
DNA Methylation Biomarkers for Prediction of Response to Platinum-Based Chemotherapy: Where Do We Stand?

 ,
,  ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Platinum-Based Chemotherapy: Brief Definition and Mechanisms of Action
1.2. A Brief Introduction to DNA Methylation
1.3. DNA Methylation Regulates Transcription and Affects Protein Levels
1.4. Epigenetic-Based Cancer Biomarkers
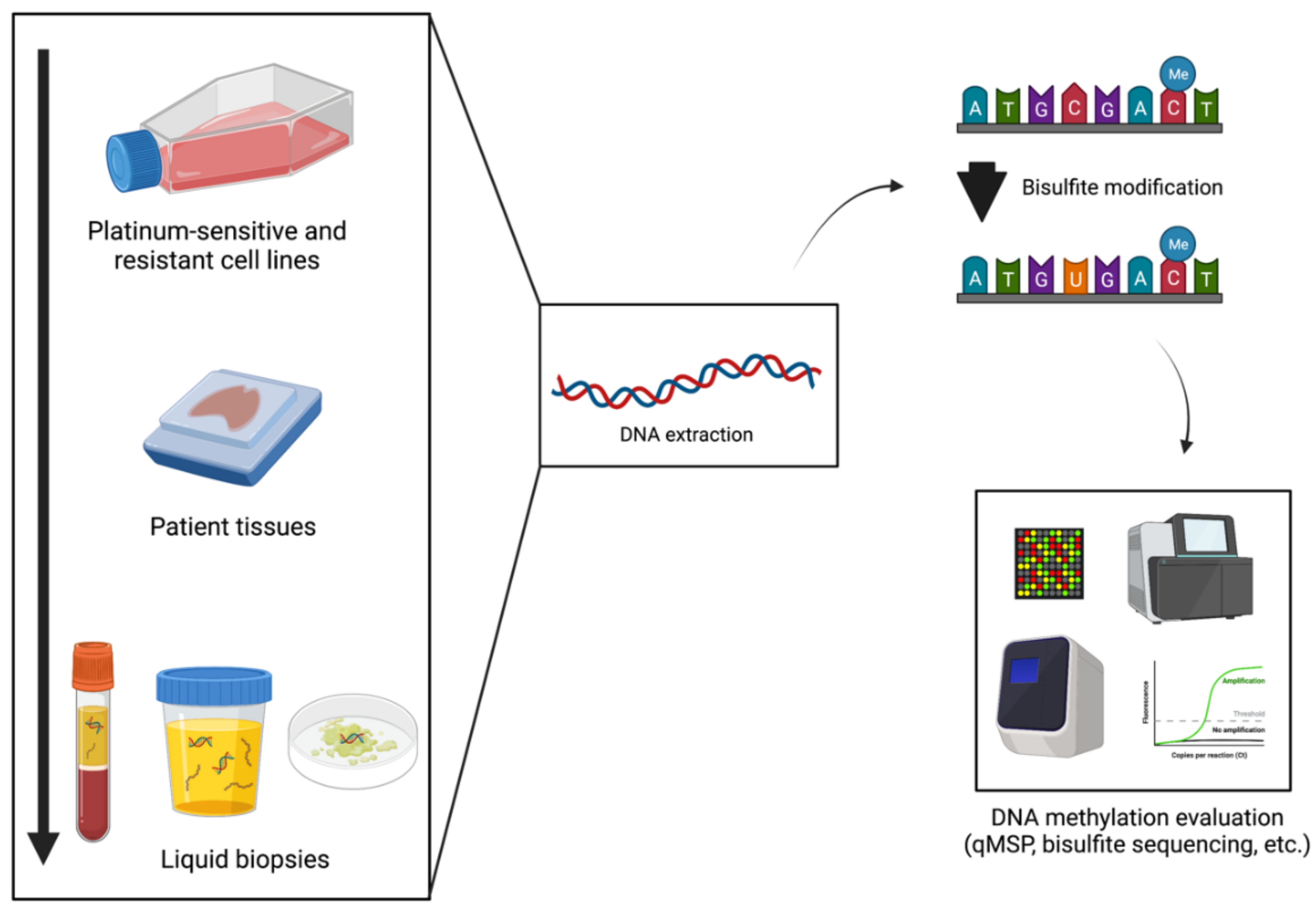
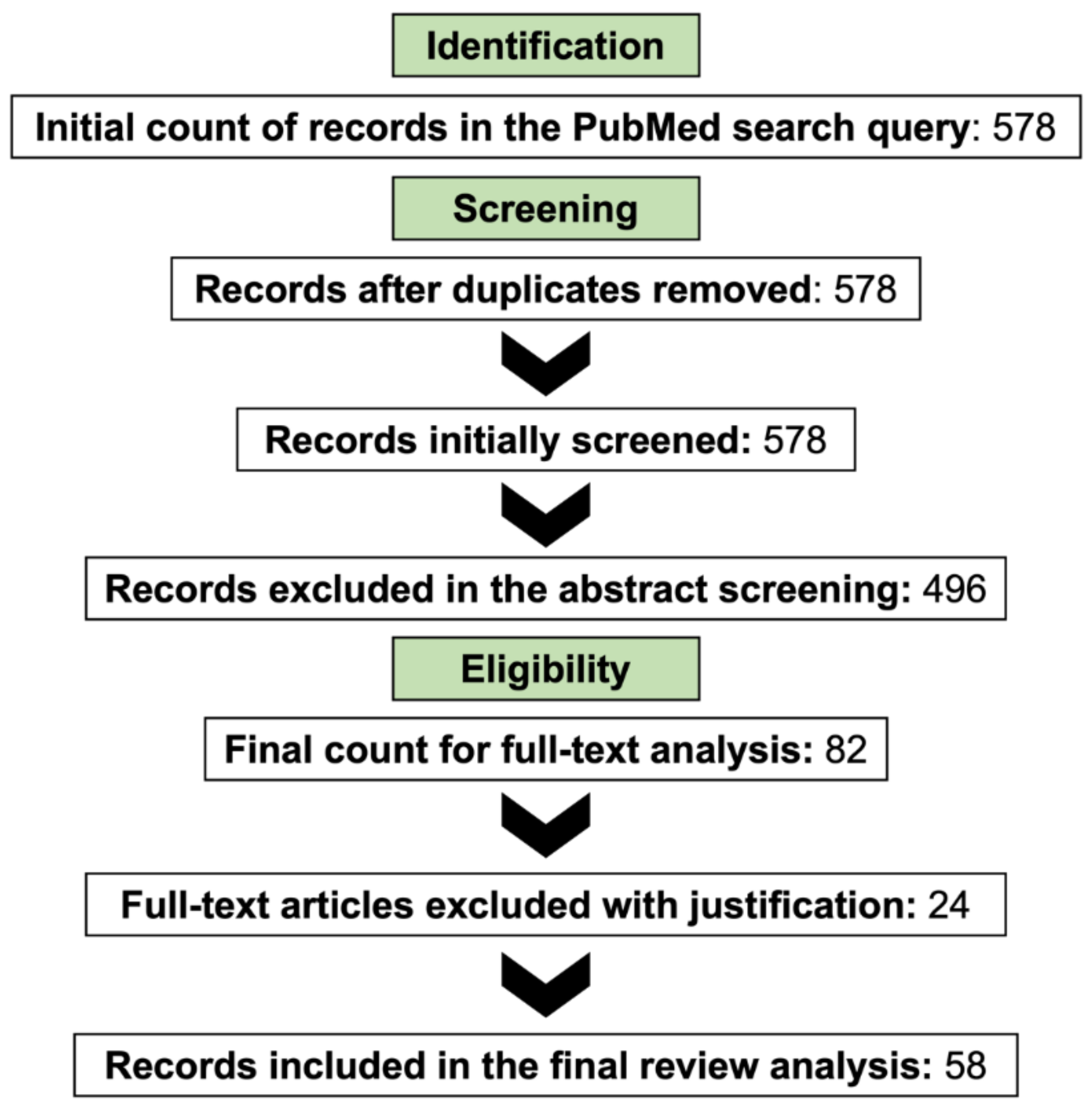
2. Research Methodology
3. Discussion
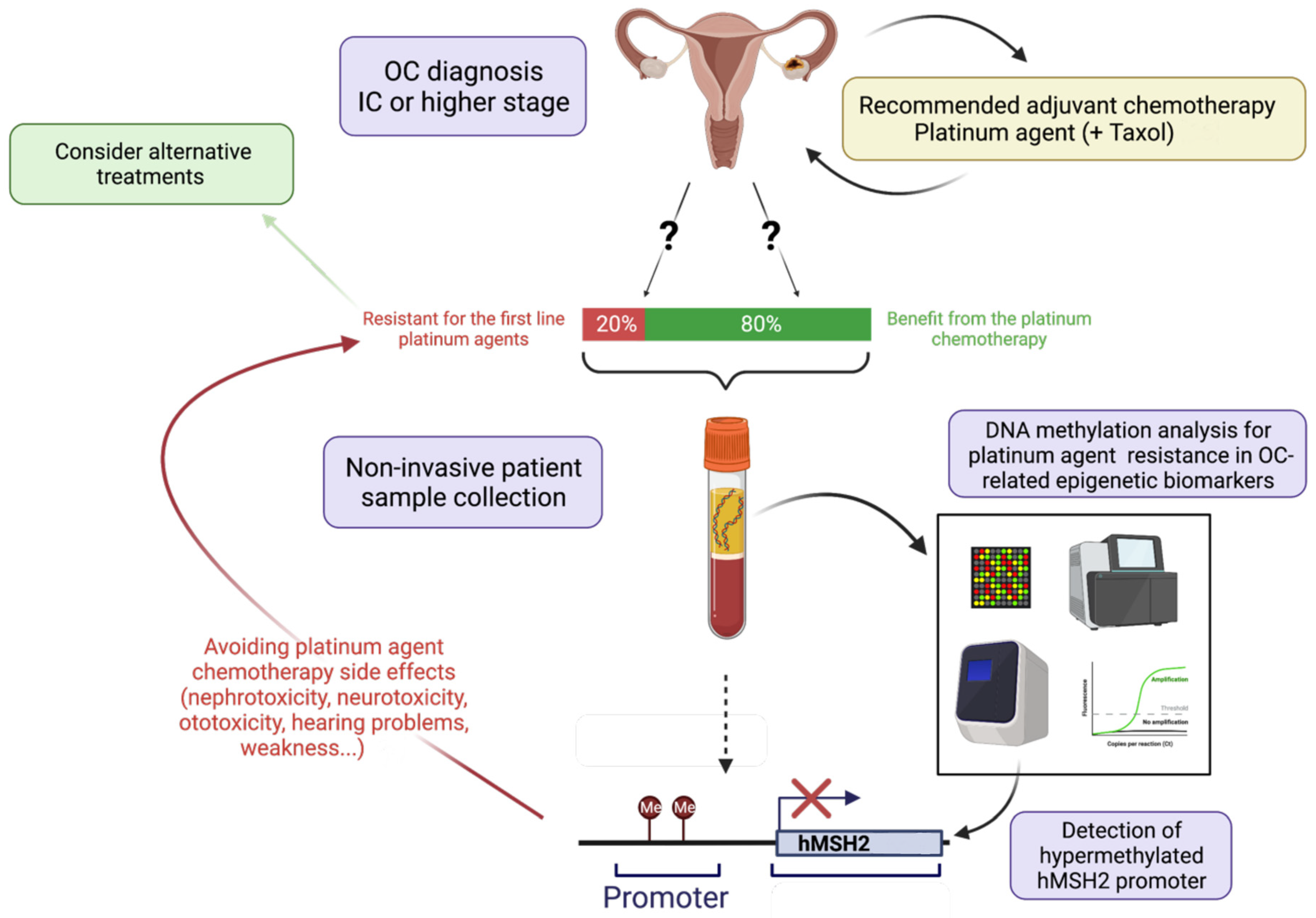
DNA Methylation and Platinum Resistance
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lobo, J.; Gillis, A.J.M.; van den Berg, A.; Dorssers, L.C.J.; Belge, G.; Dieckmann, K.P.; Roest, H.P.; van der Laan, L.J.W.; Gietema, J.; Hamilton, R.J.; et al. Identification and Validation Model for Informative Liquid Biopsy-Based microRNA Biomarkers: Insights from Germ Cell Tumor in Vitro, in Vivo and Patient-Derived Data. Cells 2019, 8, 1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amin, S.; Bathe, O.F. Response biomarkers: Re-envisioning the approach to tailoring drug therapy for cancer. BMC Cancer 2016, 16, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shariat, S.F.; Lotan, Y.; Vickers, A.; Karakiewicz, P.I.; Schmitz-Drager, B.J.; Goebell, P.J.; Malats, N. Statistical consideration for clinical biomarker research in bladder cancer. Urol. Oncol. 2010, 28, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, B.; Vancamp, L.; Trosko, J.E.; Mansour, V.H. Platinum Compounds: A New Class of Potent Antitumour Agents. Nature 1969, 222, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Amable, L. Cisplatin resistance and opportunities for precision medicine. Pharmacol. Res. 2016, 106, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Harrap, K.R. Preclinical studies identifying carboplatin as a viable cisplatin alternative. Cancer Treat. Rev. 1985, 12 (Suppl. A), 21–33. [Google Scholar] [CrossRef]
- Machover, D.; Diaz-Rubio, E.; de Gramont, A.; Schilf, A.; Gastiaburu, J.J.; Brienza, S.; Itzhaki, M.; Metzger, G.; N’Daw, D.; Vignoud, J.; et al. Two consecutive phase II studies of oxaliplatin (L-OHP) for treatment of patients with advanced colorectal carcinoma who were resistant to previous treatment with fluoropyrimidines. Ann. Oncol. 1996, 7, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharm. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Desoize, B.; Madoulet, C. Particular aspects of platinum compounds used at present in cancer treatment. Crit. Rev. Oncol./Hematol. 2002, 42, 317–325. [Google Scholar] [CrossRef]
- Brown, A.; Kumar, S.; Tchounwou, P.B. Cisplatin-Based Chemotherapy of Human Cancers. J. Cancer Sci. Ther. 2019, 11, 97. [Google Scholar]
- Skowron, M.A.; Oing, C.; Bremmer, F.; Ströbel, P.; Murray, M.J.; Coleman, N.; Amatruda, J.F.; Honecker, F.; Bokemeyer, C.; Albers, P.; et al. The developmental origin of cancers defines basic principles of cisplatin resistance. Cancer Lett. 2021, 519, 199–210. [Google Scholar] [CrossRef]
- Singh, R.; Fazal, Z.; Corbet, A.K.; Bikorimana, E.; Rodriguez, J.C.; Khan, E.M.; Shahid, K.; Freemantle, S.J.; Spinella, M.J. Epigenetic Remodeling through Downregulation of Polycomb Repressive Complex 2 Mediates Chemotherapy Resistance in Testicular Germ Cell Tumors. Cancers 2019, 11, 796. [Google Scholar] [CrossRef] [Green Version]
- Albers, P.; Albrecht, W.; Algaba, F.; Bokemeyer, C.; Cohn-Cedermark, G.; Fizazi, K.; Horwich, A.; Laguna, M.P.; Nicolai, N.; Oldenburg, J.; et al. Guidelines on Testicular Cancer: 2015 Update. Eur. Urol. 2015, 68, 1054–1068. [Google Scholar] [CrossRef]
- Lobo, J.; Constancio, V.; Guimaraes-Teixeira, C.; Leite-Silva, P.; Miranda-Goncalves, V.; Sequeira, J.P.; Pistoni, L.; Guimaraes, R.; Cantante, M.; Braga, I.; et al. Promoter methylation of DNA homologous recombination genes is predictive of the responsiveness to PARP inhibitor treatment in testicular germ cell tumors. Mol. Oncol. 2021, 15, 846–865. [Google Scholar] [CrossRef]
- Lobo, J.; Gillis, A.J.M.; Jeronimo, C.; Henrique, R.; Looijenga, L.H.J. Human Germ Cell Tumors are Developmental Cancers: Impact of Epigenetics on Pathobiology and Clinic. Int. J. Mol. Sci. 2019, 20, 258. [Google Scholar] [CrossRef] [Green Version]
- Kalavska, K.; Conteduca, V.; De Giorgi, U.; Mego, M. Molecular Mechanisms of Resistance in Testicular Germ Cell Tumors-clinical Implications. Curr. Cancer Drug Targets 2018, 18, 967–978. [Google Scholar] [CrossRef]
- Lobo, J.; Jeronimo, C.; Henrique, R. Morphological and molecular heterogeneity in testicular germ cell tumors: Implications for dedicated investigations. Virchows Arch. 2021, 479, 865–866. [Google Scholar] [CrossRef]
- Helfenstein, S.; Riesterer, O.; Meier, U.R.; Papachristofilou, A.; Kasenda, B.; Pless, M.; Rothschild, S.I. 3-weekly or weekly cisplatin concurrently with radiotherapy for patients with squamous cell carcinoma of the head and neck—A multicentre, retrospective analysis. Radiat. Oncol. 2019, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Arriagada, R.; Bergman, B.; Dunant, A.; Le Chevalier, T.; Pignon, J.P.; Vansteenkiste, J.; International Adjuvant Lung Cancer Trial Collaborative Group. Cisplatin-based adjuvant chemotherapy in patients with completely resected non-small-cell lung cancer. N. Engl. J. Med. 2004, 350, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Gong, Y.-B.; Xu, H.-M. Neoadjuvant therapy strategies for advanced gastric cancer: Current innovations and future challenges. Chronic Dis. Transl. Med. 2020, 6, 147–157. [Google Scholar] [CrossRef]
- Goel, S.; Sinha, R.J.; Bhaskar, V.; Aeron, R.; Sharma, A.; Singh, V. Role of gemcitabine and cisplatin as neoadjuvant chemotherapy in muscle invasive bladder cancer: Experience over the last decade. Asian J. Urol. 2019, 6, 222–229. [Google Scholar] [CrossRef]
- Rabik, C.A.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat. Rev. 2007, 33, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Chovanec, M.; Abu Zaid, M.; Hanna, N.; El-Kouri, N.; Einhorn, L.H.; Albany, C. Long-term toxicity of cisplatin in germ-cell tumor survivors. Ann. Oncol. 2017, 28, 2670–2679. [Google Scholar] [CrossRef]
- Santabarbara, G.; Maione, P.; Rossi, A.; Gridelli, C. Pharmacotherapeutic options for treating adverse effects of Cisplatin chemotherapy. Expert Opin. Pharmacother. 2016, 17, 561–570. [Google Scholar] [CrossRef]
- Hellesnes, R.; Myklebust, T.Å.; Fosså, S.D.; Bremnes, R.M.; Karlsdottir, Á.; Kvammen, Ø.; Tandstad, T.; Wilsgaard, T.; Negaard, H.F.S.; Haugnes, H.S. Testicular Cancer in the Cisplatin Era: Causes of Death and Mortality Rates in a Population-Based Cohort. J. Clin. Oncol. 2021, 39, 3561–3573. [Google Scholar] [CrossRef]
- Rocha, C.R.R.; Silva, M.M.; Quinet, A.; Cabral-Neto, J.B.; Menck, C.F.M. DNA repair pathways and cisplatin resistance: An intimate relationship. Clinics 2018, 73, e478s. [Google Scholar] [CrossRef] [PubMed]
- Blair, B.G.; Larson, C.A.; Safaei, R.; Howell, S.B. Copper Transporter 2 Regulates the Cellular Accumulation and Cytotoxicity of Cisplatin and Carboplatin. Clin. Cancer Res. 2009, 15, 4312–4321. [Google Scholar] [CrossRef] [Green Version]
- Ishida, S.; Lee, J.; Thiele, D.J.; Herskowitz, I. Uptake of the anticancer drug cisplatin mediated by the copper transporter Ctr1 in yeast and mammals. Proc. Natl. Acad. Sci. USA 2002, 99, 14298–14302. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Krishnamurthy, S. Cellular Responses to Cisplatin-Induced DNA Damage. J. Nucleic Acids 2010, 2010, 201367. [Google Scholar] [CrossRef] [Green Version]
- Yimit, A.; Adebali, O.; Sancar, A.; Jiang, Y. Differential damage and repair of DNA-adducts induced by anti-cancer drug cisplatin across mouse organs. Nat. Commun. 2019, 10, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faivre, S.; Chan, D.; Salinas, R.; Woynarowska, B.; Woynarowski, J.M. DNA strand breaks and apoptosis induced by oxaliplatin in cancer cells. Biochem. Pharmacol. 2003, 66, 225–237. [Google Scholar] [CrossRef]
- Lemaire, M.A.; Schwartz, A.; Rahmouni, A.R.; Leng, M. Interstrand cross-links are preferentially formed at the d(GC) sites in the reaction between cis-diamminedichloroplatinum (II) and DNA. Proc. Natl. Acad. Sci. USA 1991, 88, 1982–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Cui, J.; Wen, J.; Guo, Y.; Zhang, L.; Chen, X. Cisplatin induces HepG2 cell cycle arrest through targeting specific long noncoding RNAs and the p53 signaling pathway. Oncol. Lett. 2016, 12, 4605–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Ju, J.F.; Danenberg, K.D.; Danenberg, P.V. Inhibition of pre-mRNA splicing by cisplatin and platinum analogs. Int. J. Oncol. 2003, 23, 785–789. [Google Scholar] [CrossRef]
- Velma, V.; Dasari, S.R.; Tchounwou, P.B. Low Doses of Cisplatin Induce Gene Alterations, Cell Cycle Arrest, and Apoptosis in Human Promyelocytic Leukemia Cells. Biomark. Insights 2016, 11, BMI.S39445. [Google Scholar] [CrossRef] [Green Version]
- Sooriyaarachchi, M.; Narendran, A.; Gailer, J. Comparative hydrolysis and plasma protein binding of cis-platin and carboplatin in human plasma in vitro. Metallomics 2010, 3, 49–55. [Google Scholar] [CrossRef]
- Go, R.S.; Adjei, A.A. Review of the Comparative Pharmacology and Clinical Activity of Cisplatin and Carboplatin. J. Clin. Oncol. 1999, 17, 409. [Google Scholar] [CrossRef]
- Kiyonari, S.; Iimori, M.; Matsuoka, K.; Watanabe, S.; Morikawa-Ichinose, T.; Miura, D.; Niimi, S.; Saeki, H.; Tokunaga, E.; Oki, E.; et al. The 1,2-Diaminocyclohexane Carrier Ligand in Oxaliplatin Induces p53-Dependent Transcriptional Repression of Factors Involved in Thymidylate Biosynthesis. Mol. Cancer Ther. 2015, 14, 2332–2342. [Google Scholar] [CrossRef] [Green Version]
- Alian, O.M.; Azmi, A.S.; Mohammad, R.M. Network insights on oxaliplatin anti-cancer mechanisms. Clin. Transl. Med. 2012, 1, 26. [Google Scholar] [CrossRef] [Green Version]
- Saltz, L.B.; Clarke, S.; Díaz-Rubio, E.; Scheithauer, W.; Figer, A.; Wong, R.; Koski, S.; Lichinitser, M.; Yang, T.-S.; Rivera, F.; et al. Bevacizumab in Combination With Oxaliplatin-Based Chemotherapy As First-Line Therapy in Metastatic Colorectal Cancer: A Randomized Phase III Study. J. Clin. Oncol. 2008, 26, 2013–2019. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.; Mushin, M.; Kirkpatrick, P. Oxaliplatin. Nat. Rev. Drug Discov. 2004, 3, 11–12. [Google Scholar] [CrossRef]
- Comella, P.; Casaretti, R.; Sandomenico, C.; Avallone, A.; Franco, L. Role of oxaliplatin in the treatment of colorectal cancer. Clin. Risk Manag. 2009, 5, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Rossi, A.; Garcia-Manero, G. Epigenetic therapy of leukemia: An update. Int. J. Biochem. Cell Biol. 2009, 41, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.-H.; Waterland, R.A.; Zhang, P.; Schady, D.; Chen, M.-H.; Guan, Y.; Gadkari, M.; Shen, L. Targeted p16Ink4a epimutation causes tumorigenesis and reduces survival in mice. J. Clin. Investig. 2014, 124, 3708–3712. [Google Scholar] [CrossRef]
- Ward, R.L.; Dobbins, T.; Lindor, N.M.; Rapkins, R.W.; Hitchins, M.P. Identification of constitutional MLH1 epimutations and promoter variants in colorectal cancer patients from the Colon Cancer Family Registry. Genet. Med. 2013, 15, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019505. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Shin, W.; Lee, J.; Do, J. CpG and Non-CpG Methylation in Epigenetic Gene Regulation and Brain Function. Genes 2017, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- Baylin, S.; Fearon, E.; Vogelstein, B.; De Bustros, A.; Sharkis, S.; Burke, P.; Staal, S.; Nelkin, B. Hypermethylation of the 5′ region of the calcitonin gene is a property of human lymphoid and acute myeloid malignancies. Blood 1987, 70, 412–417. [Google Scholar] [CrossRef] [Green Version]
- Gaudet, F.; Hodgson, J.G.; Eden, A.; Jackson-Grusby, L.; Dausman, J.; Gray, J.W.; Leonhardt, H.; Jaenisch, R. Induction of Tumors in Mice by Genomic Hypomethylation. Science 2003, 300, 489–492. [Google Scholar] [CrossRef]
- Mass, M.J.; Wang, L. Arsenic alters cytosine methylation patterns of the promoter of the tumor suppressor gene p53 in human lung cells: A model for a mechanism of carcinogenesis. Mutat. Res. Rev. Mutat. Res. 1997, 386, 263–277. [Google Scholar] [CrossRef]
- Wang, R.Y.H.; Kuo, K.C.; Gehrke, C.W.; Huang, L.-H.; Ehrlich, M. Heat- and alkali-induced deamination of 5-methylcytosine and cytosine residues in DNA. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 1982, 697, 371–377. [Google Scholar] [CrossRef]
- Schuebel, K.E.; Chen, W.; Cope, L.; Glöckner, S.C.; Suzuki, H.; Yi, J.-M.; Chan, T.A.; Neste, L.V.; Criekinge, W.V.; Bosch, S.v.d.; et al. Comparing the DNA Hypermethylome with Gene Mutations in Human Colorectal Cancer. PLoS Genet. 2007, 3, e157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA Methyltransferases in Cancer: Biology, Paradox, Aberrations, and Targeted Therapy. Cancers 2020, 12, 2123. [Google Scholar] [CrossRef] [PubMed]
- Ulanovskaya, O.A.; Zuhl, A.M.; Cravatt, B.F. NNMT promotes epigenetic remodeling in cancer by creating a metabolic methylation sink. Nat. Chem. Biol. 2013, 9, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Campagna, R.; Pozzi, V.; Sartini, D.; Salvolini, E.; Brisigotti, V.; Molinelli, E.; Campanati, A.; Offidani, A.; Emanuelli, M. Beyond Nicotinamide Metabolism: Potential Role of Nicotinamide N-Methyltransferase as a Biomarker in Skin Cancers. Cancers 2021, 13, 4943. [Google Scholar] [CrossRef]
- Institute, N.C. Definition of Biomarker. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/biomarker (accessed on 20 February 2022).
- Mikeska, T.; Craig, J. DNA Methylation Biomarkers: Cancer and Beyond. Genes 2014, 5, 821–864. [Google Scholar] [CrossRef] [Green Version]
- Wians, F.H. Clinical Laboratory Tests: Which, Why, and What Do The Results Mean? Lab. Med. 2009, 40, 105–113. [Google Scholar] [CrossRef]
- Coppedè, F. Genetic and epigenetic biomarkers for diagnosis, prognosis and treatment of colorectal cancer. World J. Gastroenterol. 2014, 20, 943. [Google Scholar] [CrossRef]
- Watson, C.N.; Belli, A.; Di Pietro, V. Small Non-coding RNAs: New Class of Biomarkers and Potential Therapeutic Targets in Neurodegenerative Disease. Front. Genet. 2019, 10, 364. [Google Scholar] [CrossRef]
- Talens, R.P.; Boomsma, D.I.; Tobi, E.W.; Kremer, D.; Jukema, J.W.; Willemsen, G.; Putter, H.; Slagboom, P.E.; Heijmans, B.T. Variation, patterns, and temporal stability of DNA methylation: Considerations for epigenetic epidemiology. FASEB J. 2010, 24, 3135–3144. [Google Scholar] [CrossRef]
- Shen, D.-W.; Pouliot, L.M.; Hall, M.D.; Gottesman, M.M. Cisplatin Resistance: A Cellular Self-Defense Mechanism Resulting from Multiple Epigenetic and Genetic Changes. Pharmacol. Rev. 2012, 64, 706–721. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Peng, Y.; Gao, A.; Du, C.; Herman, J.G. Epigenetic heterogeneity in cancer. Biomark. Res. 2019, 7, 23. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- How Kit, A.; Nielsen, H.M.; Tost, J. DNA methylation based biomarkers: Practical considerations and applications. Biochimie 2012, 94, 2314–2337. [Google Scholar] [CrossRef]
- Bonito, N.A.; Borley, J.; Wilhelm-Benartzi, C.S.; Ghaem-Maghami, S.; Brown, R. Epigenetic Regulation of the Homeobox Gene MSX1 Associates with Platinum-Resistant Disease in High-Grade Serous Epithelial Ovarian Cancer. Clin. Cancer Res. 2016, 22, 3097–3104. [Google Scholar] [CrossRef] [Green Version]
- Kurdyukov, S.; Bullock, M. DNA Methylation Analysis: Choosing the Right Method. Biology 2016, 5, 3. [Google Scholar] [CrossRef]
- Wang, D.; O’Rourke, D.; Sanchez-Garcia, J.F.; Cai, T.; Scheuenpflug, J.; Feng, Z. Development of a liquid biopsy based purely quantitative digital droplet PCR assay for detection of MLH1 promoter methylation in colorectal cancer patients. BMC Cancer 2021, 21, 797. [Google Scholar] [CrossRef]
- Van Ginkel, J.H.; Huibers, M.M.H.; Van Es, R.J.J.; De Bree, R.; Willems, S.M. Droplet digital PCR for detection and quantification of circulating tumor DNA in plasma of head and neck cancer patients. BMC Cancer 2017, 17, 428. [Google Scholar] [CrossRef]
- Kader, F.; Ghai, M.; Maharaj, L. The effects of DNA methylation on human psychology. Behav. Brain Res. 2018, 346, 47–65. [Google Scholar] [CrossRef]
- Bock, C. Epigenetic biomarker development. Epigenomics 2009, 1, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Chen, Y.; Huang, Y.; Liu, Z.; Li, G. In vitro study of human mutL homolog 1 hypermethylation in inducing drug resistance of esophageal carcinoma. Irish J. Med. Sci. 2017, 186, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Yokomizo, A.; Shiota, M.; Tsunoda, T.; Plass, C.; Naito, S. Aberrant DNA methylation of T-cell leukemia, homeobox 3 modulates cisplatin sensitivity in bladder cancer. Int. J. Oncol. 2011, 39, 727–733. [Google Scholar] [CrossRef]
- Xylinas, E.; Hassler, M.; Zhuang, D.; Krzywinski, M.; Erdem, Z.; Robinson, B.; Elemento, O.; Clozel, T.; Shariat, S. An Epigenomic Approach to Improving Response to Neoadjuvant Cisplatin Chemotherapy in Bladder Cancer. Biomolecules 2016, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Guida, E.; Inokawa, Y.; Goldberg, R.; Reis, L.O.; Ooki, A.; Pilli, M.; Sadhukhan, P.; Woo, J.; Choi, W.; et al. GULP1 regulates the NRF2-KEAP1 signaling axis in urothelial carcinoma. Sci. Signal. 2020, 13, eaba0443. [Google Scholar] [CrossRef]
- Chen, C.C.; Lee, K.D.; Pai, M.Y.; Chu, P.Y.; Hsu, C.C.; Chiu, C.C.; Chen, L.T.; Chang, J.Y.; Hsiao, S.H.; Leu, Y.W. Changes in DNA methylation are associated with the development of drug resistance in cervical cancer cells. Cancer Cell Int. 2015, 15, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosof, L.; Yerram, S.; Armstrong, T.; Chu, N.; Danilova, L.; Yanagisawa, B.; Hidalgo, M.; Azad, N.; Herman, J.G. GPX3 promoter methylation predicts platinum sensitivity in colorectal cancer. Epigenetics 2017, 12, 540–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, B.; Lu, R.; Lou, W.; Bao, Y.; Qiao, L.; Hu, Y.; Liu, K.; Chen, J.; Bao, D.; Ye, M.; et al. KIF18b-dependent hypomethylation of PARPBP gene promoter enhances oxaliplatin resistance in colorectal cancer. Exp. Cell Res. 2021, 407, 112827. [Google Scholar] [CrossRef]
- He, T.; Zhang, M.; Zheng, R.; Zheng, S.; Linghu, E.; Herman, J.G.; Guo, M. Methylation of SLFN11 is a marker of poor prognosis and cisplatin resistance in colorectal cancer. Epigenomics 2017, 9, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.-P.; Li, L.; Yan, J.; Hou, X.-X.; Jia, Y.-X.; Chang, Z.-W.; Guan, X.-Y.; Qin, Y.-R. Down-Regulation of CIDEA Promoted Tumor Growth and Contributed to Cisplatin Resistance by Regulating the JNK-p21/Bad Signaling Pathways in Esophageal Squamous Cell Carcinoma. Front. Oncol. 2021, 10, 3294. [Google Scholar] [CrossRef]
- Iwabu, J.; Yamashita, S.; Takeshima, H.; Kishino, T.; Takahashi, T.; Oda, I.; Koyanagi, K.; Igaki, H.; Tachimori, Y.; Daiko, H.; et al. FGF5 methylation is a sensitivity marker of esophageal squamous cell carcinoma to definitive chemoradiotherapy. Sci. Rep. 2019, 9, 13347. [Google Scholar] [CrossRef] [Green Version]
- Kurimoto, K.; Hayashi, M.; Guerrero-Preston, R.; Koike, M.; Kanda, M.; Hirabayashi, S.; Tanabe, H.; Takano, N.; Iwata, N.; Niwa, Y.; et al. PAX5 gene as a novel methylation marker that predicts both clinical outcome and cisplatin sensitivity in esophageal squamous cell carcinoma. Epigenetics 2017, 12, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, T.; Zouridis, H.; Wu, Y.; Cheng, L.L.; Tan, I.B.; Gopalakrishnan, V.; Ooi, C.H.; Lee, J.; Qin, L.; Wu, J.; et al. Integrated epigenomics identifies BMP4 as a modulator of cisplatin sensitivity in gastric cancer. Gut 2013, 62, 22–33. [Google Scholar] [CrossRef]
- Maeda, O.; Ando, T.; Ohmiya, N.; Ishiguro, K.; Watanabe, O.; Miyahara, R.; Hibi, Y.; Nagai, T.; Yamada, K.; Goto, H. Alteration of gene expression and DNA methylation in drug-resistant gastric cancer. Oncol. Rep. 2014, 31, 1883–1890. [Google Scholar] [CrossRef] [Green Version]
- Subhash, V.V.; Tan, S.H.; Tan, W.L.; Yeo, M.S.; Xie, C.; Wong, F.Y.; Kiat, Z.Y.; Lim, R.; Yong, W.P. GTSE1 expression represses apoptotic signaling and confers cisplatin resistance in gastric cancer cells. BMC Cancer 2015, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Wang, L.; Wu, L.; Zhang, L.; Nie, G.; Guo, M. Methylation of SLFN11 promotes gastric cancer growth and increases gastric cancer cell resistance to cisplatin. J. Cancer 2019, 10, 6124–6134. [Google Scholar] [CrossRef]
- Wermann, H.; Stoop, H.; Gillis, A.J.; Honecker, F.; van Gurp, R.J.; Ammerpohl, O.; Richter, J.; Oosterhuis, J.W.; Bokemeyer, C.; Looijenga, L.H. Global DNA methylation in fetal human germ cells and germ cell tumours: Association with differentiation and cisplatin resistance. J. Pathol. 2010, 221, 433–442. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Shao, C.-J.; Chen, F.-R.; Kwan, A.-L.; Chen, Z.-P. Role of ERCC1 promoter hypermethylation in drug resistance to cisplatin in human gliomas. Int. J. Cancer 2010, 126, 1944–1954. [Google Scholar] [CrossRef]
- Fujiyoshi, S.; Honda, S.; Minato, M.; Ara, M.; Suzuki, H.; Hiyama, E.; Taketomi, A. Hypermethylation of CSF3R is a novel cisplatin resistance marker and predictor of response to postoperative chemotherapy in hepatoblastoma. Hepatol. Res. 2020, 50, 598–606. [Google Scholar] [CrossRef]
- Chen, B.; Rao, X.; House, M.G.; Nephew, K.P.; Cullen, K.J.; Guo, Z. GPx3 promoter hypermethylation is a frequent event in human cancer and is associated with tumorigenesis and chemotherapy response. Cancer Lett. 2011, 309, 37–45. [Google Scholar] [CrossRef]
- Chen, B.; Chen, J.; House, M.G.; Cullen, K.J.; Nephew, K.P.; Guo, Z. Role of neurofilament light polypeptide in head and neck cancer chemoresistance. Mol. Cancer Res. 2012, 10, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Viet, C.T.; Dang, D.; Achdjian, S.; Ye, Y.; Katz, S.G.; Schmidt, B.L. Decitabine Rescues Cisplatin Resistance in Head and Neck Squamous Cell Carcinoma. PLoS ONE 2014, 9, e112880. [Google Scholar] [CrossRef]
- Faller, W.J.; Rafferty, M.; Hegarty, S.; Gremel, G.; Ryan, D.; Fraga, M.F.; Esteller, M.; Dervan, P.A.; Gallagher, W.M. Metallothionein 1E is methylated in malignant melanoma and increases sensitivity to cisplatin-induced apoptosis. Melanoma Res. 2010, 20, 392–400. [Google Scholar] [CrossRef]
- Shen, L.; Kondo, Y.; Ahmed, S.; Boumber, Y.; Konishi, K.; Guo, Y.; Chen, X.; Vilaythong, J.N.; Issa, J.P. Drug sensitivity prediction by CpG island methylation profile in the NCI-60 cancer cell line panel. Cancer Res. 2007, 67, 11335–11343. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Song, J.; Lai, Q.; Liu, B.; Wang, H.; Xu, Y.; Feng, X.; Sun, X.; Du, Z. Hypermethylation of ATP-binding cassette B1 (ABCB1) multidrug resistance 1 (MDR1) is associated with cisplatin resistance in the A549 lung adenocarcinoma cell line. Int. J. Exp. Pathol. 2016, 97, 412–421. [Google Scholar] [CrossRef]
- Wu, J.-E.; Wu, Y.-Y.; Tung, C.-H.; Tsai, Y.-T.; Chen, H.-Y.; Chen, Y.-L.; Hong, T.-M. DNA methylation maintains the CLDN1-EPHB6-SLUG axis to enhance chemotherapeutic efficacy and inhibit lung cancer progression. Theranostics 2020, 10, 8903–8923. [Google Scholar] [CrossRef]
- Zhao, J.; Xue, X.; Fu, W.; Dai, L.; Jiang, Z.; Zhong, S.; Deng, B.; Yin, J. Epigenetic activation of FOXF1 confers cancer stem cell properties to cisplatinresistant nonsmall cell lung cancer. Int. J. Oncol. 2020, 56, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- Ibanez de Caceres, I.; Cortes-Sempere, M.; Moratilla, C.; Machado-Pinilla, R.; Rodriguez-Fanjul, V.; Manguán-García, C.; Cejas, P.; López-Ríos, F.; Paz-Ares, L.; de CastroCarpeño, J.; et al. IGFBP-3 hypermethylation-derived deficiency mediates cisplatin resistance in non-small-cell lung cancer. Oncogene 2010, 29, 1681–1690. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, P.; Zhang, L.; Tessema, M.; Bai, L.; Xu, X.; Li, Q.; Zheng, X.; Saxton, B.; Chen, W.; et al. Epigenetic Regulation of RIP3 Suppresses Necroptosis and Increases Resistance to Chemotherapy in NonSmall Cell Lung Cancer. Transl. Oncol. 2020, 13, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-W.; Zheng, Y.; Wang, J.-Z.; Lu, X.-X.; Wang, Z.; Chen, L.-B.; Guan, X.-X.; Tong, J.-D. Integrated analysis of DNA methylation and mRNA expression profiling reveals candidate genes associated with cisplatin resistance in non-small cell lung cancer. Epigenetics 2014, 9, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Li, X.; Li, S.; Zhou, S.; Zhou, Q. Cisplatin-induced downregulation of SOX1 increases drug resistance by activating autophagy in non-small cell lung cancer cell. Biochem. Biophys. Res. Commun. 2013, 439, 187–190. [Google Scholar] [CrossRef]
- Park, K.S.; Kim, H.K.; Lee, J.H.; Choi, Y.B.; Park, S.Y.; Yang, S.H.; Kim, S.Y.; Hong, K.M. Transglutaminase 2 as a cisplatin resistance marker in non-small cell lung cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 493–502. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Yang, W.; Qin, X.; Wang, F.; Li, H.; Lin, C.; Li, W.; Gu, C.; Zhang, Y.; Ran, Y. ECRG4 acts as a tumor suppressor and as a determinant of chemotherapy resistance in human nasopharyngeal carcinoma. Cell. Oncol. 2015, 38, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Coley, H.M.; Safuwan, N.A.; Chivers, P.; Papacharalbous, E.; Giannopoulos, T.; Butler-Manuel, S.; Madhuri, K.; Lovell, D.P.; Crook, T. The cyclin-dependent kinase inhibitor p57(Kip2) is epigenetically regulated in carboplatin resistance and results in collateral sensitivity to the CDK inhibitor seliciclib in ovarian cancer. Br. J. Cancer 2012, 106, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Zeller, C.; Dai, W.; Steele, N.L.; Siddiq, A.; Walley, A.J.; Wilhelm-Benartzi, C.S.M.; Rizzo, S.; Van Der Zee, A.; Plumb, J.A.; Brown, R. Candidate DNA methylation drivers of acquired cisplatin resistance in ovarian cancer identified by methylome and expression profiling. Oncogene 2012, 31, 4567–4576. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, L.J.; Smith, P.R.; Hiller, L.; Szlosarek, P.W.; Kimberley, C.; Sehouli, J.; Koensgen, D.; Mustea, A.; Schmid, P.; Crook, T. Epigenetic silencing of argininosuccinate synthetase confers resistance to platinum-induced cell death but collateral sensitivity to arginine auxotrophy in ovarian cancer. Int. J. Cancer 2009, 125, 1454–1463. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Zhang, J.R.; Li, S.D.; He, Y.Y.; Yang, Y.X.; Liu, X.L.; Wan, X.P. Aberrant methylation of breast and ovarian cancer susceptibility gene 1 in chemosensitive human ovarian cancer cells does not involve the phosphatidylinositol 3′-kinase-Akt pathway. Cancer Sci. 2010, 101, 1618–1623. [Google Scholar] [CrossRef]
- Chou, J.-L.; Su, H.-Y.; Chen, L.-Y.; Liao, Y.-P.; Hartman-Frey, C.; Lai, Y.-H.; Yang, H.-W.; Deatherage, D.E.; Kuo, C.-T.; Huang, Y.-W.; et al. Promoter hypermethylation of FBXO32, a novel TGF-β/SMAD4 target gene and tumor suppressor, is associated with poor prognosis in human ovarian cancer. Lab. Investig. 2010, 90, 414–425. [Google Scholar] [CrossRef]
- Soto, J.A.; Rodríguez-Antolín, C.; Vera, O.; Pernía, O.; Esteban-Rodríguez, I.; Dolores Diestro, M.; Benitez, J.; Sánchez-Cabo, F.; Alvarez, R.; De Castro, J.; et al. Transcriptional epigenetic regulation of Fkbp1/Pax9 genes is associated with impaired sensitivity to platinum treatment in ovarian cancer. Clin. Epigenet. 2021, 13, 167. [Google Scholar] [CrossRef]
- Strathdee, G.; MacKean, M.J.; Illand, M.; Brown, R. A role for methylation of the hMLH1 promoter in loss of hMLH1 expression and drug resistance in ovarian cancer. Oncogene 1999, 18, 2335–2341. [Google Scholar] [CrossRef] [Green Version]
- Ha, Y.N.; Sung, H.Y.; Yang, S.D.; Chae, Y.J.; Ju, W.; Ahn, J.H. Epigenetic modification of alpha-N-acetylgalactosaminidase enhances cisplatin resistance in ovarian cancer. Korean J. Physiol. Pharm. 2018, 22, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.D.; Ahn, S.H.; Kim, J.I. 3-Oxoacid CoA transferase 1 as a therapeutic target gene for cisplatin-resistant ovarian cancer. Oncol. Lett. 2018, 15, 2611–2618. [Google Scholar] [CrossRef] [PubMed]
- Su, H.Y.; Lai, H.C.; Lin, Y.W.; Liu, C.Y.; Chen, C.K.; Chou, Y.C.; Lin, S.P.; Lin, W.C.; Lee, H.Y.; Yu, M.H. Epigenetic silencing of SFRP5 is related to malignant phenotype and chemoresistance of ovarian cancer through Wnt signaling pathway. Int. J. Cancer 2010, 127, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Nogales, V.; Reinhold, W.C.; Varma, S.; Martinez-Cardus, A.; Moutinho, C.; Moran, S.; Heyn, H.; Sebio, A.; Barnadas, A.; Pommier, Y.; et al. Epigenetic inactivation of the putative DNA/RNA helicase SLFN11 in human cancer confers resistance to platinum drugs. Oncotarget 2016, 7, 3084–3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Leon, M.; Cardenas, H.; Vieth, E.; Emerson, R.; Segar, M.; Liu, Y.; Nephew, K.; Matei, D. Transmembrane protein 88 (TMEM88) promoter hypomethylation is associated with platinum resistance in ovarian cancer. Gynecol. Oncol. 2016, 142, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Kritsch, D.; Hoffmann, F.; Steinbach, D.; Jansen, L.; Mary Photini, S.; Gajda, M.; Mosig, A.S.; Sonnemann, J.; Peters, S.; Melnikova, M.; et al. Tribbles 2 mediates cisplatin sensitivity and DNA damage response in epithelial ovarian cancer. Int. J. Cancer 2017, 141, 1600–1614. [Google Scholar] [CrossRef]
- Xu, J.; Sun, T.; Guo, X.; Wang, Y.; Jing, M. Estrogen receptor-α promoter methylation is a biomarker for outcome prediction of cisplatin resistance in triple-negative breast cancer. Oncol. Lett. 2017, 15, 2855–2862. [Google Scholar] [CrossRef] [Green Version]
- Bunch, B.; Krishnan, N.; Greenspan, R.D.; Ramakrishnan, S.; Attwood, K.; Yan, L.; Qi, Q.; Wang, D.; Morrison, C.; Omilian, A.; et al. TAp73 expression and P1 promoter methylation, a potential marker for chemoresponsiveness to cisplatin therapy and survival in muscle-invasive bladder cancer (MIBC). Cell Cycle 2019, 18, 2055–2066. [Google Scholar] [CrossRef]
- Ebert, M.P.; Tanzer, M.; Balluff, B.; Burgermeister, E.; Kretzschmar, A.K.; Hughes, D.J.; Tetzner, R.; Lofton-Day, C.; Rosenberg, R.; Reinacher-Schick, A.C.; et al. TFAP2E-DKK4 and chemoresistance in colorectal cancer. N. Engl. J. Med. 2012, 366, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yang, Y.; Lu, Y.; Herman, J.G.; Brock, M.V.; Zhao, P.; Guo, M. Predictive value of CHFR and MLH1 methylation in human gastric cancer. Gastric Cancer 2015, 18, 280–287. [Google Scholar] [CrossRef]
- Koul, S.; McKiernan, J.M.; Narayan, G.; Houldsworth, J.; Bacik, J.; Dobrzynski, D.L.; Assaad, A.M.; Mansukhani, M.; Reuter, V.E.; Bosl, G.J.; et al. Role of promoter hypermethylation in Cisplatin treatment response of male germ cell tumors. Mol. Cancer 2004, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Sempere, M.; de Miguel, M.P.; Pernía, O.; Rodriguez, C.; de Castro Carpeño, J.; Nistal, M.; Conde, E.; López-Ríos, F.; Belda-Iniesta, C.; Perona, R.; et al. IGFBP-3 methylation-derived deficiency mediates the resistance to cisplatin through the activation of the IGFIR/Akt pathway in non-small cell lung cancer. Oncogene 2013, 32, 1274–1283. [Google Scholar] [CrossRef] [Green Version]
- Grasse, S.; Lienhard, M.; Frese, S.; Kerick, M.; Steinbach, A.; Grimm, C.; Hussong, M.; Rolff, J.; Becker, M.; Dreher, F.; et al. Epigenomic profiling of non-small cell lung cancer xenografts uncover LRP12 DNA methylation as predictive biomarker for carboplatin resistance. Genome Med. 2018, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Yan, L.; Xiao-Fei, L.; Hai-Yan, S.; Juan, C.; Shan, K. Hypermethylation of mismatch repair gene hMSH2 associates with platinum-resistant disease in epithelial ovarian cancer. Clin. Epigenet. 2019, 11, 153. [Google Scholar] [CrossRef]
- Chan, D.W.; Lam, W.-Y.; Chen, F.; Yung, M.M.H.; Chan, Y.-S.; Chan, W.-S.; He, F.; Liu, S.S.; Chan, K.K.L.; Li, B.; et al. Genome-wide DNA methylome analysis identifies methylation signatures associated with survival and drug resistance of ovarian cancers. Clin. Epigenet. 2021, 13, 142. [Google Scholar] [CrossRef]
- Li, X.F.; Sun, H.Y.; Hua, T.; Zhang, H.B.; Tian, Y.J.; Li, Y.; Kang, S. Promoter Methylation of the MGRN1 Gene Predicts Prognosis and Response to Chemotherapy of High-Grade Serous Ovarian Cancer Patients. Front. Oncol. 2021, 11, 659254. [Google Scholar] [CrossRef]
- Tomar, T.; Alkema, N.G.; Schreuder, L.; Meersma, G.J.; de Meyer, T.; van Criekinge, W.; Klip, H.G.; Fiegl, H.; van Nieuwenhuysen, E.; Vergote, I.; et al. Methylome analysis of extreme chemoresponsive patients identifies novel markers of platinum sensitivity in high-grade serous ovarian cancer. BMC Med. 2017, 15, 116. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.S.; Teaberry, V.S.; Bland, A.E.; Huang, Z.; Whitaker, R.S.; Baba, T.; Fujii, S.; Secord, A.A.; Berchuck, A.; Murphy, S.K. Elevated MAL expression is accompanied by promoter hypomethylation and platinum resistance in epithelial ovarian cancer. Int. J. Cancer 2010, 126, 1378–1389. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, X.-X.; Li, M.-C.; Cao, C.-H.; Wan, D.-Y.; Xi, B.-X.; Tan, J.-H.; Wang, J.; Yang, Z.-Y.; Feng, X.-X.; et al. C/EBPβ enhances platinum resistance of ovarian cancer cells by reprogramming H3K79 methylation. Nat. Commun. 2018, 9, 1739. [Google Scholar] [CrossRef]
- Senga, S.S.; Grose, R.P. Hallmarks of cancer—the new testament. Open Biol. 2021, 11, 200358. [Google Scholar] [CrossRef]
- Flanagan, J.M.; Wilson, A.; Koo, C.; Masrour, N.; Gallon, J.; Loomis, E.; Flower, K.; Wilhelm-Benartzi, C.; Hergovich, A.; Cunnea, P.; et al. Platinum-Based Chemotherapy Induces Methylation Changes in Blood DNA Associated with Overall Survival in Patients with Ovarian Cancer. Clin. Cancer Res. 2017, 23, 2213–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawant, A.; Kothandapani, A.; Zhitkovich, A.; Sobol, R.W.; Patrick, S.M. Role of mismatch repair proteins in the processing of cisplatin interstrand cross-links. DNA Repair. 2015, 35, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- No, J.H.; Kim, Y.B.; Song, Y.S. Targeting nrf2 signaling to combat chemoresistance. J. Cancer Prev. 2014, 19, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Wu, R.; Guo, Y.; Kong, A.N. Regulation of Keap1-Nrf2 signaling: The role of epigenetics. Curr. Opin. Toxicol. 2016, 1, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Tossetta, G.; Fantone, S.; Montanari, E.; Marzioni, D.; Goteri, G. Role of NRF2 in Ovarian Cancer. Antioxidants 2022, 11, 663. [Google Scholar] [CrossRef]
- Kilgour, E.; Rothwell, D.G.; Brady, G.; Dive, C. Liquid Biopsy-Based Biomarkers of Treatment Response and Resistance. Cancer Cell 2020, 37, 485–495. [Google Scholar] [CrossRef]
- Takahashi, T.; Matsuda, Y.; Yamashita, S.; Hattori, N.; Kushima, R.; Lee, Y.-C.; Igaki, H.; Tachimori, Y.; Nagino, M.; Ushijima, T. Estimation of the Fraction of Cancer Cells in a Tumor DNA Sample Using DNA Methylation. PLoS ONE 2013, 8, e82302. [Google Scholar] [CrossRef]
- Gupta, S.; Venkatesh, A.; Ray, S.; Srivastava, S. Challenges and prospects for biomarker research: A current perspective from the developing world. Biochim. Biophys. Acta 2014, 1844, 899–908. [Google Scholar] [CrossRef]
- McDermott, J.E.; Wang, J.; Mitchell, H.; Webb-Robertson, B.-J.; Hafen, R.; Ramey, J.; Rodland, K.D. Challenges in biomarker discovery: Combining expert insights with statistical analysis of complex omics data. Expert Opin. Med. Diagn. 2013, 7, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Ertel, A.; Verghese, A.; Byers, S.W.; Ochs, M.; Tozeren, A. Pathway-specific differences between tumor cell lines and normal and tumor tissue cells. Mol. Cancer 2006, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.D.; Li, L.; Wang, Y.; Thoburn, C.; Afsari, B.; Danilova, L.; Douville, C.; Javed, A.A.; Wong, F.; Mattox, A.; et al. Detection and localization of surgically resectable cancers with a multi-analyte blood test. Science 2018, 359, 926–930. [Google Scholar] [CrossRef] [Green Version]
- Alix-Panabières, C.; Pantel, K. Clinical Applications of Circulating Tumor Cells and Circulating Tumor DNA as Liquid Biopsy. Cancer Discov. 2016, 6, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Workman, P.; Aboagye, E.O.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.J.; Double, J.A.; Everitt, J.; Farningham, D.A.H.; Glennie, M.J.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1577. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zheng, W.; Wang, H.; Cheng, Y.; Fang, Y.; Wu, F.; Sun, G.; Sun, G.; Lv, C.; Hui, B. Application of Animal Models in Cancer Research: Recent Progress and Future Prospects. Cancer Manag. Res. 2021, 13, 2455–2475. [Google Scholar] [CrossRef]
- Mai, A.; Altucci, L. Epi-drugs to fight cancer: From chemistry to cancer treatment, the road ahead. Int. J. Biochem. Cell Biol. 2009, 41, 199–213. [Google Scholar] [CrossRef]
- Nebbioso, A.; Carafa, V.; Benedetti, R.; Altucci, L. Trials with ‘epigenetic’ drugs: An update. Mol. Oncol. 2012, 6, 657–682. [Google Scholar] [CrossRef] [Green Version]
- Matulonis, U.; Berlin, S.; Lee, H.; Whalen, C.; Obermayer, E.; Penson, R.; Liu, J.; Campos, S.; Krasner, C.; Horowitz, N. Phase I study of combination of vorinostat, carboplatin, and gemcitabine in women with recurrent, platinum-sensitive epithelial ovarian, fallopian tube, or peritoneal cancer. Cancer Chemother. Pharmacol. 2015, 76, 417–423. [Google Scholar] [CrossRef]
- Crabb, S.J.; Danson, S.; Catto, J.W.F.; Hussain, S.; Chan, D.; Dunkley, D.; Downs, N.; Marwood, E.; Day, L.; Saunders, G.; et al. Phase I Trial of DNA Methyltransferase Inhibitor Guadecitabine Combined with Cisplatin and Gemcitabine for Solid Malignancies Including Urothelial Carcinoma (SPIRE). Clin. Cancer Res. 2021, 27, 1882–1892. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene(s) | Platinum Compound | Tumor Model | Cell Line(s) | Methylation Detection Method | Key Findings | Ref. |
|---|---|---|---|---|---|---|
| TLX3 | CDDP | BLCA | T24 and KK27 | RLGS; COBRA; bisulfite sequencing | TLX3 is involved in BLCA cell proliferation. TLX3 gene promoter is hypermethylated in CDDP-resistant BLCA cell lines and hypomethylated in sensitive cells. TLX3 methylation status in CDDP-resistant cells is 78.6%. | [73] |
| HOXA9 | CDDP | BLCA (MIBC) | BC-3C, 647V, JO’N, BFTC-905, UM-UC14, RT4, 97-1, and 96-1 | EpiTYPER™ assay | HOXA9 promoter methylation status is related to response to CDDP-based chemotherapy in BLCA cell lines and metastatic BLCA. Demethylating agent-induced in vitro sensitization in resistant BLCA cell lines. | [74] |
| GULP1 | CDDP | BLCA (UC) | SW780, UM-UC-3, BFTC909, RT4, 5637, BFTC905, HT1376, J82, T24, and HUC-1 | MSP | CDDP-resistant T24 cell line discloses reduced endogenous GULP1 expression. These cells have longer survival in response to CDDP, indicating a possible association between GULP1 silencing and CDDP resistance. | [75] |
| Casp8AP2 [A] | Oxaliplatin | CC | SiHa and S3 | Differential methylation hybridization (DMH) microarray; qMSP; restriction with methylation-sensitive enzymes | There are global and individual loci methylation changes in resistant cells. Expression of Casp8AP2 in oxaliplatin-resistant cells was reduced and associated with increased promoter methylation. After exposure to the demethylating agent, the sensitivity of resistant S3 cells was restored to the same level as in untreated SiHa cells. | [76] |
| GPx3 | CDDP, oxaliplatin | CRC | RKO, SW48, LOVO, HCT116, SW480, SW620, COLO205, CACO2, and HT29 | MSP | Cell lines tested showed different sensitivity to CDDP, and MSP analysis disclosed a correlation between GPx3 methylation status and mRNA expression levels. Cell lines with GPx3 promoter methylation and downregulation had increased sensitivity to platinum. | [77] |
| PARPBP | Oxaliplatin | CRC | HCT116 and HCT116L | Bisulfite sequencing | The methylation level of PARPBP promoter is decreased in the resistant cell line. A mechanism in resistant CRC cells was suggested, with KIF18b inhibiting the interaction between SP1 and DNMT3b through binding to SP1, resulting in hypomethylation of PARPBP promoter and consequent promotion of PARPBP expression. Then PARPBP promoted PARP1 to enhance DNA repair in oxaliplatin-resistant cells. | [78] |
| SLFN11 | CDDP | CRC | RKO, DLD1, SW620, LOVO, Ls180, and DKO | MSP; bisulfite sequencing | The expression of SLFN11 is silenced by DNA methylation in CRC cell lines. SLFN11 suppresses CRC cell proliferation and promotes chemosensitivity of CRC cells to CDDP in vitro. | [79] |
| hMLH1 [R] | CDDP | EC | EC9706, EC1, EC9706-DDP, and EC1-DDP | MSP | hMLH1 methylation in cell lines significantly increased after the acquisition of CDDP resistance. Resistance was reversed by exposure to a demethylating agent. | [72] |
| CIDEA [A] | CDDP | ESCC | KYSE30, KYSE140, KYSE150, KYSE180, KYSE410, KYSE510, and EC109 | Bisulfite sequencing; MSP | KYSE410 cells with upregulated CIDEA disclose lower promoter methylation levels compared to KYSE30 and KYSE150 cells with downregulation of CIDEA. The 5-Aza-dC treatment restored the cellular expression of CIDEA. Gene downregulation was associated with promoter hypermethylation and the introduction of CIDEA enhanced sensitivity to CDDP. | [80] |
| FGF5 | CDDP | ESCC | KYSE-30, 50, 140, 170, 180, 220, 270, 410, 450, 510, 520, and TE-15 | Infinium® HumanMethylation450K BeadChip; bisulfite sequencing | FGF5 methylation is associated with response to chemoradiotherapy with CDDP. FGF5 expression was induced by CDDP treatment in three unmethylated cell lines, but not in two methylated cell lines. Exogenous FGF5 overexpression in a cell line with FGF5 promoter methylation conferred resistance to CDDP. | [81] |
| PAX5 | CDDP | ESCC | NUEC1 and TE3 | qMSP | PAX5-silenced cells showed relatively higher cell proliferation and cell cycle promotion, suggesting acquisition of CDDP resistance due to methylation-associated gene silencing. | [82] |
| BMP4 | CDDP | GC | AGS, Kato III, Hs746T, FU97, Ist1, MKN1, MKN7, MKN4, MKN28, MKN45, IM95, TMK1, AZ521, SCH, YCC3, YCC7, YCC10, YCC11, and YCC16 | Bisulfite sequencing; MSP | Bisulfite sequencing of the BMP4 199 region confirmed that all five CpG sites within the region were fully methylated in CDDP-sensitive lines (YCC10, YCC11, YCC16, FU97) but unmethylated in CDDP-resistant cell lines (MKN45, AZ521, Kato III). BMP4 was found methylated in sensitive, but not in resistant cells. | [83] |
| CPT1C, KLK13, ETV7, FSCN1, NOTCH3 | CDDP | GC | Wild-type AGS and CDDP-resistant AGS | Infinium® HumanMethylation450K BeadChip; bisulfite pyrosequencing | Expression of KLK13, ETV7, FSCN1, CPT1C, and NOTCH3 before and after CDDP chemotherapy differed due to promoter methylation. These alterations may be associated with mechanisms of GC drug resistance and may be used as biomarkers to predict drug sensitivity. | [84] |
| GTSE1 | CDDP | GC | AZ521, OCUM-1, SNU610, and SNU719 | MSP | All hypomethylated cell lines depicted higher GTSE1 expression. Loss of GTSE1 expression significantly enhanced sensitivity to CDDP treatment as shown by a ~5-fold decrease in IC50 values in AZ521-kd cells. GTSE1 knockdown in GC cells disclosed it as the major cause of CDDP resistance. | [85] |
| SLFN11 | CDDP | GC | NUGC3, SNU5, SNU16, PHM82, NCI-N87, BGC823, MCG803, and AGS | MSP; bisulfite sequencing | SLFN11 loss of expression was associated with promoter hypermethylation. Seven out of eight cell lines expressed SLFN11 and the promoter region was methylated. SLFN11 re-expression suppressed proliferation in SNU16 and MGC803 cell lines and sensitized cells to CDDP. | [86] |
| CFLAR [A], ERBB2, KLF11 | CDDP | GCT | TCam-2 | HumanMethylation27 DNA Analysis BeadChip (high-throughput methylation profiling) | Global methylation changes were determinant of the acquisition of resistance to CDDP, but methylation of some genes (CFLAR, ERBB2, KLF11) stood as the most promising markers to predict drug resistance. | [87] |
| ERCC1 [R] | CDDP | Glioma | T98-G, UW28, MGR1, MGR2, and SF767 | Genomic DNA methylation sequencing; MSP; real-time MSP | CDDP-sensitive MGR2 and SF767 cell lines disclosed methylation of ERCC1 promoter CpG island (5.4 Kb upstream). CDDP inhibition rate was slightly reduced and CDDP killing efficiency was lower. | [88] |
| CSF3R | CDDP | HBL | HuH6 (wild-type and CDDP-resistant variant) | Infinium® HumanMethylation450K BeadChip; bisulfite pyrosequencing | CSF3R was upregulated in CDDP-resistant cells after CDDP exposure compared to CDDP-sensitive cells. It was associated with methylation status. | [89] |
| GPx3 | CDDP | HNC | PCI13, HN17B, HN22A, SCC25, SCC25cp, HN38, PCI51, FaDu, O11, and O12 | Bisulfite DNA sequencing; MSP | Cell lines with GPx3 promoter methylation depicted gene expression downregulation or total silencing. In GPx3 methylated cells, 5aza-dC restored gene expression. CDDP-resistant and -sensitive cells significantly differed in GPx3 promoter methylation levels. There was complete or partial GPx3 methylation in 85% of CDDP-resistant HNC cells. | [90] |
| NEFL | CDDP | HNC | HaCaT, PCI13, O29, HN17B, HN22A, O12, HN38, O13, SCC25, O11, O22, PCI51, FaDu, SCC25cp, HN17Bcp, and O28 | MSP | NEFL expression was observed in all CDDP-sensitive HNC cell lines and NEFL expression was absent or greatly reduced in all five cell lines displaying the highest level of CDDP resistance and in 2/5 of moderately resistant cell lines. The other 3 moderately sensitive lines showed high NEFL expression. | [91] |
| CRIP1, G0S2, MLH1 [R], OPN3, S100, TUBB2A | CDDP | HNSCC | SCC-25 and SCC-25/CP | Methylight PCR | Methylation of these genes is associated with CDDP resistance. Decitabine treatment restored CDDP sensitivity in SCC-25/CP cells and significantly reduced the dose of CDDP required to induce apoptosis (sensitivity 67%, specificity 100%). | [92] |
| MT1E | CDDP | Melanoma | WM793, WM793-P1, WM793-P2, and 1205Lu | Bisulfite sequencing; MSP | MT1E promoter methylation is common in human melanoma and might be considered a biomarker. Gene silencing was suggested to play a role in the resistance of melanoma to chemotherapy. | [93] |
| p73 [A] | CDDP, carboplatin | Multiple tumor types (CNS cancer, CRC, leukemia, melanoma, NSCLC, OC, PC, BC, RCC) | NCI-60 (panel of 58 cancer cell lines) | COBRA; MSP | A functional link between p73 and alkylating agent (CDDP) sensitivity was confirmed, as in several cancer cell lines tested; downregulation of p73 increased sensitivity to commonly used alkylating agents (CDDP and carboplatin). | [94] |
| ABCB1 [T] | CDDP | LC | A549, A549/DDP | Bisulfite sequencing | ABCB1 promoter methylation levels are significantly higher in CDDP-resistant cells compared to A549 cells. | [95] |
| CLDN1 | CDDP | LC | CL1-0 and CL1-5 | Bisulfite sequencing; MSP; pyrosequencing of CpG regions | CLDN1 represses cancer progression via a feedback loop involving the CLDN1-EPHB6-ERK1/2-SLUG axis, which represses drug resistance and sensitizes lung adenocarcinoma cells to chemotherapy. DNA methylation maintains CLDN1 expression. As CLDN1 expression improves the efficacy of chemotherapy, it might constitute a biomarker predictive of response to chemotherapy. | [96] |
| FOXF1 | CDDP | NSCLC | A549, A549/DDP, H1299, and 16HBE | Infinium® HumanMethylation450K BeadChip; pyrosequencing | FOXF1 promoter methylation levels are decreased in CDDP-resistant cells. FOXF1 overexpression decreased CDDP-induced apoptosis of sensitive cells and FOXF1 knockdown increased apoptosis of resistant cells. | [97] |
| IGFBP-3 | CDDP | NSCLC | H23R, H460R, and 41R | Bisulfite sequencing | IGFBP-3 is silenced by promoter hypermethylation in 41R and H23R-resistant cells compared with their parental sensitive cell lines, with marked IGFBP-3 basal expression. | [98] |
| RIP3 | CDDP | NSCLC | A549, H1568, H1299, H460, H23, H2009, H2023, H1689, HCC4006, Calu-3, and Calu-6 | COBRA; Infinium® HumanMethylation450K BeadChip | Hypermethylation of RIP3 promoter region was detected in all LC cell lines but not in primary human bronchial epithelial cells. RIP3 mRNA and protein expression increased after demethylating agent treatment in LC cell lines with methylated promoters, but not in those without methylation. Restored RIP3 expression sensitized cells to CDDP. | [99] |
| S100P, GDA, WISP2, LOXL1, TIMP4, ICAM1, CLMP, HSP8, GAS1, BMP2 | CDDP | NSCLC | A549 and A549/DDP | Infinium® HumanMethylation450K BeadChip; qMSP | All candidate genes were hypermethylated in A549/DDP cells compared with parental A549 cells. In vivo studies also showed that GAS1 downregulation by methylation was associated with CDDP resistance. | [100] |
| SOX1 | CDDP | NSCLC | A549, A549/cis, H358, and H358/cis | Bisulfite genomic sequencing | SOX1 is hypermethylated in CDDP-resistant cell lines compared to the parental cells. The expression of SOX1 was upregulated in CDDP-resistant cells after treatment with demethylating agent. SOX1 silencing enhanced CDDP-mediated autophagy in NSCLCs. | [101] |
| TGM2 | CDDP | NSCLC | HCC-95, HCC-1588, NCI-H23, HCC-1195, NCI-H1299, HCC-2279, SK-MES-1, SK-LU-1, and HCC-1171 | Bisulfite genomic sequencing | CDDP sensitivity was higher in TGM2 promoter-methylated LC cell lines (HCC-95/1588) than in non-methylated ones (NCI-H1299 and HCC-1195). TGM2 overexpression decreased sensitivity to CDDP and decreased TGM2 expression, with siRNA in non-methylated cell lines increased sensitivity to CDDP. | [102] |
| ECRG4 | CDDP | NPC | HNE1, HONE1, CNE1, SUNE1, CNE2, 6-10B, and C666-1 | Bisulfite sequencing; MSP | Demethylation with 5-aza-dC induced reactivation of methylated and silenced ECRG4 in NPC cell lines. NPC-derived cell line CNE1 was used for exogenous ECRG4 overexpression, which increased tumor cell death when exposed to cisplatin. | [103] |
| p57Kip2 | Carboplatin | OC (EOC) | PEO1, PEO1CisR, and PEO1CarbR | MSP; pyrosequencing | p57Kip2 is epigenetically downregulated in a carboplatin-resistant cell line (PEO1CarbR). MSP analysis of the CpG island located at the 5’ region of the p57Kip2 gene disclosed that methylation level was significantly higher for PEO1CarbR than for PEO1. | [104] |
| ARHGDIB, PSMB9, HSPA1A, ARMCX2, MEST, FLNC, MLH1 [R], MDK, GLUL, FLNA, NTS, COL1A1, NEFL | CDDP | OC | A2780p5, A2780p6, A2780/cp70, A2780/MCP1, A2780/MCP6, PEO1, PEO4, PEO14, PEO23, PEA1, and PEA2 | Array-based methylation profiling; pyrosequencing | Thirteen genes were consistently hypermethylated in CDDP-resistant A2780 cells; 5/13 genes (ARMCX2, COL1A1, MDK, MEST, and MLH1) acquired methylation in drug-resistant, OC-sustaining cells. MLH1 gene was found to have a direct role in conferring CDDP sensitivity when reintroduced to cells in vitro. | [105] |
| ASS1 | CDDP, carboplatin | OC | JAMA2, OVCA433, TR175, SKOV3, OVCAR3, 1847, A2780, and A2780 CisR | Bisulfite sequencing; MSP | There were methylated CpG dinucleotides in ASS1 promoter of the CDDP-resistant A2780 CisR cell line whereas the parent A2780 cell line was not methylated. When ASS1 was expressed in A2780 CisR and JAMA2 cell lines, the sensitivity to CDDP increased. | [106] |
| BRCA1 [R] | CDDP | OC | COC1, COC1/DDP, and SKOV-3 | qMSP; bisulfite genomic sequencing | CDDP-sensitive cells were found to harbor higher BRCA1 promoter methylation levels than cells with inherent and acquired resistance. Treatment of cell lines with a demethylating agent decreased sensitivity to CDDP. | [107] |
| FBXO32 | CDDP | OC | IOSE, HeyC2, SKOV3, MCP3, MCP2, A2780, and CP70 | COBRA; MSP; real-time qMSP | FBXO32 is downregulated in OC cells and its re-expression reduced tumor growth in vitro and in vivo. When restored in drug-resistant CP70 cells, FBXO32 re-sensitized cells to CDDP and enhanced apoptosis, although, in more resistant HeyC2 cells, the re-expression only caused decreased cell cycle progression. | [108] |
| FKBP1B, PAX9 | CDDP | OC | A2780 and OVCAR3 (and matched resistant variants) | Bisulfite sequencing; MSP; qMSP; whole-genome bisulfite sequencing; Infinium® HumanMethylation450K BeadChip | PAX9 and FKBP1B showed higher methylation levels in OVCAR3-resistant cell line compared to WT, control ovarian tissues, and PBMCs. There was a 4.7-fold increase in FKBP1B methylation comparing the resistant and sensitive variants of OVCAR3 and a 6-fold increase in PAX9. Moreover, FKBP1B overexpression caused increased CDDP sensitivity. | [109] |
| hMLH1 [R] | CDDP | OC | A2780 parental cell line and 10 CDDP-resistant A2780 derivative cell lines | Promoter DNA restriction with methylation-sensitive (HpaII) and methylation-insensitive (MspI) endonucleases; Southern blot analysis | hMLH1 promoter methylation was confirmed and loss of protein expression was observed. The CDDP-sensitive parental cell line was methylated only in one of the hMLH1 promoter alleles, whereas the resistant one was methylated in both alleles. | [110] |
| NAGA | CDDP | OC, NSCLC | PA-1, TOV-21G, TOV-112D, Caov-3, A2780, A2780cis, MDAH2774, ES-2, OVCAR-3, OV-90, and SK-OV-3 | Infinium® HumanMethylation450K BeadChip | NAGA mRNA downregulation correlated with specific NAGA promoter CpG site hypermethylation in CDDP-resistant OC cells. Demethylating agent restored expression and CDDP cytotoxicity increased, whereas loss of NAGA induced increased chemoresistance in sensitive and resistant cells. | [111] |
| OXCT1 | CDDP | OC | SK-OV-3, PA-1, Caov-3, TOV-21G, A2780, TOV-112D, OV-90, and OVCAR-3 | Infinium® HumanMethylation450K BeadChip | OXCT1 downregulation by hypermethylation of CGI within the promoter region is significantly higher in CDDP-resistant cell lines than in the sensitive ones. In the most resistant SKOV3 OC cell line, OXCT1 overexpression improved sensitivity to CDDP. | [112] |
| SFRP5 | CDDP | OC | SKOV3, A2780s, CP70, and OVCAR3 | MSP; bisulfite sequencing | All tested OC cell lines disclosed SFRP5 hypermethylation. Treatment with methylation inhibitor restored SFRP5 mRNA expression. Epigenetic silencing of SFRP5 affected tumor growth, invasion, tumorigenicity, and chemosensitivity of OC cells. | [113] |
| SLFN11 | CDDP, carboplatin | OC, NSCLC | SK-OV-3 and NCI-H23 | Bisulfite sequencing; Infinium® HumanMethylation450K BeadChip | When SLFN11 was downregulated by shRNA, both cell lines showed significantly increased IC50 values for platinum treatment compared to control cells, indicating a role for SLFN11 in platinum resistance. | [114] |
| TMEM88 | Carboplatin | OC | A2780 (injected in mice) | Infinium® HumanMethylation450K BeadChip | In mice injected with A2780 cells and treated with carboplatin hypomethylation of TMEM88, gene promoter in resistant tumors was observed. It was confirmed that TMEM88 mRNA expression levels are increased in resistant tumors versus controls, which is consistent with gene promoter hypomethylation in those tumors. | [115] |
| TRIB2 | CDDP | OC | A2780, SKOV3, and HeyA8 | Microarray-based methylation analysis | Analysis of TRIB2 confirmed an indirect contribution of hypermethylation to gene silencing and the functional impact of this gene on A2780 chemosensitivity. TRIB2 overexpression in resistant cells led to reduced IC50, and shRNA-mediated silencing of TRIB2 in parental sensitive A2780 cells increased their resistance. | [116] |
| Gene(s) | Platinum Compound | Tumor Model | Sample Type | Sample Grouping and Size | Patients’ Gender and Mean Age | Methylation Detection Method | Key Findings | Ref. |
|---|---|---|---|---|---|---|---|---|
| ERα | CDDP | BC (TNBC) | Tumor tissue samples | 35 patient samples | All ♀ (median age 47 y.o., range 27–69 y.o.) | MSP | Tumor samples with ERα methylation were resistant to CDDP. Furthermore, ERα methylation was related to increased BRCA1 expression, indicating a possible resistance mechanism. | [117] |
| TLX3 | CDDP | BLCA | Tumor tissue samples | 110 patient samples | n.m. | RLGS; COBRA; bisulfite sequencing | TLX3 is hypermethylated in tumors resistant to CDDP. Methylation in patient samples and cell lines was congruent, indicating a role for TLX3 as a biomarker of response to CDDP. | [73] |
| HOXA9 | CDDP | BLCA (MIBC) | Tumor tissue samples from vesical transurethral resections | 18 patient samples | 15 ♂ and 3 ♀; mean age 69 y.o. at the time of cystectomy (median 71, range 60 to 77 y.o.) | EpiTYPER™ assay | HOXA9 promoter methylation status was associated with response to CDDP-based chemotherapy in MIBC. HOXA9 promoter methylation might be used to predict sensitivity or resistance to CDDP-based chemotherapy in BLCA patients. | [74] |
| GULP1 | CDDP | BLCA (UC) | Tumor tissue and urine samples | 46 urine samples from individuals without neoplastic disease; 58 diagnosed with UCB; 20 primary tumors and matched normal samples; 76 primary tumors | n.m. | MSP | The qMSP in tumor samples showed a significantly higher frequency of GULP1 promoter methylation in tumors than in matched normal tissues. The results were confirmed in urine samples and TCGA-BLCA dataset. GULP1 might be a biomarker of resistance to CDDP. | [75] |
| p73 [A] | CDDP | BLCA (MIBC) | Tumor tissue samples | 14 patient samples (8 low and 6 high methylation) | n.m. | Infinium® HumanMethylation450K BeadChip; pyrosequencing | The p73 promoter methylation was significantly related to worse OS (high methylation: 13.5 months vs. low methylation: 30 months). The p73 promoter hypermethylation might be a predictive biomarker for CDDP response in BLCA patients. | [118] |
| SLFN11 | CDDP | CRC | Tumor tissue samples | 133 patient samples (128 primary CRC cases and 5 noncancerous colorectal mucosae) | 84 ♂ and 44 ♀ (30 < 50 y.o. and 98 ≥ 50 y.o.) | MSP; bisulfite sequencing | SLFN11 was found methylated in 55.47% of human CRC samples, regulating gene expression. SLFN11 methylation is significantly associated with age, poor 5-year OS, and RFS. | [79] |
| TFAP2E | Oxaliplatin | CRC | Tumor tissue samples | 74 patient samples (metastatic CRC) | n.m. | MethylLight | The cohort treated with oxaliplatin disclosed a negative association between methylation and treatment response: higher response rates among patients with hypomethylated TFAP2E (3/20 patients with hypermethylated TFAP2E responded to treatment, whereas 33/54 patients with hypomethylated TFAP2E responded). | [119] |
| FGF5 | CDDP | ESCC | Tumor tissue samples | 117 patient tumor samples of responders and non-responders (41 patients in screening set, 44 patients in validation set, 42 patients in re-validation set) | Screening set: 34 ♂, 7 ♀ (mean age 64.6 y.o.); validation set: 28 ♂, 6 ♀ (mean age 66.8 y.o.); re-validation set: 30 ♂, 9 ♀ (mean age 65.9 y.o.) | Infinium® HumanMethylation450K BeadChip; bisulfite sequencing | FGF5 methylation might be a biomarker predictive of sensitivity to dCRT (with CDDP). Methylome screening identified the specificity of FGF5 expression and associated promoter methylation levels with the response (45% sensitivity and 90% specificity in the combined validation and re-validation sets, n = 76). | [81] |
| PAX5 | CDDP | ESCC | Tumor tissue surgical samples | 156 ESCC patient samples (78 tumor and 78 normal adjacent) | 62 ♂ and 16 ♀, 37 ≥ 65 y.o. and 41 < 65 y.o. | qMSP | PAX5 methylation was frequent and highly tumor specific in ESCC. Methylation was significantly associated with low protein expression in tumors. PAX5 silencing correlated with increased cancer cell proliferation and CDDP resistance and might associate with poor RFS. | [82] |
| BMP4 | CDDP | GC | Tumor tissue samples | 197 patient samples | n.m. | Bisulfite sequencing; MSP | A significant correlation between BMP4 methylation status and mRNA expression was found across tumors investigated. BMP4- expressing tumors were associated with poor GC prognosis and possible resistance to CDDP. | [83] |
| MLH1 [R] | Oxaliplatin | GC | FFPE tumor tissue samples | 53 oxaliplatin-treated patient samples | 72 ♂ and 30 ♀, median age 53 y.o. | Nested MSP | In oxaliplatin-treated patients, MLH1 methylation was found in 30.2% of cases. OS was higher in the unmethylated MLH1 group vs. methylated group (p = 0.046). Patients with methylated MLH1 promoters were found to be resistant to oxaliplatin. MLH1 methylation might be an oxaliplatin-resistance biomarker in GC. | [120] |
| SLFN11 | CDDP | GC | Tumor tissue samples | 209 patient samples (201 GC samples and 8 normal gastric mucosa samples) | 157 ♂ and 44 ♀ (39 patients < 50 y.o. and 162 patients ≥ 50 y.o.) | MSP; bisulfite sequencing | SLFN11 was found methylated in 29.9% of human GC samples, and SLFN11 expression was regulated by promoter methylation. Additionally, SLFN11 methylation was significantly associated with tumor size. | [86] |
| ERCC1 [R] | CDDP | Glioma | Tumor tissue surgical samples | 32 patient samples | 1 8♂ and 14 ♀ (median age 29 y.o.) | MSP; real-time MSP | Aberrant ERCC1 promoter methylation was found in primary glioma samples. ERCC1 mRNA and protein expression levels, as well as response to CDDP in glioma, were associated with ERCC1 promoter methylation levels. | [88] |
| APC, RASSF1A [A], HIC1, BRCA1 [R], MGMT, RARB, FHIT, FANCF [R], ECAD | CDDP | Male GCT | Tumor tissue samples | 70 patient samples (31 CDDP-sensitive and 39 resistant) | n.m. | MSP | One or more genes were methylated in 59% of tested tumors. The top hypermethylated genes were RASSF1A (35.7%), HIC1 (31.9%), BRCA1 (26.1%), and APC (24.3%). RASSF1A and HIC1 inactivation by promoter hypermethylation might constitute biomarkers for platinum resistance. | [121] |
| CSF3R | CDDP | HBL | Fresh-frozen tumor samples | 43 patient samples (38 CDDP-sensitive and 5 resistant) | n.m. | Infinium® HumanMethylation450K BeadChip; bisulfite pyrosequencing | CSF3R hypermethylation was evaluated in CDDP-resistant hepatoblastoma. CSF3R hypermethylation was associated with CDDP resistance and might assist in selecting ion of HBL patients for postoperative chemotherapy. | [89] |
| GPx3 | CDDP | HNC | Frozen tumor tissue samples | 46 patient samples | 34 ♂ and 12 ♀; mean age: 43.8 ± 24.3 y.o. | Bisulfite sequencing; qMSP; MSP | 61% of tested HNC primary tumors harbored GPx3 methylation vs. only 8% of normal tissue samples. In cases with complete or partial response to chemotherapy, 82.6% of samples were not methylated and 59% of patients with no response to chemotherapy disclosed methylated GPx3. | [90] |
| NEFL | CDDP | HNC | Frozen tumor tissues | 51 patient samples (25 methylated and 26 unmethylated for NEFL) | 39 ♂ and 12 ♀ | Bisulfite DNA sequencing; MSP; real-time MSP | Patients with methylated NEFL promoter were nearly 3 times more likely to endure resistance to CDDP-based chemotherapy. NEFL methylation also predicted reduced OS and disease-free survival in HNC patients who received CDDP-based chemotherapy. | [91] |
| CRIP1, G0S2 [A], MLH1 [R], OPN3, S100, TUBB2A | CDDP | HNSCC | FFPE tumor samples | 19 patient samples (10 progressed, 2 stable, and 7 complete remission 6 months post-CDDP treatment) | 12 ♂ and 7 ♀; mean age 57.95 | Methylight PCR | The genes tested disclosed higher promoter methylation in CDDP-resistant than CDDP-sensitive tumors. Genes were assembled into a classifier, which might be used to categorize sensitivity to CDDP (67% sensitivity, 100% specificity). | [92] |
| GDA, S100P, WISP2, LOXL1, TIMP4, ICAM1, HSP8, GAS1 | CDDP | NSCLC | Primary tumor samples | 40 patient samples (20 CDDP-resistant and 20-sensitive) | n.m. | Infinium® HumanMethylation450K BeadChip platform; qMSP | The genes listed were found to disclose higher methylation levels in CDDP-resistant NSCLC samples compared to sensitive tumors. | [100] |
| IGFBP-3 | CDDP | NSCLC | Paraffin-embedded surgical specimens | 36 patient samples (19 CDDP-resistant and 17 sensitive); 10 control biopsies | 34 ♂ and 2 ♀; mean age 65.8 y.o. | Bisulfite sequencing; MSP | Most CpG dinucleotides were methylated in resistant but not in sensitive primary tumors, indicating a significant association between IGFBP-3 methylation and CDDP chemosensitivity. | [98] |
| IGFBP-3 | CDDP | NSCLC | Paraffin-embedded surgical specimens | 25 patient samples | 23 ♂ and 2 ♀; mean age 63.7 y.o. | MSP | IGFBP-3 promoter methylation and IGFIR/AKT phosphorylation occurred only in CDDP-resistant NSCLC patients. IGFBP-3 deficiency due to methylation might mediate the resistance to CDDP through activation of IGFIR/AKT pathway. | [122] |
| LRP12 | Carboplatin | NSCLC | FFPE primary tumor samples and frozen tumor tissue samples | PDX models derived from 22 NSCLC patients and validation in an independent cohort of 35 patient FFPE samples | n.m. | Me-DIP Seq; targeted bisulfite sequencing; MSP | LRP12 hypermethylation correlated with increased resistance to carboplatin. LRP12 methylation was significantly higher in patients with relapse (13.9% vs. 7.4%). A threshold of 8.3% was determined, allowing us to classify tumors into responders and non-responders to carboplatin (80% sensitivity, 84% specificity). | [123] |
| RIP3 | CDDP | NSCLC | Frozen tumor tissue samples | 16 NSCLC patients (both normal and tumor tissues) | n.m. | COBRA; Infinium® HumanMethylation450K BeadChip | The quantitative methylation data for probes located within the RIP3 promoter CpG island revealed significantly higher methylation in 25% of tumors. When RIP3 promoter was methylated, protein expression was suppressed, correlating with increased resistance to CDDP. | [99] |
| hMSH2 [R] | CDDP, carboplatin | OC (EOC) | Patient tissues | 40 patient samples (18 CDDP-resistant and 22-sensitive) | Median age 56 y.o. (years old) | RRBS; MALDI-TOF mass spectrometry | A specific promoter region containing CpGs was significantly hypermethylated in platinum-resistant patients. High hMSH2 promoter methylation levels are associated with poor prognosis in patients submitted to CDDP treatment. | [124] |
| DLG2, OR51L1, OR51I1, OR51F1, OR51B6, HBBP1, TMEM200A [T] | CDDP | OC (HGSOC) | Frozen cryosections of tumor tissue samples | 30 patient samples | 10 platinum-sensitive cases (60% did not recur in 5 years); 20 platinum-resistant cases (5% did not recur in 5 years after treatment) | Illumina 850K methylation assay | The platinum-sensitive group depicted lower methylation levels than the platinum-resistant group. In an epigenome-wide association study, differentially methylated probes helped to identify hypermethylated genes in platinum-resistant patients. | [125] |
| EGR1, MGRN1 | CDDP | OC (HGSOC) | Tumor tissue surgical samples | 96 patient samples (55 platinum-sensitive and 41 platinum-resistant) | 34 < 50 y.o. and 62 ≥ 50 y.o. | RRBS; MALDI-TOF mass spectrometry | The promoters of MGRN1, EGR1 were significantly hypermethylated in cancer tissues from platinum-resistant HGSOC. Lower MGRN1 and EGR1 expressions due to hypermethylation were associated with clinical outcomes. | [126] |
| FZD10 | CDDP | OC (HGSOC) | Frozen tumor tissue samples | 70 patient samples divided by 2 patient groups (group 1: 18 advanced-stage HGSOC samples; group 2: 21 responder and 31 non-responder samples) | Group 1: median age 61 y.o.; group 2: median age 62.25 y.o. | MethylCap-seq; bisulfite pyrosequencing | FZD10 was the most differentially methylated gene among two of the chemoresponsive-related groups. FZD10 expression was significantly lower due to promoter methylation in the extreme responder HGSOC patient group compared to the non-responder group. | [127] |
| ARHGDIB, PSMB9, HSPA1A, ARMCX2, MEST, FLNC, MLH1 [R], MDK, GLUL, FLNA, NTS, COL1A1, NEFL | CDDP | OC | Tumor tissue samples | 14 patient samples (7 matched tumor samples before chemotherapy and at relapse) | n.m. | Array-based methylation profiling; pyrosequencing | CpG sites of 9/13 genes (ARHGDIB, ARMCX2, COL1A, FLNA, FLNC, MEST, MLH1, NTS, and PSMB9) acquired methylation in relapsed ovarian tumors after chemotherapy with CDDP. | [105] |
| ASS1 | CDDP | OC | Frozen tumor tissue samples | 54 patients (treated with surgery followed by post-operative CDDP chemotherapy, tissue sample at initial diagnosis and relapse) | n.m. | MSP | In a group of patients with methylated ASS1 promoter at diagnosis, there were significantly more cases with partial clinical response, RFS < 12 months, or progressive disease; 34 patients relapsed during the study and, in 53% of them, methylation was present at diagnosis and in 74% at relapse. ASS1 methylation at diagnosis was associated with significantly reduced RFS. | [106] |
| FBXO32 | CDDP | OC | Tumor tissue surgical samples | 96 OC patient samples and 5 normal benign gynecological disease cells | Median age 52 y.o. (18 to 90 y.o.) | COBRA; MSP; real-time qMSP | High FBXO32 methylation level was significantly associated with higher stage and shorter PFS. Samples with higher FBXO32 methylation disclosed lower expression. | [108] |
| MAL | CDDP | OC | Frozen tumor tissue samples | 61 serous epithelial OC patient samples (60 III/IV stage, 26 living > 7 years, and 34 living < 3 years post-diagnosis; and 1 I/II stage cancer) | n.m. | Quantitative bisulfite sequencing; bisulfite sequencing; pyrosequencing; MSP | There was an average increase in MAL expression in III-IV stage ovarian tumors and transcript levels of short-term survivors compared to long-term survivors treated with CDDP. This was associated with CDDP resistance. | [128] |
| SFRP5 | CDDP | OC | Frozen primary tumor biopsies | 105 patient samples | n.m. | MSP; bisulfite sequencing | SFRP5 methylation status was found to correlate with CDDP resistance in OC patients. The patients with no SFRP5 methylation had a significantly better response to chemotherapy. | [113] |
| SLFN11 | CDDP, carboplatin | OC, NSCLC | Tumor tissue samples | 63 patient samples (41 in OC cohort and 22 in NSCLC cohort) | OC cohort: 5 ♀ < 50 y.o., 21 ♀ > 5 0 y.o., 15 ♀ unknown; NSCLC cohort: 10 ♂, 12 ♀ (2 < 50 y.o., 20 > 50 y.o.) | Infinium® HumanMethylation450K BeadChip; bisulfite sequencing | SLFN11 hypermethylation was associated with shorter OS and PFS. Clinical results paralleled those of cancer cell lines. | [114] |
| PAX9 | CDDP | OC | Fresh frozen and FFPE tumor tissue samples | 189 patient samples (129 FFPE and 57 frozen samples) | n.m. | Bisulfite sequencing; MSP; qMSP; whole-genome bisulfite sequencing; Infinium® HumanMethylation450K BeadChip | Kaplan–Meier analysis showed that resistant/PAX9-methylated patients had reduced OS compared to cases without methylation. Moreover, patients with low PAX9 expression disclosed shorter OS and recurrent disease. | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavares, N.T.; Gumauskaitė, S.; Lobo, J.; Jerónimo, C.; Henrique, R. DNA Methylation Biomarkers for Prediction of Response to Platinum-Based Chemotherapy: Where Do We Stand? Cancers 2022, 14, 2918. https://doi.org/10.3390/cancers14122918
Tavares NT, Gumauskaitė S, Lobo J, Jerónimo C, Henrique R. DNA Methylation Biomarkers for Prediction of Response to Platinum-Based Chemotherapy: Where Do We Stand? Cancers. 2022; 14(12):2918. https://doi.org/10.3390/cancers14122918
Chicago/Turabian StyleTavares, Nuno Tiago, Saulė Gumauskaitė, João Lobo, Carmen Jerónimo, and Rui Henrique. 2022. "DNA Methylation Biomarkers for Prediction of Response to Platinum-Based Chemotherapy: Where Do We Stand?" Cancers 14, no. 12: 2918. https://doi.org/10.3390/cancers14122918
APA StyleTavares, N. T., Gumauskaitė, S., Lobo, J., Jerónimo, C., & Henrique, R. (2022). DNA Methylation Biomarkers for Prediction of Response to Platinum-Based Chemotherapy: Where Do We Stand? Cancers, 14(12), 2918. https://doi.org/10.3390/cancers14122918