
Targeting mTOR as a Cancer Therapy: Recent Advances in Natural Bioactive Compounds and Immunotherapy

 , , , , ,
, , , , ,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
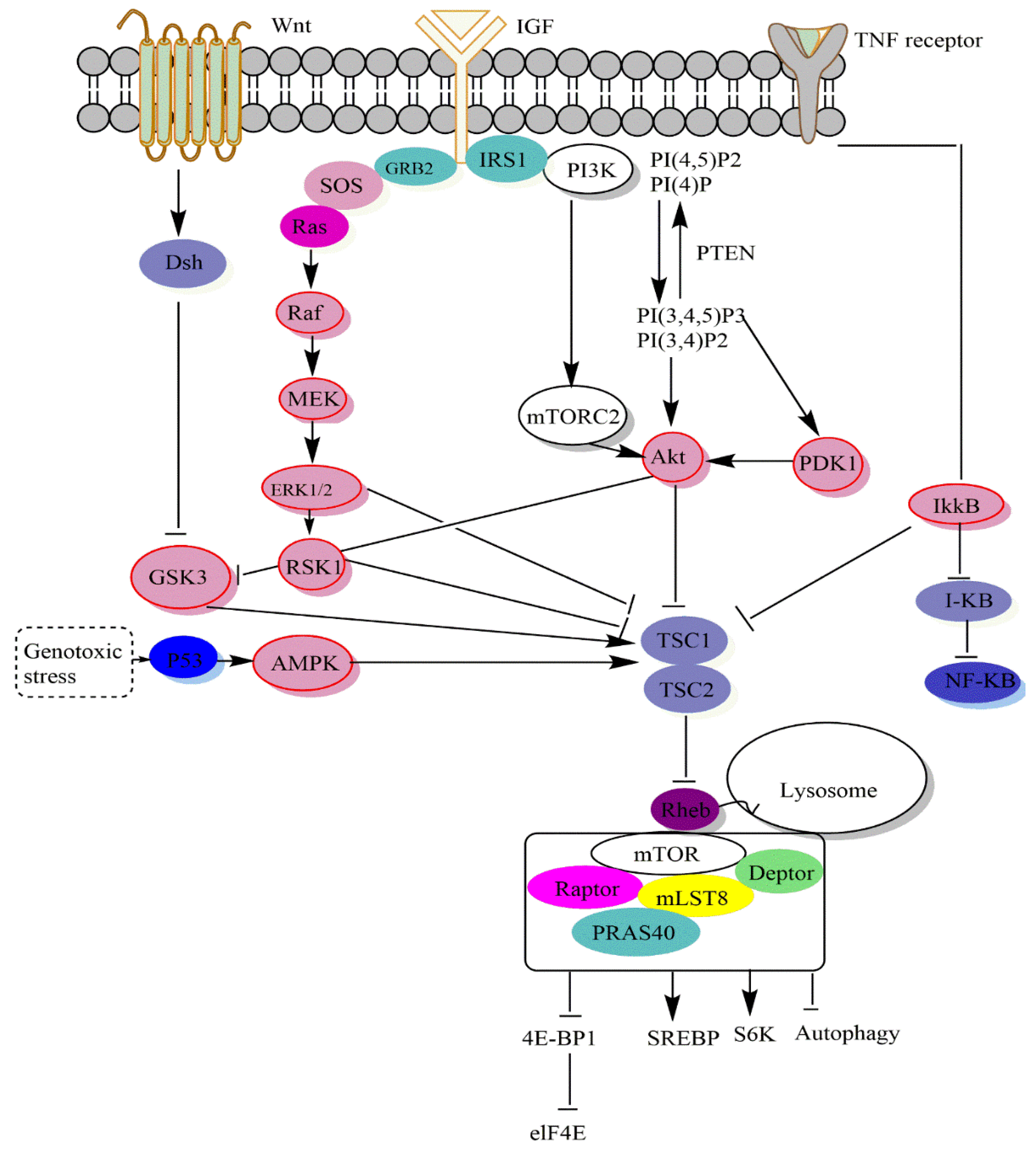
2. mTOR Signaling Pathways
2.1. Structure of mTOR
2.2. Role of mTOR
2.2.1. Role of mTORC1
2.2.2. Autophagy and mTOR
2.2.3. mTOR and Synthesis of Lipid
2.2.4. mTOR and Synthesis of Proteins
2.2.5. mTOR: A Master Regulator of mRNA Translation
2.2.6. mTOR and Biogenesis Ribosome
3. mTOR and Cancer
3.1. Signalling Upstream of mTOR
3.2. Signaling Downstream of mTOR
3.3. Breast Cancer
3.4. Lung Cancer
3.5. Colon Cancer
3.6. Head and Neck Cancer
3.7. Cervical Cancer
3.8. Reproductive Cancer
3.9. Bone Cancer
3.10. Cancers Caused by Different Type of Viruses
3.11. Nasopharyngeal Carcinoma
3.12. DLBCL
3.13. Different Type of Lymphomas
3.14. Non-Mantle Hodgkin’s Cell Lymphoma (MCL)
4. Targeting mTOR Signaling Pathways by Natural Products

{kind=link}
{kind=link}
| Compounds | Methods | Key Results | References |
|---|---|---|---|
| Apigetrin | AGS human gastric cancer cells | Induced extrinsic apoptosis Induced autophagy Caused G2/M phase cell cycle arrest through PI3K/AKT/mTOR pathway | [7] |
| Dihydromyricetin | HepG2 cells | Induced significantly autophagosome characteristics Promoted LC3-II and Beclin-1 expressions Suppressed mTOR activation Induced autophagy Downregulated the cell proliferation | [198] |
| Licochalcone A | Human cervical cancer cells | Induced mitochondria dependent apoptosis Decreased Bcl-2 expression Induced autophagy Inhibited of PI3K/Akt/ mTOR Inhibited tumor growth | [199] |
| Piperlongumine | DMH/DSS induced experimental colon cancer | Inhibited tumor cell growth Inhibited Ras and PI3K proteins levels Suppressed activity of Akt/NF-κB, c-Myc, and cyclin D1 Arrested cell cycle progression Induced apoptosis | [200] |
| Atractylodin | Cholangiocarcinoma cell line | Regulated PI3K/AKT/mTOR and p38MAPK signalling pathways Inhibited cell growth Inhibited the migration and invasion Induced autophagy Increased SB202190 and 3-MA Reduced the rate of ATD-induced autophagy Inhibited the phosphorylation of PI3K, protein kinase B/AKT, mTOR, mitogen-activated protein kinase (p38MAPK) Elevated Beclin-1 expression and LC3 conversion Decreased p-AKT/AKT, p-mTOR/mTOR, and p-p38MAPK/p38MAPK | [201] |
| Cannabisin B | HepG2 human hepatoblastoma cells | Induced cell death Induced S phase cell cycle arrest Inhibited survival signaling Blocked AKT activation Down-stream mTOR targets | [202] |
| Thymoquinone (2-isopropyl-5-methyl-1,4-benzoquinone) | Gastric cancer cells (MGC80-3 and SGC-7901) | Exhibited significant growth inhibitory effects Inhibited cell migration ability Downregulated mesenchymal gene expression Inhibition of PI3K/Akt/mTOR signalling pathway key proteins | [203] |
| Curcumin | Human renal carcinoma caki cells | Downregulated Mcl-1 protein expression Decreased Bcl-2 mRNA and protein expression Induced apoptosis | [204] |
| Gallic acid | Acute myeloid leukemia (AML) | Induced apoptosis Inhibited mitochondrial respiration Reduced ATP production and oxidative stress | [205] |
| Gartanin | Human urinary bladder cancer cell lines (T24 and RT4) | Suppressed p70S6 and 4E-BP1 expressions Inducted autophagy Downregulated Bcl-2 expression Activated the p53 pathway | [206] |
| Kavalactones yangonin | Bladder cancer cells (RT4, T24, UMUC3, HT1376, and HT 1197 cell lines) | Elevated beclin and ATG5 expression Increased LKB1 expression Decreased the phosphorylation of Akt, PRAS40, rpS6, p70S6K, and 4E-BP1 Reduced the viability of bladder cancer cell lines | [207] |
| Resveratrol | Breast cancer cell line | Downregulated cell growth Prevented mTORC1 signaling pathways Activated the autophagy pathway Modulated Akt/mTOR signaling pathway Regulated microRNA expression | [208] |
| Resveratrol | Bladder cancer cell line | Activated the autophagy pathway Regulated microRNA expression Modulated the Akt/mTOR signaling pathway | [209] |
| Glycyrrhizic acid | TF-1 leukemia cells | Induced apoptosis Blocked Akt, mTOR, and STAT3 phosphorylation signaling | [210] |
| Oleanolic acid | Cancer cell lines (A549, MCF-7, U2OS, BXPC3, PANC-1, and PC-3 cells) | Induced autophagy Activated JNK Inhibited mTOR | [211] |
| Poricoic acid A | SKOV3 ovarian cancer cells lines | Suppressed SKOV3 cellular viability, migration, and invasion Induced SKOV3 cell apoptosis Increased LC3-II/LC3-I ratio Inhibited mTOR and p70s6k phosphorylation Reduced the xenograft tumor weight | [212] |
| Echinatin | Esophageal squamous cell carcinoma (ESCC) | Inducted autophagy Inactivated the AKT/mTOR | [213] |
| Vitexin | Human renal carcinoma cell lines (OS-RC-2 and ACHN cell) | Inhibited cell growth Induced apoptosis and hyperautophagy Upregulated the AMPK/mTOR and JNK pathways Downregulated PI3K/AKT/mTOR pathways | [214] |
| Cryptotanshinone | CT26 colon cancer cell lines | Induced cell autophagy and apoptosis Inhibited PI3K-Akt-mTOR signaling pathway Diminished p-PI3K, p-Akt, and p-mTOR expressions | [215] |
| Tanshinone IIA | KBM-5 leukemia cells | Induced autophagic cell death Induced AMPK phosphorylation Induced mTOR dephosphorylation Activated the Raf/ERK/p90 RSK signaling | [216] |
| Tomentosin | Human leukemia cancer cells | Induced cell autophagy and apoptosis Downregulated mTOR and p-mTOR proteins and PI3K/Akt protein expressions | [217] |
| Parthenolide | MDA-T32 papillary thyroid carcinoma cells and mouse tumor xenografts | Induced cell apoptosis Increased LC3-II, and beclin-1 expression Inhibited the mTOR/PI3K/AKT cascade Inhibited mouse xenograft tumor growth | [218] |
| Zingiberene | Human colon cancer HT-29 cell line | Inhibited colon cancer cell proliferation Induced autophagy Increased LC3-II expression Decreased p62 expression Inhibited mTOR/PI3K/AKT signalling pathway | [219] |
| Urolithin A | Pancreatic ductal adenocarcinoma (PDAC) | Inhibited PDAC cell proliferation and migration Enhanced apoptosis Downregulated PI3K/mTOR pathway Suppressed pancreatic tumor growth Reduced phosphorylation of AKT and p70S6K | [220] |
| Rotundic acid | Human hepatocellular carcinoma cell lines (HepG2, SMMC-7721) HepG2 xenograft mouse model | Inhibited HCC cell proliferation Induced cell cycle arrest, DNA damage, and apoptosis Inhibited tumor growth Regulated the expression of the proteins involved in PI3K/AKT/mTOR and MAPK pathways | [223] |
| Arnicolide D | MDA-MB-231 and MDA-MB-468 triple-negative breast cancer (TNBC) cell lines | Reduced cell viability Induced G2/M cell cycle arrest and apoptosis Reduced cell viability Induced G2/M cell cycle arrest and apoptosis | [221] |
| Atractylenolide I (ATL-1) | COLO205 and HCT116 CRC colorectal cancer cell lines | Inhibited CRC cell invasion Downregulated the phosphorylation of proteins related to the AKT/mTOR pathway | [150] |
| Falcarindiol | Human oral squamous cell carcinomas (OSCCs) cell lines | Suppressed cell growth Induced apoptosis Induced PI3K, AKT, mTOR, and p70S6K dephosphorylation Induced autophagy Induced antimetastatic effects | [222] |
| Eupafolin | Breast cancer cell lines | Decreased p-PI3K, p-Akt, and p-mTOR protein levels Increased apoptosis rate Increased the protein levels of Bax and cleaved caspase 3 Decreased Bcl-2 Inhibited cell migration and invasion Promoted cell apoptosis Induced G0/G1 phase arrest | [224] |
| Chaetocochin J | Colorectal cancer cell lines SW480, HCT116, and RKO | Induced apoptosis and growth inhibition Induced apoptosis and autophagy Inhibited PI3K/AKT/mTOR signaling pathways | [225] |
| Rhein | Human CRC cell lines HCT116, HCT15, and DLD1 and xenograft mice model | Inhibited cell growth Induced S phase cell cycle arrest and apoptosis Inhibited CRC cell migration and invasion ability Suppressed the mTOR/p70S6K signaling pathway | [226] |
5. Synthetic Compounds as Cancer Preventive and Therapeutic Agents by Targeting the mTOR Signaling Pathway
| Inhibitors | Brand Name | Drug’s Form | Administration | Year of FDA Approved | Conditions |
|---|---|---|---|---|---|
| Rapamycin | Rapamune | Tablets or solution | Oral route | 1999 | Lymphangioleiomyomatosis, a rare lung pathology known by abnormal proliferation of smooth muscle-like cells and lymphatic treatment of facial angiofibroma linked with tuberous sclerosis. |
| Hyftor™ | Gel | Topical route | 2022 | ||
| Everolimus | Afinitor® | Tablets | Oral route | 2009 | Advanced hormone receptor, epidermal growth factor receptor breast cancer; progressive neuroendocrine tumors of pancreatic origin; progressive neuroendocrine tumors of lung origin or gastrointestinal; subependymal giant cell astrocytoma and renal angiomyolipomas linked with tuberous sclerosis; advanced renal cell carcinoma. |
| Disperz® | Tablets for suspension | Oral route | 2012 | ||
| Zortress® | Tablets | Oral route | 2010 | ||
| Temsirolimus | Torisel® | Solution | Intravenous route | 2007 | Advanced renal cell carcinoma |
| Nanoparticle rapamycin bound to albumin | Fyarro™ | Suspension | Intravenous route | 2021 | Treatment of perivascular epithelioid cell tumor |
6. Immunotherapy Targeted mTOR
6.1. mTOR and Immunological Cells
6.2. T Cells
6.3. TAMs
6.4. MDSCs
6.5. Combination of mTOR Inhibitors with Therapies
6.6. Inhibition of mTOR in Combination with Tumor Vaccines
6.7. mTOR Inhibitors and Immune Checkpoint Modulation
7. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Brown, E.J.; Albers, M.W.; Bum Shin, T.; Keith, C.T.; Lane, W.S.; Schreiber, S.L. A Mammalian Protein Targeted by G1-Arresting Rapamycin–Receptor Complex. Nature 1994, 369, 756–758. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and Regulation of Akt/PKB by the Rictor-MTOR Complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laplante, M.; Sabatini, D.M. MTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin Passes the Torch: A New Generation of MTOR Inhibitors. Nat. Rev. Drug Discov. 2011, 10, 868–880. [Google Scholar] [CrossRef]
- Foster, D.A. Phosphatidic Acid and Lipid-Sensing by MTOR. Trends Endocrinol. Metab. 2013, 24, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Fingar, D.C. Growing Knowledge of the MTOR Signaling Network. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 36, pp. 79–90. [Google Scholar]
- Kim, S.M.; Vetrivel, P.; Ha, S.E.; Kim, H.H.; Kim, J.-A.; Kim, G.S. Apigetrin Induces Extrinsic Apoptosis, Autophagy and G2/M Phase Cell Cycle Arrest through PI3K/AKT/MTOR Pathway in AGS Human Gastric Cancer Cell. J. Nutr. Biochem. 2020, 83, 108427. [Google Scholar] [CrossRef]
- Khan, K.; Quispe, C.; Javed, Z.; Iqbal, M.J.; Sadia, H.; Raza, S.; Irshad, A.; Salehi, B.; Reiner, Ž.; Sharifi-Rad, J. Resveratrol, Curcumin, Paclitaxel and MiRNAs Mediated Regulation of PI3K/Akt/MTOR Pathway: Go Four Better to Treat Bladder Cancer. Cancer Cell Int. 2020, 20, 560. [Google Scholar] [CrossRef]
- Mafi, S.; Mansoori, B.; Taeb, S.; Sadeghi, H.; Abbasi, R.; Cho, W.C.; Rostamzadeh, D. MTOR-Mediated Regulation of Immune Responses in Cancer and Tumor Microenvironment. Front. Immunol. 2022, 12, 5724. [Google Scholar] [CrossRef]
- Thomson, A.W.; Turnquist, H.R.; Raimondi, G. Immunoregulatory Functions of MTOR Inhibition. Nat. Rev. Immunol. 2009, 9, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Weichhart, T.; Hengstschläger, M.; Linke, M. Regulation of Innate Immune Cell Function by MTOR. Nat. Rev. Immunol. 2015, 15, 599–614. [Google Scholar] [CrossRef]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Weber, J. Immune Checkpoint Proteins: A New Therapeutic Paradigm for Cancer—Preclinical Background: CTLA-4 and PD-1 Blockade. In Seminars in Oncology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 37, pp. 430–439. [Google Scholar]
- Sabatini, D.M.; Erdjument-Bromage, H.; Lui, M.; Tempst, P.; Snyder, S.H. RAFT1: A Mammalian Protein That Binds to FKBP12 in a Rapamycin-Dependent Fashion and Is Homologous to Yeast TORs. Cell 1994, 78, 35–43. [Google Scholar] [CrossRef]
- Sabers, C.J.; Martin, M.M.; Brunn, G.J.; Williams, J.M.; Dumont, F.J.; Wiederrecht, G.; Abraham, R.T. Isolation of a Protein Target of the FKBP12-Rapamycin Complex in Mammalian Cells∗. J. Biol. Chem. 1995, 270, 815–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingras, A.-C.; Raught, B.; Sonenberg, N. Regulation of Translation Initiation by FRAP/MTOR. Genes Dev. 2001, 15, 807–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornu, M.; Albert, V.; Hall, M.N. MTOR in Aging, Metabolism, and Cancer. Curr. Opin. Genet. Dev. 2013, 23, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Maruki, Y.; Long, X.; Yoshino, K.; Oshiro, N.; Hidayat, S.; Tokunaga, C.; Avruch, J.; Yonezawa, K. Raptor, a Binding Partner of Target of Rapamycin (TOR), Mediates TOR Action. Cell 2002, 110, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Sarbassov, D.D.; Ali, S.M.; King, J.E.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. MTOR Interacts with Raptor to Form a Nutrient-Sensitive Complex That Signals to the Cell Growth Machinery. Cell 2002, 110, 163–175. [Google Scholar] [CrossRef] [Green Version]
- Schalm, S.S.; Fingar, D.C.; Sabatini, D.M.; Blenis, J. TOS Motif-Mediated Raptor Binding Regulates 4E-BP1 Multisite Phosphorylation and Function. Curr. Biol. 2003, 13, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Schalm, S.S.; Blenis, J. Identification of a Conserved Motif Required for MTOR Signaling. Curr. Biol. 2002, 12, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Hay, N.; Sonenberg, N. Upstream and Downstream of MTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef]
- Jansson, T.; Aye, I.L.; Goberdhan, D.C. The Emerging Role of MTORC1 Signaling in Placental Nutrient-Sensing. Placenta 2012, 33, e23–e29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoki, K.; Li, Y.; Zhu, T.; Wu, J.; Guan, K.-L. TSC2 Is Phosphorylated and Inhibited by Akt and Suppresses MTOR Signalling. Nat. Cell Biol. 2002, 4, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Tee, A.R.; Logsdon, M.N.; Blenis, J.; Cantley, L.C. Identification of the Tuberous Sclerosis Complex-2 Tumor Suppressor Gene Product Tuberin as a Target of the Phosphoinositide 3-Kinase/Akt Pathway. Mol. Cell 2002, 10, 151–162. [Google Scholar] [CrossRef]
- Ballif, B.A.; Roux, P.P.; Gerber, S.A.; MacKeigan, J.P.; Blenis, J.; Gygi, S.P. Quantitative Phosphorylation Profiling of the ERK/P90 Ribosomal S6 Kinase-Signaling Cassette and Its Targets, the Tuberous Sclerosis Tumor Suppressors. Proc. Natl. Acad. Sci. USA 2005, 102, 667–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, P.P.; Ballif, B.A.; Anjum, R.; Gygi, S.P.; Blenis, J. Tumor-Promoting Phorbol Esters and Activated Ras Inactivate the Tuberous Sclerosis Tumor Suppressor Complex via P90 Ribosomal S6 Kinase. Proc. Natl. Acad. Sci. USA 2004, 101, 13489–13494. [Google Scholar] [CrossRef] [Green Version]
- Tee, A.R.; Anjum, R.; Blenis, J. Inactivation of the Tuberous Sclerosis Complex-1 and-2 Gene Products Occurs by Phosphoinositide 3-Kinase/Akt-Dependent and-Independent Phosphorylation of Tuberin. J. Biol. Chem. 2003, 278, 37288–37296. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.G.; Buel, G.R.; Blenis, J. Nutrient Regulation of the MTOR Complex 1 Signaling Pathway. Mol. Cells 2013, 35, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Avivar-Valderas, A.; Bobrovnikova-Marjon, E.; Alan Diehl, J.; Bardeesy, N.; Debnath, J.; Aguirre-Ghiso, J.A. Regulation of Autophagy during ECM Detachment Is Linked to a Selective Inhibition of MTORC1 by PERK. Oncogene 2013, 32, 4932–4940. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zhang, S.; He, L.; Rong, Y.; Brier, L.W.; Sun, Q.; Liu, R.; Fan, W.; Chen, S.; Yue, Z. MTORC1-Mediated NRBF2 Phosphorylation Functions as a Switch for the Class III PtdIns3K and Autophagy. Autophagy 2017, 13, 592–607. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.-X.; Russell, R.C.; Guan, K.-L. Regulation of PIK3C3/VPS34 Complexes by MTOR in Nutrient Stress-Induced Autophagy. Autophagy 2013, 9, 1983–1995. [Google Scholar] [CrossRef]
- Martina, J.A.; Chen, Y.; Gucek, M.; Puertollano, R. MTORC1 Functions as a Transcriptional Regulator of Autophagy by Preventing Nuclear Transport of TFEB. Autophagy 2012, 8, 903–914. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Kakuta, S.; Watanabe, T.M.; Kitamura, A.; Sekito, T.; Kondo-Kakuta, C.; Ichikawa, R.; Kinjo, M.; Ohsumi, Y. Atg9 Vesicles Are an Important Membrane Source during Early Steps of Autophagosome Formation. J. Cell Biol. 2012, 198, 219–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and MTOR Regulate Autophagy through Direct Phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, R.M.; Zoncu, R. The Lysosome as a Regulatory Hub. Annu. Rev. Cell Dev. Biol. 2016, 32, 223–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.Y.-S.; Shin, H.R.; Berdan, C.A.; Ford, B.; Ward, C.C.; Olzmann, J.A.; Zoncu, R.; Nomura, D.K. Covalent Targeting of the Vacuolar H+-ATPase Activates Autophagy via MTORC1 Inhibition. Nat. Chem. Biol. 2019, 15, 776–785. [Google Scholar] [CrossRef]
- Suk, J.; Kwak, S.S.; Lee, J.H.; Choi, J.H.; Lee, S.-H.; Lee, D.H.; Byun, B.; Lee, G.-H.; Joe, C.O. Alkaline Stress-Induced Autophagy Is Mediated by MTORC1 Inactivation. J. Cell Biochem. 2011, 112, 2566–2573. [Google Scholar] [CrossRef]
- Cybulski, N.; Polak, P.; Auwerx, J.; Rüegg, M.A.; Hall, M.N. MTOR Complex 2 in Adipose Tissue Negatively Controls Whole-Body Growth. Proc. Natl. Acad. Sci. USA 2009, 106, 9902–9907. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Harris, T.E.; Keller, S.R.; Choi, K.M.; Magnuson, M.A.; Lawrence, J.C., Jr. Muscle-Specific Deletion of Rictor Impairs Insulin-Stimulated Glucose Transport and Enhances Basal Glycogen Synthase Activity. Mol. Cell Biol. 2008, 28, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wendel, A.A.; Keogh, M.R.; Harris, T.E.; Chen, J.; Coleman, R.A. Glycerolipid Signals Alter MTOR Complex 2 (MTORC2) to Diminish Insulin Signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 1667–1672. [Google Scholar] [CrossRef]
- Morita, M.; Gravel, S.-P.; Hulea, L.; Larsson, O.; Pollak, M.; St-Pierre, J.; Topisirovic, I. MTOR Coordinates Protein Synthesis, Mitochondrial Activity and Proliferation. Cell Cycle 2015, 14, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, M.J.; Rasmussen, B.B. Leucine-Enriched Nutrients and the Regulation of MTOR Signalling and Human Skeletal Muscle Protein Synthesis. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Conn, C.S.; Qian, S.-B. MTOR Signaling in Protein Homeostasis: Less Is More? Cell Cycle 2011, 10, 1940–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, A.A.; Proud, C.G. The Rapid Activation of Protein Synthesis by Growth Hormone Requires Signaling through MTOR. Am. J. Physiol.-Endocrinol. Metab. 2007, 292, E1647–E1655. [Google Scholar] [CrossRef] [Green Version]
- Thoreen, C.C. Many Roads from MTOR to EIF4F. Biochem. Soc. Trans. 2013, 41, 913–916. [Google Scholar] [CrossRef] [Green Version]
- Miloslavski, R.; Cohen, E.; Avraham, A.; Iluz, Y.; Hayouka, Z.; Kasir, J.; Mudhasani, R.; Jones, S.N.; Cybulski, N.; Rüegg, M.A. Oxygen Sufficiency Controls TOP MRNA Translation via the TSC-Rheb-MTOR Pathway in a 4E-BP-Independent Manner. J. Mol. Cell Biol. 2014, 6, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Patursky-Polischuk, I.; Stolovich-Rain, M.; Hausner-Hanochi, M.; Kasir, J.; Cybulski, N.; Avruch, J.; Rüegg, M.A.; Hall, M.N.; Meyuhas, O. The TSC-MTOR Pathway Mediates Translational Activation of TOP MRNAs by Insulin Largely in a Raptor-or Rictor-Independent Manner. Mol. Cell Biol. 2009, 29, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Stolovich, M.; Tang, H.; Hornstein, E.; Levy, G.; Cohen, R.; Bae, S.S.; Birnbaum, M.J.; Meyuhas, O. Transduction of Growth or Mitogenic Signals into Translational Activation of TOP MRNAs Is Fully Reliant on the Phosphatidylinositol 3-Kinase-Mediated Pathway but Requires Neither S6K1 nor RpS6 Phosphorylation. Mol. Cell Biol. 2002, 22, 8101–8113. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Hornstein, E.; Stolovich, M.; Levy, G.; Livingstone, M.; Templeton, D.; Avruch, J.; Meyuhas, O. Amino Acid-Induced Translation of TOP MRNAs Is Fully Dependent on Phosphatidylinositol 3-Kinase-Mediated Signaling, Is Partially Inhibited by Rapamycin, and Is Independent of S6K1 and RpS6 Phosphorylation. Mol. Cell Biol. 2001, 21, 8671–8683. [Google Scholar] [CrossRef] [Green Version]
- Musa, J.; Orth, M.F.; Dallmayer, M.; Baldauf, M.; Pardo, C.; Rotblat, B.; Kirchner, T.; Leprivier, G.; Grünewald, T.G.P. Eukaryotic Initiation Factor 4E-Binding Protein 1 (4E-BP1): A Master Regulator of MRNA Translation Involved in Tumorigenesis. Oncogene 2016, 35, 4675–4688. [Google Scholar] [CrossRef]
- Mamane, Y.; Petroulakis, E.; Martineau, Y.; Sato, T.-A.; Larsson, O.; Rajasekhar, V.K.; Sonenberg, N. Epigenetic Activation of a Subset of MRNAs by EIF4E Explains Its Effects on Cell Proliferation. PLoS ONE 2007, 2, e242. [Google Scholar] [CrossRef] [PubMed]
- Eliseeva, I.; Vasilieva, M.; Ovchinnikov, L.P. Translation of Human β-Actin MRNA Is Regulated by MTOR Pathway. Genes 2019, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of Translation Initiation in Eukaryotes: Mechanisms and Biological Targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancak, Y.; Peterson, T.R.; Shaul, Y.D.; Lindquist, R.A.; Thoreen, C.C.; Bar-Peled, L.; Sabatini, D.M. The Rag GTPases Bind Raptor and Mediate Amino Acid Signaling to MTORC1. Science 2008, 320, 1496–1501. [Google Scholar] [CrossRef] [Green Version]
- Ben-Sahra, I.; Howell, J.J.; Asara, J.M.; Manning, B.D. Stimulation of de Novo Pyrimidine Synthesis by Growth Signaling through MTOR and S6K1. Science 2013, 339, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Mayer, C.; Grummt, I. Ribosome Biogenesis and Cell Growth: MTOR Coordinates Transcription by All Three Classes of Nuclear RNA Polymerases. Oncogene 2006, 25, 6384–6391. [Google Scholar] [CrossRef] [Green Version]
- Mayer, C.; Zhao, J.; Yuan, X.; Grummt, I. MTOR-Dependent Activation of the Transcription Factor TIF-IA Links RRNA Synthesis to Nutrient Availability. Genes Dev. 2004, 18, 423–434. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Yang, Y.; Zhang, J.; Chen, J.; Wei, X.; He, J.; Luo, L. Ribosome Biogenesis Protein Urb1 Acts Downstream of MTOR Complex 1 to Modulate Digestive Organ Development in Zebrafish. J. Genet. Genom. 2017, 44, 567–576. [Google Scholar] [CrossRef]
- Binal, Z.; Açıkgöz, E.; Kızılay, F.; Öktem, G.; Altay, B. Cross-Talk between Ribosome Biogenesis, Translation, and MTOR in CD133+ 4/CD44+ Prostate Cancer Stem Cells. Clin. Transl. Oncol. 2020, 22, 1040–1048. [Google Scholar] [CrossRef]
- Kazyken, D.; Kaz, Y.; Kiyan, V.; Zhylkibayev, A.A.; Chen, C.-H.; Agarwal, N.K.; Sarbassov, D.D. The Nuclear Import of Ribosomal Proteins Is Regulated by MTOR. Oncotarget 2014, 5, 9577. [Google Scholar] [CrossRef]
- Ping, W.; Liu, X.; Lei, D.; Zhang, X. MTOR Signaling-Related MicroRNAs and Cancer Involvement. J. Cancer 2018, 9, 667. [Google Scholar]
- Feng, Z.; Zhang, H.; Levine, A.J.; Jin, S. The Coordinate Regulation of the P53 and MTOR Pathways in Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 8204–8209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the P53 Network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef]
- Feng, Z.; Hu, W.; De Stanchina, E.; Teresky, A.K.; Jin, S.; Lowe, S.; Levine, A.J. The Regulation of AMPK Β1, TSC2, and PTEN Expression by P53: Stress, Cell and Tissue Specificity, and the Role of These Gene Products in Modulating the IGF-1-AKT-MTOR Pathways. Cancer Res. 2007, 67, 3043–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Chen, X.; Ma, J.; Peng, H.; Wang, F.; Zha, X.; Wang, Y.; Jing, Y.; Yang, H.; Chen, R. Mammalian Target of Rapamycin Up-Regulation of Pyruvate Kinase Isoenzyme Type M2 Is Critical for Aerobic Glycolysis and Tumor Growth. Proc. Natl. Acad. Sci. USA 2011, 108, 4129–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, G.G.; Abraham, R.T. Targeting the MTOR Signaling Network in Cancer. Trends Mol. Med. 2007, 13, 433–442. [Google Scholar] [CrossRef]
- Jhanwar-Uniyal, M.; Jeevan, D.; Neil, J.; Shannon, C.; Albert, L.; Murali, R. Deconstructing MTOR Complexes in Regulation of Glioblastoma Multiforme and Its Stem Cells. Adv. Biol. Regul. 2013, 53, 202–210. [Google Scholar] [CrossRef]
- Nagahashi, M.; Hait, N.C.; Maceyka, M.; Avni, D.; Takabe, K.; Milstien, S.; Spiegel, S. Sphingosine-1-Phosphate in Chronic Intestinal Inflammation and Cancer. Adv. Biol. Regul. 2014, 54, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Pyne, N.J.; Ohotski, J.; Bittman, R.; Pyne, S. The Role of Sphingosine 1-Phosphate in Inflammation and Cancer. Adv. Biol. Regul. 2014, 54, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Pyne, N.J.; Tonelli, F.; Lim, K.G.; Long, J.; Edwards, J.; Pyne, S. Targeting Sphingosine Kinase 1 in Cancer. Adv. Biol. Regul. 2012, 52, 31–38. [Google Scholar] [CrossRef]
- Liu, L.; Li, F.; Cardelli, J.A.; Martin, K.A.; Blenis, J.; Huang, S. Rapamycin Inhibits Cell Motility by Suppression of MTOR-Mediated S6K1 and 4E-BP1 Pathways. Oncogene 2006, 25, 7029–7040. [Google Scholar] [CrossRef] [PubMed]
- Vivanco, I.; Sawyers, C.L. The Phosphatidylinositol 3-Kinase–AKT Pathway in Human Cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Sansal, I.; Sellers, W.R. The Biology and Clinical Relevance of the PTEN Tumor Suppressor Pathway. J. Clin. Oncol. 2004, 22, 2954–2963. [Google Scholar] [CrossRef] [PubMed]
- Stiles, B.; Groszer, M.; Wang, S.; Jiao, J.; Wu, H. PTENless Means More. Dev. Biol. 2004, 273, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wymann, M.P.; Zvelebil, M.; Laffargue, M. Phosphoinositide 3-Kinase Signalling–Which Way to Target? Trends Pharmacol. Sci. 2003, 24, 366–376. [Google Scholar] [CrossRef]
- Menon, S.; Manning, B.D. Common Corruption of the MTOR Signaling Network in Human Tumors. Oncogene 2008, 27, S43–S51. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Chen, Z.; Erdjument-Bromage, H.; Tempst, P.; Pandolfi, P.P. Phosphorylation and Functional Inactivation of TSC2 by Erk: Implications for Tuberous Sclerosisand Cancer Pathogenesis. Cell 2005, 121, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Potter, C.J.; Pedraza, L.G.; Xu, T. Akt Regulates Growth by Directly Phosphorylating Tsc2. Nat. Cell Biol. 2002, 4, 658–665. [Google Scholar] [CrossRef]
- Lee, D.-F.; Kuo, H.-P.; Chen, C.-T.; Hsu, J.-M.; Chou, C.-K.; Wei, Y.; Sun, H.-L.; Li, L.-Y.; Ping, B.; Huang, W.-C. IKKβ Suppression of TSC1 Links Inflammation and Tumor Angiogenesis via the MTOR Pathway. Cell 2007, 130, 440–455. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Inoki, K.; Vacratsis, P.; Guan, K.-L. The P38 and MK2 Kinase Cascade Phosphorylates Tuberin, the Tuberous Sclerosis 2 Gene Product, and Enhances Its Interaction with 14-3-3. J. Biol. Chem. 2003, 278, 13663–13671. [Google Scholar] [CrossRef] [Green Version]
- Dalle Pezze, P.; Ruf, S.; Sonntag, A.G.; Langelaar-Makkinje, M.; Hall, P.; Heberle, A.M.; Razquin Navas, P.; Van Eunen, K.; Tölle, R.C.; Schwarz, J.J. A Systems Study Reveals Concurrent Activation of AMPK and MTOR by Amino Acids. Nat. Commun. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Efeyan, A.; Zoncu, R.; Chang, S.; Gumper, I.; Snitkin, H.; Wolfson, R.L.; Kirak, O.; Sabatini, D.D.; Sabatini, D.M. Regulation of MTORC1 by the Rag GTPases Is Necessary for Neonatal Autophagy and Survival. Nature 2013, 493, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghislat, G.; Patron, M.; Rizzuto, R.; Knecht, E. Withdrawal of Essential Amino Acids Increases Autophagy by a Pathway Involving Ca2+/Calmodulin-Dependent Kinase Kinase-β (CaMKK-β). J. Biol. Chem. 2012, 287, 38625–38636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andjelkovic, M.; Alessi, D.R.; Meier, R.; Fernandez, A.; Lamb, N.J.; Frech, M.; Cron, P.; Cohen, P.; Lucocq, J.M.; Hemmings, B.A. Role of Translocation in the Activation and Function of Protein Kinase B. J. Biol. Chem. 1997, 272, 31515–31524. [Google Scholar] [CrossRef] [Green Version]
- Frech, M.; Andjelkovic, M.; Ingley, E.; Reddy, K.K.; Falck, J.R.; Hemmings, B.A. High Affinity Binding of Inositol Phosphates and Phosphoinositides to the Pleckstrin Homology Domain of RAC/Protein Kinase B and Their Influence on Kinase Activity. J. Biol. Chem. 1997, 272, 8474–8481. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.M.; Blenis, J. Molecular Mechanisms of MTOR-Mediated Translational Control. Nat. Rev. Mol. Cell Biol. 2009, 10, 307–318. [Google Scholar] [CrossRef]
- Brunn, G.J.; Hudson, C.C.; Sekulic, A.; Williams, J.M.; Hosoi, H.; Houghton, P.J.; Lawrence, J.C., Jr.; Abraham, R.T. Phosphorylation of the Translational Repressor PHAS-I by the Mammalian Target of Rapamycin. Science 1997, 277, 99–101. [Google Scholar] [CrossRef]
- Hara, K.; Yonezawa, K.; Kozlowski, M.T.; Sugimoto, T.; Andrabi, K.; Weng, Q.-P.; Kasuga, M.; Nishimoto, I.; Avruch, J. Regulation of EIF-4E BP1 Phosphorylation by MTOR. J. Biol. Chem. 1997, 272, 26457–26463. [Google Scholar] [CrossRef] [Green Version]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.R.; Chung, Y.-L.; Schulze, A. SREBP Activity Is Regulated by MTORC1 and Contributes to Akt-Dependent Cell Growth. Cell Metab. 2008, 8, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Castilho, R.M.; Squarize, C.H.; Chodosh, L.A.; Williams, B.O.; Gutkind, J.S. MTOR Mediates Wnt-Induced Epidermal Stem Cell Exhaustion and Aging. Cell Stem Cell 2009, 5, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Laplante, M.; Sabatini, D.M. An Emerging Role of MTOR in Lipid Biosynthesis. Curr. Biol. 2009, 19, R1046–R1052. [Google Scholar] [CrossRef] [PubMed]
- Zoncu, R.; Efeyan, A.; Sabatini, D.M. MTOR: From Growth Signal Integration to Cancer, Diabetes and Ageing. Nat. Rev. Mol. Cell Biol. 2011, 12, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, P.E.; Barrow, R.K.; Cohen, N.A.; Snyder, S.H.; Sabatini, D.M. RAFT1 Phosphorylation of the Translational Regulators P70 S6 Kinase and 4E-BP1. Proc. Natl. Acad. Sci. USA 1998, 95, 1432–1437. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.J.; Beal, P.A.; Keith, C.T.; Chen, J.; Bum Shin, T.; Schreiber, S.L. Control of P70 S6 Kinase by Kinase Activity of FRAP in Vivo. Nature 1995, 377, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Gingras, A.-C.; Raught, B.; Sonenberg, N. EIF4 Initiation Factors: Effectors of MRNA Recruitment to Ribosomes and Regulators of Translation. Annu. Rev. Biochem. 1999, 68, 913–963. [Google Scholar] [CrossRef]
- Wang, X.; Beugnet, A.; Murakami, M.; Yamanaka, S.; Proud, C.G. Distinct Signaling Events Downstream of MTOR Cooperate to Mediate the Effects of Amino Acids and Insulin on Initiation Factor 4E-Binding Proteins. Mol. Cell Biol. 2005, 25, 2558–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yellen, P.; Saqcena, M.; Salloum, D.; Feng, J.; Preda, A.; Xu, L.; Rodrik-Outmezguine, V.; Foster, D.A. High-Dose Rapamycin Induces Apoptosis in Human Cancer Cells by Dissociating MTOR Complex 1 and Suppressing Phosphorylation of 4E-BP1. Cell Cycle 2011, 10, 3948–3956. [Google Scholar] [CrossRef] [Green Version]
- Bärlund, M.; Monni, O.; Kononen, J.; Cornelison, R.; Torhorst, J.; Sauter, G.; Kallioniemi, O.-P.; Kallioniemi, A. Multiple Genes at 17q23 Undergo Amplification and Overexpression in Breast Cancer. Cancer Res. 2000, 60, 5340–5344. [Google Scholar]
- Surace, E.I.; Lusis, E.; Haipek, C.A.; Gutmann, D.H. Functional Significance of S6K Overexpression in Meningioma Progression. Ann. Neurol. Off. J. Am. Neurol. Assoc. Child Neurol. Soc. 2004, 56, 295–298. [Google Scholar] [CrossRef]
- Kim, J.E.; Chen, J. Regulation of Peroxisome Proliferator–Activated Receptor-γ Activity by Mammalian Target of Rapamycin and Amino Acids in Adipogenesis. Diabetes 2004, 53, 2748–2756. [Google Scholar] [CrossRef] [Green Version]
- Armour, S.M.; Baur, J.A.; Hsieh, S.N.; Land-Bracha, A.; Thomas, S.M.; Sinclair, D.A. Inhibition of Mammalian S6 Kinase by Resveratrol Suppresses Autophagy. Aging 2009, 1, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingar, D.C.; Salama, S.; Tsou, C.; Harlow, E.D.; Blenis, J. Mammalian Cell Size Is Controlled by MTOR and Its Downstream Targets S6K1 and 4EBP1/EIF4E. Genes Dev. 2002, 16, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sheng, Z.; Cai, Y. Effects of MicroRNA-513b on Cell Proliferation, Apoptosis, Invasion, and Migration by Targeting HMGB3 through Regulation of MTOR Signaling Pathway in Non-Small-Cell Lung Cancer. J. Cell Physiol. 2019, 234, 10934–10941. [Google Scholar] [CrossRef]
- Sun, J.; Chen, Z.; Tan, X.; Zhou, F.; Tan, F.; Gao, Y.; Sun, N.; Xu, X.; Shao, K.; He, J. MicroRNA-99a/100 Promotes Apoptosis by Targeting MTOR in Human Esophageal Squamous Cell Carcinoma. Med. Oncol. 2013, 30, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Zhang, Y. MicroRNA-381 Facilitates Autophagy and Apoptosis in Prostate Cancer Cells via Inhibiting the RELN-Mediated PI3K/AKT/MTOR Signaling Pathway. Life Sci. 2020, 254, 117672. [Google Scholar] [CrossRef]
- Liu, T.; Ye, P.; Ye, Y.; Han, B. MicroRNA-216b Targets HK2 to Potentiate Autophagy and Apoptosis of Breast Cancer Cells via the MTOR Signaling Pathway. Int. J. Biol. Sci. 2021, 17, 2970. [Google Scholar] [CrossRef]
- Li, X.J.; Luo, X.Q.; Han, B.W.; Duan, F.T.; Wei, P.P.; Chen, Y.Q. MicroRNA-100/99a, Deregulated in Acute Lymphoblastic Leukaemia, Suppress Proliferation and Promote Apoptosis by Regulating the FKBP51 and IGF1R/MTOR Signalling Pathways. Br. J. Cancer 2013, 109, 2189–2198. [Google Scholar] [CrossRef] [Green Version]
- Ueng, S.-H.; Chen, S.-C.; Chang, Y.-S.; Hsueh, S.; Lin, Y.-C.; Chien, H.-P.; Lo, Y.-F.; Shen, S.-C.; Hsueh, C. Phosphorylated MTOR Expression Correlates with Poor Outcome in Early-Stage Triple Negative Breast Carcinomas. Int. J. Clin. Exp. Pathol. 2012, 5, 806. [Google Scholar]
- Xu, K.; Liu, P.; Wei, W. MTOR Signaling in Tumorigenesis. Biochim. Biophys. Acta BBA-Rev. Cancer 2014, 1846, 638–654. [Google Scholar] [CrossRef] [Green Version]
- Krieger, K.L.; Hu, W.-F.; Ripperger, T.; Woods, N.T. Functional Impacts of the BRCA1-MTORC2 Interaction in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5876. [Google Scholar] [CrossRef] [Green Version]
- Woods, N.T.; Mesquita, R.D.; Sweet, M.; Carvalho, M.A.; Li, X.; Liu, Y.; Nguyen, H.; Thomas, C.E.; Iversen, E.S., Jr.; Marsillac, S. Charting the Landscape of Tandem BRCT Domain–Mediated Protein Interactions. Sci. Signal. 2012, 5, rs6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wazir, U.; Newbold, R.F.; Jiang, W.G.; Sharma, A.K.; Mokbel, K. Prognostic and Therapeutic Implications of MTORC1 and Rictor Expression in Human Breast Cancer. Oncol. Rep. 2013, 29, 1969–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogan, F.; Avci, C.B. Correlation between Telomerase and MTOR Pathway in Cancer Stem Cells. Gene 2018, 641, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Hynes, N.E.; Boulay, A. The MTOR Pathway in Breast Cancer. J. Mammary Gland Biol. Neoplasia 2006, 11, 53–61. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Montalto, G.; Cervello, M.; Nicoletti, F.; Fagone, P.; Malaponte, G.; Mazzarino, M.C. Mutations and Deregulation of Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/MTOR Cascades Which Alter Therapy Response. Oncotarget 2012, 3, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strimpakos, A.S.; Karapanagiotou, E.M.; Saif, M.W.; Syrigos, K.N. The Role of MTOR in the Management of Solid Tumors: An Overview. Cancer Treat. Rev. 2009, 35, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.; Flanagan, L.; Quinn, C.; Evoy, D.; McDermott, E.W.; Pierce, A.; Duffy, M.J. MTOR in Breast Cancer: Differential Expression in Triple-Negative and Non-Triple-Negative Tumors. Breast 2012, 21, 178–182. [Google Scholar] [CrossRef]
- Droog, M.; Beelen, K.; Linn, S.; Zwart, W. Tamoxifen Resistance: From Bench to Bedside. Eur. J. Pharmacol. 2013, 717, 47–57. [Google Scholar] [CrossRef]
- Viedma-Rodríguez, R.; Baiza-Gutman, L.; Salamanca-Gómez, F.; Diaz-Zaragoza, M.; Martínez-Hernández, G.; Ruiz Esparza-Garrido, R.; Velázquez-Flores, M.A.; Arenas-Aranda, D. Mechanisms Associated with Resistance to Tamoxifen in Estrogen Receptor-Positive Breast Cancer. Oncol. Rep. 2014, 32, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Hare, S.H.; Harvey, A.J. MTOR Function and Therapeutic Targeting in Breast Cancer. Am. J. Cancer Res. 2017, 7, 383. [Google Scholar]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.W.; Zhang, J.; Tsai, M.-H.; Yu, D. PI3K-Independent MTOR Activation Promotes Lapatinib Resistance and IAP Expression That Can Be Effectively Reversed by MTOR and Hsp90 Inhibition. Cancer Biol. Ther. 2015, 16, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Margariti, N.; Fox, S.B.; Bottini, A.; Generali, D. “Overcoming Breast Cancer Drug Resistance with MTOR Inhibitors”. Could It Be a Myth or a Real Possibility in the Short-Term Future? Breast Cancer Res. Treat. 2011, 128, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mao, C.; Wang, M.; Liu, N.; Ouyang, L.; Liu, S.; Tang, H.; Cao, Y.; Liu, S.; Wang, X. Cancer Progression Is Mediated by Proline Catabolism in Non-Small Cell Lung Cancer. Oncogene 2020, 39, 2358–2376. [Google Scholar] [CrossRef]
- Schettino, C.; Bareschino, M.A.; Sacco, P.C.; Maione, P.; Rossi, A.; Casaluce, F.; Sgambato, A.; Gridelli, C. New Molecular Targets in the Treatment of NSCLC. Curr. Pharm. Des. 2013, 19, 5333–5343. [Google Scholar] [CrossRef]
- Pothongsrisit, S.; Pongrakhananon, V. Targeting the PI3K/AKT/MTOR Signaling Pathway in Lung Cancer: An Update Regarding Potential Drugs and Natural Products. Molecules 2021, 26, 4100. [Google Scholar]
- Ekman, S.; Wynes, M.W.; Hirsch, F.R. The MTOR Pathway in Lung Cancer and Implications for Therapy and Biomarker Analysis. J. Thorac. Oncol. 2012, 7, 947–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Getz, G.; Wheeler, D.A.; Mardis, E.R.; McLellan, M.D.; Cibulskis, K.; Sougnez, C.; Greulich, H.; Muzny, D.M.; Morgan, M.B. Somatic Mutations Affect Key Pathways in Lung Adenocarcinoma. Nature 2008, 455, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the Phosphoinositide 3-Kinase Pathway in Cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Marcus, A.I.; Vertino, P.M. Dysregulation of MTOR Activity through LKB1 Inactivation. Chin. J. Cancer 2013, 32, 427. [Google Scholar] [CrossRef] [Green Version]
- Mamane, Y.; Petroulakis, E.; Rong, L.; Yoshida, K.; Ler, L.W.; Sonenberg, N. EIF4E–from Translation to Transformation. Oncogene 2004, 23, 3172–3179. [Google Scholar] [CrossRef] [Green Version]
- Rosenwald, I.B.; Hutzler, M.J.; Wang, S.; Savas, L.; Fraire, A.E. Expression of Eukaryotic Translation Initiation Factors 4E and 2α Is Increased Frequently in Bronchioloalveolar but Not in Squamous Cell Carcinomas of the Lung. Cancer 2001, 92, 2164–2171. [Google Scholar] [CrossRef]
- Seki, N.; Takasu, T.; Mandai, K.; Nakata, M.; Saeki, H.; Heike, Y.; Takata, I.; Segawa, Y.; Hanafusa, T.; Eguchi, K. Expression of Eukaryotic Initiation Factor 4E in Atypical Adenomatous Hyperplasia and Adenocarcinoma of the Human Peripheral Lung. Clin. Cancer Res. 2002, 8, 3046–3053. [Google Scholar] [PubMed]
- Frankel, S.K.; Moats-Staats, B.M.; Cool, C.D.; Wynes, M.W.; Stiles, A.D.; Riches, D.W. Human Insulin-like Growth Factor-IA Expression in Transgenic Mice Promotes Adenomatous Hyperplasia but Not Pulmonary Fibrosis. Am. J. Physiol.-Lung Cell Mol. Physiol. 2005, 288, L805–L812. [Google Scholar] [CrossRef] [PubMed]
- Conde, E.; Angulo, B.; Tang, M.; Morente, M.; Torres-Lanzas, J.; Lopez-Encuentra, A.; Lopez-Rios, F.; Sanchez-Cespedes, M. Molecular Context of the EGFR Mutations: Evidence for the Activation of MTOR/S6K Signaling. Clin. Cancer Res. 2006, 12, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Faoro, L.; Singleton, P.A.; Cervantes, G.M.; Lennon, F.E.; Choong, N.W.; Kanteti, R.; Ferguson, B.D.; Husain, A.N.; Tretiakova, M.S.; Ramnath, N. EphA2 Mutation in Lung Squamous Cell Carcinoma Promotes Increased Cell Survival, Cell Invasion, Focal Adhesions, and Mammalian Target of Rapamycin Activation. J. Biol. Chem. 2010, 285, 18575–18585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobashi, Y.; Suzuki, S.; Matsubara, H.; Kimura, M.; Endo, S.; Ooi, A. Critical and Diverse Involvement of Akt/Mammalian Target of Rapamycin Signaling in Human Lung Carcinomas. Cancer 2009, 115, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.; Ding, J.; Huang, Y.; Liu, J.; Liu, N.; Ao, Y.; Hong, Y.; Wang, L.; Zhang, L. Targeting MTOR Suppressed Colon Cancer Growth through 4EBP1/EIF4E/PUMA Pathway. Cancer Gene Ther. 2020, 27, 448–460. [Google Scholar] [CrossRef]
- Francipane, M.G.; Lagasse, E. MTOR Pathway in Colorectal Cancer: An Update. Oncotarget 2014, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Gulhati, P.; Cai, Q.; Li, J.; Liu, J.; Rychahou, P.G.; Qiu, S.; Lee, E.Y.; Silva, S.R.; Bowen, K.A.; Gao, T. Targeted Inhibition of Mammalian Target of Rapamycin Signaling Inhibits Tumorigenesis of Colorectal CancerTargeting MTOR Signaling in Colorectal Cancer. Clin. Cancer Res. 2009, 15, 7207–7216. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Zheng, X.; Li, M.; Zhang, L.; Yu, J. MTOR Inhibitors Induce Apoptosis in Colon Cancer Cells via CHOP-Dependent DR5 Induction on 4E-BP1 Dephosphorylation. Oncogene 2016, 35, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-J.; Dai, Q.; Sun, D.-F.; Xiong, H.; Tian, X.-Q.; Gao, F.-H.; Xu, M.-H.; Chen, G.-Q.; Han, Z.-G.; Fang, J.-Y. MTOR Signaling Pathway Is a Target for the Treatment of Colorectal Cancer. Ann. Surg. Oncol. 2009, 16, 2617–2628. [Google Scholar] [CrossRef] [PubMed]
- Roulin, D.; Cerantola, Y.; Dormond-Meuwly, A.; Demartines, N.; Dormond, O. Targeting MTORC2 Inhibits Colon Cancer Cell Proliferation in Vitro and Tumor Formation in Vivo. Mol. Cancer 2010, 9, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Gulhati, P.; Bowen, K.A.; Liu, J.; Stevens, P.D.; Rychahou, P.G.; Chen, M.; Lee, E.Y.; Weiss, H.L.; O’Connor, K.L.; Gao, T. MTORC1 and MTORC2 Regulate EMT, Motility, and Metastasis of Colorectal Cancer via RhoA and Rac1 Signaling PathwaysmTOR Signaling in EMT and Metastasis of CRC. Cancer Res. 2011, 71, 3246–3256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Tamai, Y.; Horiike, S.; Oshima, M.; Taketo, M.M. Colonic Polyposis Caused by MTOR-Mediated Chromosomal Instability in Apc+/Δ716 Cdx2+/- Compound Mutant Mice. Nat. Genet. 2003, 35, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xu, C.; Yang, W.; Chen, J.; Ou, Y.; Guan, Y.; Guan, J.; Liu, Y. E3 Ligase RNF167 and Deubiquitinase STAMBPL1 Modulate MTOR and Cancer Progression. Mol. Cell 2022, 82, 770–784. [Google Scholar] [CrossRef]
- Weng, M.; Chen, W.; Chen, X.; Lu, H.; Sun, Z.; Yu, Q.; Sun, P.; Xu, Y.; Zhu, M.; Jiang, N. Fasting Inhibits Aerobic Glycolysis and Proliferation in Colorectal Cancer via the Fdft1-Mediated AKT/MTOR/HIF1α Pathway Suppression. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Huang, W.; Sang, X.; Wu, X.; Shan, Q.; Tang, D.; Xu, X.; Cao, G. Atractylenolide I Inhibits Colorectal Cancer Cell Proliferation by Affecting Metabolism and Stemness via AKT/MTOR Signaling. Phytomedicine 2020, 68, 153191. [Google Scholar] [CrossRef]
- Roper, J.; Richardson, M.P.; Wang, W.V.; Richard, L.G.; Chen, W.; Coffee, E.M.; Sinnamon, M.J.; Lee, L.; Chen, P.-C.; Bronson, R.T. The Dual PI3K/MTOR Inhibitor NVP-BEZ235 Induces Tumor Regression in a Genetically Engineered Mouse Model of PIK3CA Wild-Type Colorectal Cancer. PLoS ONE 2011, 6, e25132. [Google Scholar] [CrossRef] [Green Version]
- Belmont, P.J.; Jiang, P.; McKee, T.D.; Xie, T.; Isaacson, J.; Baryla, N.E.; Roper, J.; Sinnamon, M.J.; Lee, N.V.; Kan, J.L. Resistance to Dual Blockade of the Kinases PI3K and MTOR in KRAS-Mutant Colorectal Cancer Models Results in Combined Sensitivity to Inhibition of the Receptor Tyrosine Kinase EGFR. Sci. Signal. 2014, 7, ra107. [Google Scholar] [CrossRef]
- Shen, J.; Wang, A.; Wang, Q.; Gurvich, I.; Siegel, A.B.; Remotti, H.; Santella, R.M. Exploration of Genome-Wide Circulating MicroRNA in Hepatocellular Carcinoma: MiR-483-5p as a Potential Biomarker. Cancer Epidemiol. Prev. Biomark. 2013, 22, 2364–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, F.H.; Bai, Y.; Saintigny, P.; Darido, C. MTOR Signalling in Head and Neck Cancer: Heads Up. Cells 2019, 8, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Li, X.; Zhang, J. MTOR Signaling in Cancer and MTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.-J.; Zhang, L.; Hall, B.; Bian, Y.; Gutkind, J.S.; Kulkarni, A.B. Chemopreventive and Chemotherapeutic Actions of MTOR Inhibitor in Genetically Defined Head and Neck Squamous Cell Carcinoma Mouse ModelRapamycin Prevents Tumorigenesis in HNSCC Mouse Model. Clin. Cancer Res. 2012, 18, 5304–5313. [Google Scholar] [CrossRef] [Green Version]
- Moore, E.C.; Cash, H.A.; Caruso, A.M.; Uppaluri, R.; Hodge, J.W.; Van Waes, C.; Allen, C.T. Enhanced Tumor Control with Combination MTOR and PD-L1 Inhibition in Syngeneic Oral Cavity CancersCombination MTOR and PD-L1 Checkpoint Inhibition. Cancer Immunol. Res. 2016, 4, 611–620. [Google Scholar] [CrossRef]
- Bozec, A.; Ebran, N.; Radosevic-Robin, N.; Sudaka, A.; Monteverde, M.; Toussan, N.; Etienne-Grimaldi, M.-C.; Nigro, C.L.; Merlano, M.; Penault-Llorca, F. Combination of m TOR and EGFR Targeting in an Orthotopic Xenograft Model of Head and Neck Cancer. Laryngoscope 2016, 126, E156–E163. [Google Scholar] [CrossRef]
- Li, Z.; Yang, Z.; Passaniti, A.; Lapidus, R.G.; Liu, X.; Cullen, K.J.; Dan, H.C. A Positive Feedback Loop Involving EGFR/Akt/MTORC1 and IKK/NF-ΚB Regulates Head and Neck Squamous Cell Carcinoma Proliferation. Oncotarget 2016, 7, 31892. [Google Scholar] [CrossRef] [Green Version]
- Suda, T.; Hama, T.; Kondo, S.; Yuza, Y.; Yoshikawa, M.; Urashima, M.; Kato, T.; Moriyama, H. Copy Number Amplification of the PIK3CA Gene Is Associated with Poor Prognosis in Non-Lymph Node Metastatic Head and Neck Squamous Cell Carcinoma. BMC Cancer 2012, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Keysar, S.B.; Le, P.N.; Miller, B.; Jackson, B.C.; Eagles, J.R.; Nieto, C.; Kim, J.; Tang, B.; Glogowska, M.J.; Morton, J.J. Regulation of Head and Neck Squamous Cancer Stem Cells by PI3K and SOX2. J. Natl. Cancer Inst. 2017, 109, djw189. [Google Scholar] [CrossRef]
- Pedrero, J.M.G.; Carracedo, D.G.; Pinto, C.M.; Zapatero, A.H.; Rodrigo, J.P.; Nieto, C.S.; Gonzalez, M.V. Retracted: Frequent Genetic and Biochemical Alterations of the PI 3-K/AKT/PTEN Pathway in Head and Neck Squamous Cell Carcinoma. Int. J. Cancer 2005, 114, 242–248. [Google Scholar] [CrossRef]
- Georgy, S.R.; Cangkrama, M.; Srivastava, S.; Partridge, D.; Auden, A.; Dworkin, S.; McLean, C.A.; Jane, S.M.; Darido, C. Identification of a Novel Proto-Oncogenic Network in Head and Neck Squamous Cell Carcinoma. JNCI J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Y.; Hall, B.; Sun, Z.-J.; Molinolo, A.; Chen, W.; Gutkind, J.S.; Waes, C.V.; Kulkarni, A.B. Loss of TGF-β Signaling and PTEN Promotes Head and Neck Squamous Cell Carcinoma through Cellular Senescence Evasion and Cancer-Related Inflammation. Oncogene 2012, 31, 3322–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiaris, H.; Spandidos, D.A.; Jones, A.S.; Vaughan, E.D.; Field, J.K. Mutations, Expression and Genomic Instability of the H-Ras Proto-Oncogene in Squamous Cell Carcinomas of the Head and Neck. Br. J. Cancer 1995, 72, 123–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruicci, K.M.; Pinto, N.; Khan, M.I.; Yoo, J.; Fung, K.; MacNeil, D.; Mymryk, J.S.; Barrett, J.W.; Nichols, A.C. ERK-TSC2 Signalling in Constitutively-Active HRAS Mutant HNSCC Cells Promotes Resistance to PI3K Inhibition. Oral Oncol. 2018, 84, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Duan, X.; Liu, J.; Xiao, J.; Brown, R.E. Morphoproteomic Evidence of Constitutively Activated and Overexpressed MTOR Pathway in Cervical Squamous Carcinoma and High Grade Squamous Intraepithelial Lesions. Int. J. Clin. Exp. Pathol. 2009, 2, 249. [Google Scholar]
- Harada, H.; Andersen, J.S.; Mann, M.; Terada, N.; Korsmeyer, S.J. P70S6 Kinase Signals Cell Survival as Well as Growth, Inactivating the pro-Apoptotic Molecule BAD. Proc. Natl. Acad. Sci. USA 2001, 98, 9666–9670. [Google Scholar] [CrossRef]
- Ji, J.; Zheng, P.-S. Activation of MTOR Signaling Pathway Contributes to Survival of Cervical Cancer Cells. Gynecol. Oncol. 2010, 117, 103–108. [Google Scholar] [CrossRef]
- Faried, L.S.; Faried, A.; Kanuma, T.; Sano, T.; Nakazato, T.; Tamura, T.; Kuwano, H.; Minegishi, T. Predictive and Prognostic Role of Activated Mammalian Target of Rapamycin in Cervical Cancer Treated with Cisplatin-Based Neoadjuvant Chemotherapy. Oncol. Rep. 2006, 16, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Tan, W.H.; Liu, W.; Jin, Y.X.; Dong, D.D.; Zhao, X.J.; Liu, Q. Effects of MiR-214 on Cervical Cancer Cell Proliferation, Apoptosis and Invasion via Modulating PI3K/AKT/MTOR Signal Pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1891–1898. [Google Scholar]
- Liu, L.; Wang, Y.; Geng, C.; Wang, A.; Han, S.; You, X.; Sun, Y.; Zhang, J.; Lu, W.; Zhang, Y. CD155 Promotes the Progression of Cervical Cancer Cells Through AKT/MTOR and NF-ΚB Pathways. Front. Oncol. 2021, 2153. [Google Scholar] [CrossRef]
- Deng, J.; Bai, X.; Feng, X.; Ni, J.; Beretov, J.; Graham, P.; Li, Y. Inhibition of PI3K/Akt/MTOR Signaling Pathway Alleviates Ovarian Cancer Chemoresistance through Reversing Epithelial-Mesenchymal Transition and Decreasing Cancer Stem Cell Marker Expression. BMC Cancer 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Lv, X.; Liu, D.; Guo, H.; Yao, G.; Wang, L.; Liang, X.; Yang, Y. METTL3-Mediated Maturation of MiR-126-5p Promotes Ovarian Cancer Progression via PTEN-Mediated PI3K/Akt/MTOR Pathway. Cancer Gene Ther. 2021, 28, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tu, Z.; Zhang, J.; Shen, X.; Wan, B.; Li, Y.; Wang, A.; Zhao, L.; Hu, J.; Ma, N. Follicular Fluid-Derived Exosomal MicroRNA-18b-5p Regulates PTEN-Mediated PI3K/Akt/MTOR Signaling Pathway to Inhibit Polycystic Ovary Syndrome Development. Mol. Neurobiol. 2022, 59, 2520–2531. [Google Scholar] [CrossRef]
- Chang, L.; Graham, P.H.; Hao, J.; Ni, J.; Bucci, J.; Cozzi, P.J.; Kearsley, J.H.; Li, Y. Acquisition of Epithelial–Mesenchymal Transition and Cancer Stem Cell Phenotypes Is Associated with Activation of the PI3K/Akt/MTOR Pathway in Prostate Cancer Radioresistance. Cell Death Dis. 2013, 4, e875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-MTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef] [PubMed]
- Crumbaker, M.; Khoja, L.; Joshua, A.M. AR Signaling and the PI3K Pathway in Prostate Cancer. Cancers 2017, 9, 34. [Google Scholar] [CrossRef]
- Malik, S.N.; Brattain, M.; Ghosh, P.M.; Troyer, D.A.; Prihoda, T.; Bedolla, R.; Kreisberg, J.I. Immunohistochemical Demonstration of Phospho-Akt in High Gleason Grade Prostate Cancer. Clin. Cancer Res. 2002, 8, 1168–1171. [Google Scholar] [PubMed]
- Bertoldo, F.; Silvestris, F.; Ibrahim, T.; Cognetti, F.; Generali, D.; Ripamonti, C.I.; Amadori, D.; Colleoni, M.A.; Conte, P.; Del Mastro, L. Targeting Bone Metastatic Cancer: Role of the MTOR Pathway. Biochim. Biophys. Acta BBA—Rev. Cancer 2014, 1845, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, L.; Wang, H.; Su, Z.; Pang, X. Neuroinflammation and Central PI3K/Akt/MTOR Signal Pathway Contribute to Bone Cancer Pain. Mol. Pain 2019, 15, 1744806919830240. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Li, M.; Hock, J.; Yu, X. Hyperactivation of MTOR Critically Regulates Abnormal Osteoclastogenesis in Neurofibromatosis Type 1. J. Orthop. Res. 2012, 30, 144–152. [Google Scholar] [CrossRef]
- Ding, L.; Congwei, L.; Bei, Q.; Tao, Y.; Ruiguo, W.; Heze, Y.; Bo, D.; Zhihong, L. MTOR: An Attractive Therapeutic Target for Osteosarcoma? Oncotarget 2016, 7, 50805. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Gutkind, J.S. Human Tumor-Associated Viruses and New Insights into the Molecular Mechanisms of Cancer. Oncogene 2008, 27, S31–S42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, P.S.; Chang, Y. Why Do Viruses Cause Cancer? Highlights of the First Century of Human Tumour Virology. Nat. Rev. Cancer 2010, 10, 878–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wu, J.; Ling, M.T.; Zhao, L.; Zhao, K.-N. The Role of the PI3K/Akt/MTOR Signalling Pathway in Human Cancers Induced by Infection with Human Papillomaviruses. Mol. Cancer 2015, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewell, A.; Brown, B.; Biktasova, A.; Mills, G.B.; Lu, Y.; Tyson, D.R.; Issaeva, N.; Yarbrough, W.G. Reverse-Phase Protein Array Profiling of Oropharyngeal Cancer and Significance of PIK3CA Mutations in HPV-Associated Head and Neck CancerMutant PIK3CA in Head and Neck Cancer. Clin. Cancer Res. 2014, 20, 2300–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surviladze, Z.; Sterk, R.T.; DeHaro, S.A.; Ozbun, M.A. Cellular Entry of Human Papillomavirus Type 16 Involves Activation of the Phosphatidylinositol 3-Kinase/Akt/MTOR Pathway and Inhibition of Autophagy. J. Virol. 2013, 87, 2508–2517. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, S.; Bhanja Chowdhury, J.; Steele, R.; Ray, R.; Ray, R.B. Hepatitis C Virus Upregulates Beclin1 for Induction of Autophagy and Activates MTOR Signaling. J. Virol. 2012, 86, 8705–8712. [Google Scholar] [CrossRef]
- Aravinth, S.P.; Rajendran, S.; Li, Y.; Wu, M.; Wong, A.H.Y.; Schwarz, H. Epstein–Barr Virus-Encoded LMP1 Induces Ectopic CD137 Expression on Hodgkin and Reed–Sternberg Cells via the PI3K-AKT-MTOR Pathway. Leuk. Lymphoma 2019, 60, 2697–2704. [Google Scholar] [CrossRef]
- Wang, W.; Wen, Q.; Xu, L.; Xie, G.; Li, J.; Luo, J.; Chu, S.; Shi, L.; Huang, D.; Li, J. Activation of Akt/MTOR Pathway Is Associated with Poor Prognosis of Nasopharyngeal Carcinoma. PLoS ONE 2014, 9, e106098. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Yao, N. Correlation of the AKT/MTOR Signaling Pathway with the Clinicopathological Features and Prognosis of Nasopharyngeal Carcinoma. Eur. J. Histochem. EJH 2021, 65, 3304. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, Y.; Sun, X.; Cui, Q.; Bai, X.; Dong, G.; Gao, Z.; Wang, Y.; Gao, C.; Sun, S. The PI3K/AKT/MTOR Signaling Pathway Is Aberrantly Activated in Primary Central Nervous System Lymphoma and Correlated with a Poor Prognosis. BMC Cancer 2022, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, M.; Ippolito, T.; Mavis, C.; Gu, J.; Cairo, M.S.; Lim, M.S.; Hernandez-Ilizaliturri, F.; Barth, M.J. Pre-Clinical Activity of Targeting the PI3K/Akt/MTOR Pathway in Burkitt Lymphoma. Oncotarget 2018, 9, 21820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindel, A.; McConnell, I.; Windle, J.; Sabo, R.; Chesney, A.; Lai, G.; Mauro, A.; Al-Juhaishi, T.; Rahmani, M.; Zweit, J. Role of the PI3K Pathway in the Pathogenesis of Marginal Zone Lymphoma. Blood 2018, 132, 4125. [Google Scholar] [CrossRef]
- Okosun, J.; Wolfson, R.L.; Wang, J.; Araf, S.; Wilkins, L.; Castellano, B.M.; Escudero-Ibarz, L.; Al Seraihi, A.F.; Richter, J.; Bernhart, S.H. Recurrent MTORC1-Activating RRAGC Mutations in Follicular Lymphoma. Nat. Genet. 2016, 48, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Zhang, Y.; Chen, G.; Xie, Y.; Xu, Z.; Chang, S.; Hu, L.; Li, B.; Bu, W.; Wang, Y. Targeting the PI3K/Akt/MTOR Signaling Pathway by Pterostilbene Attenuates Mantle Cell Lymphoma Progression. Acta Biochim. Biophys. Sin. 2018, 50, 782–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Guo, S.; Fang, T.; Feng, D.; Zhang, X.; Zhang, Q.; Liu, J.; Liu, B.; Li, M.; Zhu, R. Dihydromyricetin Induces Autophagy in HepG2 Cells Involved in Inhibition of MTOR and Regulating Its Upstream Pathways. Food Chem. Toxicol. 2014, 66, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.-P.; Lee, C.-H.; Ying, T.-H.; Lin, C.-L.; Lin, C.-L.; Hsueh, J.-T.; Hsieh, Y.-H. Licochalcone A Induces Autophagy through PI3K/Akt/MTOR Inactivation and Autophagy Suppression Enhances Licochalcone A-Induced Apoptosis of Human Cervical Cancer Cells. Oncotarget 2015, 6, 28851–28866. [Google Scholar] [CrossRef]
- Kumar, S.; Agnihotri, N. Piperlongumine, a Piper Alkaloid Targets Ras/PI3K/Akt/MTOR Signaling Axis to Inhibit Tumor Cell Growth and Proliferation in DMH/DSS Induced Experimental Colon Cancer. Biomed. Pharmacother. 2019, 109, 1462–1477. [Google Scholar] [CrossRef]
- Acharya, B.; Chaijaroenkul, W.; Na-Bangchang, K. Atractylodin Inhibited the Migration and Induced Autophagy in Cholangiocarcinoma Cells via PI3K/AKT/MTOR and P38MAPK Signalling Pathways. J. Pharm. Pharmacol. 2021, 73, 1191–1200. [Google Scholar] [CrossRef]
- Chen, T.; Hao, J.; He, J.; Zhang, J.; Li, Y.; Liu, R.; Li, L. Cannabisin B Induces Autophagic Cell Death by Inhibiting the AKT/MTOR Pathway and S Phase Cell Cycle Arrest in HepG2 Cells. Food Chem. 2013, 138, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.-M.; Wang, X.-F.; Huang, Q.-X. Thymoquinone Induces Cytotoxicity and Reprogramming of EMT in Gastric Cancer Cells by Targeting PI3K/Akt/MTOR Pathway. J. Biosci. 2017, 42, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.R.; Min, K.-J.; Cho, I.J.; Kim, S.C.; Kwon, T.K. Curcumin Significantly Enhances Dual PI3K/Akt and MTOR Inhibitor NVP-BEZ235-Induced Apoptosis in Human Renal Carcinoma Caki Cells through down-Regulation of P53-Dependent Bcl-2 Expression and Inhibition of Mcl-1 Protein Stability. PLoS ONE 2014, 9, e95588. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.; Zhang, M.; Meng, H.; Xu, D.; Xie, Y. Gallic Acid Targets Acute Myeloid Leukemia via Akt/MTOR-Dependent Mitochondrial Respiration Inhibition. Biomed. Pharmacother. 2018, 105, 491–497. [Google Scholar] [CrossRef]
- Liu, Z.; Antalek, M.; Nguyen, L.; Li, X.; Tian, X.; Le, A.; Zi, X. The Effect of Gartanin, a Naturally- Occurring Xanthone in Mangosteen Juice, on the MTOR Pathway, Autophagy, Apoptosis and the Growth of Human Urinary Bladder Cancer Cell Lines. Nutr. Cancer 2013, 65, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ha, U.-S.; Yu, K.; Wu, C.; Yokoyama, N.; Zi, X. Kavalactone Yangonin Induces Autophagy and Sensitizes Bladder Cancer Cells to Flavokawain A and Docetaxel via Inhibition of the MTOR Pathway. J. Biomed. Res. 2017, 31, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Alayev, A.; Salamon, R.S.; Schwartz, N.S.; Berman, A.Y.; Wiener, S.L.; Holz, M.K. Combination of Rapamycin and Resveratrol for Treatment of Bladder Cancer. J. Cell. Physiol. 2017, 232, 436–446. [Google Scholar] [CrossRef]
- Zhou, C.; Ding, J.; Wu, Y. Resveratrol Induces Apoptosis of Bladder Cancer Cells via MiR-21 Regulation of the Akt/Bcl-2 Signaling Pathway. Mol. Med. Rep. 2014, 9, 1467–1473. [Google Scholar] [CrossRef]
- He, S.-Q.; Gao, M.; Fu, Y.-F.; Zhang, Y.-N. Glycyrrhizic Acid Inhibits Leukemia Cell Growth and Migration via Blocking AKT/MTOR/STAT3 Signaling. Int. J. Clin. Exp. Pathol. 2015, 8, 5175–5181. [Google Scholar]
- Liu, J.; Zheng, L.; Zhong, J.; Wu, N.; Liu, G.; Lin, X. Oleanolic Acid Induces Protective Autophagy in Cancer Cells through the JNK and MTOR Pathways. Oncol. Rep. 2014, 32, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Zhang, Z.; Xu, J.; Liang, X.; Zhao, Q. Poricoic Acid A Induces Apoptosis and Autophagy in Ovarian Cancer via Modulating the MTOR/P70s6k Signaling Axis. Braz. J. Med. Biol. Res. 2021, 54, e11183. [Google Scholar] [CrossRef]
- Hong, P.; Liu, Q.-W.; Xie, Y.; Zhang, Q.-H.; Liao, L.; He, Q.-Y.; Li, B.; Xu, W.W. Echinatin Suppresses Esophageal Cancer Tumor Growth and Invasion through Inducing AKT/MTOR-Dependent Autophagy and Apoptosis. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, Q.; Li, H.; Yang, B.; Wang, M. Vitexin Suppresses Renal Cell Carcinoma by Regulating MTOR Pathways. Transl. Androl. Urol. 2020, 9, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, C.; Duanmu, J.; Wu, Y.; Tao, J.; Yang, A.; Yin, X.; Xiong, B.; Gu, J.; Li, C.; et al. Cryptotanshinone Inhibits the Growth and Invasion of Colon Cancer by Suppressing Inflammation and Tumor Angiogenesis through Modulating MMP/TIMP System, PI3K/Akt/MTOR Signaling and HIF-1alpha Nuclear Translocation. Int. Immunopharmacol. 2018, 65, 429–437. [Google Scholar] [CrossRef]
- Yun, S.-M.; Jung, J.H.; Jeong, S.-J.; Sohn, E.J.; Kim, B.; Kim, S.-H. Tanshinone IIA Induces Autophagic Cell Death via Activation of AMPK and ERK and Inhibition of MTOR and P70 S6K in KBM-5 Leukemia Cells. Phytother. Res. 2013, 28, 458–464. [Google Scholar] [CrossRef]
- Yang, L.; Xie, J.; Almoallim, H.S.; Alharbi, S.A.; Chen, Y. Tomentosin Inhibits Cell Proliferation and Induces Apoptosis in MOLT-4 Leukemia Cancer Cells through the Inhibition of MTOR/PI3K/Akt Signaling Pathway. J. Biochem. Mol. Toxicol. 2021, 35, e22719. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, Y.; Cai, Y.; Shui, C.; Liu, W.; Wang, X.; Jiang, J.; Zeng, D.; Gui, C.; Sun, R. Parthenolide Inhibits the Proliferation of MDA-T32 Papillary Thyroid Carcinoma Cells in Vitro and in Mouse Tumor Xenografts and Activates Autophagy and Apoptosis by Downregulation of the Mammalian Target of Rapamycin (MTOR)/PI3K/AKT Signaling Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 5054. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tang, X.; Liu, T.; Jing, L.; Wu, J. Zingiberene Inhibits in Vitro and in Vivo Human Colon Cancer Cell Growth via Autophagy Induction, Suppression of PI3K/AKT/MTOR Pathway and Caspase 2 Deactivation. J. Buon. 2019, 24, 1470–1475. [Google Scholar]
- Totiger, T.M.; Srinivasan, S.; Jala, V.R.; Lamichhane, P.; Dosch, A.R.; Gaidarski, A.A.; Joshi, C.; Rangappa, S.; Castellanos, J.; Vemula, P.K. Urolithin A, a Novel Natural Compound to Target PI3K/AKT/MTOR Pathway in Pancreatic CancerTargeting PI3K/AKT/MTOR Pathway in Pancreatic Cancer. Mol. Cancer Ther. 2019, 18, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Lin, Y.; Mok, D.K.-W.; Bian, Q.; Tai, W.C.-S.; Chen, S. Arnicolide D Inhibits Triple Negative Breast Cancer Cell Proliferation by Suppression of Akt/MTOR and STAT3 Signaling Pathways. Int. J. Med. Sci. 2020, 17, 1482. [Google Scholar] [CrossRef]
- Park, K.-R.; Leem, H.H.; Kwon, Y.-J.; Kwon, I.K.; Hong, J.T.; Yun, H.-M. Falcarindiol Stimulates Apoptotic and Autophagic Cell Death to Attenuate Cell Proliferation, Cell Division, and Metastasis through the PI3K/AKT/MTOR/P70S6K Pathway in Human Oral Squamous Cell Carcinomas. Am. J. Chin. Med. 2022, 50, 295–311. [Google Scholar] [CrossRef]
- Roy, G.; Guan, S.; Liu, H.; Zhang, L. Rotundic Acid Induces DNA Damage and Cell Death in Hepatocellular Carcinoma through AKT/MTOR and MAPK Pathways. Front. Oncol. 2019, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, X.; Pan, H.; He, S.; Yuan, B.; Liu, Q.; Zhang, J.; Ding, Y. Eupafolin Inhibits Breast Cancer Cell Proliferation and Induces Apoptosis by Inhibiting the PI3K/Akt/MTOR Pathway. Oncol. Lett. 2021, 21, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yin, J.; Yan, S.; Hu, P.; Huang, J.; Zhang, G.; Wang, F.; Tong, Q.; Zhang, Y. Chaetocochin J, an Epipolythiodioxopiperazine Alkaloid, Induces Apoptosis and Autophagy in Colorectal Cancer via AMPK and PI3K/AKT/MTOR Pathways. Bioorganic Chem. 2021, 109, 104693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yi, J.-K.; Huang, H.; Park, S.; Park, S.; Kwon, W.; Kim, E.; Jang, S.; Kim, S.-Y.; Choi, S.-K. Rhein Suppresses Colorectal Cancer Cell Growth by Inhibiting the MTOR Pathway In Vitro and In Vivo. Cancers 2021, 13, 2176. [Google Scholar] [CrossRef]
- Karbowniczek, M.; Spittle, C.S.; Morrison, T.; Wu, H.; Henske, E.P. MTOR Is Activated in the Majority of Malignant Melanomas. J. Investig. Dermatol. 2008, 128, 980–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera Aguilera, J.; Rao, R.D.; Allred, J.B.; Suman, V.J.; Windschitl, H.E.; Kaur, J.S.; Maples, W.J.; Lowe, V.J.; Creagan, E.T.; Erickson, L.A. Phase II Study of Everolimus in Metastatic Malignant Melanoma (NCCTG-N0377, Alliance). Oncologist 2018, 23, 887-e94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, L.; Xu, X.; Kong, Y.; Flaherty, K.T.; Chi, Z.; Cui, C.; Sheng, X.; Li, S.; Dai, J.; Yu, W. Major Response to Everolimus in Melanoma with Acquired Imatinib Resistance. J. Clin. Oncol. 2012, 30, e37–e40. [Google Scholar] [CrossRef]
- Hainsworth, J.D.; Infante, J.R.; Spigel, D.R.; Peyton, J.D.; Thompson, D.S.; Lane, C.M.; Clark, B.L.; Rubin, M.S.; Trent, D.F.; Burris III, H.A. Bevacizumab and Everolimus in the Treatment of Patients with Metastatic Melanoma: A Phase 2 Trial of the Sarah Cannon Oncology Research Consortium. Cancer 2010, 116, 4122–4129. [Google Scholar] [CrossRef]
- Niessner, H.; Kosnopfel, C.; Sinnberg, T.; Beck, D.; Krieg, K.; Wanke, I.; Lasithiotakis, K.; Bonin, M.; Garbe, C.; Meier, F. Combined Activity of Temozolomide and the MTOR Inhibitor Temsirolimus in Metastatic Melanoma Involves DKK1. Exp. Dermatol. 2017, 26, 598–606. [Google Scholar] [CrossRef]
- Rangwala, R.; Chang, Y.C.; Hu, J.; Algazy, K.M.; Evans, T.L.; Fecher, L.A.; Schuchter, L.M.; Torigian, D.A.; Panosian, J.T.; Troxel, A.B. Combined MTOR and Autophagy Inhibition: Phase I Trial of Hydroxychloroquine and Temsirolimus in Patients with Advanced Solid Tumors and Melanoma. Autophagy 2014, 10, 1391–1402. [Google Scholar] [CrossRef]
- Kolev, V.N.; Wright, Q.G.; Vidal, C.M.; Ring, J.E.; Shapiro, I.M.; Ricono, J.; Weaver, D.T.; Padval, M.V.; Pachter, J.A.; Xu, Q. PI3K/MTOR Dual Inhibitor VS-5584 Preferentially Targets Cancer Stem Cells. Cancer Res. 2015, 75, 446–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickland, L.R.; Pal, H.C.; Elmets, C.A.; Afaq, F. Targeting Drivers of Melanoma with Synthetic Small Molecules and Phytochemicals. Cancer Lett. 2015, 359, 20–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schadendorf, D.; Hauschild, A. Melanoma—The Run of Success Continues. Nat. Rev. Clin. Oncol. 2014, 11, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Bao, Q.; Jiang, F.; Qian, H.; Fang, Q.; Hu, X. VS-5584, a Novel PI3K-MTOR Dual Inhibitor, Inhibits Melanoma Cell Growth in Vitro and in Vivo. PLoS ONE 2015, 10, e0132655. [Google Scholar] [CrossRef]
- Webster, R.M.; Mentzer, S.E. The Malignant Melanoma Landscape. Nat. Rev. Drug Discov. 2014, 13, 491. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.; Wen, J.; Ma, F.; Wang, F.; Yu, K.; Tang, M.; Wu, W.; Dong, Y.; Cheng, X. SKLB-M8 Induces Apoptosis through the AKT/MTOR Signaling Pathway in Melanoma Models and Inhibits Angiogenesis with Decrease of ERK1/2 Phosphorylation. J. Pharmacol. Sci. 2014, 14077FP. [Google Scholar] [CrossRef] [Green Version]
- Head, S.A.; Shi, W.; Zhao, L.; Gorshkov, K.; Pasunooti, K.; Chen, Y.; Deng, Z.; Li, R.; Shim, J.S.; Tan, W. Antifungal Drug Itraconazole Targets VDAC1 to Modulate the AMPK/MTOR Signaling Axis in Endothelial Cells. Proc. Natl. Acad. Sci. USA 2015, 112, E7276–E7285. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Liu, M.; Wang, Q.; Shen, Y.; Mei, H.; Li, D.; Liu, W. Itraconazole Exerts Its Anti-Melanoma Effect by Suppressing Hedgehog, Wnt, and PI3K/MTOR Signaling Pathways. Oncotarget 2017, 8, 28510. [Google Scholar] [CrossRef] [Green Version]
- Oleksak, P.; Nepovimova, E.; Chrienova, Z.; Musilek, K.; Patocka, J.; Kuca, K. Contemporary MTOR Inhibitor Scaffolds to Diseases Breakdown: A Patent Review (2015–2021). Eur. J. Med. Chem. 2022, 238, 114498. [Google Scholar] [CrossRef]
- Hua, H.; Kong, Q.; Zhang, H.; Wang, J.; Luo, T.; Jiang, Y. Targeting MTOR for Cancer Therapy. J. Hematol. Oncol. 2019, 12, 1–19. [Google Scholar] [CrossRef]
- Xie, J.; Wang, X.; Proud, C.G. MTOR Inhibitors in Cancer Therapy. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Ye, L.; Araki, K.; Ahmed, R. MTOR, Linking Metabolism and Immunity. In Seminars in Immunology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, pp. 429–435. [Google Scholar]
- Zeng, H. MTOR Signaling in Immune Cells and Its Implications for Cancer Immunotherapy. Cancer Lett. 2017, 408, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Pollizzi, K.N.; Patel, C.H.; Sun, I.-H.; Oh, M.-H.; Waickman, A.T.; Wen, J.; Delgoffe, G.M.; Powell, J.D. MTORC1 and MTORC2 Selectively Regulate CD8+ T Cell Differentiation. J. Clin. Investig. 2015, 125, 2090–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Chi, H. MTOR and Metabolic Pathways in T Cell Quiescence and Functional Activation. In Seminars in Immunology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 24, pp. 421–428. [Google Scholar]
- Delgoffe, G.M.; Kole, T.P.; Zheng, Y.; Zarek, P.E.; Matthews, K.L.; Xiao, B.; Worley, P.F.; Kozma, S.C.; Powell, J.D. The MTOR Kinase Differentially Regulates Effector and Regulatory T Cell Lineage Commitment. Immunity 2009, 30, 832–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgoffe, G.M.; Pollizzi, K.N.; Waickman, A.T.; Heikamp, E.; Meyers, D.J.; Horton, M.R.; Xiao, B.; Worley, P.F.; Powell, J.D. The Kinase MTOR Regulates the Differentiation of Helper T Cells through the Selective Activation of Signaling by MTORC1 and MTORC2. Nat. Immunol. 2011, 12, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.Z.; Wang, R.; Huang, G.; Vogel, P.; Neale, G.; Green, D.R.; Chi, H. HIF1α–Dependent Glycolytic Pathway Orchestrates a Metabolic Checkpoint for the Differentiation of TH17 and Treg Cells. J. Exp. Med. 2011, 208, 1367–1376. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J. The Transcription Factor Myc Controls Metabolic Reprogramming upon T Lymphocyte Activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Templeton, A.J.; Dutoit, V.; Cathomas, R.; Rothermundt, C.; Bärtschi, D.; Dröge, C.; Gautschi, O.; Borner, M.; Fechter, E.; Stenner, F. Phase 2 Trial of Single-Agent Everolimus in Chemotherapy-Naive Patients with Castration-Resistant Prostate Cancer (SAKK 08/08). Eur. Urol. 2013, 64, 150–158. [Google Scholar] [CrossRef]
- Kurebayashi, Y.; Nagai, S.; Ikejiri, A.; Ohtani, M.; Ichiyama, K.; Baba, Y.; Yamada, T.; Egami, S.; Hoshii, T.; Hirao, A. PI3K-Akt-MTORC1-S6K1/2 Axis Controls Th17 Differentiation by Regulating Gfi1 Expression and Nuclear Translocation of RORγ. Cell Rep. 2012, 1, 360–373. [Google Scholar] [CrossRef]
- Chapman, N.M.; Chi, H. MTOR Signaling, Tregs and Immune Modulation. Immunotherapy 2014, 6, 1295–1311. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, W.; Beckett, O.; Ma, Q.; Paik, J.; DePinho, R.A.; Li, M.O. Foxo Proteins Cooperatively Control the Differentiation of Foxp3+ Regulatory T Cells. Nat. Immunol. 2010, 11, 618–627. [Google Scholar] [CrossRef]
- Huynh, A.; DuPage, M.; Priyadharshini, B.; Sage, P.T.; Quiros, J.; Borges, C.M.; Townamchai, N.; Gerriets, V.A.; Rathmell, J.C.; Sharpe, A.H. Control of PI (3) Kinase in Treg Cells Maintains Homeostasis and Lineage Stability. Nat. Immunol. 2015, 16, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Yang, K.; Guy, C.; Vogel, P.; Neale, G.; Chi, H. Treg Cells Require the Phosphatase PTEN to Restrain TH1 and TFH Cell Responses. Nat. Immunol. 2015, 16, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, I.-H.; Oh, M.-H.; Zhao, L.; Patel, C.H.; Arwood, M.L.; Xu, W.; Tam, A.J.; Blosser, R.L.; Wen, J.; Powell, J.D. MTOR Complex 1 Signaling Regulates the Generation and Function of Central and Effector Foxp3+ Regulatory T Cells. J. Immunol. 2018, 201, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 3151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, F.; Zhou, P.; Wang, Q.; Xu, C.; Li, Y.; Bian, L.; Liu, Y.; Zhou, J.; Wang, F. The MTOR Signaling Pathway Regulates Macrophage Differentiation from Mouse Myeloid Progenitors by Inhibiting Autophagy. Autophagy 2019, 15, 1150–1162. [Google Scholar] [CrossRef]
- Jiang, H.; Westerterp, M.; Wang, C.; Zhu, Y.; Ai, D. Macrophage MTORC1 Disruption Reduces Inflammation and Insulin Resistance in Obese Mice. Diabetologia 2014, 57, 2393–2404. [Google Scholar] [CrossRef]
- Mercalli, A.; Calavita, I.; Dugnani, E.; Citro, A.; Cantarelli, E.; Nano, R.; Melzi, R.; Maffi, P.; Secchi, A.; Sordi, V. Rapamycin Unbalances the Polarization of Human Macrophages to M 1. Immunology 2013, 140, 179–190. [Google Scholar] [CrossRef]
- Byles, V.; Covarrubias, A.J.; Ben-Sahra, I.; Lamming, D.W.; Sabatini, D.M.; Manning, B.D.; Horng, T. The TSC-MTOR Pathway Regulates Macrophage Polarization. Nat. Commun. 2013, 4, 1–11. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-Derived Suppressor Cells as Regulators of the Immune System. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Conciatori, F.; Bazzichetto, C.; Falcone, I.; Pilotto, S.; Bria, E.; Cognetti, F.; Milella, M.; Ciuffreda, L. Role of MTOR Signaling in Tumor Microenvironment: An Overview. Int. J. Mol. Sci. 2018, 19, 2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, R.; Asif, M.; Singh, V.; Dubey, P.; Malik, S.A.; Tewari, B.N.; Baghel, K.S.; Pal, S.; Nagar, G.K.; Chattopadhyay, N. M2 Polarization of Macrophages by Oncostatin M in Hypoxic Tumor Microenvironment Is Mediated by MTORC2 and Promotes Tumor Growth and Metastasis. Cytokine 2019, 118, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Welte, T.; Kim, I.S.; Tian, L.; Gao, X.; Wang, H.; Li, J.; Holdman, X.B.; Herschkowitz, J.I.; Pond, A.; Xie, G. Oncogenic MTOR Signalling Recruits Myeloid-Derived Suppressor Cells to Promote Tumour Initiation. Nat. Cell Biol. 2016, 18, 632–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryzhov, S.V.; Pickup, M.W.; Chytil, A.; Gorska, A.E.; Zhang, Q.; Owens, P.; Feoktistov, I.; Moses, H.L.; Novitskiy, S.V. Role of TGF-β Signaling in Generation of CD39+ CD73+ Myeloid Cells in Tumors. J. Immunol. 2014, 193, 3155–3164. [Google Scholar] [CrossRef] [Green Version]
- Farkona, S.; Diamandis, E.P.; Blasutig, I.M. Cancer Immunotherapy: The Beginning of the End of Cancer? BMC Med. 2016, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chen, M.; Nie, H.; Yuan, Y. PD-1 and PD-L1 in Cancer Immunotherapy: Clinical Implications and Future Considerations. Hum. Vaccines Immunother. 2019, 15, 1111–1122. [Google Scholar] [CrossRef]
- Li, J.-X.; Huang, J.-M.; Jiang, Z.-B.; Li, R.-Z.; Sun, A.; Lai-Han Leung, E.; Yan, P.-Y. Current Clinical Progress of PD-1/PD-L1 Immunotherapy and Potential Combination Treatment in Non–Small Cell Lung Cancer. Integr. Cancer Ther. 2019, 18, 1534735419890020. [Google Scholar] [CrossRef] [PubMed]
- McDermott, D.F.; Atkins, M.B. PD-1 as a Potential Target in Cancer Therapy. Cancer Med. 2013, 2, 662–673. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Smyth, M.J.; Teng, M.W. Acquired Resistance to Anti-PD1 Therapy: Checkmate to Checkpoint Blockade? Genome Med. 2016, 8, 1–3. [Google Scholar] [CrossRef]
- Murciano-Goroff, Y.R.; Warner, A.B.; Wolchok, J.D. The Future of Cancer Immunotherapy: Microenvironment-Targeting Combinations. Cell Res. 2020, 30, 507–519. [Google Scholar] [CrossRef]
- Ventola, C.L. Cancer Immunotherapy, Part 3: Challenges and Future Trends. Pharm. Ther. 2017, 42, 514. [Google Scholar]
- Chang, J.T.; Wherry, E.J.; Goldrath, A.W. Molecular Regulation of Effector and Memory T Cell Differentiation. Nat. Immunol. 2014, 15, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Hargadon, K.M.; Johnson, C.E.; Williams, C.J. Immune Checkpoint Blockade Therapy for Cancer: An Overview of FDA-Approved Immune Checkpoint Inhibitors. Int. Immunopharmacol. 2018, 62, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Akdis, M.; Blaser, K.; Akdis, C.A. Mechanisms of Allergen Specific Immunotherapy–T-Cell Tolerance and More. Allergy 2006, 61, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-C.; Hung, S.-K.; Lin, H.-Y.; Chiou, W.-Y.; Lee, M.-S.; Liao, H.-F.; Huang, H.-B.; Ho, H.-C.; Su, Y.-C. Targeting the PI3K/AKT/MTOR Signaling Pathway as an Effectively Radiosensitizing Strategy for Treating Human Oral Squamous Cell Carcinoma in Vitro and in Vivo. Oncotarget 2017, 8, 68641. [Google Scholar] [CrossRef]
- Yu, L.; Wei, J.; Liu, P. Attacking the PI3K/Akt/MTOR Signaling Pathway for Targeted Therapeutic Treatment in Human Cancer. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Wang, Y.; Wang, X.Y.; Subjeck, J.R.; Shrikant, P.A.; Kim, H.L. Temsirolimus, an MTOR Inhibitor, Enhances Anti-Tumour Effects of Heat Shock Protein Cancer Vaccines. Br. J. Cancer 2011, 104, 643–652. [Google Scholar] [CrossRef]
- Diken, M.; Kreiter, S.; Vascotto, F.; Selmi, A.; Attig, S.; Diekmann, J.; Huber, C.; Türeci, Ö.; Sahin, U. MTOR Inhibition Improves Antitumor Effects of Vaccination with Antigen-Encoding RNAmTOR Inhibition by Rapamycin Augments RNA Vaccine Efficacy. Cancer Immunol. Res. 2013, 1, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Chaoul, N.; Fayolle, C.; Desrues, B.; Oberkampf, M.; Tang, A.; Ladant, D.; Leclerc, C. Rapamycin Impairs Antitumor CD8+ T-Cell Responses and Vaccine-Induced Tumor EradicationRapamycin Impedes Vaccine-Induced Tumor Rejection. Cancer Res. 2015, 75, 3279–3291. [Google Scholar] [CrossRef] [Green Version]
- Langdon, S.; Hughes, A.; Taylor, M.A.; Kuczynski, E.A.; Mele, D.A.; Delpuech, O.; Jarvis, L.; Staniszewska, A.; Cosulich, S.; Carnevalli, L.S. Combination of Dual MTORC1/2 Inhibition and Immune-Checkpoint Blockade Potentiates Anti-Tumour Immunity. Oncoimmunology 2018, 7, e1458810. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sparwasser, T.L.; Figlin, R.; Kim, H.L. Abstract LB-260: Combination of Foxp3+ T Cells Depletion and MTOR Inhibitor Enhances Immune Memory and Controls Cancer Growth. Cancer Res. 2014, 74, LB-260. [Google Scholar] [CrossRef]
- Lastwika, K.J.; Wilson, W.; Li, Q.K.; Norris, J.; Xu, H.; Ghazarian, S.R.; Kitagawa, H.; Kawabata, S.; Taube, J.M.; Yao, S. Control of PD-L1 Expression by Oncogenic Activation of the AKT–MTOR Pathway in Non–Small Cell Lung CancerControl of PD-L1 by Oncogenic Activation of AKT/MTOR in NSCLC. Cancer Res. 2016, 76, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Li, X.; Liu, S.; Guo, L.; Zhang, B.; Zhang, J.; Ye, Q. PD-1 Checkpoint Blockade in Combination with an MTOR Inhibitor Restrains Hepatocellular Carcinoma Growth Induced by Hepatoma Cell-Intrinsic PD-1. Hepatology 2017, 66, 1920–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Weiss, J.M.; Back, T.; Chan, T.; Ortaldo, J.R.; Guichard, S.; Wiltrout, R.H. MTOR Kinase Inhibitor AZD8055 Enhances the Immunotherapeutic Activity of an Agonist CD40 Antibody in Cancer TreatmentAntitumor Effect of AZD8055/ACD40. Cancer Res. 2011, 71, 4074–4084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhao, X.; Sun, Q.; Jiang, Y.; Zhang, W.; Luo, J.; Li, Y. Synergic Effect of PD-1 Blockade and Endostar on the PI3K/AKT/MTOR-Mediated Autophagy and Angiogenesis in Lewis Lung Carcinoma Mouse Model. Biomed. Pharmacother. 2020, 125, 109746. [Google Scholar] [CrossRef]
- Ingels, A.; Zhao, H.; Thong, A.E.; Saar, M.; Valta, M.P.; Nolley, R.; Santos, J.; Peehl, D.M. Preclinical Trial of a New Dual MTOR Inhibitor, MLN0128, Using Renal Cell Carcinoma Tumorgrafts. Int. J. Cancer 2014, 134, 2322–2329. [Google Scholar] [CrossRef]
- Beziaud, L.; Boullerot, L.; Tran, T.; Mansi, L.; Marie-Joseph, E.L.; Ravel, P.; Johannes, L.; Bayry, J.; Tartour, E.; Adotévi, O. Rapalog Combined with CCR4 Antagonist Improves Anticancer Vaccines Efficacy. Int. J. Cancer 2018, 143, 3008–3018. [Google Scholar] [CrossRef] [Green Version]
- Knutson, K.L.; Disis, M.L. Tumor Antigen-Specific T Helper Cells in Cancer Immunity and Immunotherapy. Cancer Immunol. Immunother. 2005, 54, 721–728. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Lin, H.-W.; Sun, N.-Y.; Yie, J.-C.; Hung, H.-C.; Chen, C.-A.; Sun, W.-Z.; Cheng, W.-F. MTOR Inhibitors Can Enhance the Anti-Tumor Effects of DNA Vaccines through Modulating Dendritic Cell Function in the Tumor Microenvironment. Cancers 2019, 11, 617. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, H.L. Vaccines for Melanoma and Renal Cell Carcinoma. In Seminars in Oncology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 39, pp. 263–275. [Google Scholar]
- O’Donnell, J.S.; Massi, D.; Teng, M.W.; Mandala, M. PI3K-AKT-MTOR Inhibition in Cancer Immunotherapy, Redux. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 48, pp. 91–103. [Google Scholar]
- El Hage, A.; Dormond, O. Combining MTOR Inhibitors and T Cell-Based Immunotherapies in Cancer Treatment. Cancers 2021, 13, 1359. [Google Scholar] [CrossRef]
- Shor, B.; Zhang, W.-G.; Toral-Barza, L.; Lucas, J.; Abraham, R.T.; Gibbons, J.J.; Yu, K. A New Pharmacologic Action of CCI-779 Involves FKBP12-Independent Inhibition of MTOR Kinase Activity and Profound Repression of Global Protein Synthesis. Cancer Res. 2008, 68, 2934–2943. [Google Scholar] [CrossRef]
- Yang, H.; Rudge, D.G.; Koos, J.D.; Vaidialingam, B.; Yang, H.J.; Pavletich, N.P. MTOR Kinase Structure, Mechanism and Regulation. Nature 2013, 497, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotwals, P.; Cameron, S.; Cipolletta, D.; Cremasco, V.; Crystal, A.; Hewes, B.; Mueller, B.; Quaratino, S.; Sabatos-Peyton, C.; Petruzzelli, L. Prospects for Combining Targeted and Conventional Cancer Therapy with Immunotherapy. Nat. Rev. Cancer 2017, 17, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Rowshanravan, B.; Halliday, N.; Sansom, D.M. CTLA-4: A Moving Target in Immunotherapy. Blood J. Am. Soc. Hematol. 2018, 131, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Pedicord, V.A.; Cross, J.R.; Montalvo-Ortiz, W.; Miller, M.L.; Allison, J.P. Friends Not Foes: CTLA-4 Blockade and MTOR Inhibition Cooperate during CD8+ T Cell Priming to Promote Memory Formation and Metabolic Readiness. J. Immunol. 2015, 194, 2089–2098. [Google Scholar] [CrossRef] [Green Version]
- Alsaab, H.O.; Sau, S.; Alzhrani, R.; Tatiparti, K.; Bhise, K.; Kashaw, S.K.; Iyer, A.K. PD-1 and PD-L1 Checkpoint Signaling Inhibition for Cancer Immunotherapy: Mechanism, Combinations, and Clinical Outcome. Front. Pharmacol. 2017, 8, 561. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Han, X. Anti-PD-1PD-L1 Ther. Hum. Cancer Past Present Future. J. Clin. Investig. 2015, 125, 3384–3391. [Google Scholar]
- Lastwika, K.J. Regulation and Consequences of PD-L1 Expression in Non-Small Cell Lung Cancer. Ph.D. Thesis, The George Washington University, Washington, DC, USA, 2014. [Google Scholar]
- Mittendorf, E.A.; Philips, A.V.; Meric-Bernstam, F.; Qiao, N.; Wu, Y.; Harrington, S.; Su, X.; Wang, Y.; Gonzalez-Angulo, A.M.; Akcakanat, A. PD-L1 Expression in Triple-Negative Breast Cancer. Cancer Immunol. Res. 2014, 2, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.; Yi, M.; Qin, S.; Chu, Q.; Zheng, X.; Wu, K. The Efficacy and Safety of Combination of PD-1 and CTLA-4 Inhibitors: A Meta-Analysis. Exp. Hematol. Oncol. 2019, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jarosz-Biej, M.; Smolarczyk, R.; Cichoń, T.; Ku\lach, N. Tumor Microenvironment as a “Game Changer” in Cancer Radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212. [Google Scholar] [CrossRef] [Green Version]
- Lindau, D.; Gielen, P.; Kroesen, M.; Wesseling, P.; Adema, G.J. The Immunosuppressive Tumour Network: Myeloid-Derived Suppressor Cells, Regulatory T Cells and Natural Killer T Cells. Immunology 2013, 138, 105–115. [Google Scholar] [CrossRef]


| Study | Cancer | Conclusion | References |
|---|---|---|---|
| Temsirolimus, (mTOR inhibitor) in combination with heat shock protein cancer vaccines | Renal cell carcinoma (RCC) and melanoma (B16) | Temsirolimus enhanced the anti-tumor activity of cancer vaccines. The enhanced anti-tumor activity associated with temsirolimus was immune-mediated. | [281] |
| The impact of rapamycin on the in vivo primed RNA vaccine | B16 melanoma | Treatment with rapamycin skews the vaccine-induced immune response toward the formation of a superior memory pool and results in a better response. | [282] |
| Impact of rapamycin on the antitumor efficacy of a human papilloma virus E7 peptide vaccine (CyaA-E7) | Cervical cancer TC-1 | In animals vaccinated with CyaA-E7, rapamycin administration had a negative impact on the recruitment of CD8+ T cells into TC-1 tumors along as well as the ability of the vaccine to reduce infiltration of T regulatory cells. | [283] |
| Effect of mTORC1/2 dual kinase inhibitor vistusertib (AZD2014) in combination with anti-CTLA-4 (αCTLA-4), αPD-1 or αPD-L1 immune checkpoint blockade | MC-38 or CT-26 | Vistusertib/immune checkpoint combination has been shown to reduce the occurrence of exhausted phenotype tumor-infiltrating lymphocytes (TILs), as well as increasing frequencies of activated Th1 polarized T-cells in tumors. | [284] |
| Study of Treg control in addition to mTOR inhibition in preclinical models. | B16-gp100 | The combination therapy enhanced CD8 memory formation and function. | [285] |
| Control of PD-L1 Expression by oncogenic activation of the AKT-mTOR pathway in non-small cell lung cancer | Lung adenocarcinomas and squamous cell carcinomas | In vitro and in vivo PD-L1 expression is regulated by the activation of the AKT-mTOR pathway. It has been demonstrated that mTOR is required for PD-L1 induction by both oncogenic and IFN-mediated pathways. | [286] |
| Combining mTOR and PD-L1 inhibition improves tumor control in syngeneic oral cavity cancers | Oral cancer | Rapamycin has been found to increase IFNγ production capacity in peripheral and tumor-infiltrating CD8 T cells. | [157] |
| The combination of rapamycin and programmed cell death-1 (PD-1) checkpoint inhibition prevents the growth of hepatocellular carcinoma | Hepatocellular carcinoma | Targeting mammalian target of rapamycin pathways in combination with PD-1 may result in increased antitumor efficacy in cancer patients. | [287] |
| Combination of αCD40 agonistic antibody and the ATP-competitive mTOR kinase inhibitory drug AZD8055 elicited synergistic anti-tumor responses. | Metastatic renal cell carcinoma | When combined with CD40 therapy, the ATP-competitive mTOR kinase inhibitor AZD8055 can cause a remodeling of the tumor immune milieu and result in the regression of a metastatic cancer that has already spread. | [288] |
| Lewis lung cancer mouse model: synergistic effects of PD-1 inhibition and endostar on PI3K/AKT/mTOR-mediated autophagy and angiogenesis | Lewis lung carcinoma | IFN-secretion increased, myeloid-derived suppressor cell (MDSC) accumulation decreased, pro-inflammatory cytokine IL-17 and immunosuppressive factor TGF-1 levels fell, and CD8+ T cell suppression was reversed as a result of this synergistic impact. | [289] |
| PI3K/AKT/mTOR pathway targeting natural compound urolithin A for pancreatic cancer. | Pancreatic ductal adenocarcinoma (PDAC) | Treatment of PDAC cells with Uro A improved survival of Ptf1aCre/+; LSL-KrasG12D/+; Tgfbr2flox/flox (PKT) mice compared to vehicle or gemcitabine therapy alone, successfully suppressed the formation of tumor xenografts, and prevented the phosphorylation of AKT and p70S6K in vitro. | [220] |
| Comparison of the powerful second-generation mTOR inhibitor MLN0128 with the drug temsirolimus | RCC growth and metastasis | Our findings showed that MLN0128 outperformed temsirolimus in preventing both metastases and primary RCC growth, providing strong evidence in favor of additional clinical research into dual mTOR inhibitors for the treatment of RCC. | [290] |
| Examined the interaction between temsirolimus and anticancer vaccinations. using a range of cancer vaccine designs (short and long peptides or the B subunit of Shiga toxin as an antigen delivery vector) | Melanoma, lung, and colon cancer. | We demonstrated that temsirolimus treatment effectively reduced tumor development and improved tumor-specific CD8 T-cell responses brought on by vaccination. | [291] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouyahya, A.; El Allam, A.; Aboulaghras, S.; Bakrim, S.; El Menyiy, N.; Alshahrani, M.M.; Al Awadh, A.A.; Benali, T.; Lee, L.-H.; El Omari, N.; et al. Targeting mTOR as a Cancer Therapy: Recent Advances in Natural Bioactive Compounds and Immunotherapy. Cancers 2022, 14, 5520. https://doi.org/10.3390/cancers14225520
Bouyahya A, El Allam A, Aboulaghras S, Bakrim S, El Menyiy N, Alshahrani MM, Al Awadh AA, Benali T, Lee L-H, El Omari N, et al. Targeting mTOR as a Cancer Therapy: Recent Advances in Natural Bioactive Compounds and Immunotherapy. Cancers. 2022; 14(22):5520. https://doi.org/10.3390/cancers14225520
Chicago/Turabian StyleBouyahya, Abdelhakim, Aicha El Allam, Sara Aboulaghras, Saad Bakrim, Naoual El Menyiy, Mohammed Merae Alshahrani, Ahmed Abdullah Al Awadh, Taoufiq Benali, Learn-Han Lee, Nasreddine El Omari, and et al. 2022. "Targeting mTOR as a Cancer Therapy: Recent Advances in Natural Bioactive Compounds and Immunotherapy" Cancers 14, no. 22: 5520. https://doi.org/10.3390/cancers14225520
APA StyleBouyahya, A., El Allam, A., Aboulaghras, S., Bakrim, S., El Menyiy, N., Alshahrani, M. M., Al Awadh, A. A., Benali, T., Lee, L. -H., El Omari, N., Goh, K. W., Ming, L. C., & Mubarak, M. S. (2022). Targeting mTOR as a Cancer Therapy: Recent Advances in Natural Bioactive Compounds and Immunotherapy. Cancers, 14(22), 5520. https://doi.org/10.3390/cancers14225520