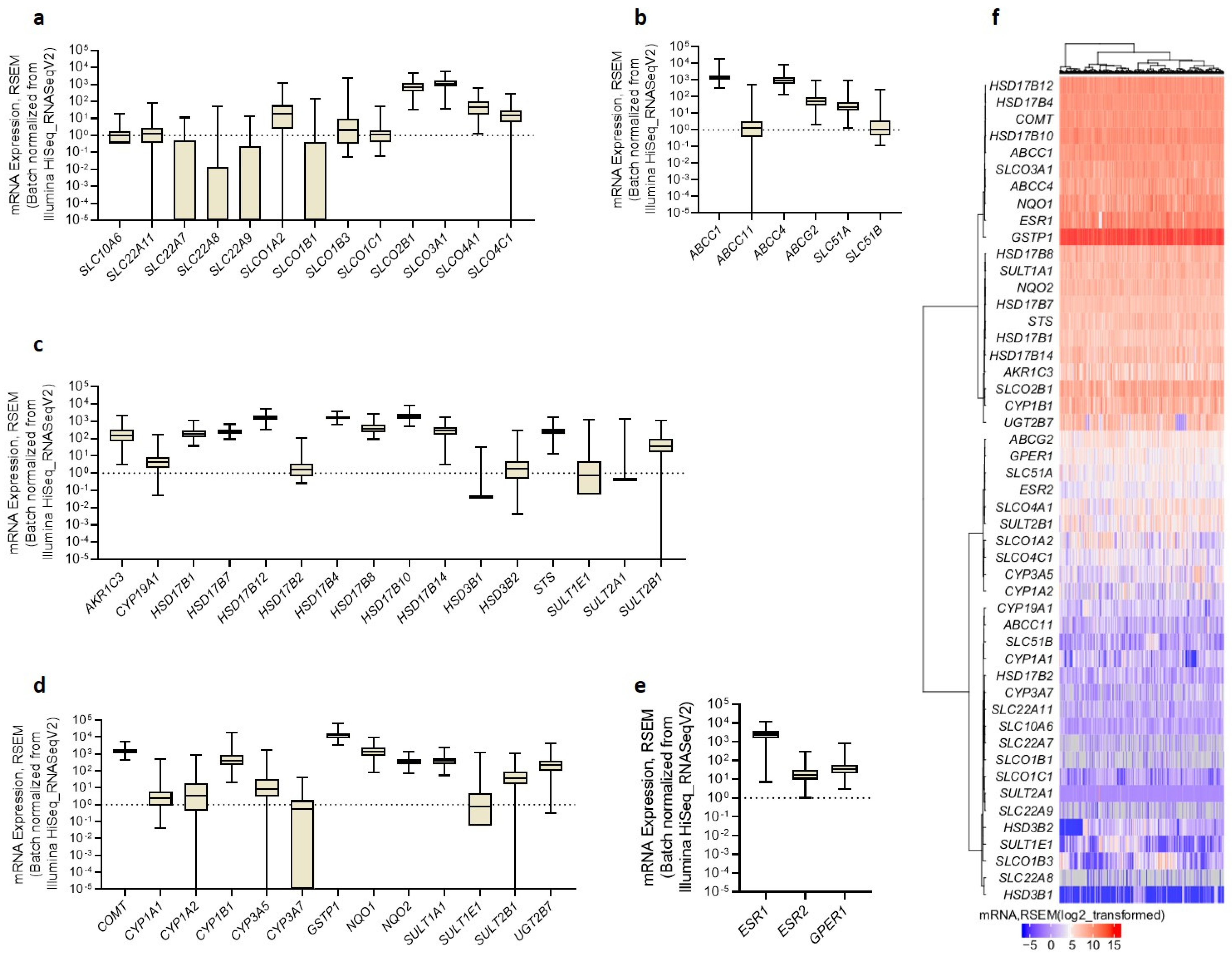
3.4. Gene Expression in Tissues Is in Line with Poor Prognosis of HGSOC
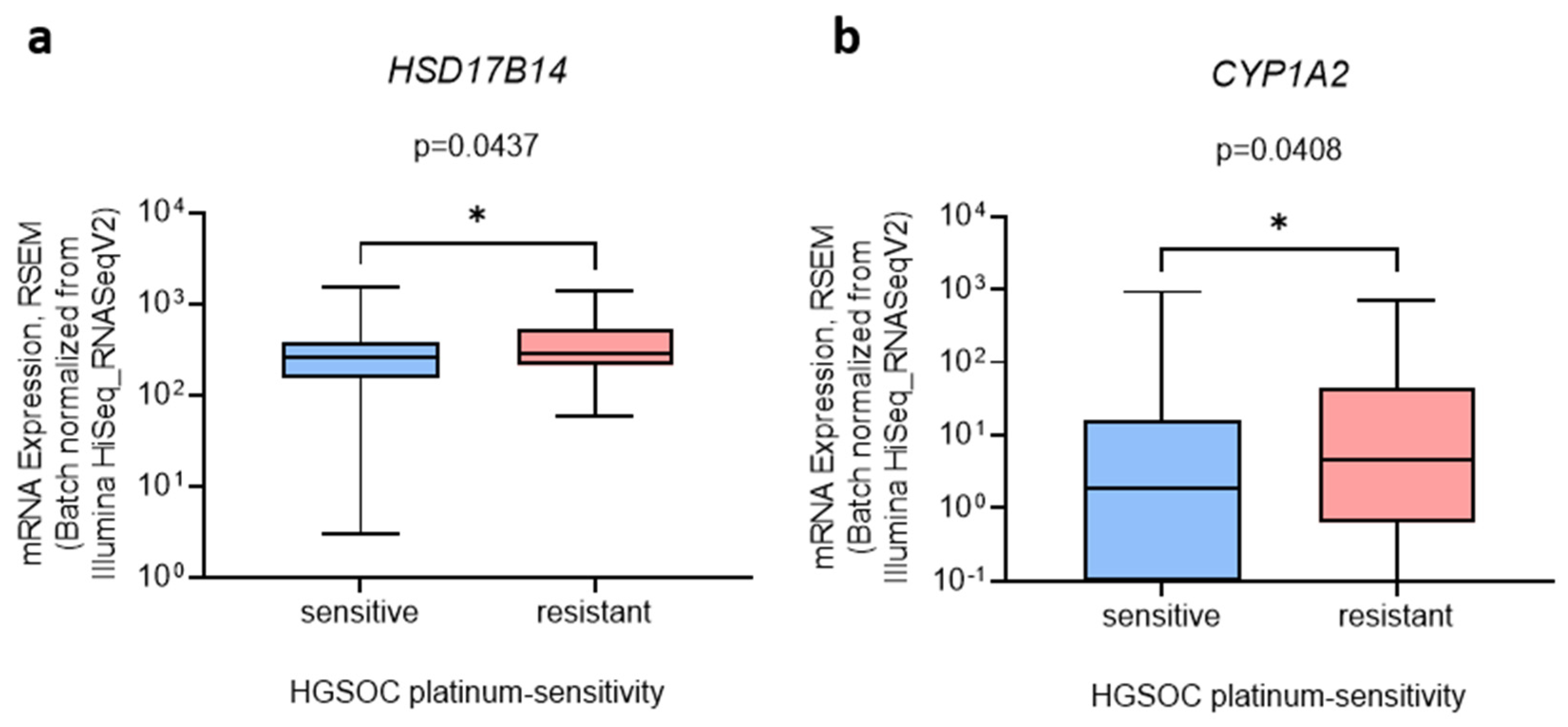
3.5. HSD17B14 and CYP1A2 Are Associated with Chemoresistance in HGSOC Tissues
3.6. Protein Levels in Tissues Suggest Differences in Estrogen Biosynthesis between HGSOC Subtypes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.; Ralyea, C.; Lockwood, S. Ovarian Cancer: An Integrated Review. Semin. Oncol. Nurs. 2019, 35, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.; Xu, J.; Aysola, K.; Qin, Y.; Okoli, C.; Hariprasad, R.; Chinemerem, U.; Gates, C.; Reddy, A.; Danner, O.; et al. Epithelial ovarian cancer: An overview. World J. Transl. Med. 2014, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kossaï, M.; Leary, A.; Scoazec, J.Y.; Genestie, C. Ovarian Cancer: A Heterogeneous Disease. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2018, 85, 41–49. [Google Scholar] [CrossRef]
- Vaughan, S.; Coward, J.I.; Bast, R.C., Jr.; Berchuck, A.; Berek, J.S.; Brenton, J.D.; Coukos, G.; Crum, C.C.; Drapkin, R.; Etemadmoghadam, D.; et al. Rethinking ovarian cancer: Recommendations for improving outcomes. Nat. Rev. Cancer 2011, 11, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Salazar, C.; Campbell, I.G.; Gorringe, K.L. When Is “Type I” Ovarian Cancer Not “Type I”? Indications of an Out-Dated Dichotomy. Front. Oncol. 2018, 8, 654. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Integrated genomic analyses of ovarian carcinoma. Nature 2011, 474, 609. [Google Scholar] [CrossRef]
- Hunn, J.; Rodriguez, G.C. Ovarian cancer: Etiology, risk factors, and epidemiology. Clin. Obstet. Gynecol. 2012, 55, 3–23. [Google Scholar] [CrossRef]
- Chura, J.C.; Ryu, H.S.; Simard, M.; Poirier, D.; Tremblay, Y.; Brooker, D.C.; Blomquist, C.H.; Argenta, P.A. Steroid-converting enzymes in human ovarian carcinomas. Mol. Cell Endocrinol. 2009, 301, 51–58. [Google Scholar] [CrossRef]
- Song, J.; Fadiel, A.; Edusa, V.; Chen, Z.; So, J.; Sakamoto, H.; Fishman, D.A.; Naftolin, F. Estradiol-induced ezrin overexpression in ovarian cancer: A new signaling domain for estrogen. Cancer Lett. 2005, 220, 57–65. [Google Scholar] [CrossRef]
- Park, S.H.; Cheung, L.W.; Wong, A.S.; Leung, P.C. Estrogen regulates Snail and Slug in the down-regulation of E-cadherin and induces metastatic potential of ovarian cancer cells through estrogen receptor alpha. Mol. Endocrinol. 2008, 22, 2085–2098. [Google Scholar] [CrossRef]
- Lee, Y.C.; Lheureux, S.; Oza, A.M. Treatment strategies for endometrial cancer: Current practice and perspective. Curr. Opin. Obstet. Gynecol. 2017, 29, 47–58. [Google Scholar] [CrossRef]
- Chan, K.K.; Leung, T.H.; Chan, D.W.; Wei, N.; Lau, G.T.; Liu, S.S.; Siu, M.K.; Ngan, H.Y. Targeting estrogen receptor subtypes (ERα and ERβ) with selective ER modulators in ovarian cancer. J. Endocrinol. 2014, 221, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Brandenberger, A.W.; Tee, M.K.; Jaffe, R.B. Estrogen receptor alpha (ER-alpha) and beta (ER-beta) mRNAs in normal ovary, ovarian serous cystadenocarcinoma and ovarian cancer cell lines: Down-regulation of ER-beta in neoplastic tissues. J. Clin. Endocrinol. Metab. 1998, 83, 1025–1028. [Google Scholar]
- Tan, J.; Song, C.; Wang, D.; Hu, Y.; Liu, D.; Ma, D.; Gao, Q. Expression of hormone receptors predicts survival and platinum sensitivity of high-grade serous ovarian cancer. Biosci. Rep. 2021, 41, BSR20210478. [Google Scholar] [CrossRef]
- Matsumura, S.; Ohta, T.; Yamanouchi, K.; Liu, Z.; Sudo, T.; Kojimahara, T.; Seino, M.; Narumi, M.; Tsutsumi, S.; Takahashi, T.; et al. Activation of estrogen receptor α by estradiol and cisplatin induces platinum-resistance in ovarian cancer cells. Cancer Biol. Ther. 2017, 18, 730–739. [Google Scholar] [CrossRef] [Green Version]
- Voutsadakis, I.A. Hormone Receptors in Serous Ovarian Carcinoma: Prognosis, Pathogenesis, and Treatment Considerations. Clinical Medicine Insights. Oncology 2016, 10, 17–25. [Google Scholar]
- Treeck, O.; Pfeiler, G.; Mitter, D.; Lattrich, C.; Piendl, G.; Ortmann, O. Estrogen receptor β1 exerts antitumoral effects on SK-OV-3 ovarian cancer cells. J. Endocrinol. 2007, 193, 421–433. [Google Scholar] [CrossRef]
- Pinton, G.; Nilsson, S.; Moro, L. Targeting estrogen receptor beta (ERβ) for treatment of ovarian cancer: Importance of KDM6B and SIRT1 for ERβ expression and functionality. Oncogenesis 2018, 7, 421–433. [Google Scholar] [CrossRef]
- Pavlič, R.; Gjorgoska, M.; Hafner, E.; Sinreih, M.; Gajser, K.; Poschner, S.; Jäger, W.; Rižner, T.L. In the Model Cell Lines of Moderately and Poorly Differentiated Endometrial Carcinoma, Estrogens Can Be Formed via the Sulfatase Pathway. Front. Mol. Biosci. 2021, 8, 1055. [Google Scholar] [CrossRef]
- Rižner, T.L.; Thalhammer, T.; Özvegy-Laczka, C. The Importance of Steroid Uptake and Intracrine Action in Endometrial and Ovarian Cancers. Front. Pharmacol. 2017, 8, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, B.; Edwards, R.P. Diagnosis and Treatment of Ovarian Cancer. Hematol./Oncol. Clin. N. Am. 2018, 32, 943–964. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.-S.; Chen, G.D.; Tsai, H.J.; Lee, M.S.; Wang, P.H.; Liu, F.S. Mechanisms Involved in Chemoresistance in Ovarian Cancer. Taiwan. J. Obstet. Gynecol. 2005, 44, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Penning, T.M.; Jonnalagadda, S.; Trippier, P.C.; Rižner, T.L. Aldo-Keto Reductases and Cancer Drug Resistance. Pharmacol. Rev. 2021, 73, 1150–1171. [Google Scholar] [CrossRef]
- Haley, J.; Tomar, S.; Pulliam, N.; Xiong, S.; Perkins, S.M.; Karpf, A.R.; Mitra, S.; Nephew, K.P.; Mitra, A.K. Functional characterization of a panel of high-grade serous ovarian cancer cell lines as representative experimental models of the disease. Oncotarget 2016, 7, 32810–32820. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.L.; Godwin, A.K.; Xu, X.X. Tumor necrosis factor-alpha-induced matrix proteolytic enzyme production and basement membrane remodeling by human ovarian surface epithelial cells: Molecular basis linking ovulation and cancer risk. Cancer Res. 2004, 64, 1534–1540. [Google Scholar] [CrossRef] [Green Version]
- Yanagibashi, T.; Gorai, I.; Nakazawa, T.; Miyagi, E.; Hirahara, F.; Kitamura, H.; Minaguchi, H. Complexity of expression of the intermediate filaments of six new human ovarian carcinoma cell lines: New expression of cytokeratin 20. Br. J. Cancer 1997, 76, 829–835. [Google Scholar] [CrossRef] [Green Version]
- Motoyama, T. Biological characterization including sensitivity to mitomycin C of cultured human ovarian cancers (author’s transl). Nihon Sanka Fujinka Gakkai Zasshi 1981, 33, 1197–1204. [Google Scholar]
- van den Berg-Bakker, C.A.; Hagemeijer, A.; Franken-Postma, E.M.; Smit, V.T.; Kuppen, P.J.; van Ravenswaay Claasen, H.H.; Cornelisse, C.J.; Schrier, P.I. Establishment and characterization of 7 ovarian carcinoma cell lines and one granulosa tumor cell line: Growth features and cytogenetics. Int. J. Cancer 1993, 53, 613–620. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.H.; Zhang, B.; McDermott, J.E.; Zhou, J.Y.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated Proteogenomic Characterization of Human High-Grade Serous Ovarian Cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Pan, J.; Shah, P.; Ao, M.; Thomas, S.N.; Liu, Y.; Chen, L.; Schnaubelt, M.; Clark, D.J.; Rodriguez, H.; et al. Integrated Proteomic and Glycoproteomic Characterization of Human High-Grade Serous Ovarian Carcinoma. Cell Rep. 2020, 33, 108276. [Google Scholar] [CrossRef]
- R Studio Team. RStudio: Integrated Development Environment for R; R Studio Team: Boston, MA, USA, 2016. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Poschner, S.; Wackerlig, J.; Castillo-Tong, D.C.; Wolf, A.; Decken, I.V.; Rižner, T.L.; Pavlič, R.; Meshcheryakova, A.; Mechtcheriakova, D.; Fritzer-Szekeres, M.; et al. Metabolism of Estrogens: Turnover Differs Between Platinum-Sensitive and -Resistant High-Grade Serous Ovarian Cancer Cells. Cancers 2020, 12, 279. [Google Scholar] [CrossRef] [Green Version]
- Kreitzburg, K.M.; Fehling, S.C.; Landen, C.N.; Gamblin, T.L.; Vance, R.B.; Arend, R.C.; Katre, A.A.; Oliver, P.G.; van Waardenburg, R.C.A.M.; Alvarez, R.D.; et al. FTY720 enhances the anti-tumor activity of carboplatin and tamoxifen in a patient-derived xenograft model of ovarian cancer. Cancer Lett. 2018, 436, 75–86. [Google Scholar] [CrossRef]
- Brandon, A.D.H.; Box, G.; Hallsworth, A.; Court, W.; Matthews, N.; Herodek, B.; Arteagabeitia, A.B.; Valenti, M.; Kirkin, V. Identification of ovarian high-grade serous carcinoma cell lines that show estrogen-sensitive growth as xenografts in immunocompromised mice. Sci. Rep. 2020, 10, 10799. [Google Scholar] [CrossRef]
- Li, Q.; Shu, Y. Role of solute carriers in response to anticancer drugs. Mol. Cell. Ther. 2014, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, T.; Li, G.; Lin, S.X. An unprecedented endocrine target for ovarian cancer: Inhibiting 17β-HSD7 supresses cancer cell proliferation and arrests G2/M cycle. Am. J. Cancer Res. 2021, 11, 5358. [Google Scholar]
- Liu, H.; Yan, Y.; Wen, H.; Jiang, X.; Cao, X.; Zhang, G.; Liu, G. A novel estrogen receptor GPER mediates proliferation induced by 17β-estradiol and selective GPER agonist G-1 in estrogen receptor α (ERα)-negative ovarian cancer cells. Cell Biol. Int. 2014, 38, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Sawers, L.; Ferguson, M.J.; Ihrig, B.R.; Young, H.C.; Chakravarty, P.; Wolf, C.R.; Smith, G. Glutathione S-transferase P1 (GSTP1) directly influences platinum drug chemosensitivity in ovarian tumour cell lines. Br. J. Cancer 2014, 111, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Domcke, S.; Sinha, R.; Levine, D.A.; Sander, C.; Schultz, N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat. Commun. 2013, 4, 2126. [Google Scholar] [CrossRef] [PubMed]
- Pavlič, R.; Vidic, S.; Anko, M.; Knific, T.; Büdefeld, T.; Marton, K.; Sinreih, M.; Poschner, S.; Jäger, W.; Frković-Grazio, S.; et al. Altered Profile of E1-S Transporters in Endometrial Cancer: Lower Protein Levels of ABCG2 and OSTβ and Up-Regulation of SLCO1B3 Expression. Int. J. Mol. Sci. 2021, 22, 3819. [Google Scholar] [CrossRef] [PubMed]
- Ballatori, N.; Christian, W.V.; Wheeler, S.G.; Hammond, C.L. The heteromeric organic solute transporter, OSTα–OSTβ/SLC51: A transporter for steroid-derived molecules. Mol. Aspects Med. 2013, 34, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svoboda, M.; Mungenast, F.; Gleiss, A.; Vergote, I.; Vanderstichele, A.; Sehouli, J.; Braicu, E.; Mahner, S.; Jäger, W.; Mechtcheriakova, D.; et al. Clinical Significance of Organic Anion Transporting Polypeptide Gene Expression in High-Grade Serous Ovarian Cancer. Front. Pharmacol. 2018, 9, 842. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, M.; Wlcek, K.; Taferner, B.; Hering, S.; Stieger, B.; Tong, D.; Zeillinger, R.; Thalhammer, T.; Jäger, W. Expression of organic anion-transporting polypeptides 1B1 and 1B3 in ovarian cancer cells: Relevance for paclitaxel transport. Biomed. Pharmacother. 2011, 65, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Reader, J.C.; Fan, C.; Ory, E.C.; Ju, J.; Lee, R.; Vitolo, M.I.; Smith, P.; Wu, S.; Ching, M.M.N.; Asiedu, E.B.; et al. Microtentacle Formation in Ovarian Carcinoma. Cancers 2022, 14, 800. [Google Scholar] [CrossRef]
- Kovacsics, D.; Patik, I.; Özvegy, L.C. The role of organic anion transporting polypeptides in drug absorption, distribution, excretion and drug-drug interactions. Expert Opin. Drug Metab. Toxicol. 2017, 13, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Mu, Y.; Qi, C.; Wang, J.; Xi, G.; Guo, J.; Mi, R.; Zhao, F. CYP1B1 enhances the resistance of epithelial ovarian cancer cells to paclitaxel in vivo and in vitro. Int. J. Mol. Med. 2015, 35, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Leung, Y.K.; Lau, K.M.; Mobley, J.; Jiang, Z.; Ho, S.M. Overexpression of cytochrome P450 1A1 and its novel spliced variant in ovarian cancer cells: Alternative subcellular enzyme compartmentation may contribute to carcinogenesis. Cancer Res. 2005, 65, 3726–3734. [Google Scholar] [CrossRef] [Green Version]
- Bagnoli, M.; Beretta, G.L.; Gatti, L.; Pilotti, S.; Alberti, P.; Tarantino, E.; Barbareschi, M.; Canevari, S.; Mezzanzanica, D.; Perego, P. Clinicopathological impact of ABCC1/MRP1 and ABCC4/MRP4 in epithelial ovarian carcinoma. BioMed Res. Int. 2013, 2013, 143202. [Google Scholar] [CrossRef]
- Elsnerova, K.; Mohelnikova-Duchonova, B.; Cerovska, E.; Ehrlichova, M.; Gut, I.; Rob, L.; Skapa, P.; Hruda, M.; Bartakova, A.; Bouda, J. Gene expression of membrane transporters: Importance for prognosis and progression of ovarian carcinoma. Oncol. Rep. 2022, 35, 2159–2170. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, Y.; Yue, W.; Zhang, L.; Gu, M.; Wang, Y. ABCC4 is required for cell proliferation and tumorigenesis in non-small cell lung cancer. OncoTargets Ther. 2014, 7, 343–351. [Google Scholar]
- Yang, X.; Xu, Y.; Guo, F.; Ning, Y.; Zhi, X.; Yin, L.; Li, X. Hydroxysteroid sulfotransferase SULT2B1b promotes hepatocellular carcinoma cells proliferation in vitro and in vivo. PLoS ONE 2013, 8, e60853. [Google Scholar] [CrossRef]
- Hernández-Silva, C.D.; Villegas-Pineda, J.C.; Pereira-Suárez, A.L. Expression and Role of the G Protein-Coupled Estrogen Receptor (GPR30/GPER) in the Development and Immune Response in Female Reproductive Cancers. Front. Endocrinol. 2020, 11, 544. [Google Scholar] [CrossRef]
- Puranen, T.; Poutanen, M.; Ghosh, D.; Vihko, P.; Vihko, R. Characterization of structural and functional properties of human 17 beta-hydroxysteroid dehydrogenase type 1 using recombinant enzymes and site-directed mutagenesis. Mol. Endocrinol. 1997, 11, 77–86. [Google Scholar]
- Wu, L.; Einstein, M.; Geissler, W.M.; Chan, H.K.; Elliston, K.O.; Andersson, S. Expression cloning and characterization of human 17 beta-hydroxysteroid dehydrogenase type 2, a microsomal enzyme possessing 20 alpha-hydroxysteroid dehydrogenase activity. J. Biol. Chem. 1993, 268, 12964–12969. [Google Scholar] [CrossRef]
- Luu-The, V.; Tremblay, P.; Labrie, F. Characterization of type 12 17beta-hydroxysteroid dehydrogenase, an isoform of type 3 17beta-hydroxysteroid dehydrogenase responsible for estradiol formation in women. Mol. Endocrinol. 2006, 20, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Lukacik, P.; Keller, B.; Bunkoczi, G.; Kavanagh, K.L.; Lee, W.H.; Adamski, J.; Oppermann, U. Structural and biochemical characterization of human orphan DHRS10 reveals a novel cytosolic enzyme with steroid dehydrogenase activity. Biochem. J. 2007, 402, 419–427. [Google Scholar] [CrossRef]
- Kemiläinen, H.; Huhtinen, K.; Auranen, A.; Carpén, O.; Strauss, L.; Poutanen, M. The Expression of HSD17B12 Is Associated with COX-2 Expression and Is Increased in High-Grade Epithelial Ovarian Cancer. Oncology 2018, 94, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Saito, H.; Sekizawa, A.; Shimizu, Y.; Akamatsu, T.; Kushima, M.; Yanaihara, T.; Okai, T.; Farina, A. Steroid sulfatase expression in ovarian clear cell adenocarcinoma: Immunohistochemical study. Gynecol. Oncol. 2001, 82, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Foster, P.A. Steroid Sulphatase and Its Inhibitors: Past, Present, and Future. Molecules 2021, 26, 2852. [Google Scholar] [CrossRef] [PubMed]
- Mungenast, F.; Aust, S.; Vergote, I.; Vanderstichele, A.; Sehouli, J.; Braicu, E.; Mahner, S.; Castillo-Tong, D.C.; Zeillinger, R.; Thalhammer, T. Clinical significance of the estrogen-modifying enzymes steroid sulfatase and estrogen sulfotransferase in epithelial ovarian cancer. Oncol. Lett. 2017, 13, 4047–4054. [Google Scholar] [CrossRef] [Green Version]
- Zahid, M.; Beseler, C.L.; Hall, J.B.; LeVan, T.; Cavalieri, E.L.; Rogan, E.G. Unbalanced estrogen metabolism in ovarian cancer. Int. J. Cancer 2014, 134, 2414–2423. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Li, L.; Yan, G.; Meng, K.; Lin, Z.; Nan, Y.; Jin, G.; Li, C. High expression of NQO1 is associated with poor prognosis in serous ovarian carcinoma. BMC Cancer 2015, 15, 244. [Google Scholar] [CrossRef] [Green Version]
- Bossard, C.; Busson, M.; Vindrieux, D.; Gaudin, F.; Machelon, V.; Brigitte, M.; Jacquard, C.; Pillon, A.; Balaguer, P.; Balabanian, K.; et al. Potential role of estrogen receptor beta as a tumor suppressor of epithelial ovarian cancer. PLoS ONE 2012, 7, e44787. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.K.L.; Siu, M.K.Y.; Jiang, Y.X.; Wang, J.J.; Wang, Y.; Leung, T.H.Y.; Liu, S.S.; Cheung, A.N.Y.; Ngan, H.Y.S. Differential expression of estrogen receptor subtypes and variants in ovarian cancer: Effects on cell invasion, proliferation and prognosis. BMC Cancer 2017, 17, 606. [Google Scholar] [CrossRef]
- Drummond, A.E.; Fuller, P.J. The importance of ERbeta signalling in the ovary. J. Endocrinol. 2010, 205, 15–23. [Google Scholar] [CrossRef]
- Jansson, A.K.; Gunnarsson, C.; Cohen, M.; Sivik, T.; Stål, O. 17beta-hydroxysteroid dehydrogenase 14 affects estradiol levels in breast cancer cells and is a prognostic marker in estrogen receptor-positive breast cancer. Cancer Res. 2006, 66, 11471–11477. [Google Scholar] [CrossRef] [Green Version]
- Sivik, T.; Gunnarsson, C.; Fornander, T.; Nordenskjöld, B.; Skoog, L.; Stål, O.; Jansson, A. 17β-Hydroxysteroid dehydrogenase type 14 is a predictive marker for tamoxifen response in oestrogen receptor positive breast cancer. PLoS ONE 2012, 7, e40568. [Google Scholar] [CrossRef] [Green Version]
- Tian, Z.; Li, Y.L.; Zhao, L.; Zhang, C.L. Role of CYP1A2 1F polymorphism in cancer risk: Evidence from a meta-analysis of 46 case-control studies. Gene 2013, 524, 168–174. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Z.; Han, S.; Lu, Y.; Feng, F.; Yuan, J. CYP1A2 rs762551 polymorphism contributes to cancer susceptibility: A meta-analysis from 19 case-control studies. BMC Cancer 2012, 12, 528. [Google Scholar] [CrossRef] [Green Version]
- Szajnik, M.; Szczepanski, M.J.; Elishaev, E.; Visus, C.; Lenzner, D.; Zabel, M.; Glura, M.; DeLeo, A.B.; Whiteside, T.L. 17β Hydroxysteroid dehydrogenase type 12 (HSD17B12) is a marker of poor prognosis in ovarian carcinoma. Gynecol. Oncol. 2012, 127, 587–594. [Google Scholar] [CrossRef] [Green Version]
- McFadyen, M.C.E.; Cruickshank, M.E.; Miller, I.D.; McLeod, H.L.; Melvin, W.T.; Haites, N.E.; Parkin, D.; Murray, G.I. Cytochrome P450 CYP1B1 over-expression in primary and metastatic ovarian cancer. Br. J. Cancer 2001, 85, 242–246. [Google Scholar] [CrossRef]
- Maecker, B.; Sherr, D.H.; Vonderheide, R.H.; von Bergwelt-Baildon, M.S.; Hirano, N.; Anderson, K.S.; Xia, Z.; Butler, M.O.; Wucherpfennig, K.W.; O’Hara, C.; et al. The shared tumor-associated antigen cytochrome P450 1B1 is recognized by specific cytotoxic T cells. Blood 2003, 102, 3287–3294. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Name | Assay | Assay ID or Sequences or Primers and/or Probes (5′ to 3′) |
|---|---|---|---|
| ABCC1 | Multidrug-resistance-associated protein 1 | SYBR Green | Forward primer: GGACTCAGGAGCACACGAAA |
| Reverse primer: ACGGCGATCCCTTGTGAAAT | |||
| ABCC11 | ATP-binding cassette subfamily C member 11 | SYBR Green | Forward primer: TCTCCATATATCCTGTTAAT |
| Reverse primer: TATAGTTCTCCAGTCTCTTG | |||
| ABCC4 | Multidrug-resistance-associated protein 4 | SYBR Green | Forward primer: AACTGCAACTTTCACGGATG |
| Reverse primer: AATGACTTTTCCCAGGCGTA | |||
| ABCG2 | Broad substrate specificity ATP-binding cassette transporter ABCG2 | SYBR Green | Forward primer: GGGTTTGGAACTGTGGGTAG |
| Reverse primer: AGATGATTCTGACGCACACC | |||
| AKR1C3 | Aldo-keto reductase family 1, member C3 (17β-hydroxysteroid dehydrogenase type 5) | TaqMan | Forward primer: GTTGCCTATAGTGCTCTGGGATCT |
| Reverse primer: GGACTGGGTC CTCCAAGAGG | |||
| Fluorescent MGB-NFQ probe: CACCCATCGTTTGTCTC FAM | |||
| COMT | Catechol-O-methyltransferase | Taqman | Hs00241349_m1 |
| CYP19A1 | Cytochrome P450, family 19, subfamily A | Taqman | Hs00240671_m1 |
| CYP1A1 | Cytochrome P450, family 1, subfamily A, polypeptide 1 | Taqman | Hs00153120_m1 |
| CYP1A2 | Cytochrome P450, family 1, subfamily A, polypeptide 2 | Taqman | Hs00167927_m1 |
| CYP1B1 | Cytochrome P450, family 1, subfamily B, polypeptide 1 | Taqman | Hs00164383_m1 |
| CYP3A5 | Cytochrome P450, family 3, subfamily A, polypeptide 5 | Taqman | Hs00241417_m1 |
| CYP3A7 | Cytochrome P450, family 3, subfamily A, polypeptide 7 | Taqman | Hs00426361_m1 |
| ESR1 | Estrogen receptor 1 (α) | TaqMan | Hs00174860_m1 |
| ESR2 | Estrogen receptor 2 (β) | TaqMan | Hs00230957_m1 |
| GPER v2 | G-protein–coupled estrogen receptor 1 (gene variant 2) | Taqman | Hs00173506_m1 |
| GPER v3, v4 | G-protein–coupled estrogen receptor 1 (gene variants 3 and 4) | Taqman | Hs01116133_m1 |
| GSTP1 | Glutathione S-transferase pi 1 | Taqman | Hs00168310_m1 |
| HSD17B1 | Hydroxysteroid (17β) dehydrogenase 1 | TaqMan | Hs00166219_g1 |
| HSD17B10 | Hydroxysteroid (17β) dehydrogenase 10 | TaqMan | Hs00189576_m1 |
| HSD17B12 | Hydroxysteroid (17β) dehydrogenase 12 | TaqMan | Hs00275054_m1 |
| HSD17B14 | Hydroxysteroid (17β) dehydrogenase 14 | Taqman | Hs00212233_m1 |
| HSD17B2 | Hydroxysteroid (17β) dehydrogenase 2 | TaqMan | Hs00157993_m1 |
| HSD17B4 | Hydroxysteroid (17β) dehydrogenase 4 | TaqMan | Hs00264973_m1 |
| HSD17B7 | Hydroxysteroid (17β) dehydrogenase 7 | Taqman | Hs00367686_m1 |
| HSD17B8 | Hydroxysteroid (17β) dehydrogenase 8 | TaqMan | Hs00367151_m1 |
| HSD3B1 | Hydroxy-delta-5-steroid dehydrogenase, 3β, and steroid delta-isomerase 1 | Taqman | Hs00426435 |
| HSD3B2 | Hydroxy-delta-5-steroid dehydrogenase, 3β, and steroid delta-isomerase 2 | Taqman | Hs00605123_m1 |
| NQO1 | NAD(P)H dehydrogenase, quinone 1 | Taqman | Hs00168547_m1 |
| NQO2 | NAD(P)H dehydrogenase, quinone 2 | Taqman | Hs00168552_m1 |
| POLR2A * | DNA-directed RNA polymerase II subunit RPB1 | TaqMan and SYBR Green | Hs00172187_m1 (TaqMan) |
| Forward primer: CAAGTTCAACCAAGCCATTG (SYBR) | |||
| Reverse primer: GTGGCAGGTTCTCCAAGG (SYBR) | |||
| RPLP0 * | 60S acidic ribosomal protein P0 | TaqMan and SYBR Green | Hs99999902_m1 (TaqMan) |
| Forward primer: AATGTGGGCTCCAAGCAGAT (SYBR) | |||
| Reverse primer: TTCTTGCCCATCAGCACCAC (SYBR) | |||
| SLC10A6 | Solute carrier family 10 member 6 | SYBR Green | Forward primer: TATGACAACCTGTTCCACCG |
| Reverse primer: GAATGGTCAGGCACACAAGG | |||
| SLC22A11 | Solute carrier family 22 member 11 | SYBR Green | Forward primer: CTCACCTTCATCCTCCCCTG |
| Reverse primer: CCATTGTCCAGCATGTGTGT | |||
| SLC22A7 | Solute carrier family 22 member 7 | SYBR Green | Forward primer: CCTCCAGAGTCCAAGGGTCT |
| Reverse primer: ATGCTGCTCACCCACCAAAT | |||
| SLC22A8 | Solute carrier family 22 member 8 | SYBR Green | Forward primer: TACGCTGGTTGGTCTTGTCT |
| Reverse primer: CTCCCTCTTCCTTCTTGCCA | |||
| SLC22A9 | Solute carrier family 22 member 9 | SYBR Green | Forward primer: CGGCTACCTATCTGACCCCA |
| Reverse primer: TCTTGACGACTGTGCTTCCC | |||
| SLC51A | Organic solute transporter subunit alpha | SYBR Green | Forward primer: GCCCTTTCCAATACGCCTTC |
| Reverse primer: TCTGCTGGGTCATAGATGCC | |||
| SLC51B | Organic solute transporter subunit beta | SYBR Green | Forward primer: GTGCTGTCAGTTTTCCTTCCG |
| Reverse primer: TCATGTGTCTGGCTTAGGATGG | |||
| SLCO1A2 | Solute carrier organic anion transporter family member 1A2 | SYBR Green | Forward primer: GTTGGCATCATTCTGTGCAAATGTT |
| Reverse primer: AACGAGTGTCAGTGGGAGTTATGAT | |||
| SLCO1B1 | Solute carrier organic anion transporter family member 1B1 | SYBR Green | Forward primer: CAAATTCTCATGTTTTACTG |
| Reverse primer: GATTATTTCCATCATAGGTC | |||
| SLCO1B3 | Solute carrier organic anion transporter family member 1B3 | SYBR Green | Forward primer: TCCAGTCATTGGCTTTGCAC |
| Reverse primer: TCCAACCCAACGAGAGTCCT | |||
| SLCO1C1 | Solute carrier organic anion transporter family member 1C1 | SYBR Green | Forward primer: CACACAGACTACCAAACACCC |
| Reverse primer: TCACCATGCCGAACAGAGAA | |||
| SLCO2B1 | Solute carrier organic anion transporter family member 2B1 | SYBR Green | Forward primer: AGAGCCCTGTGTTCCATTCT |
| Reverse primer: CTCTTGCTCCAGAAATGGCC | |||
| SLCO3A1 | Solute carrier organic anion transporter family member 3A1 | SYBR Green | Forward primer: CTACGACAATGTGGTCTAC |
| Reverse primer: TTTTGATGTAGCGTTTATAG | |||
| SLCO4A1 | Solute carrier organic anion transporter family member 4A1 | SYBR Green | Forward primer: ATGCACCAGTTGAAGGACAG |
| Reverse primer: AACAAGGTGGCAGCTTCTGAG | |||
| SULT1A1 | Sulfotransferase family 1A, member 1 | Taqman | Hs00738644_m1 |
| SULT1E1 | Sulfotransferase family 1E, estrogen-preferring, member 1 | Taqman | Hs00193690_m1 |
| SLCO4C1 | Solute carrier organic anion transporter family member 4C1 | SYBR Green | Forward primer: CCAGGAGCCCCAGAAGTC |
| Reverse primer: AACTCGGACAGCGACAGTG | |||
| STS | Steroid sulfatase (microsomal), isozyme S | TaqMan | Hs00165853_m1 |
| SULT2A1 | Sulfotransferase family, cytosolic, 2A, dehydroepiandrosterone-preferring, member 1 | Taqman | Hs00234219_m1 |
| SULT2B1 | Sulfotransferase family, cytosolic, 2B, member 1 | Taqman | Hs00190268_m1 |
| UGT2B7 | UDP glucuronosyltransferase 2 family, polypeptide B7 | Taqman | Hs00426592_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlič, R.; Gjorgoska, M.; Rižner, T.L. Model Cell Lines and Tissues of Different HGSOC Subtypes Differ in Local Estrogen Biosynthesis. Cancers 2022, 14, 2583. https://doi.org/10.3390/cancers14112583
Pavlič R, Gjorgoska M, Rižner TL. Model Cell Lines and Tissues of Different HGSOC Subtypes Differ in Local Estrogen Biosynthesis. Cancers. 2022; 14(11):2583. https://doi.org/10.3390/cancers14112583
Chicago/Turabian StylePavlič, Renata, Marija Gjorgoska, and Tea Lanišnik Rižner. 2022. "Model Cell Lines and Tissues of Different HGSOC Subtypes Differ in Local Estrogen Biosynthesis" Cancers 14, no. 11: 2583. https://doi.org/10.3390/cancers14112583
APA StylePavlič, R., Gjorgoska, M., & Rižner, T. L. (2022). Model Cell Lines and Tissues of Different HGSOC Subtypes Differ in Local Estrogen Biosynthesis. Cancers, 14(11), 2583. https://doi.org/10.3390/cancers14112583