
New Visions on Natural Products and Cancer Therapy: Autophagy and Related Regulatory Pathways

 ,
, 
 ,
, 
 , , , ,
, , , ,  ,
, 
 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Autophagy
3. Autophagy Databases
4. Physiological and Pathophysiological Roles of the Autophagic Machinery in Cancer
4.1. Autophagy in the Initial Stages of Cancer
4.2. Autophagy in Advanced Tumors
4.3. Functional Crosstalk between Apoptosis and Autophagy in Cancer
4.4. Functional Relationship between Autophagy and the UPR in Cancer
4.5. Interaction between the Immune System and Autophagy in Cancer Treatment
5. Natural Compounds and Cancer
5.1. The Effects of Natural Compounds on the Autophagy Signaling Pathway
5.1.1. Alkaloids
5.1.2. Terpenoids
5.1.3. Flavonoids and Phenolic Compounds
5.1.4. Other Natural Compounds
5.2. The Effects of Natural Compounds on the Apoptosis Signaling Pathway
5.2.1. Alkaloids
5.2.2. Flavonoids and Phenolic Compounds
5.2.3. Terpenoids
5.2.4. Other Natural Compounds
5.3. The Effects of Natural Compounds on the UPR Signaling Pathway
5.3.1. Alkaloids
5.3.2. Flavonoids and Phenolic Compounds
5.3.3. Terpenoids
5.3.4. Other Natural Compounds
6. Animal Studies
7. Clinical Trials
8. Drawbacks of Natural Compounds
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3-MA | 3-methyladenine |
| 5-FU | 5-fluorouracil |
| ABCB1 | ATP binding cassette subfamily B member1 |
| ACAC | acetyl-CoA carboxylase |
| ACD | autophagic cell death |
| AIFM | apoptosis-inducing factor mitochondria associated |
| AKT/Protein kinase B | AKT serine/threonine kinase |
| AMP | adenosine monophosphate |
| AMPK | 5′-AMP-activated protein kinase |
| AP-1 | activator protein 1 |
| AR | androgen receptor |
| ATF | activating transcription factor |
| ATG | autophagy related |
| ATG16L1 | autophagy-related 16 like 1 |
| ATP | adenosine triphosphate |
| ATP2A1 | ATPase sarcoplasmic/endoplasmic reticulum Ca2+ transporting 1 |
| AVOs | acidic vesicular organelles |
| BAD | BCL2 associated agonist of cell death |
| BAX | BCL2 associated X, apoptosis regulator |
| BBC3/PUMA | BCL2 binding component 3/p53 upregulated modulator of apoptosis |
| BCL2 | B-cell lymphoma 2 |
| BDMC | bisdemethoxycurcumin |
| BID | BH3 interacting domain death agonist |
| BIRC5 | baculoviral IAP repeat containing 5 |
| BNIP3 | BCL2 interacting protein 3 |
| BRAFi | potent inhibitors of v-Raf murine sarcoma viral oncogene homolog B |
| CALR | calreticulin |
| CAPN2 | calpain 2 |
| CASP3 | caspase 3 |
| CCNA | cyclin A |
| CCNB | cyclin B |
| CCND | cyclin D |
| CCNE | cyclin E |
| CDK2 | cyclin dependent kinase 2 |
| CDKN1A | cyclin dependent kinase inhibitor 1A |
| CMA | chaperone-mediated autophagy |
| CQ | chloroquine |
| CRPC | castration-resistant prostate cancer |
| CTL | cytotoxic T lymphocyte |
| CTNNB1 | catenin beta 1 |
| CYCS | cytochrome c, somatic |
| ΔΨm | mitochondrial membrane potential |
| DAPK | death associated protein kinase 1 |
| DDIT3/CHOP/GADD | DNA damage inducible transcript 3 |
| DMC | demethoxycurcumin |
| EAC | Ehrlich ascites carcinoma |
| EDEM | ER degradation enhancing alpha-mannosidase like protein |
| EHMT2 | euchromatic histone lysine methyltransferase 2 |
| EIF2A/eIF2α | eukaryotic translation initiation factor 2A |
| EIF2AK3/PERK | eukaryotic translation initiation factor 2 alpha kinase 3 |
| ER | endoplasmic reticulum |
| ERN1/IRE1 | endoplasmic reticulum to nucleus signaling 1 |
| FADD | Fas associated via death domain |
| FAS | Fas cell surface death receptor |
| FASLG | Fas ligand |
| FOXO1 | forkhead box O1 |
| GLB1 | galactosidase beta 1 |
| GSK3B | glycogen synthase kinase 3 beta |
| HepG2 | hepatocarcinoma cell line |
| HIF1A/HIF-1α | hypoxia inducible factor 1 subunit alpha |
| HK2 | hexokinase 2 |
| HNSCC | head and neck squamous cell carcinoma |
| HRAS | HRas proto-oncogene, GTPase |
| HSPA5/GRP78/BiP | heat shock protein family A (Hsp70) member 5 |
| HSPA8/HSC70 | heat shock protein family A (Hsp70) member 8 |
| HSP90B1/GRP94 | heat shock protein 90 beta family member 1 |
| IL6 | interleukin 6 |
| KRAS | KRAS proto-oncogene, GTPase |
| LAMP2 | lysosomal associated membrane protein 2 |
| LMS | leiomyosarcoma |
| LV | leucovorin calcium |
| MAP1LC3/LC3 | microtubule associated protein 1 light chain 3 |
| MAPK8/JNK1 | mitogen-activated protein kinase 8/Jun N-terminal kinase |
| MCL1 | MCL1 apoptosis regulator, BCL2 family member |
| MEG3 | maternally expressed 3 |
| MEKi | mitogen-activated protein kinase inhibitor |
| MGMT | O-6-methylguanine-DNA methyltransferase |
| MMP9 | matrix metallopeptidase 9 |
| MTA1 | metastasis associated 1 |
| MT-CO2 | mitochondrially encoded cytochrome c oxidase II |
| MTOR | mechanistic target of rapamycin kinase |
| MTORC1 | MTOR complex 1 |
| MTORC2 | MTOR complex 2 |
| NAC | N-acetyl-L-cysteine |
| NFE2L2/nrf2 | NFE2 like bZIP transcription factor 2 |
| NKT | natural killer T cell |
| NLR | NOD-like receptor |
| NOTCH1 | notch receptor 1 |
| NPC | nasopharyngeal carcinoma |
| NSCLC | non-small cell lung cancer |
| OSCC | oral squamous cell carcinoma |
| p-EGFR | phosphorylated epidermal growth factor receptor |
| PARP | poly(ADP-ribose) polymerase |
| PDE4A | phosphodiesterase 4A |
| PDI | protein disulfide isomerase |
| PDPK1 | 3-phosphoinositide dependent protein kinase 1 |
| PI3K | class I phosphoinositide 3-kinase |
| PIK3C3 | phosphatidylinositol 3-kinase catalytic subunit type 3 |
| PMAIP1/NOXA | phorbol-12-myristate-13-acetate-induced protein 1/(Latin for damage) |
| PPP1R15A/GADD34 | protein phosphatase 1 regulatory subunit 15A |
| PRKCD/PKCδ | protein kinase C delta |
| PtdIns3K | class III phosphatidylinositol 3-kinase |
| PtdIns3P | phosphatidylinositol-3-phosphate |
| PTEN | phosphatase and tensin homolog |
| RECK | reversion inducing cysteine rich protein with kazal motifs |
| RELA | RELA proto-oncogene, NF-kB subunit |
| RELB | RELB proto-oncogene, NF-kB subunit |
| RICTOR | RPTOR independent companion of mTOR complex 2 |
| RNA | ribonucleic acid |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| RPS6KA1 | ribosomal protein S6 kinase A1 |
| RPS6KB | ribosomal protein S6 kinase B |
| SCC | squamous cell carcinoma |
| SERPINB4 | serpin family B member 4 |
| SH3GLB1/Bif-1 | SH3 domain containing GRB2 like, endophilin B1 |
| siRNA | small-interfering ribonucleic acid |
| SIRT1 | sirtuin 1 |
| SQSTM1/p62 | sequestosome 1 |
| SMPD | sphingomyelin phosphodiesterase |
| STIM1 | stromal interaction molecule 1 |
| STK11 | serine/threonine kinase 11 |
| TAMs | tumor-associated macrophages |
| TFEB | transcription factor EB |
| TGM2 | transglutaminase 2 |
| TLR | toll like receptor |
| TNBC | triple-negative breast cancer |
| TNFRSF10A/TRAIL/Apo2L1 | TNF receptor superfamily member 10a |
| TRAF2 | TNF receptor associated factor 2 |
| Treg | regulatory T cell |
| TSC2 | TSC complex subunit 2 |
| ULK1 | unc-51 like autophagy activating kinase 1 |
| UPR | unfolded protein response |
| UV | ultraviolet |
| UVRAG | UV radiation resistance associated |
| VEGF | vascular endothelial growth factor |
| WIPI1 | WD repeat domain, phosphoinositide interacting 1 |
| WIPI2 | WD repeat domain, phosphoinositide interacting 2 |
| WNT | Wnt family member |
| XBP1 | X-box binding protein 1 |
References
- Xu, D.; Lao, Y.; Xu, N.; Hu, H.; Fu, W.; Tan, H.; Gu, Y.; Song, Z.; Cao, P.; Xu, H. Identification and characterization of anticancer compounds targeting apoptosis and autophagy from Chinese native Garcinia species. Planta Med. 2015, 81, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, G.; Sha, T.; Guo, J.; Li, W.; Lu, J.; Chen, X. Cucurbitacin B induces DNA damage and autophagy mediated by reactive oxygen species (ROS) in MCF-7 breast cancer cells. J. Nat. Med. 2015, 69, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, J.L. Plants Used Against Cancer: A Survey; Quarterman Publications: Lawrence, MA, USA, 1982. [Google Scholar]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalvand, A.; da Silva Rosa, S.C.; Ghavami, S.; Marzban, H. Potential role of TGFBeta and autophagy in early crebellum development. Biochem. Biophys. Rep. 2022, 32, 101358. [Google Scholar] [PubMed]
- Boya, P.; Reggiori, F.; Codogno, P. Emerging regulation and functions of autophagy. Nat. Cell Biol. 2013, 15, 713–720. [Google Scholar] [CrossRef]
- Sridhar, S.; Botbol, Y.; Macian, F.; Cuervo, A.M. Autophagy and disease: Always two sides to a problem. J. Pathol. 2012, 226, 255–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrbod, P.; Ande, S.R.; Alizadeh, J.; Rahimizadeh, S.; Shariati, A.; Malek, H.; Hashemi, M.; Glover, K.K.M.; Sher, A.A.; Coombs, K.M.; et al. The roles of apoptosis, autophagy and unfolded protein response in arbovirus, influenza virus, and HIV infections. Virulence 2019, 10, 376–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shojaei, S.; Koleini, N.; Samiei, E.; Aghaei, M.; Cole, L.K.; Alizadeh, J.; Islam, M.I.; Vosoughi, A.R.; Albokashy, M.; Butterfield, Y.; et al. Simvastatin increases temozolomide-induced cell death by targeting the fusion of autophagosomes and lysosomes. FASEB J. 2020, 287, 1005–1034. [Google Scholar] [CrossRef]
- Eghtedardoost, M.; Ghazanfari, T.; Sadeghipour, A.; Hassan, Z.M.; Ghanei, M.; Ghavami, S. Delayed effects of sulfur mustard on autophagy suppression in chemically-injured lung tissue. Int. Immunopharmacol. 2020, 80, 105896. [Google Scholar] [CrossRef]
- Deter, R.L.; De Duve, C. Influence of glucagon, an inducer of cellular autophagy, on some physical properties of rat liver lysosomes. J. Cell Biol. 1967, 33, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Harnett, M.M.; Pineda, M.A.; Latre de Late, P.; Eason, R.J.; Besteiro, S.; Harnett, W.; Langsley, G. From Christian de Duve to Yoshinori Ohsumi: More to autophagy than just dining at home. Biomed. J. 2017, 40, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Eshraghi, M.; Adlimoghaddam, A.; Mahmoodzadeh, A.; Sharifzad, F.; Yasavoli-Sharahi, H.; Lorzadeh, S.; Albensi, B.C.; Ghavami, S. Alzheimer’s Disease Pathogenesis: Role of Autophagy and Mitophagy Focusing in Microglia. Int. J. Mol. Sci. 2021, 22, 3330. [Google Scholar] [CrossRef]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 2010, 22, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J. Autophagy: From phenomenology to molecular understanding in less than a decade. Nat. Rev. Mol. Cell Biol. 2007, 8, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, M.; Zmarzly, N.; Grabarek, B.; Mazurek, U.; Muc-Wierzgon, M. Genes involved in the regulation of different types of autophagy and their participation in cancer pathogenesis. Oncotarget 2018, 9, 34413–34428. [Google Scholar] [CrossRef] [Green Version]
- Shojaei, S.; Suresh, M.; Klionsky, D.J.; Labouta, H.I.; Ghavami, S. Autophagy and SARS-CoV-2 infection: Apossible smart targeting of the autophagy pathway. Virulence 2020, 11, 805–810. [Google Scholar] [CrossRef]
- Marzban, H.; Del Bigio, M.R.; Alizadeh, J.; Ghavami, S.; Zachariah, R.M.; Rastegar, M. Cellular commitment in the developing cerebellum. Front. Cell. Neurosci. 2014, 8, 450. [Google Scholar] [CrossRef] [Green Version]
- Zeki, A.A.; Yeganeh, B.; Kenyon, N.J.; Post, M.; Ghavami, S. Autophagy in airway diseases: A new frontier in human asthma? Allergy 2016, 71, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, J.; Shojaei, S.; Sepanjnia, A.; Hashemi, M.; Eftekharpour, E.; Ghavami, S. Simultaneous detection of autophagy and epithelial to mesenchymal transition in the non-small cell lung cancer cells. Methods Mol. Biol. 2019, 1854, 87–103. [Google Scholar]
- Alizadeh, J.; Kochan, M.M.; Stewart, V.D.; Drewnik, D.A.; Hannila, S.S.; Ghavami, S. Inhibition of Autophagy Flux Promotes Secretion of Chondroitin Sulfate Proteoglycans in Primary Rat Astrocytes. Mol. Neurobiol. 2021, 58, 6077–6091. [Google Scholar] [CrossRef]
- Hanna, R.A.; Quinsay, M.N.; Orogo, A.M.; Giang, K.; Rikka, S.; Gustafsson, A.B. Microtubule-associated protein 1 light chain 3 (LC3) interacts with Bnip3 protein to selectively remove endoplasmic reticulum and mitochondria via autophagy. J. Biol. Chem. 2012, 287, 19094–19104. [Google Scholar] [CrossRef]
- Shpilka, T.; Weidberg, H.; Pietrokovski, S.; Elazar, Z. Atg8: An autophagy-related ubiquitin-like protein family. Genome Biol. 2011, 12, 226. [Google Scholar] [CrossRef] [PubMed]
- da Silva Rosa, S.C.; Martens, M.D.; Field, J.T.; Nguyen, L.; Kereliuk, S.M.; Hai, Y.; Chapman, D.; Diehl-Jones, W.; Aliani, M.; West, A.R.; et al. BNIP3L/Nix-induced mitochondrial fission, mitophagy, and impaired myocyte glucose uptake are abrogated by PRKA/PKA phosphorylation. Autophagy 2020, 17, 2257–2272. [Google Scholar] [CrossRef] [PubMed]
- Sattler, T.; Mayer, A. Cell-free reconstitution of microautophagic vacuole invagination and vesicle formation. J. Cell Biol. 2000, 151, 238–529. [Google Scholar] [CrossRef] [PubMed]
- Kunz, J.B.; Schwarz, H.; Mayer, A. Determination of four sequential stages during microautophagy in vitro. J. Biol. Chem. 2004, 279, 9987–9996. [Google Scholar] [CrossRef] [Green Version]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Microautophagy in mammalian cells: Revisiting a 40-year-old conundrum. Autophagy 2011, 7, 673–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, P.C.; Bartlett, J.J.; Pulinilkunnil, T. Lysosomal biology and function: Modern view of cellular debris bin. Cells 2020, 9, 1131. [Google Scholar] [CrossRef]
- Mortimore, G.E.; Lardeux, B.R.; Adams, C.E. Regulation of microautophagy and basal protein turnover in rat liver. Effects of short-term starvation. J. Biol. Chem. 1988, 263, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Kohler, V.; Aufschnaiter, A.; Buttner, S. Closing the gap: Membrane contact sites in the regulation of autophagy. Cells 2020, 9, 1184. [Google Scholar] [CrossRef] [PubMed]
- Dice, J.F. Peptide sequences that target cytosolic proteins for lysosomal proteolysis. Trends Biochem. Sci. 1990, 15, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Chun, Y.; Kim, J. Autophagy: An essential degradation program for cellular homeostasis and life. Cells 2018, 7, 278. [Google Scholar] [CrossRef]
- Saha, S.; Panigrahi, D.P.; Patil, S.; Bhutia, S.K. Autophagy in health and disease: A comprehensive review. Biomed. Pharmacother. 2018, 104, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Bar-Yosef, T.; Damri, O.; Agam, G. Dual role of autophagy in diseases of the central nervous system. Front. Cell. Neurosci. 2019, 13, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorburn, A. Autophagy and disease. J. Biol. Chem. 2018, 293, 5425–5430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Mizushima, N. Autophagy and human diseases. Cell Res. 2014, 24, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghavami, S.; Gupta, S.; Ambrose, E.; Hnatowich, M.; Freed, D.H.; Dixon, I.M. Autophagy and heart disease: Implications for cardiac ischemia-reperfusion damage. Curr. Mol. Med. 2014, 14, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Golbabapour, S.; Bagheri-Lankarani, K.; Ghavami, S.; Geramizadeh, B. Autoimmune hepatitis and stellate cells; an insight into the role of autophagy. Curr. Med. Chem. 2020, 27, 6073–6095. [Google Scholar] [CrossRef]
- Mokarram, P.; Ahmadi, M.; Shojaei, S.; Ghavami, S. Autophagy and cancer research in Iran. Autophagy 2019, 15, 2039–2042. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.; Yeganeh, B.; Zeki, A.A.; Shojaei, S.; Kenyon, N.J.; Ott, S.; Samali, A.; Patterson, J.; Alizadeh, J.; Moghadam, A.R.; et al. Autophagy and the unfolded protein response promote profibrotic effects of TGF-beta1 in human lung fibroblasts. Am. J. Physiol. Lung C 2018, 314, L493–L504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankarani, K.B.; Sepehrimanesh, M.; Seghatoleslam, S.F.; Hoseini, S.E.; Ghavami, S. Autophagy-related protein 7 level in patients with ulcerative colitis. Scand. J. Gastroenterol. 2017, 52, 468. [Google Scholar] [CrossRef] [PubMed]
- Yeganeh, B.; Rezaei Moghadam, A.; Alizadeh, J.; Wiechec, E.; Alavian, S.M.; Hashemi, M.; Geramizadeh, B.; Samali, A.; Bagheri Lankarani, K.; Post, M.; et al. Hepatitis B and C virus-induced hepatitis: Apoptosis, autophagy, and unfolded protein response. World J. Gastroenterol. 2015, 21, 13225–13239. [Google Scholar] [CrossRef]
- Ghavami, S.; Shojaei, S.; Yeganeh, B.; Ande, S.R.; Jangamreddy, J.R.; Mehrpour, M.; Christoffersson, J.; Chaabane, W.; Moghadam, A.R.; Kashani, H.H.; et al. Autophagy and apoptosis dysfunction in neurodegenerative disorders. Prog. Neurobiol. 2014, 112, 24–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavian, S.M.; Ande, S.R.; Coombs, K.M.; Yeganeh, B.; Davoodpour, P.; Hashemi, M.; Los, M.; Ghavami, S. Virus-triggered autophagy in viral hepatitis-possible novel strategies for drug development. J. Viral. Hepat. 2011, 18, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasik, A.M.; Grabarek, J.; Pantovic, A.; Cieslar-Pobuda, A.; Asgari, H.R.; Bundgaard-Nielsen, C.; Rafat, M.; Dixon, I.M.; Ghavami, S.; Los, M.J. Reprogramming and carcinogenesis-parallels and distinctions. Int. Rev. Cell Mol. Biol. 2014, 308, 167–203. [Google Scholar]
- Wang, P.; Zhu, L.; Sun, D.; Gan, F.; Gao, S.; Yin, Y.; Chen, L. Natural products as modulator of autophagy with potential clinical prospects. Apoptosis 2017, 22, 325–356. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Biological functions of autophagy genes: A disease perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [Green Version]
- Homma, K.; Suzuki, K.; Sugawara, H. The autophagy database: An all-inclusive information resource on autophagy that provides nourishment for research. Nucleic Acids Res. 2011, 39, D986–D990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turei, D.; Foldvari-Nagy, L.; Fazekas, D.; Modos, D.; Kubisch, J.; Kadlecsik, T.; Demeter, A.; Lenti, K.; Csermely, P.; Vellai, T.; et al. Autophagy regulatory network-a systems-level bioinformatics resource for studying the mechanism and regulation of autophagy. Autophagy 2015, 11, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhu, L.; Cai, H.; Wang, G.; Liu, B. Autophagic compound database: A resource connecting autophagy-modulating compounds, their potential targets and relevant diseases. Cell Proliferat. 2018, 51, e12403. [Google Scholar] [CrossRef] [Green Version]
- Nanduri, R.; Kalra, R.; Bhagyaraj, E.; Chacko, A.P.; Ahuja, N.; Tiwari, D.; Kumar, S.; Jain, M.; Parkesh, R.; Gupta, P. AutophagySMDB: A curated database of small molecules that modulate protein targets regulating autophagy. Autophagy 2019, 15, 1280–1295. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Dong, J.; Zhang, L.; Ouyang, D.; Cheng, Y.; Chen, A.F.; Lu, A.P.; Cao, D.S. HAMdb: A database of human autophagy modulators with specific pathway and disease information. J. Cheminformatics 2018, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh, M.; Ashtari, N.; Jiao, X.; Rahimi Balaei, M.; Marzban, A.; Qiyami-Hour, F.; Kong, J.; Ghavami, S.; Marzban, H. Alteration of the dopamine receptors’ expression in the cerebellum of the lysosomal acid phosphatase 2 mutant (naked-ataxia (NAX)) mouse. Int. J. Mol. Sci. 2020, 21, 2914. [Google Scholar] [CrossRef]
- Alizadeh, J.; Glogowska, A.; Thliveris, J.; Kalantari, F.; Shojaei, S.; Hombach-Klonisch, S.; Klonisch, T.; Ghavami, S. Autophagy modulates transforming growth factor beta 1 induced epithelial to mesenchymal transition in non-small cell lung cancer cells. BBA Mol. Cell Res. 2018, 1865, 749–768. [Google Scholar] [CrossRef] [PubMed]
- Rikiishi, H. Novel insights into the interplay between apoptosis and autophagy. Int. J. Cell Biol. 2012, 2012, 317645. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Mei, Y.; Sinha, S. Role of the crosstalk between autophagy and apoptosis in cancer. J. Oncol. 2013, 2013, 102735. [Google Scholar] [CrossRef] [Green Version]
- Cuervo, A.M. Autophagy: Many paths to the same end. Mol. Cell. Biochem. 2004, 263, 55–72. [Google Scholar] [CrossRef]
- Shinozaki, D.; Merkulova, E.A.; Naya, L.; Horie, T.; Kanno, Y.; Seo, M.; Ohsumi, Y.; Masclaux-Daubresse, C.; Yoshimoto, K. Autophagy increases zinc bioavailability to avoid light-mediated reactive oxygen species production under zinc deficiency. Plant Physiol. 2020, 182, 1284–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Ding, L.; Chen, Y.; Shi, J. Augmenting tumor-starvation therapy by cancer cell autophagy inhibition. Adv. Sci. 2020, 7, 1902847. [Google Scholar] [CrossRef] [Green Version]
- Reggiori, F.; Klionsky, D.J. Autophagy in the eukaryotic cell. Eukaryot. Cell 2002, 1, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami, A.; Shojaei, S.; da Silva Rosa, S.C.; Aghaei, M.; Samiei, E.; Vosoughi, A.R.; Kalantari, F.; Kawalec, P.; Thliveris, J.; Sharma, P.; et al. Mechanisms of simvastatin myotoxicity: The role of autophagy flux inhibition. Eur. J. Pharmacol. 2019, 862, 172616. [Google Scholar] [CrossRef]
- Moghadam, A.R.; da Silva Rosa, S.C.; Samiei, E.; Alizadeh, J.; Field, J.; Kawalec, P.; Thliveris, J.; Akbari, M.; Ghavami, S.; Gordon, J.W. Autophagy modulates temozolomide-induced cell death in alveolar Rhabdomyosarcoma cells. Cell Death Discov. 2018, 4, 52. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Marino, G.; Levine, B. Autophagy and the integrated stress response. Mol. Cell Biochem. 2010, 40, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Karsli-Uzunbas, G.; Poillet-Perez, L.; Sawant, A.; Hu, Z.S.; Zhao, Y.; Moore, D.; Hu, W.; White, E. Autophagy promotes mammalian survival by suppressing oxidative stress and p53. Genes Dev. 2020, 34, 688–700. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H. Astaxanthin modulation of signaling pathways that regulate autophagy. Mar. Drugs 2019, 17, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surviladze, Z.; Sterk, R.T.; DeHaro, S.A.; Ozbun, M.A. Cellular entry of human papillomavirus type 16 involves activation of the phosphatidylinositol 3-kinase/Akt/mTOR pathway and inhibition of autophagy. J. Virol. 2013, 87, 2508–2517. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg-Lerner, A.; Kimchi, A. The paradox of autophagy and its implication in cancer etiology and therapy. Apoptosis 2009, 14, 376–391. [Google Scholar] [CrossRef] [Green Version]
- Ghavami, S.; Mutawe, M.M.; Sharma, P.; Yeganeh, B.; McNeill, K.D.; Klonisch, T.; Unruh, H.; Kashani, H.H.; Schaafsma, D.; Los, M.; et al. Mevalonate cascade regulation of airway mesenchymal cell autophagy and apoptosis: A dual role for p53. PLoS ONE 2011, 6, e16523. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Autophagy in the pathogenesis of disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghavami, S.; Mutawe, M.M.; Schaafsma, D.; Yeganeh, B.; Unruh, H.; Klonisch, T.; Halayko, A.J. Geranylgeranyl transferase 1 modulates autophagy and apoptosis in human airway smooth muscle. Am. J. Physiol. Lung C 2012, 302, L420–L428. [Google Scholar] [CrossRef] [Green Version]
- Rubinsztein, D.C.; Codogno, P.; Levine, B. Autophagy modulation as a potential therapeutic target for diverse diseases. Nat. Rev. Drug Discov. 2012, 11, 709–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brun, P.; Tarricone, E.; Di Stefano, A.; Mattiuzzo, E.; Mehrbod, P.; Ghavami, S.; Leonardi, A. The regulatory activity of autophagy in conjunctival fibroblasts and its possible role in vernal keratoconjunctivitis. J. Allergy Clin. Immunol. 2020, 146, 1210–1212. [Google Scholar] [CrossRef] [PubMed]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katheder, N.S.; Khezri, R.; O'Farrell, F.; Schultz, S.W.; Jain, A.; Rahman, M.M.; Schink, K.O.; Theodossiou, T.A.; Johansen, T.; Juhasz, G.; et al. Microenvironmental autophagy promotes tumour growth. Nature 2017, 541, 417–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giampieri, F.; Afrin, S.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Cianciosi, D.; Reboredo-Rodriguez, P.; Varela-Lopez, A.; Quiles, J.L.; Battino, M. Autophagy in human health and disease: Novel therapeutic opportunities. Antioxid. Redox Signal. 2019, 30, 577–634. [Google Scholar] [CrossRef] [PubMed]
- Santana-Codina, N.; Mancias, J.D.; Kimmelman, A.C. The role of autophagy in cancer. Annu. Rev. Cancer Biol. 2017, 1, 19–39. [Google Scholar] [CrossRef]
- Kondapuram, S.K.; Sarvagalla, S.; Coumar, M.S. Targeting autophagy with small molecules for cancer therapy. J. Cancer Metastasis Treat. 2019, 5, 32. [Google Scholar] [CrossRef]
- White, E. Deconvoluting the context-dependent role for autophagy in cancer. Nat. Rev. Cancer 2012, 12, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Kung, C.P.; Budina, A.; Balaburski, G.; Bergenstock, M.K.; Murphy, M. Autophagy in tumor suppression and cancer therapy. Crit. Rev. Eukar. Gene 2011, 21, 71–100. [Google Scholar] [CrossRef] [Green Version]
- Vega-Rubín-de-Celis, S. The role of Beclin 1-dependent autophagy in cancer. Biology 2019, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Bauvy, C.; Tonelli, G.; Yue, W.; Delomenie, C.; Nicolas, V.; Zhu, Y.; Domergue, V.; Marin-Esteban, V.; Tharinger, H.; et al. Beclin 1 and autophagy are required for the tumorigenicity of breast cancer stem-like/progenitor cells. Oncogene 2013, 32, 2261–2272, 2272e 1–11. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Yu, J.; Bhagat, G.; Furuya, N.; Hibshoosh, H.; Troxel, A.; Rosen, J.; Eskelinen, E.L.; Mizushima, N.; Ohsumi, Y.; et al. Promotion of tumorigenesis by heterozygous disruption of the beclin 1 autophagy gene. J. Clin. Investig. 2003, 112, 1809–1820. [Google Scholar] [CrossRef]
- Shen, Y.; Li, D.D.; Wang, L.L.; Deng, R.; Zhu, X.F. Decreased expression of autophagy-related proteins in malignant epithelial ovarian cancer. Autophagy 2008, 4, 1067–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, Y.; Coppola, D.; Matsushita, N.; Cualing, H.D.; Sun, M.; Sato, Y.; Liang, C.; Jung, J.U.; Cheng, J.Q.; Mule, J.J.; et al. Bif-1 interacts with Beclin 1 through UVRAG and regulates autophagy and tumorigenesis. Nat. Cell Biol. 2007, 9, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Lorzadeh, S.; Kohan, L.; Ghavami, S.; Azarpira, N. Autophagy and the Wnt signaling pathway: A focus on Wnt/beta-catenin signaling. BBA Mol. Cell Res. 2021, 1868, 118926. [Google Scholar] [CrossRef]
- He, S.; Zhao, Z.; Yang, Y.; O’Connell, D.; Zhang, X.; Oh, S.; Ma, B.; Lee, J.H.; Zhang, T.; Varghese, B.; et al. Truncating mutation in the autophagy gene UVRAG confers oncogenic properties and chemosensitivity in colorectal cancers. Nat. Commun. 2015, 6, 7839. [Google Scholar] [CrossRef] [Green Version]
- Takamura, A.; Komatsu, M.; Hara, T.; Sakamoto, A.; Kishi, C.; Waguri, S.; Eishi, Y.; Hino, O.; Tanaka, K.; Mizushima, N. Autophagy-deficient mice develop multiple liver tumors. Gene Dev. 2011, 25, 795–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmelman, A.C. The dynamic nature of autophagy in cancer. Gene Dev. 2011, 25, 1999–2010. [Google Scholar] [CrossRef] [Green Version]
- Errafiy, R.; Aguado, C.; Ghislat, G.; Esteve, J.M.; Gil, A.; Loutfi, M.; Knecht, E. PTEN increases autophagy and inhibits the ubiquitin-proteasome pathway in glioma cells independently of its lipid phosphatase activity. PLoS ONE 2013, 8, e83318. [Google Scholar] [CrossRef]
- Noguchi, M.; Hirata, N.; Suizu, F. The links between AKT and two intracellular proteolytic cascades: Ubiquitination and autophagy. Biochim. Biophys. Acta. 2014, 1846, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Maiuri, M.C.; Le Toumelin, G.; Criollo, A.; Rain, J.C.; Gautier, F.; Juin, P.; Tasdemir, E.; Pierron, G.; Troulinaki, K.; Tavernarakis, N.; et al. Functional and physical interaction between Bcl-X(L) and a BH3-like domain in Beclin-1. EMBO J. 2007, 26, 2527–2539. [Google Scholar] [CrossRef]
- Singh, S.S.; Vats, S.; Chia, A.Y.; Tan, T.Z.; Deng, S.; Ong, M.S.; Arfuso, F.; Yap, C.T.; Goh, B.C.; Sethi, G.; et al. Dual role of autophagy in hallmarks of cancer. Oncogenesis 2018, 37, 1142–1158. [Google Scholar] [CrossRef] [PubMed]
- Bellot, G.; Garcia-Medina, R.; Gounon, P.; Chiche, J.; Roux, D.; Pouyssegur, J.; Mazure, N.M. Hypoxia-induced autophagy is mediated through hypoxia-inducible factor induction of BNIP3 and BNIP3L via their BH3 domains. Mol. Cell. Biol. 2009, 29, 2570–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.; Mukhopadhyay, S.; Tung, K.; Patel, D.; Foster, D.A. Rapamycin-induced G1 cell cycle arrest employs both TGF-beta and Rb pathways. Cancer Lett. 2015, 360, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, T.; Tsuno, N.H.; Okaji, Y.; Sunami, E.; Shuno, Y.; Sasaki, K.; Hongo, K.; Kaneko, M.; Hiyoshi, M.; Kawai, K.; et al. The inhibition of autophagy potentiates anti-angiogenic effects of sulforaphane by inducing apoptosis. Angiogenesis 2010, 13, 227–238. [Google Scholar] [CrossRef]
- Kenific, C.M.; Thorburn, A.; Debnath, J. Autophagy and metastasis: Another double-edged sword. Curr. Opin. Cell Biol. 2010, 22, 241–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nencioni, A.; Cea, M.; Montecucco, F.; Longo, V.D.; Patrone, F.; Carella, A.M.; Holyoake, T.L.; Helgason, G.V. Autophagy in blood cancers: Biological role and therapeutic implications. Haematologica 2013, 98, 1335–1343. [Google Scholar] [CrossRef] [PubMed]
- Ferro, F.; Servais, S.; Besson, P.; Roger, S.; Dumas, J.-F.; Brisson, L. Autophagy and mitophagy in cancer metabolic remodelling. Semin. Cell Dev. Biol. 2020, 98, 129–138. [Google Scholar] [CrossRef]
- Xu, J.L.; Yuan, L.; Tang, Y.C.; Xu, Z.Y.; Xu, H.D.; Cheng, X.D.; Qin, J.J. The role of autophagy in gastric cancer chemoresistance: Friend or foe? Front. Cell Dev. Biol. 2020, 8, 621428. [Google Scholar] [CrossRef] [PubMed]
- Belounis, A.; Nyalendo, C.; Le Gall, R.; Imbriglio, T.V.; Mahma, M.; Teira, P.; Beaunoyer, M.; Cournoyer, S.; Haddad, E.; Vassal, G.; et al. Autophagy is associated with chemoresistance in neuroblastoma. BMC Cancer 2016, 16, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [PubMed]
- Read, A.; Schröder, M. The unfolded protein response: An overview. Biology 2021, 10, 384. [Google Scholar] [CrossRef] [PubMed]
- Mariño, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrouj, H.; Seghatoleslam, A.; Mokarram, P.; Ghavami, S. Effect of casein kinase 1alpha inhibition on autophagy flux and the AKT/phospho-beta-catenin (S552) axis in HCT116, a RAS-mutated colorectal cancer cell line. Can. J. Physiol. Pharm. 2021, 99, 284–293. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef]
- Elmore, S.P.; Qian, T.; Grissom, S.F.; Lemasters, J.J. The mitochondrial permeability transition initiates autophagy in rat hepatocytes. FASEB J. 2001, 15, 2286–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.M.; Ni, J.D.; Song, D.; Ding, M.; Huang, J. Interplay between unfolded protein response and autophagy promotes tumor drug resistance. Oncol. Lett. 2015, 10, 1959–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastghaib, S.; Kumar, P.S.; Aftabi, S.; Damera, G.; Dalvand, A.; Sepanjnia, A.; Kiumarsi, M.; Aghanoori, M.R.; Sohal, S.S.; Ande, S.R.; et al. Mechanisms targeting the unfolded protein response in asthma. Am. J. Resp. Cell Mol. 2021, 64, 29–38. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Y. JAK2/STAT3 inhibitor reduced 5-FU resistance and autophagy through ATF6-mediated ER stress. J. Recept. Signal. Transduct. Res. 2022, 42, 206–213, (Preprint). [Google Scholar] [CrossRef] [PubMed]
- Dastghaib, S.; Shojaei, S.; Mostafavi-Pour, Z.; Sharma, P.; Patterson, J.B.; Samali, A.; Mokarram, P.; Ghavami, S. Simvastatin Induces Unfolded Protein Response and Enhances Temozolomide-Induced Cell Death in Glioblastoma Cells. Cells 2020, 9, 2339. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Aghaei, M.; Dastghaib, S.; Aftabi, S.; Aghanoori, M.R.; Alizadeh, J.; Mokarram, P.; Mehrbod, P.; Ashrafizadeh, M.; Zarrabi, A.; McAlinden, K.D.; et al. The ER Stress/UPR Axis in Chronic Obstructive Pulmonary Disease and Idiopathic Pulmonary Fibrosis. Life 2020, 11, 1. [Google Scholar] [CrossRef]
- Kouroku, Y.; Fujita, E.; Tanida, I.; Ueno, T.; Isoai, A.; Kumagai, H.; Ogawa, S.; Kaufman, R.J.; Kominami, E.; Momoi, T. ER stress (PERK/eIF2alpha phosphorylation) mediates the polyglutamine-induced LC3 conversion, an essential step for autophagy formation. Cell Death Differ. 2007, 14, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, M.; Hino, S.; Saito, A.; Morikawa, K.; Kondo, S.; Kanemoto, S.; Murakami, T.; Taniguchi, M.; Tanii, I.; Yoshinaga, K.; et al. Autophagy is activated for cell survival after endoplasmic reticulum stress. Mol. Cell. Biol. 2006, 26, 9220–9231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, H.P.; Zhang, Y.; Bertolotti, A.; Zeng, H.; Ron, D. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Wang, J.; Kang, R.; Huang, H.; Xi, X.; Wang, B.; Wang, J.; Zhao, Z. Hepatitis C virus core protein activates autophagy through EIF2AK3 and ATF6 UPR pathway-mediated MAP1LC3B and ATG12 expression. Autophagy 2014, 10, 766–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- B’Chir, W.; Maurin, A.C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013, 41, 7683–7699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalckvar, E.; Berissi, H.; Mizrachy, L.; Idelchuk, Y.; Koren, I.; Eisenstein, M.; Sabanay, H.; Pinkas-Kramarski, R.; Kimchi, A. DAP-kinase-mediated phosphorylation on the BH3 domain of beclin 1 promotes dissociation of beclin 1 from Bcl-XL and induction of autophagy. EMBO Rep. 2009, 10, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.; Thompson, M.D.; Cohen, R.A.; Tong, X. Endoplasmic reticulum stress and related pathological processes. J. Pharmacol. Biomed. Anal. 2013, 1, 1000107. [Google Scholar]
- Avril, T.; Vauléon, E.; Chevet, E. Endoplasmic reticulum stress signaling and chemotherapy resistance in solid cancers. Oncogenesis 2017, 6, e373. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Liu, H.; Jiang, C.-C.; Fang, L.; Chen, C.; Zhang, X.-D.; Jiang, Z.-W. Connecting endoplasmic reticulum stress to autophagy through IRE1/JNK/beclin-1 in breast cancer cells. Int. J. Mol. Med. 2014, 34, 772–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinković, M.; Šprung, M.; Buljubašić, M.; Novak, I. Autophagy modulation in cancer: Current knowledge on action and therapy. Oxid. Med. Cell Longev. 2018, 2018, 8023821. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.-M.; Tan, Y.; Wang, H.; Peng, L.; Chen, H.-T.; Meng, X.-J.; Li, L.-L.; Liu, Y.; Li, W.-F.; Shan, H. The relationship between autophagy and the immune system and its applications for tumor immunotherapy. Mol. Cancer 2019, 18, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirone, M.; Gilardini Montani, M.S.; Granato, M.; Garufi, A.; Faggioni, A.; D’Orazi, G. Autophagy manipulation as a strategy for efficient anticancer therapies: Possible consequences. J. Exp. Clin. Cancer Res. 2019, 38, 262. [Google Scholar] [CrossRef]
- Arnold, J.; Murera, D.; Arbogast, F.; Fauny, J.-D.; Muller, S.; Gros, F. Autophagy is dispensable for B-cell development but essential for humoral autoimmune responses. Cell Death Differ. 2016, 23, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xia, P.; Huang, G.; Zhu, P.; Liu, J.; Ye, B.; Du, Y.; Fan, Z. FoxO1-mediated autophagy is required for NK cell development and innate immunity. Nat. Commun. 2016, 7, 11023. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Chen, X.; Ren, X.; Yang, J.M.; Cheng, Y. Emerging role of autophagy in anti-tumor immunity: Implications for the modulation of immunotherapy resistance. Drug Resist. Updates 2021, 56, 100752. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.S.C.; Gonçalves, L.B.; Lepique, A.P.; de Araujo-Souza, P.S. The role of autophagy in tumor immunology-Complex mechanisms that may be explored therapeutically. Front. Oncol. 2020, 10, 603661. [Google Scholar] [CrossRef]
- Martino, E.; Casamassima, G.; Castiglione, S.; Cellupica, E.; Pantalone, S.; Papagni, F.; Rui, M.; Siciliano, A.M.; Collina, S. Vinca alkaloids and analogues as anti-cancer agents: Looking back, peering ahead. Bioorg. Med. Chem. Lett. 2018, 28, 2816–2826. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Bao, J.; Zhao, W.; Hu, Y.; Lu, J.; Chen, X. Natural autophagy regulators in cancer therapy: A review. Phytochem. Rev. 2015, 14, 137–154. [Google Scholar] [CrossRef]
- Moudi, M.; Go, R.; Yien, C.Y.S.; Nazre, M. Vinca alkaloids. Int. J. Prev. Med. 2013, 4, 1231. [Google Scholar]
- Adiseshaiah, P.P.; Clogston, J.D.; McLeland, C.B.; Rodriguez, J.; Potter, T.M.; Neun, B.W.; Skoczen, S.L.; Shanmugavelandy, S.S.; Kester, M.; Stern, S.T.; et al. Synergistic combination therapy with nanoliposomal C6-ceramide and vinblastine is associated with autophagy dysfunction in hepatocarcinoma and colorectal cancer models. Cancer Lett. 2013, 337, 254–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.; Cheng, C.; Shao, Q.; Lin, Z.; Lu, S.; Chen, Y. CD24 promotes HCC progression via triggering notch-related EMT and modulation of tumor microenvironment. Tumor. Biol. 2016, 37, 6073–6084. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Cui, X.; Liang, H.; Hao, C.; Yang, Z.; Li, X.; Yang, X.; Han, C. CD24 is a potential biomarker for prognosis in human breast carcinoma. Cell Physiol. Biochem. 2018, 48, 111–119. [Google Scholar] [CrossRef]
- Deng, J.; Gao, G.; Wang, L.; Wang, T.; Yu, J.; Zhao, Z. CD24 expression as a marker for predicting clinical outcome in human gliomas. J. Biotechnol. Biomed. 2012, 2012, 517172. [Google Scholar] [CrossRef]
- Deng, X.; Apple, S.; Zhao, H.; Song, J.; Lee, M.; Luo, W.; Wu, X.; Chung, D.; Pietras, R.J.; Chang, H.R. CD24 Expression and differential resistance to chemotherapy in triple-negative breast cancer. Oncotarget 2017, 8, 38294. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Yao, Y.; Xu, G.; Zhou, C.; Zhang, Y.; Sun, J.; Jiang, R.; Shao, Q.; Chen, Y. CD24 regulates sorafenib resistance via activating autophagy in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 646. [Google Scholar] [CrossRef] [Green Version]
- Ono, Y.J.; Tanabe, A.; Tanaka, T.; Tanaka, Y.; Hayashi, M.; Terai, Y.; Ohmichi, M. Met signaling cascade is amplified by the recruitment of phosphorylated met to lipid rafts via CD24 and leads to drug resistance in endometrial cancer cell lines. Mol. Cancer Ther. 2015, 14, 2353–2363. [Google Scholar] [CrossRef] [Green Version]
- Eyvazi, S.; Kazemi, B.; Dastmalchi, S.; Bandehpour, M. Involvement of CD24 in multiple cancer related pathways makes it an interesting new target for cancer therapy. Curr. Cancer Drug Targets 2018, 18, 328–336. [Google Scholar] [CrossRef]
- Sun, J.; Feng, D.; Xi, H.; Luo, J.; Zhou, Z.; Liu, Q.; Chen, Y.; Shao, Q. CD24 blunts the sensitivity of retinoblastoma to vincristine by modulating autophagy. Mol. Oncol. 2020, 14, 1740–1759. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.E.; Wani, M.C.; Cook, C.; Palmer, K.H.; McPhail, A.a.; Sim, G. Plant antitumor agents. I. The isolation and structure of camptothecin, a novel alkaloidal leukemia and tumor inhibitor from camptotheca acuminata. J. Am. Chem. Soc. 1966, 88, 3888–3890. [Google Scholar] [CrossRef]
- Zhang, J.W.; Zhang, S.S.; Song, J.R.; Sun, K.; Zong, C.; Zhao, Q.D.; Liu, W.T.; Li, R.; Wu, M.C.; Wei, L.X. Autophagy inhibition switches low-dose camptothecin-induced premature senescence to apoptosis in human colorectal cancer cells. Biochem. Pharmacol. 2014, 90, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Jayasooriya, R.G.P.T.; Dilshara, M.G.; Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Choi, Y.H.; Kim, G.-Y. Camptothecin enhances c-Myc-mediated endoplasmic reticulum stress and leads to autophagy by activating Ca2+-mediated AMPK. Food Chem. Toxicol. 2018, 121, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-H.; Hsu, S.-H.; Hsu, H.-W.; Huang, K.-C.; Liu, W.; Wu, C.-Y.; Huang, W.-P.; Chen, J.Y.F.; Chen, B.-H.; Chiu, C.-C. Human non-small cell lung cancer cells can be sensitized to camptothecin by modulating autophagy. Int. J. Oncol. 2018, 53, 1967–1979. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, J.; Wang, Y.; Xing, R.; Yi, C.; Zhu, H.; Chen, X.; Guo, J.; Guo, W.; Li, W. Matrine, a novel autophagy inhibitor, blocks trafficking and the proteolytic activation of lysosomal proteases. Carcinogenesis 2013, 34, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Gu, D.; Li, J.; Cui, K.; Zhang, Y. Effects and action mechanisms of berberine and Rhizoma coptidis on gut microbes and obesity in high-fat diet-fed C57BL/6J mice. PLoS ONE 2011, 6, e24520. [Google Scholar] [CrossRef] [Green Version]
- Guamán Ortiz, L.M.; Croce, A.L.; Aredia, F.; Sapienza, S.; Fiorillo, G.; Syeda, T.M.; Buzzetti, F.; Lombardi, P.; Scovassi, A.I. Effect of new berberine derivatives on colon cancer cells. Acta Bioch. Bioph. Sin. 2015, 47, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.-H.; Tan, J.-Q.; Durairajan, S.S.K.; Liu, L.-F.; Zhang, Z.-H.; Ma, L.; Shen, H.-M.; Chan, H.E.; Li, M. Isorhynchophylline, a natural alkaloid, promotes the degradation of alpha-synuclein in neuronal cells via inducing autophagy. Autophagy 2012, 8, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Jang, B.-C.; Lim, K.-J.; Paik, J.-H.; Cho, J.-W.; Baek, W.-K.; Suh, M.-H.; Park, J.-B.; Kwon, T.K.; Park, J.-W.; Kim, S.-P. Tetrandrine-induced apoptosis is mediated by activation of caspases and PKC-δ in U937 cells. Biochem. Pharmacol. 2004, 67, 1819–1829. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, T.; Yu, M.; Li, K.; Li, W. The plant alkaloid tetrandrine inhibits metastasis via autophagy-dependent Wnt/β-catenin and metastatic tumor antigen 1 signaling in human liver cancer cells. J. Exp. Clin. Cancer Res. 2018, 37, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, B.Y.K.; Mok, S.W.F.; Chan, W.K.; Xu, S.W.; Wu, A.G.; Yao, X.J.; Wang, J.R.; Liu, L.; Wong, V.K.W. Hernandezine, a novel AMPK activator induces autophagic cell death in drug-resistant cancers. Oncotarget 2016, 7, 8090. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.P.; Seki, A.; Horiguchi, K.; Shoji, T.; Arai, T.; Nugroho, A.E.; Hirasawa, Y.; Sato, F.; Kaneda, T.; Morita, H. Bisleuconothine A induces autophagosome formation by interfering with AKT-mTOR signaling pathway. J. Nat. Prod. 2015, 78, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Utaipan, T.; Athipornchai, A.; Suksamrarn, A.; Jirachotikoon, C.; Yuan, X.; Lertcanawanichakul, M.; Chunglok, W. Carbazole alkaloids from Murraya koenigii trigger apoptosis and autophagic flux inhibition in human oral squamous cell carcinoma cells. J. Nat. Med. 2017, 71, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Garufi, A.; Pistritto, G.; Cirone, M.; D’Orazi, G. Reactivation of mutant p53 by capsaicin, the major constituent of peppers. J. Exp. Clin. Cancer Res. 2016, 35, 136. [Google Scholar] [CrossRef] [Green Version]
- Ellington, A.A.; Berhow, M.; Singletary, K.W. Induction of macroautophagy in human colon cancer cells by soybean B-group triterpenoid saponins. Carcinogenesis 2005, 26, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Q.; Yu, X.; Xiao, L.; Hu, Z.; Luo, X.; Tao, Y.; Yang, L.; Liu, X.; Chen, H.; Ding, Z. Neoalbaconol induces energy depletion and multiple cell death in cancer cells by targeting PDK1-PI3-K/Akt signaling pathway. Cell Death Dis. 2013, 4, e804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Lao, Y.; Xu, N.; Wang, X.; Tan, H.; Fu, W.; Lin, Z.; Xu, H. Guttiferone K induces autophagy and sensitizes cancer cells to nutrient stress-induced cell death. Phytomedicine 2015, 22, 902–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, E.; Nagao, Y.; Node, M.; Kaneko, K.; Nakazawa, S.; Kuroda, H. Antitumor activity of the Isodon diterpenoids: Structural requirements for the activity. Experientia 1976, 32, 203–206. [Google Scholar] [CrossRef]
- Yao, Z.; Xie, F.; Li, M.; Liang, Z.; Xu, W.; Yang, J.; Liu, C.; Li, H.; Zhou, H.; Qu, L.-H. Oridonin induces autophagy via inhibition of glucose metabolism in p53-mutated colorectal cancer cells. Cell Death Dis. 2017, 8, e2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croteau, R.; Ketchum, R.E.B.; Long, R.M.; Kaspera, R.; Wildung, M.R. Taxol biosynthesis and molecular genetics. Phytochem. Rev. 2006, 5, 75–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.-A.; Li, W.-H.; Chen, P.-H.; He, C.-L.; Chang, Y.-H.; Chuang, C.-M. Intraperitoneal delivery of a novel liposome-encapsulated paclitaxel redirects metabolic reprogramming and effectively inhibits cancer stem cells in Taxol®-resistant ovarian cancer. Am. J. Transl. Res. 2015, 7, 841. [Google Scholar] [PubMed]
- D’Antona, L.; Amato, R.; Talarico, C.; Ortuso, F.; Menniti, M.; Dattilo, V.; Iuliano, R.; Gigliotti, F.; Artese, A.; Costa, G.; et al. SI113, a specific inhibitor of the Sgk1 kinase activity that counteracts cancer cell proliferation. Cell Physiol. Biochem. 2015, 35, 2006–2018. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Gong, F.; Chen, Y.; Jiang, Y.; Liu, J.; Yu, M.; Zhang, S.; Wang, M.; Xiao, G.; Liao, H. Autophagy promotes paclitaxel resistance of cervical cancer cells: Involvement of Warburg effect activated hypoxia-induced factor 1-alpha-mediated signaling. Cell Death Dis. 2014, 5, e1367. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Shi, W. Autophagy regulates resistance of non-small cell lung cancer cells to paclitaxel. Tumor. Biol. 2016, 37, 10539–10544. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.H.; Du, X.; Lin, H.; Wang, P.C.; Li, M. Paclitaxel inhibits the progression of cervical cancer by inhibiting autophagy via lncRNARP11-381N20.2. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3010–3017. [Google Scholar] [PubMed]
- Yang, J.-C.; Lu, M.-C.; Lee, C.-L.; Chen, G.-Y.; Lin, Y.-Y.; Chang, F.-R.; Wu, Y.-C. Selective targeting of breast cancer cells through ROS-mediated mechanisms potentiates the lethality of paclitaxel by a novel diterpene, gelomulide K. Free Radical. Biol. Med. 2011, 51, 641–657. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.M.; Jung, J.H.; Jeong, S.J.; Sohn, E.J.; Kim, B.; Kim, S.H. Tanshinone IIA induces autophagic cell death via activation of AMPK and ERK and inhibition of mTOR and p70 S6K in KBM-5 leukemia cells. Phytother. Res. 2014, 28, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.B.; Fu, P.Y.; Ky, N.; Zhu, H.S.; Feng, X.; Li, J.; Srinivasan, K.G.; Hamza, M.S.; Zhao, Y. NF-κB p65 repression by the sesquiterpene lactone, Helenalin, contributes to the induction of autophagy cell death. BMC Complement Altern Med. 2012, 12, 93. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, P.; Hong, H.; Wang, L.; Zhou, Y.; Lang, Y. JNK pathway mediates curcumin-induced apoptosis and autophagy in osteosarcoma MG63 cells. Exp. Ther. Med. 2017, 14, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, C.P.; Lima, C.F.; Pedro, D.F.; Wilson, J.M.; Kristiansen, K.; Pereira-Wilson, C. Ursolic acid induces cell death and modulates autophagy through JNK pathway in apoptosis-resistant colorectal cancer cells. J. Nutr. Biochem. 2013, 24, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Meng, Y.; Sun, Q.; Zhang, Z.; Guo, X.; Sheng, X.; Tai, G.; Cheng, H.; Zhou, Y. Ginsenoside compound K sensitizes human colon cancer cells to TRAIL-induced apoptosis via autophagy-dependent and-independent DR5 upregulation. Cell Death Dis. 2016, 7, e2334. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.O.; Shastina, V.V.; Shin, S.W.; Xu, Q.; Park, J.I.; Rasskazov, V.A.; Avilov, S.A.; Fedorov, S.N.; Stonik, V.A.; Kwak, J.Y. Differential effects of triterpene glycosides, frondoside A and cucumarioside A2-2 isolated from sea cucumbers on caspase activation and apoptosis of human leukemia cells. FEBS Lett. 2009, 583, 697–702. [Google Scholar] [CrossRef] [Green Version]
- Dyshlovoy, S.A.; Madanchi, R.; Hauschild, J.; Otte, K.; Alsdorf, W.H.; Schumacher, U.; Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; Honecker, F.; et al. The marine triterpene glycoside frondoside A induces p53-independent apoptosis and inhibits autophagy in urothelial carcinoma cells. BMC Cancer 2017, 17, 93. [Google Scholar] [CrossRef]
- Lao, Y.; Wan, G.; Liu, Z.; Wang, X.; Ruan, P.; Xu, W.; Xu, D.; Xie, W.; Zhang, Y.; Xu, H.; et al. The natural compound oblongifolin C inhibits autophagic flux and enhances antitumor efficacy of nutrient deprivation. Autophagy 2014, 10, 736–749. [Google Scholar] [CrossRef] [Green Version]
- Ranelletti, F.O.; Maggiano, N.; Serra, F.G.; Ricci, R.; Larocca, L.M.; Lanza, P.; Scambia, G.; Fattorossi, A.; Capelli, A.; Piantelli, M. Quercetin inhibits p21-RAS expression in human colon cancer cell lines and in primary colorectal tumors. Int. J. Cancer 2000, 85, 438–445. [Google Scholar] [CrossRef]
- Avila, M.A.; Velasco, J.A.; Cansado, J.; Notario, V. Quercetin mediates the down-regulation of mutant p53 in the human breast cancer cell line MDA-MB468. Cancer Res. 1994, 54, 2424–2428. [Google Scholar] [PubMed]
- Srinivasan, A.; Thangavel, C.; Liu, Y.; Shoyele, S.; Den, R.B.; Selvakumar, P.; Lakshmikuttyamma, A. Quercetin regulates β-catenin signaling and reduces the migration of triple negative breast cancer. Mol. Carcinog. 2016, 55, 743–756. [Google Scholar] [CrossRef]
- Psahoulia, F.H.; Moumtzi, S.; Roberts, M.L.; Sasazuki, T.; Shirasawa, S.; Pintzas, A. Quercetin mediates preferential degradation of oncogenic Ras and causes autophagy in Ha-RAS-transformed human colon cells. Carcinogenesis 2007, 28, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR-and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klappan, A.K.; Hones, S.; Mylonas, I.; Brüning, A. Proteasome inhibition by quercetin triggers macroautophagy and blocks mTOR activity. Histochem. Cell Biol. 2012, 137, 25–36. [Google Scholar] [CrossRef] [PubMed]
- De, A.; De, A.; Papasian, C.; Hentges, S.; Banerjee, S.; Haque, I.; Banerjee, S.K. Emblica officinalis extract induces autophagy and inhibits human ovarian cancer cell proliferation, angiogenesis, growth of mouse xenograft tumors. PLoS ONE 2013, 8, e72748. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.-K.; Lin-Shiau, S.-Y.; Lin, J.-K. Induction of apoptosis by apigenin and related flavonoids through cytochrome c release and activation of caspase-9 and caspase-3 in leukaemia HL-60 cells. Eur. J. Cancer 1999, 35, 1517–1525. [Google Scholar] [CrossRef]
- Wang, W.; Heideman, L.; Chung, C.S.; Pelling, J.C.; Koehler, K.J.; Birt, D.F. Cell-cycle arrest at G2/M and growth inhibition by apigenin in human colon carcinoma cell lines. Mol. Carcinog. 2000, 28, 102–110. [Google Scholar] [CrossRef]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer 2000, 87, 595–600. [Google Scholar] [CrossRef]
- Ruela-de-Sousa, R.; Fuhler, G.M.; Blom, N.; Ferreira, C.V.; Aoyama, H.; Peppelenbosch, M.P. Cytotoxicity of apigenin on leukemia cell lines: Implications for prevention and therapy. Cell Death Dis. 2010, 1, e19. [Google Scholar] [CrossRef] [Green Version]
- Jyoti, A.S.; Saxena, S.; Sharma, A. Phytoestrogen “genistein”: Its extraction and isolation from soybean seeds. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1121–1126. [Google Scholar]
- Gossner, G.; Choi, M.; Tan, L.; Fogoros, S.; Griffith, K.A.; Kuenker, M.; Liu, J.R. Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecol. Oncol. 2007, 105, 23–30. [Google Scholar] [CrossRef]
- Christian, F.; Anthony, D.F.; Vadrevu, S.; Riddell, T.; Day, J.P.; McLeod, R.; Adams, D.R.; Baillie, G.S.; Houslay, M.D. p62 (SQSTM1) and cyclic AMP phosphodiesterase-4A4 (PDE4A4) locate to a novel, reversible protein aggregate with links to autophagy and proteasome degradation pathways. Cell Signal. 2010, 22, 1576–1596. [Google Scholar] [CrossRef]
- Rafatian, G.; Khodagholi, F.; Farimani, M.M.; Abraki, S.B.; Gardaneh, M. Increase of autophagy and attenuation of apoptosis by Salvigenin promote survival of SH-SY5Y cells following treatment with H2O2. Mol. Cell Biochem. 2012, 371, 9–22. [Google Scholar] [CrossRef]
- Xuan, H.-Z.; Zhang, J.-H.; Wang, Y.-H.; Fu, C.-L.; Zhang, W. Anti-tumor activity evaluation of novel chrysin–organotin compound in MCF-7 cells. Bioorg. Med. Chem. Lett. 2016, 26, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Chiu, Y.-W.; Lin, T.-H.; Huang, W.-S.; Teng, C.-Y.; Liou, Y.-S.; Kuo, W.-H.; Lin, W.-L.; Huang, H.-I.; Tung, J.-N.; Huang, C.-Y. Baicalein inhibits the migration and invasive properties of human hepatoma cells. Toxicol. Appl. Pharmacol. 2011, 255, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Wang, N.; Man, K.; Tsao, S.; Che, C.; Feng, Y. Autophagy-induced RelB/p52 activation mediates tumour-associated macrophage repolarisation and suppression of hepatocellular carcinoma by natural compound baicalin. Cell Death Dis. 2015, 6, e1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.-L.; Dong, J.-L.; Wu, J. Juglanin induces apoptosis and autophagy in human breast cancer progression via ROS/JNK promotion. Biomed. Pharmacother. 2017, 85, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.; Crozier, A. Plant foods and herbal sources of resveratrol. J. Agr. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef]
- Opipari, A.W.; Tan, L.; Boitano, A.E.; Sorenson, D.R.; Aurora, A.; Liu, J.R. Resveratrol-induced autophagocytosis in ovarian cancer cells. Cancer Res. 2004, 64, 696–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Chang, H.; Peng, X.; Bai, Q.; Yi, L.; Zhou, Y.; Zhu, J.; Mi, M. Resveratrol inhibits breast cancer stem-like cells and induces autophagy via suppressing Wnt/β-catenin signaling pathway. PLoS ONE 2014, 9, e102535. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Zepeda, S.P.; Garcia-Villa, E.; Diaz-Chavez, J.; Hernandez-Pando, R.; Gariglio, P. Resveratrol induces cell death in cervical cancer cells through apoptosis and autophagy. Eur. J. Cancer Prev. 2013, 22, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, T.; Zhu, C.; Sun, H.; Qiu, Y.; Zhu, X.; Li, J. Resveratrol triggers protective autophagy through the ceramide/Akt/mTOR pathway in melanoma B16 cells. Nutr. Cancer 2014, 66, 435–440. [Google Scholar] [CrossRef]
- Selvaraj, S.; Sun, Y.; Sukumaran, P.; Singh, B.B. Resveratrol activates autophagic cell death in prostate cancer cells via downregulation of STIM1 and the mTOR pathway. Mol. Carcinog. 2016, 55, 818–831. [Google Scholar] [CrossRef]
- Kueck, A.; Opipari, A.W., Jr.; Griffith, K.A.; Tan, L.; Choi, M.; Huang, J.; Wahl, H.; Liu, J.R. Resveratrol inhibits glucose metabolism in human ovarian cancer cells. Gynecol. Oncol. 2007, 107, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Suzuki, S.O.; Himeno, M. Resveratrol-induced autophagy in human U373 glioma cells. Oncol. Lett. 2010, 1, 489–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puissant, A.; Robert, G.; Fenouille, N.; Luciano, F.; Cassuto, J.-P.; Raynaud, S.; Auberger, P. Resveratrol promotes autophagic cell death in chronic myelogenous leukemia cells via JNK-mediated p62/SQSTM1 expression and AMPK activation. Cancer Res. 2010, 70, 1042–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarlatti, F.; Maffei, R.; Beau, I.; Codogno, P.; Ghidoni, R. Role of non-canonical Beclin 1-independent autophagy in cell death induced by resveratrol in human breast cancer cells. Cell Death Differ. 2008, 15, 1318–1329. [Google Scholar] [CrossRef]
- Back, J.H.; Zhu, Y.; Calabro, A.; Queenan, C.; Kim, A.S.; Arbesman, J.; Kim, A.L. Resveratrol-mediated downregulation of Rictor attenuates autophagic process and suppresses UV-induced skin carcinogenesis. Photochem. Photobiol. 2012, 88, 1165–1172. [Google Scholar] [CrossRef]
- Proikas-Cezanne, T.; Codogno, P. Beclin 1 or not Beclin 1. Autophagy 2011, 7, 671–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauthe, M.; Jacob, A.; Freiberger, S.; Hentschel, K.; Stierhof, Y.-D.; Codogno, P.; Proikas-Cezanne, T. Resveratrol-mediated autophagy requires WIPI-1-regulated LC3 lipidation in the absence of induced phagophore formation. Autophagy 2011, 7, 1448–1461. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.-C.; Ng, L.-T.; Lin, L.-T.; Richardson, C.D.; Wang, G.-H.; Lin, C.-C. Resveratrol arrests cell cycle and induces apoptosis in human hepatocellular carcinoma Huh-7 cells. J. Med. Food 2010, 13, 1415–1423. [Google Scholar] [CrossRef]
- Maher, E.A.; Furnari, F.B.; Bachoo, R.M.; Rowitch, D.H.; Louis, D.N.; Cavenee, W.K.; DePinho, R.A. Malignant glioma: Genetics and biology of a grave matter. Gene Dev. 2001, 15, 1311–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippi-Chiela, E.C.; Villodre, E.S.; Zamin, L.L.; Lenz, G. Autophagy interplay with apoptosis and cell cycle regulation in the growth inhibiting effect of resveratrol in glioma cells. PLoS ONE 2011, 6, e20849. [Google Scholar] [CrossRef]
- Lee, I.H.; Cao, L.; Mostoslavsky, R.; Lombard, D.B.; Liu, J.; Bruns, N.E.; Tsokos, M.; Alt, F.W.; Finkel, T. A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proc. Natl. Acad. Sci. USA 2008, 105, 3374–3379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.-w.; Chung, S.; Sundar, I.K.; Yao, H.; Arunachalam, G.; McBurney, M.W.; Rahman, I. Cigarette smoke-induced autophagy is regulated by SIRT1–PARP-1-dependent mechanism: Implication in pathogenesis of COPD. Arch. Biochem. Biophys. 2010, 500, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Hsu, K.-F.; Wu, C.-L.; Huang, S.-C.; Wu, C.-M.; Hsiao, J.-R.; Yo, Y.-T.; Chen, Y.-H.; Shiau, A.-L.; Chou, C.-Y. Cathepsin L mediates resveratrol-induced autophagy and apoptotic cell death in cervical cancer cells. Autophagy 2009, 5, 451–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuttan, R.; Sudheeran, P.C.; Josph, C.D. Turmeric and curcumin as topical agents in cancer therapy. Tumori 1987, 73, 29–31. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, J.; Cui, R.; Lin, J.; Ding, X. Curcumin in treating breast cancer: A review. J. Lab. Autom. 2016, 21, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinojima, N.; Yokoyama, T.; Kondo, Y.; Kondo, S. Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in curcumin-induced autophagy. Autophagy 2007, 3, 635–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, H.; Takada, Y.; Kondo, S.; Sawaya, R.; Aggarwal, B.B.; Kondo, Y. Evidence that curcumin suppresses the growth of malignant gliomas in vitro and in vivo through induction of autophagy: Role of Akt and extracellular signal-regulated kinase signaling pathways. Mol. Pharmacol. 2007, 72, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Kim, N.-Y.; Suh, Y.-A.; Lee, C. Involvement of ROS in curcumin-induced autophagic cell death. Korean J. Physiol. Pharmacol. 2011, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Cho, T.J.; Woo, B.H.; Choi, K.U.; Lee, C.H.; Ryu, M.H.; Park, H.R. Curcumin-induced autophagy contributes to the decreased survival of oral cancer cells. Arch. Oral. Biol. 2012, 57, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Jiang, J.; Guan, C.; Dong, C.; Wang, G.; Bai, L.; Sun, J.; Hu, C.; Bai, C. Curcumin induces autophagy via activating the AMPK signaling pathway in lung adenocarcinoma cells. J. Pharmacol. Sci. 2013, 123, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Takeda, T.; Tsuiji, K.; Wong, T.F.; Tadakawa, M.; Kondo, A.; Nagase, S.; Yaegashi, N. Curcumin induces cross-regulation between autophagy and apoptosis in uterine leiomyosarcoma cells. Int. J. Gynecol. Cancer 2013, 23, 803–808. [Google Scholar] [CrossRef]
- Settembre, C.; Fraldi, A.; Medina, D.L.; Ballabio, A. Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nat. Rev. Mol. Cell Biol. 2013, 14, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Settembre, C.; Di Malta, C.; Polito, V.A.; Arencibia, M.G.; Vetrini, F.; Erdin, S.; Erdin, S.U.; Huynh, T.; Medina, D.; Colella, P. TFEB links autophagy to lysosomal biogenesis. Science 2011, 332, 1429–1433. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.; Xu, J.; Lu, Y.; Jiang, J.; Wang, L.; Shen, H.-M.; Xia, D. Curcumin targets the TFEB-lysosome pathway for induction of autophagy. Oncotarget 2016, 7, 75659. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Long, M.; Li, X.; Zhu, S.; Zhang, M.; Yang, Z. Curcumin activates autophagy and attenuates oxidative damage in EA. hy926 cells via the Akt/mTOR pathway. Mol. Med. Rep. 2016, 13, 2187–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin induces autophagy, inhibits proliferation and invasion by downregulating AKT/mTOR signaling pathway in human melanoma cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Bu, S. Curcumin induces autophagy, apoptosis, and cell cycle arrest in human pancreatic cancer cells. Evid. -Based Complement. Altern. Med. 2017, 2017, 5787218. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Wang, C.; Yang, D.; Wei, Z.; Xu, J.; Hu, Z.; Zhang, Y.; Wang, W.; Yan, R.; Cai, Q. Curcumin regulates proliferation, autophagy, and apoptosis in gastric cancer cells by affecting PI3K and P53 signaling. J. Cell Physiol. 2018, 233, 4634–4642. [Google Scholar] [CrossRef] [PubMed]
- Basile, V.; Belluti, S.; Ferrari, E.; Gozzoli, C.; Ganassi, S.; Quaglino, D.; Saladini, M.; Imbriano, C. bis-Dehydroxy-Curcumin triggers mitochondrial-associated cell death in human colon cancer cells through ER-stress induced autophagy. PLoS ONE 2013, 8, e53664. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Lai, C.S.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Tetrahydrocurcumin, a major metabolite of curcumin, induced autophagic cell death through coordinative modulation of PI3K/Akt-mTOR and MAPK signaling pathways in human leukemia HL-60 cells. Mol. Nutr. Food Res. 2011, 55, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, G.; Ramalingam, S.; Subramaniam, D.; Rangarajan, P.; Protti, P.; Rammamoorthy, P.; Anant, S.; Mammen, J.M. Honokiol induces cytotoxic and cytostatic effects in malignant melanoma cancer cells. Am. J. Surg. 2012, 204, 868–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasul, A.; Yu, B.; Khan, M.; Zhang, K.; Iqbal, F.; Ma, T.; Yang, H. Magnolol, a natural compound, induces apoptosis of SGC-7901 human gastric adenocarcinoma cells via the mitochondrial and PI3K/Akt signaling pathways. Int. J. Oncol. 2012, 40, 1153–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishal, R.P.; Madhavi, G.P.; Rakesh, K.P. Development and validation of a RP-HPLC method for quantification of rottlerin in Kamala (Mallotus philppinensis). Drug Inven. Today 2009, 1, 116. [Google Scholar]
- Song, K.-S.; Kim, J.-S.; Yun, E.-J.; Kim, Y.-R.; Seo, K.-S.; Park, J.-H.; Jung, Y.-J.; Park, J.-I.; Kweon, G.-R.; Yoon, W.-H. Rottlerin induces autophagy and apoptotic cell death through a PKC-delta-independent pathway in HT1080 human fibrosarcoma cells: The protective role of autophagy in apoptosis. Autophagy 2008, 4, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Akar, U.; Ozpolat, B.; Mehta, K.; Fok, J.; Kondo, Y.; Lopez-Berestein, G. Tissue transglutaminase inhibits autophagy in pancreatic cancer cells. Mol. Cancer Res. 2007, 5, 241–249. [Google Scholar] [CrossRef]
- Balgi, A.D.; Fonseca, B.D.; Donohue, E.; Tsang, T.C.; Lajoie, P.; Proud, C.G.; Nabi, I.R.; Roberge, M. Screen for chemical modulators of autophagy reveals novel therapeutic inhibitors of mTORC1 signaling. PLoS ONE 2009, 4, e7124. [Google Scholar] [CrossRef] [Green Version]
- Torricelli, C.; Salvadori, S.; Valacchi, G.; Souček, K.; Slabáková, E.; Muscettola, M.; Volpi, N.; Maioli, E. Alternative pathways of cancer cell death by rottlerin: Apoptosis versus autophagy. Evid. -Based Complement. Altern. Med. 2012, 2012, 980658. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin-induced autophagy leads to the apoptosis in breast cancer stem cells: Molecular mechanisms. Mol. Cancer 2013, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Torricelli, C.; Daveri, E.; Salvadori, S.; Valacchi, G.; Ietta, F.; Muscettola, M.; Carlucci, F.; Maioli, E. Phosphorylation-independent mTORC1 inhibition by the autophagy inducer Rottlerin. Cancer Lett. 2015, 360, 17–27. [Google Scholar] [CrossRef]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy and apoptosis in prostate cancer stem cells via PI3K/Akt/mTOR signaling pathway. Cancer Lett. 2014, 343, 179–189. [Google Scholar] [CrossRef]
- Wang, X.; Howell, C.P.; Chen, F.; Yin, J.; Jiang, Y. Gossypol—A polyphenolic compound from cotton plant. Adv. Food Nutr. Res. 2009, 58, 215–263. [Google Scholar]
- Yuan, Y.; Tang, A.J.; Castoreno, A.B.; Kuo, S.Y.; Wang, Q.; Kuballa, P.; Xavier, R.; Shamji, A.F.; Schreiber, S.L.; Wagner, B.K. Gossypol and an HMT G9a inhibitor act in synergy to induce cell death in pancreatic cancer cells. Cell Death Dis. 2013, 4, e690. [Google Scholar] [CrossRef] [Green Version]
- Jing, Z.; Fei, W.; Zhou, J.; Zhang, L.; Chen, L.; Zhang, X.; Liang, X.; Xie, J.; Fang, Y.; Sui, X.; et al. Salvianolic acid B, a novel autophagy inducer, exerts antitumor activity as a single agent in colorectal cancer cells. Oncotarget 2016, 7, 61509–61519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, N.; Liu, P.; Chen, Q.; Situ, H.; Xie, T.; Zhang, J.; Peng, C.; Lin, Y.; Chen, J. MicroRNA-25 regulates chemoresistance-associated autophagy in breast cancer cells, a process modulated by the natural autophagy inducer isoliquiritigenin. Oncotarget 2014, 5, 7013. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Fang, Y.; Yang, Y.; Qin, Y.; Wu, P.; Wang, T.; Lai, H.; Meng, L.; Wang, D.; Zheng, Z.; et al. Elaiophylin, a novel autophagy inhibitor, exerts antitumor activity as a single agent in ovarian cancer cells. Autophagy 2015, 11, 1849–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göder, A.; Nagel, G.; Kraus, A.; Dörsam, B.; Seiwert, N.; Kaina, B.; Fahrer, J. Lipoic acid inhibits the DNA repair protein O 6-methylguanine-DNA methyltransferase (MGMT) and triggers its depletion in colorectal cancer cells with concomitant autophagy induction. Carcinogenesis 2015, 36, 817–831. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Morselli, E.; Mariño, G.; Bennetzen, M.V.; Eisenberg, T.; Megalou, E.; Schroeder, S.; Cabrera, S.; Bénit, P.; Rustin, P.; Criollo, A. Spermidine and resveratrol induce autophagy by distinct pathways converging on the acetylproteome. J. Cell Biol. 2011, 192, 615–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S. Allicin and other functional active components in garlic: Health benefits and bioavailability. Int. J. Food Prop. 2007, 10, 245–268. [Google Scholar] [CrossRef]
- Hirsch, K.; Danilenko, M.; Giat, J.; Miron, T.; Rabinkov, A.; Wilchek, M.; Mirelman, D.; Levy, J.; Sharoni, Y. Effect of purified Allicin, the major ingredient of freshly crushed garlic, on cancer cell proliferation. Nutr. Cancer 2000, 38, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Zhao, J.; Zhao, M.; Wang, K. Allicin activates autophagic cell death to alleviate the malignant development of thyroid cancer. Exp. Ther. Med. 2018, 15, 3537–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimasi, P.; Quintiero, A.; Shelkovnikova, T.A.; Buchman, V.L. Modulation of p-eIF2α cellular levels and stress granule assembly/disassembly by trehalose. Sci. Rep. 2017, 7, 44088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.-c.; Pan, Z.-h.; Liu, B.-n.; Meng, Z.-w.; Wu, X.; Zhou, Q.-h.; Xu, K. Benzyl isothiocyanate induces protective autophagy in human lung cancer cells through an endoplasmic reticulum stress-mediated mechanism. Acta Pharmacol. Sin. 2017, 38, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Ueda, J.Y.; Athikomkulchai, S.; Miyatake, R.; Saiki, I.; Esumi, H.; Awale, S. (+)-Grandifloracin, an antiausterity agent, induces autophagic PANC-1 pancreatic cancer cell death. Drug Des. Devel. Ther. 2014, 8, 39. [Google Scholar]
- Kolomeichuk, S.N.; Bene, A.; Upreti, M.; Dennis, R.A.; Lyle, C.S.; Rajasekaran, M.; Chambers, T.C. Induction of apoptosis by Vinblastine via c-Jun autoamplification and p53-independent down-regulation of p21WAF1/CIP1. Mol. Pharmacol. 2008, 73, 128–136. [Google Scholar]
- Brantley-Finley, C.; Lyle, C.S.; Du, L.; Goodwin, M.E.; Hall, T.; Szwedo, D.; Kaushal, G.P.; Chambers, T.C. The JNK, ERK and p53 pathways play distinct roles in apoptosis mediated by the antitumor agents vinblastine, doxorubicin, and etoposide. Biochem. Pharmacol. 2003, 66, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.J.P.; Salerni, B.L.; Lowrey, C.H.; Eastman, A. Vinblastine sensitizes leukemia cells to cyclin-dependent kinase inhibitors, inducing acute cell cycle phase-independent apoptosis. Cancer Biol. Ther. 2011, 12, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Groninger, E.; Boer, M.-D.; De Graaf, S.; Kamps, W.; De Bont, E. Vincristine induced apoptosis in acute lymphoblastic leukaemia cells: A mitochondrial controlled pathway regulated by reactive oxygen species? Int. J. Oncol. 2002, 21, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.-k.; Wang, P.; Zhang, X.D.; Jiang, C.C.; Chen, L.H.; Avery-Kiejda, K.A.; Watts, R.; Hersey, P. Activation of Jun N-terminal kinase is a mediator of vincristine-induced apoptosis of melanoma cells. Anti-Cancer Drug 2008, 19, 189–200. [Google Scholar] [CrossRef]
- Casado, P.; Zuazua-Villar, P.; Valle, E.d.; Martínez-Campa, C.; Lazo, P.S.; Ramos, S. Vincristine regulates the phosphorylation of the antiapoptotic protein HSP27 in breast cancer cells. Cancer Lett. 2007, 247, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Li, Y.; Tan, X.; Fu, L. Small heat shock proteins in cancers: Functions and therapeutic potential for cancer therapy. Int. J. Mol. Sci. 2020, 21, 6611. [Google Scholar] [CrossRef]
- Chen, M.B.; Shen, W.X.; Yang, Y.; Wu, X.Y.; Gu, J.H.; Lu, P.H. Activation of AMP-activated protein kinase is involved in vincristine-induced cell apoptosis in B16 melanoma cell. J. Cell Physiol. 2011, 226, 1915–1925. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Cheng, S.; Zhang, S.; Sun, H.; Xu, Z. Vincristine induces cell cycle arrest and apoptosis in SH-SY5Y human neuroblastoma cells. Int. J. Mol. Med. 2013, 31, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Song, Y.; Chen, H.; Pan, S.; Sun, X. Matrine inhibits proliferation and induces apoptosis of pancreatic cancer cells in vitro and in vivo. Biol. Pharm. Bull. 2010, 33, 1740–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-M.; Chen, Y.; Chen, J.-C.; Lin, T.-Y.; Tseng, S.-H. Tetrandrine induces apoptosis and growth suppression of colon cancer cells in mice. Cancer Lett. 2010, 287, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Liew, S.Y.; Looi, C.Y.; Paydar, M.; Cheah, F.K.; Leong, K.H.; Wong, W.F.; Mustafa, M.R.; Litaudon, M.; Awang, K. Subditine, a new monoterpenoid indole Alkaloid from bark of Nauclea subdita (Korth.) Steud. Induces apoptosis in human prostate cancer cells. PLoS ONE 2014, 9, e87286. [Google Scholar] [CrossRef]
- Xia, E.H.; Zhang, H.B.; Sheng, J.; Li, K.; Zhang, Q.J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W.; et al. The Tea tree genome provides insights into Tea flavor and independent evolution of caffeine biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiki, S.; Sasazawa, Y.; Imamichi, Y.; Kawajiri, S.; Fujimaki, T.; Tanida, I.; Kobayashi, H.; Sato, F.; Sato, S.; Ishikawa, K.-I.; et al. Caffeine induces apoptosis by enhancement of autophagy via PI3K/Akt/mTOR/p70S6K inhibition. Autophagy 2011, 7, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Sun, H.; Yu, D.; Liang, Y.; Yuan, Z.; Ge, Y. Hydroxycamptothecin induces apoptosis of human tenon's capsule fibroblasts by activating the PERK signaling pathway. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4749–4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-A.; Kim, J.-Y.; Lee, J.-Y.; Kang, C.-M.; Kwon, H.-J.; Yoo, Y.-D.; Kim, T.-W.; Lee, Y.-S.; Lee, S.-J. Induction of cell cycle arrest and apoptosis in human breast cancer cells by quercetin. Int. J. Oncol. 2001, 19, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.-T.; Huang, Y.-T.; Hwang, J.-J.; Lee, P.; Ke, F.-C.; Nair, M.P.; Kanadaswam, C.; Lee, M.-T. Blockade of the epidermal growth factor receptor tyrosine kinase activity by quercetin and luteolin leads to growth inhibition and apoptosis of pancreatic tumor cells. Anticancer Res. 2002, 22, 1615–1627. [Google Scholar] [PubMed]
- Shen, S.C.; Chen, Y.C.; Hsu, F.L.; Lee, W.R. Differential apoptosis-inducing effect of quercetin and its glycosides in human promyeloleukemic HL-60 cells by alternative activation of the caspase 3 cascade. J. Cell Biochem. 2003, 89, 1044–1055. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, Y.J. TRAIL apoptosis is enhanced by quercetin through Akt dephosphorylation. J. Cell Biochem. 2007, 100, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Manouchehri, J.M.; Kalafatis, M.; Lindner, D. Evaluation of the Efficacy of TRAIL Plus Quercetin as a Potential Breast Carcinoma Therapeutic; AACR: Philadelphia, PA, USA, 2016. [Google Scholar]
- Lee, D.-H.; Szczepanski, M.; Lee, Y.J. Role of Bax in quercetin-induced apoptosis in human prostate cancer cells. Biochem. Pharmacol. 2008, 75, 2345–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Zhang, K.; Zhang, Q.; Mei, J.; Chen, C.-j.; Feng, Z.-z.; Yu, D.-h. Effects of quercetin on the apoptosis of the human gastric carcinoma cells. Toxicol. Vitr. 2012, 26, 221–228. [Google Scholar] [CrossRef]
- Li, X.; Zhou, N.; Wang, J.; Liu, Z.; Wang, X.; Zhang, Q.; Liu, Q.; Gao, L.; Wang, R. Quercetin suppresses breast cancer stem cells (CD44+/CD24−) by inhibiting the PI3K/Akt/mTOR-signaling pathway. Life Sci. 2018, 196, 56–62. [Google Scholar] [CrossRef]
- Braicu, C.; Pileczki, V.; Pop, L.; Petric, R.C.; Chira, S.; Pointiere, E.; Achimas-Cadariu, P.; Berindan-Neagoe, I. Dual targeted therapy with p53 siRNA and Epigallocatechingallate in a triple negative breast cancer cell model. PLoS ONE 2015, 10, e0120936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Li, W.; Liu, F. Inhibition of proliferation and induction of apoptosis by genistein in colon cancer HT-29 cells. Cancer Lett. 2004, 215, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Olivera, A.; Moore, T.W.; Hu, F.; Brown, A.P.; Sun, A.; Liotta, D.C.; Snyder, J.P.; Yoon, Y.; Shim, H.; Marcus, A.I. Inhibition of the NF-κB signaling pathway by the curcumin analog, 3, 5-Bis (2-pyridinylmethylidene)-4-piperidone (EF31): Anti-inflammatory and anti-cancer properties. Int. Immunopharmacol. 2012, 12, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A.; Buhrmann, C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.L.; Wu, J.G.; Dai, N.; Yu, H.G.; Si, J.M. Curcumin reverses chemoresistance of human gastric cancer cells by downregulating the NF-κB transcription factor. Oncol. Rep. 2011, 26, 1197–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.; Fan, T.; Zhang, Y.; Jiang, Y.; Zhang, X. Curcumin potentiates the antitumor effects of 5-FU in treatment of esophageal squamous carcinoma cells through downregulating the activation of NF-κB signaling pathway in vitro and in vivo. Acta. Bioch. Bioph. Sin. 2012, 44, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Hang, Y.; Liu, J.; Hou, Y.; Wang, N.; Wang, M. Anticancer effect of curcumin inhibits cell growth through miR-21/PTEN/Akt pathway in breast cancer cell. Oncol. Lett. 2017, 13, 4825–4831. [Google Scholar] [CrossRef] [Green Version]
- Amaral, J.D.; Xavier, J.M.; Steer, C.J.; Rodrigues, C.M.P. Targeting the p53 pathway of apoptosis. Curr. Pharm. Design 2010, 16, 2493–2503. [Google Scholar] [CrossRef]
- Choudhuri, T.; Pal, S.; Agwarwal, M.L.; Das, T.; Sa, G. Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. FEBS Lett. 2002, 512, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wang, Y.; Song, Y.; Xu, L.; Zhao, J.; Fang, B. A preliminary study of the effect of curcumin on the expression of p53 protein in a human multiple myeloma cell line. Oncol. Lett. 2015, 9, 1719–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallman, K.; Aleck, K.; Dwyer, B.; Lloyd, V.; Quigley, M.; Sitto, N.; Siebert, A.E.; Dinda, S. The effects of turmeric (curcumin) on tumor suppressor protein (p53) and estrogen receptor (ERα) in breast cancer cells. Breast Cancer Target. Ther. 2017, 9, 153. [Google Scholar] [CrossRef] [Green Version]
- Shankar, S.; Chen, Q.; Sarva, K.; Siddiqui, I.; Srivastava, R.K. Curcumin enhances the apoptosis-inducing potential of TRAIL in prostate cancer cells: Molecular mechanisms of apoptosis, migration and angiogenesis. J. Mol. Signal 2007, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Takada, Y.; Singh, S.; Myers, J.N.; Aggarwal, B.B. Inhibition of growth and survival of human head and neck squamous cell carcinoma cells by curcumin via modulation of nuclear factor-κB signaling. Int. J. Cancer 2004, 111, 679–692. [Google Scholar] [CrossRef]
- Shakor, A.B.A.; Atia, M.; Ismail, I.A.; Alshehri, A.; El-Refaey, H.; Kwiatkowska, K.; Sobota, A. Curcumin induces apoptosis of multidrug-resistant human leukemia HL60 cells by complex pathways leading to ceramide accumulation. Biochim. Biophys. Acta 2014, 1841, 1672–1682. [Google Scholar] [CrossRef]
- Shakor, A.B.A.; Atia, M.; Alshehri, A.S.; Sobota, A.; Kwiatkowska, K. Ceramide generation during curcumin-induced apoptosis is controlled by crosstalk among Bcl-2, Bcl-xL, caspases and glutathione. Cell Signal. 2015, 27, 2220–2230. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.-L.; Li, J.; Qin, Z.-H.; Liang, Z.-Q. Autophagic and apoptotic mechanisms of curcumin-induced death in K562 cells. J. Asian Nat. Prod. Res. 2009, 11, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.J.; Pandey, S. Chemo-resistant melanoma sensitized by tamoxifen to low dose curcumin treatment through induction of apoptosis and autophagy. Cancer Biol. Ther. 2011, 11, 216–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.-D.; Liu, X.-E.; Huang, D.-S. Curcumin induces apoptosis of triple-negative breast cancer cells by inhibition of EGFR expression. Mol. Med. Rep. 2012, 6, 1267–1270. [Google Scholar] [CrossRef] [Green Version]
- Almasan, A.; Ashkenazi, A. Apo2L/TRAIL: Apoptosis signaling, biology, and potential for cancer therapy. Cytokine Growth Factor Rev. 2003, 14, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Park, S.; Cho, D.H.; Andera, L.; Suh, N.; Kim, I. Curcumin enhances TRAIL-induced apoptosis of breast cancer cells by regulating apoptosis-related proteins. Mol. Cell Biochem. 2013, 383, 39–48. [Google Scholar] [CrossRef]
- Koohpar, Z.K.; Entezari, M.; Movafagh, A.; Hashemi, M. Anticancer activity of curcumin on human breast adenocarcinoma: Role of Mcl-1 gene. Iran. J. Cancer Prev. 2015, 8, e2331. [Google Scholar] [CrossRef]
- Deeb, D.; Jiang, H.; Gao, X.; Hafner, M.S.; Wong, H.; Divine, G.; Chapman, R.A.; Dulchavsky, S.A.; Gautam, S.C. Curcumin sensitizes prostate cancer cells to tumor necrosis factor–related apoptosis-inducing ligand/Apo2L by inhibiting nuclear factor-κB through suppression of IκBα phosphorylation. Mol. Cancer Ther. 2004, 3, 803–812. [Google Scholar] [CrossRef]
- Andrzejewski, T.; Deeb, D.; Gao, X.; Danyluk, A.; Arbab, A.S.; Dulchavsky, S.A.; Gautam, S.C. Therapeutic efficacy of curcumin/TRAIL combination regimen for hormone-refractory prostate cancer. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2008, 17, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; Lim, J.H.; Lee, T.J.; Park, J.-W.; Choi, K.S.; Kwon, T.K. Curcumin sensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through reactive oxygen species-mediated upregulation of death receptor 5 (DR5). Carcinogenesis 2005, 26, 1905–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, H.; Tan, L.; Griffith, K.; Choi, M.; Liu, J.R. Curcumin enhances Apo2L/TRAIL-induced apoptosis in chemoresistant ovarian cancer cells. Gynecol. Oncol. 2007, 105, 104–112. [Google Scholar] [CrossRef]
- Kwon, D.; Oh, S.; Park, J.-H.; Lee, S.; Lee, S. The TRAIL sensitization effect of substituted triazolyl curcumin mimics against brain cancer cells. Bull. Korean Chem. Soc. 2014, 35, 3339–3342. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.L.; Greenshields, A.; Hill, R.; Hilchie, A.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. Curcumin-induced apoptosis in ovarian carcinoma cells is p53-independent and involves p38 mitogen-activated protein kinase activation and downregulation of Bcl-2 and survivin expression and Akt signaling. Mol. Carcinog. 2010, 49, 13–24. [Google Scholar] [CrossRef]
- Calaf, G.M.; Echiburú-Chau, C.; Roy, D.; Chai, Y.; Wen, G.; Balajee, A.S. Protective role of curcumin in oxidative stress of breast cells. Oncol. Rep. 2011, 26, 1029–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-Y.; Lin, M.-T.; Zhou, M.-J.; Yi, T.; Tang, Y.-N.; Tang, S.-L.; Yang, Z.-J.; Zhao, Z.-Z.; Chen, H.-B. Combinational treatment of curcumin and quercetin against gastric cancer MGC-803 cells in vitro. Molecules 2015, 20, 11524–11534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.T.; Huang, A.C.; Tang, N.Y.; Liu, H.C.; Liao, C.L.; Ji, B.C.; Chou, Y.C.; Yang, M.D.; Lu, H.F.; Chung, J.G. Bisdemethoxycurcumin-induced S phase arrest through the inhibition of cyclin A and E and induction of apoptosis via endoplasmic reticulum stress and mitochondria-dependent pathways in human lung cancer NCI H 460 cells. Environ. Toxicol. 2016, 31, 1899–1908. [Google Scholar] [CrossRef]
- Ko, Y.-C.; Lien, J.-C.; Liu, H.-C.; Hsu, S.-C.; Ji, B.-C.; Yang, M.-D.; Hsu, W.-H.; Chung, J.-G. Demethoxycurcumin induces the apoptosis of human lung cancer NCI-H460 cells through the mitochondrial-dependent pathway. Oncol. Rep. 2015, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhang, J.; Guo, G.; Cai, Y.; Cui, R.; Yin, C.; Liu, W.; Vinothkumar, R.; Zhang, T.; Liang, G. A mono-carbonyl analog of curcumin induces apoptosis in drug-resistant EGFR-mutant lung cancer through the generation of oxidative stress and mitochondrial dysfunction. Cancer Manag. Res. 2018, 10, 3069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semlali, A.; Contant, C.; Al-Otaibi, B.; Al-Jammaz, I.; Chandad, F. The curcumin analog (PAC) suppressed cell survival and induced apoptosis and autophagy in oral cancer cells. Sci. Rep. 2021, 11, 11701. [Google Scholar] [CrossRef]
- Miki, H.; Uehara, N.; Kimura, A.; Sasaki, T.; Yuri, T.; Yoshizawa, K.; Tsubura, A. Resveratrol induces apoptosis via ROS-triggered autophagy in human colon cancer cells. Int. J. Oncol. 2012, 40, 1020–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamin, L.L.; Filippi-Chiela, E.C.; Dillenburg-Pilla, P.; Horn, F.; Salbego, C.; Lenz, G. Resveratrol and quercetin cooperate to induce senescence-like growth arrest in C6 rat glioma cells. Cancer Sci. 2009, 100, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Cardile, V.; Scifo, C.; Russo, A.; Falsaperla, M.; Morgia, G.; Motta, M.; Renis, M.; Imbriani, E.; Silvestre, G. Involvement of HSP70 in resveratrol-induced apoptosis of human prostate cancer. Anticancer Res. 2003, 23, 4921–4926. [Google Scholar] [PubMed]
- Singh, B.N.; Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy which leads to apoptotic cell death through inhibition of PI3K/Akt/mTOR pathway in human pancreatic cancer stem cells. Biochem. Pharmacol. 2012, 84, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lim, W.; Song, G. Chrysophanol selectively represses breast cancer cell growth by inducing reactive oxygen species production and endoplasmic reticulum stress via AKT and mitogen-activated protein kinase signal pathways. Toxicol. Appl. Pharmacol. 2018, 360, 201–211. [Google Scholar] [CrossRef]
- Nishimura, R.; Tabata, K.; Arakawa, M.; Ito, Y.; Kimura, Y.; Akihisa, T.; Nagai, H.; Sakuma, A.; Kohno, H.; Suzuki, T. Isobavachalcone, a chalcone constituent of Angelica keiskei, induces apoptosis in neuroblastoma. Biol. Pharm. Bull. 2007, 30, 1878–1883. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Ma, C.-M.; Liu, C.-X.; Wei, W.; Sun, Y.; Yan, H.; Wu, Y.-L. Autophagy inhibition enhances isobavachalcone-induced cell death in multiple myeloma cells. Int. J. Mol. Med. 2012, 30, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingston, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin Q: A selective STAT3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Chiu, M.H.; Nie, R.L.; Cordell, G.A.; Qiu, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, U.; Aeri, V.; Mir, S.R. Cucurbitacins-An insight into medicinal leads from nature. Pharmacogn Rev. 2015, 9, 12–18. [Google Scholar] [PubMed] [Green Version]
- Gao, L.-F.; Xu, D.-Q.; Wen, L.-J.; Zhang, X.-Y.; Shao, Y.-T.; Zhao, X.-J. Inhibition of STAT3 expression by siRNA suppresses growth and induces apoptosis in laryngeal cancer cells. Acta Pharmacol. Sin. 2005, 26, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, M.; Zhang, H.; Sun, C.; Deng, Y. Inhibitory effects of cucurbitacin B on laryngeal squamous cell carcinoma. Eur. Arch. Otorhinol. 2008, 265, 1225–1232. [Google Scholar] [CrossRef]
- Ren, X.; Zhao, B.; Chang, H.; Xiao, M.; Wu, Y.; Liu, Y. Paclitaxel suppresses proliferation and induces apoptosis through regulation of ROS and the AKT/MAPK signaling pathway in canine mammary gland tumor cells. Mol. Med. Rep. 2018, 17, 8289–8299. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Xiao, L.; Li, Z.; Hu, J. The relationship between p38MAPK and apoptosis during paclitaxel resistance of ovarian cancer cells. J. Huazhong Univ. Sci. Technol. 2007, 27, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Boronkai, A.; Bellyei, S.; Szigeti, A.; Pozsgai, E.; Bognar, Z.; Sumegi, B.; Gallyas, F., Jr. Potentiation of paclitaxel-induced apoptosis by galectin-13 overexpression via activation of Ask-1-p38-MAP kinase and JNK/SAPK pathways and suppression of Akt and ERK1/2 activation in U-937 human macrophage cells. Eur. J. Cell Biol. 2009, 88, 753–763. [Google Scholar] [CrossRef]
- Kim, H.J.; Yim, G.W.; Nam, E.J.; Kim, Y.T. Synergistic effect of COX-2 inhibitor on paclitaxel-induced apoptosis in the human ovarian cancer cell line OVCAR-3. Cancer Res. Treat. 2014, 46, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Liu, D.; Yao, Y.; Feng, X.; Huang, X.; Tang, Y.; Yang, J.; Wang, X. Paclitaxel induces apoptosis in leukemia cells through a JNK activation-dependent pathway. Genet. Mol. Res. 2016, 15, 15013904. [Google Scholar] [CrossRef] [PubMed]
- Im-Sun, W.; Jang, H.-S.; Eun, S.Y.; Kim, H.J.; Ham, S.A.; Kim, H.J.; Lee, J.H.; Chang, K.C.; Kim, J.-H.; Han, C.W.; et al. Ran suppresses paclitaxel-induced apoptosis in human glioblastoma cells. Apoptosis 2008, 13, 1223–1231. [Google Scholar]
- Xu, J.; Su, C.; Zhao, F.; Tao, J.; Hu, D.; Shi, A.; Pan, J.; Zhang, Y. Paclitaxel promotes lung cancer cell apoptosis via MEG3-P53 pathway activation. Biochem. Bioph. Res. Commun. 2018, 504, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Razi, S.S.; Rehmani, S.; Li, X.; Park, K.; Schwartz, G.S.; Latif, M.J.; Bhora, F.Y. Antitumor activity of paclitaxel is significantly enhanced by a novel proapoptotic agent in non–small cell lung cancer. J. Surg. Res. 2015, 194, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Oommen, S.; Anto, R.J.; Srinivas, G.; Karunagaran, D. Allicin (from garlic) induces caspase-mediated apoptosis in cancer cells. Eur. J. Pharmacol. 2004, 485, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Bat-Chen, W.; Golan, T.; Peri, I.; Ludmer, Z.; Schwartz, B. Allicin purified from fresh garlic cloves induces apoptosis in colon cancer cells via Nrf2. Nutr. Cancer 2010, 62, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-Q.; Tuersun, H.; Jiao, S.-J.; Zheng, J.-H.; Xiao, J.-B.; Hasim, A. Functional role of NRF2 in cervical carcinogenesis. PLoS ONE 2015, 10, e0133876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, D. Allicin suppresses the migration and invasion in cervical cancer cells mainly by inhibiting NRF2. Exp. Ther. Med. 2019, 17, 1523–1528. [Google Scholar] [CrossRef]
- Zhang, W.; Ha, M.; Gong, Y.; Xu, Y.; Dong, N.; Yuan, Y. Allicin induces apoptosis in gastric cancer cells through activation of both extrinsic and intrinsic pathways. Oncol. Rep. 2010, 24, 1585–1592. [Google Scholar] [PubMed] [Green Version]
- Rosas-González, V.C.; Téllez-Bañuelos, M.C.; Hernández-Flores, G.; Bravo-Cuellar, A.; Aguilar-Lemarroy, A.; Jave-Suárez, L.F.; Haramati, J.; Solorzano-Ibarra, F.; Ortiz-Lazareno, P.C. Differential effects of alliin and allicin on apoptosis and senescence in luminal A and triple-negative breast cancer: Caspase, ΔΨm, and pro-apoptotic gene involvement. Fundam. Clin. Pharm. 2020, 34, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.-B.; Yoo, S.-J.; Ryu, D.-G.; Yang, J.-Y.; Rho, H.-W.; Kim, J.-S.; Park, J.-W.; Kim, H.-R.; Park, B.-H. Induction of apoptosis by diallyl disulfide through activation of caspase-3 in human leukemia HL-60 cells. Biochem. Pharmacol. 2002, 63, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.F.; Sue, C.C.; Yu, C.S.; Chen, S.C.; Chen, G.W.; Chung, J.G. Diallyl disulfide (DADS) induced apoptosis undergo caspase-3 activity in human bladder cancer T24 cells. Food Chem. Toxicol. 2004, 42, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Altonsy, M.O.; Andrews, S.C. Diallyl disulphide, a beneficial component of garlic oil, causes a redistribution of cell-cycle growth phases, induces apoptosis, and enhances butyrate-induced apoptosis in colorectal adenocarcinoma cells (HT-29). Nutr. Cancer 2011, 63, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef]
- Romero-Ramirez, L.; Cao, H.; Nelson, D.; Hammond, E.; Lee, A.H.; Yoshida, H.; Mori, K.; Glimcher, L.H.; Denko, N.C.; Giaccia, A.J.; et al. XBP1 is essential for survival under hypoxic conditions and is required for tumor growth. Cancer Res. 2004, 64, 5943–5947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, M.; Naczki, C.; Koritzinsky, M.; Fels, D.; Blais, J.; Hu, N.; Harding, H.; Novoa, I.; Varia, M.; Raleigh, J.; et al. ER stress-regulated translation increases tolerance to extreme hypoxia and promotes tumor growth. EMBO J. 2005, 24, 3470–3481. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yang, Z.Q.; Zhang, K. Endoplasmic reticulum stress response in cancer: Molecular mechanism and therapeutic potential. Am. J. Transl. Res. 2010, 2, 65–74. [Google Scholar] [PubMed]
- Sui, X.; Chen, R.; Wang, Z.; Huang, Z.; Kong, N.; Zhang, M.; Han, W.; Lou, F.; Yang, J.; Zhang, Q.; et al. Autophagy and chemotherapy resistance: A promising therapeutic target for cancer treatment. Cell Death Dis. 2013, 4, e838. [Google Scholar] [CrossRef] [Green Version]
- Raj Kumar, Y.; Soo-Wan, C.; Hyung-Ryong, K.; Chae, H.J. Endoplasmic reticulum stress and cancer. J. Cancer Prev. 2014, 19, 75–88. [Google Scholar]
- Lindholm, D.; Wootz, H.; Korhonen, L. ER stress and neurodegenerative diseases. Cell Death Differ. 2006, 13, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozen, E.; Karademir, B.; Ozer, N.K. Basic mechanisms in endoplasmic reticulum stress and relation to cardiovascular diseases. Free Radical. Biol. Med. 2015, 78, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Kelloff, G.J.; Crowell, J.A.; Steele, V.E.; Lubet, R.A.; Malone, W.A.; Boone, C.W.; Kopelovich, L.; Hawk, E.T.; Lieberman, R.; Lawrence, J.A.; et al. Progress in cancer chemoprevention: Development of diet-derived chemopreventive agents. J. Nutr. 2000, 130, 467s–471s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharmacol Res. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Limonta, P.; Moretti, R.M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Montagnani Marelli, M. Role of endoplasmic reticulum stress in the anticancer activity of natural compounds. Int. J. Mol. Med. 2019, 20, 961. [Google Scholar] [CrossRef] [PubMed]
- Madhavi, B.B.; Nath, A.R.; Banji, D.; Madhu, M.N.; Ramalingam, R.; Swetha, D. Extraction, identification, formulation and evaluation of piperine in alginate beads. Int. J. Pharm. Pharm. 2009, 1, 156–161. [Google Scholar]
- Yaffe, P.B.; Power Coombs, M.R.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1 arrest and apoptosis triggered by endoplasmic reticulum stress. Mol. Carcinog. 2015, 54, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Wolpin, B.M.; Mayer, R.J. Systemic treatment of colorectal cancer. Gastroenterology 2008, 134, 1296–1310.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-F.; Zhu, D.-J.; Chen, X.-W.; Chen, Q.-K.; Luo, Z.-T.; Liu, C.-C.; Wang, G.-X.; Zhang, W.-J.; Liao, N.-Z. Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget 2017, 8, 40264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinotti, S.; Ranzato, E.; Burlando, B. (−)-Epigallocatechin-3-gallate induces GRP78 accumulation in the ER and shifts mesothelioma constitutive UPR into proapoptotic ER stress. J. Cell Physiol. 2018, 233, 7082–7090. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tian, L.; Long, L.; Quan, M.; Liu, F.; Cao, J. Casticin potentiates TRAIL-induced apoptosis of gastric cancer cells through endoplasmic reticulum stress. PLoS ONE 2013, 8, e58855. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Shu, F.; Liang, X.; Chang, H.; Shi, L.; Peng, X.; Zhu, J.; Mi, M. Ampelopsin induces cell growth inhibition and apoptosis in breast cancer cells through ROS generation and endoplasmic reticulum stress pathway. PLoS ONE 2014, 9, e89021. [Google Scholar] [CrossRef]
- Huang, A.C.; Chang, C.L.; Yu, C.S.; Chen, P.Y.; Yang, J.S.; Ji, B.C.; Lin, T.P.; Chiu, C.F.; Yeh, S.P.; Huang, Y.P. Induction of apoptosis by curcumin in murine myelomonocytic leukemia WEHI-3 cells is mediated via endoplasmic reticulum stress and mitochondria-dependent pathways. Environ. Toxicol. 2013, 28, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Rivera, M.; Ramos, Y.; Rodriguez-Valentin, M.; López-Acevedo, S.; Cubano, L.A.; Zou, J.; Zhang, Q.; Wang, G.; Boukli, N.M. Targeting multiple pro-apoptotic signaling pathways with curcumin in prostate cancer cells. PLoS ONE 2017, 12, e0179587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.L.; Poklepovic, A.; Booth, L. Curcumin interacts with sildenafil to kill GI tumor cells via endoplasmic reticulum stress and reactive oxygen/nitrogen species. Oncotarget 2017, 8, 99451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.-H.; Hang, L.-W.; Yang, J.-S.; Chen, H.-Y.; Lin, H.-Y.; Chiang, J.-H.; Lu, C.-C.; Yang, J.-L.; Lai, T.-Y.; Ko, Y.-C. Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade-and mitochondria-dependent pathways. Anticancer Res. 2010, 30, 2125–2133. [Google Scholar] [PubMed]
- Kuo, Y.-F.; Su, Y.-Z.; Tseng, Y.-H.; Wang, S.-Y.; Wang, H.-M.; Chueh, P.J. Flavokawain B, a novel chalcone from Alpinia pricei Hayata with potent apoptotic activity: Involvement of ROS and GADD153 upstream of mitochondria-dependent apoptosis in HCT116 cells. Free Radical. Biol. Med. 2010, 49, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, M.; Chen, Y.; Wang, X.J.; Tang, X. Resveratrol: An overview of its anti-cancer mechanisms. Life Sci. 2018, 207, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-M.; Galson, D.L.; Roodman, G.D.; Ouyang, H. Resveratrol triggers the pro-apoptotic endoplasmic reticulum stress response and represses pro-survival XBP1 signaling in human multiple myeloma cells. Exp. Hematol. 2011, 39, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.R.; Kim, S.M.; Hwang, K.A.; Kang, J.H.; Choi, K.C. Resveratrol induced reactive oxygen species and endoplasmic reticulum stress-mediated apoptosis, and cell cycle arrest in the A375SM malignant melanoma cell line. Int. J. Mol. Med. 2018, 42, 1427–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, C.; Pan-Castillo, B.; Valls, C.; Pujadas, G.; Garcia-Vallve, S.; Arola, L.; Mulero, M. Resveratrol enhances palmitate-induced ER stress and apoptosis in cancer cells. PLoS ONE 2014, 9, e113929. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Dong, D.-S.; Pei, L. Synergistic antitumor activity of resveratrol and miR-200c in human lung cancer. Oncol. Rep. 2014, 31, 2293–2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, S.-E.; Kao, C.-H.; Liu, Y.-T.A.; Cheng, M.-L.; Yang, Y.-W.; Huang, Y.-K.; Hsu, C.-C.; Wang, J.-S. Resveratrol induced ER expansion and ER caspase-mediated apoptosis in human nasopharyngeal carcinoma cells. Apoptosis 2014, 19, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, C.; Jiang, X.; Zhang, Z. ROS-mediated endoplasmic reticulum stress and mitochondrial dysfunction underlie apoptosis induced by resveratrol and arsenic trioxide in A549 cells. Chem.-Biol. Interact. 2016, 245, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.-X.; Yao, X.-J.; Xu, S.W.; Wong, V.K.-W.; He, J.-X.; Ding, J.; Xue, W.-W.; Mujtaba, T.; Michelangeli, F.; Huang, M. (Z) 3, 4, 5, 4′-trans-tetramethoxystilbene, a new analogue of resveratrol, inhibits gefitinb-resistant non-small cell lung cancer via selectively elevating intracellular calcium level. Sci. Rep. 2015, 5, 16348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Choi, W.G.; Lee, P.J.; Chung, S.W.; Kim, B.S.; Chung, H.T.; Cho, S.; Kim, J.H.; Kang, B.H.; Kim, H. The novel resveratrol derivative 3, 5-diethoxy-3′, 4′-dihydroxy-trans-stilbene induces mitochondrial ROS-mediated ER stress and cell death in human hepatoma cells in vitro. Acta Pharmacol. Sin. 2017, 38, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-Y.; Wang, Y.-T.; Lin, L.-G. New insights into the anti-obesity activity of xanthones from Garcinia mangostana. Food Funct. 2015, 6, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Fujiwara, H.; Oku, H.; Ishiguro, K.; Ohizumi, Y. α-Mangostin induces Ca2+-ATPase-dependent apoptosis via mitochondrial pathway in PC12 cells. J. Pharmacol. Sci. 2004, 95, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-H.; Liu, Q.-Y.; Li, T.; Liu, J.-L.; Chen, X.; Huang, L.; Qiang, W.-A.; Chen, X.; Wang, Y.; Lin, L.-G. Garcinone E induces apoptosis and inhibits migration and invasion in ovarian cancer cells. Sci. Rep. 2017, 7, 10718. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Petiwala, S.M.; Yan, M.; Won, J.H.; Petukhov, P.A.; Johnson, J.J. Gartanin, an isoprenylated xanthone from the mangosteen fruit (Garcinia mangostana), is an androgen receptor degradation enhancer. Mol. Nutr. Food Res. 2016, 60, 1458–1469. [Google Scholar] [CrossRef] [PubMed]
- Sheu, M.L.; Liu, S.H.; Lan, K.H. Honokiol induces calpain-mediated glucose-regulated protein-94 cleavage and apoptosis in human gastric cancer cells and reduces tumor growth. PLoS ONE 2007, 2, e1096. [Google Scholar] [CrossRef]
- Cheng, A.-C.; Tsai, M.-L.; Liu, C.-M.; Lee, M.-F.; Nagabhushanam, K.; Ho, C.-T.; Pan, M.-H. Garcinol inhibits cell growth in hepatocellular carcinoma Hep3B cells through induction of ROS-dependent apoptosis. Food Funct. 2010, 1, 301–307. [Google Scholar] [CrossRef]
- Siu, F.M.; Ma, D.L.; Cheung, Y.W.; Lok, C.N.; Yan, K.; Yang, Z.; Yang, M.; Xu, S.; Ko, B.C.; He, Q.Y.; et al. Proteomic and transcriptomic study on the action of a cytotoxic saponin (Polyphyllin D): Induction of endoplasmic reticulum stress and mitochondria-mediated apoptotic pathways. Proteomics 2008, 8, 3105–3117. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-M.; Bai, L.-L.; Zhang, D.-M.; Yiu, A.; Yin, Z.-Q.; Han, W.-L.; Liu, J.-S.; Li, Y.; Fu, D.-Y.; Ye, W.-C. Saxifragifolin D induces the interplay between apoptosis and autophagy in breast cancer cells through ROS-dependent endoplasmic reticulum stress. Biochem. Pharmacol. 2013, 85, 913–926. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.Y.; Hsu, Y.L.; Ni, W.C.; Tsai, Y.M.; Yang, C.J.; Kuo, P.L.; Huang, M.S. Oxidative and endoplasmic reticulum stress signaling are involved in dehydrocostuslactone-mediated apoptosis in human non-small cell lung cancer cells. Lung Cancer 2010, 68, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Park, I.-J.; Kim, M.-J.; Park, O.J.; Choe, W.; Kang, I.; Kim, S.-S.; Ha, J. Cryptotanshinone induces ER stress-mediated apoptosis in HepG2 and MCF7 cells. Apoptosis 2012, 17, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Aghaei, M.; Ghanadian, M.; Sajjadi, S.E.; Saghafian, R.; Keyvanloo Shahrestanaki, M. Pimpinelol, a novel atypical Sesquiterpene lactone from Pimpinella haussknechtii fruits with evaluation of endoplasmic reticulum stress in breast cancer cells. Fitoterapia 2018, 129, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-S.; Dang, Y.-Y.; Guo, J.-J.; Wu, G.-S.; Lu, J.-J.; Chen, X.-P.; Wang, Y.-T. Furanodiene induces endoplasmic reticulum stress and presents antiproliferative activities in lung cancer cells. Evid. -Based Complement. Altern. Med. 2012, 2012, 426521. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, X.; Su, L. Parthenolide induces apoptosis via TNFRSF10B and PMAIP1 pathways in human lung cancer cells. J. Exp. Clin. Cancer Res. 2014, 33, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Miyamoto, S.; Yasui, Y.; Oyama, T.; Murakami, A.; Tanaka, T. Zerumbone, a tropical ginger sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int. J. Cancer 2009, 124, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Edagawa, M.; Kawauchi, J.; Hirata, M.; Goshima, H.; Inoue, M.; Okamoto, T.; Murakami, A.; Maehara, Y.; Kitajima, S. Role of activating transcription factor 3 (ATF3) in endoplasmic reticulum (ER) stress-induced sensitization of p53-deficient human colon cancer cells to tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis through up-regulation of death receptor 5 (DR5) by zerumbone and celecoxib. J. Biol. Chem. 2014, 289, 21544–21561. [Google Scholar] [PubMed] [Green Version]
- Huang, X.; Zhang, Z.; Jia, L.; Zhao, Y.; Zhang, X.; Wu, K. Endoplasmic reticulum stress contributes to vitamin E succinate-induced apoptosis in human gastric cancer SGC-7901 cells. Cancer Lett. 2010, 296, 123–131. [Google Scholar] [CrossRef]
- Huang, X.; Li, L.; Zhang, L.; Zhang, Z.; Wang, X.; Zhang, X.; Hou, L.; Wu, K. Crosstalk between endoplasmic reticulum stress and oxidative stress in apoptosis induced by α-tocopheryl succinate in human gastric carcinoma cells. Brit. J. Nutr. 2012, 109, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Alves-Fernandes, D.K.; de Oliveira, É.A.; Faião-Flores, F.; Alicea-Rebecca, G.; Weeraratna, A.T.; Smalley, K.S.; de Moraes Barros, S.B.; Maria-Engler, S.S. ER stress promotes antitumor effects in BRAFi/MEKi resistant human melanoma induced by natural compound 4-nerolidylcathecol (4-NC). Pharmacol. Res. 2019, 141, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-C.; Lai, K.-H.; Su, J.-H.; Wu, Y.-J.; Sheu, J.-H. 7-Acetylsinumaximol B induces apoptosis and autophagy in human gastric carcinoma cells through mitochondria dysfunction and activation of the PERK/eIF2α/ATF4/CHOP signaling pathway. Mar. Drugs 2018, 16, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.K.; Sanders, B.G.; Kline, K. Tocotrienols induce apoptosis in breast cancer cell lines via an endoplasmic reticulum stress-dependent increase in extrinsic death receptor signaling. Breast Cancer Res. Treat. 2010, 124, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, V.; Nepal, A.; Olaisen, C.; Bachke, S.; Hira, J.; Søgaard, C.K.; Røst, L.M.; Misund, K.; Andreassen, T.; Melø, T.M. Anti-cancer potential of homemade fresh garlic extract is related to increased endoplasmic reticulum stress. Nutrients 2018, 10, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaschula, C.H.; Hunter, R.; Cotton, J.; Tuveri, R.; Ngarande, E.; Dzobo, K.; Schäfer, G.; Siyo, V.; Lang, D.; Kusza, D.A. The garlic compound ajoene targets protein folding in the endoplasmic reticulum of cancer cells. Mol. Carcinogen 2016, 55, 1213–1228. [Google Scholar] [CrossRef]
- Rizzi Siyo, V.; Schäfer, G.; Hunter, R.; Grafov, A.; Grafova, I.; Nieger, M.; Katz, A.A.; Parker, M.I.; Kaschula, C.H. The cytotoxicity of the ajoene analogue BisPMB in WHCO1 oesophageal cancer cells is mediated by CHOP/GADD153. Molecules 2017, 22, 892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzi, F.; Naponelli, V.; Silva, A.; Modernelli, A.; Ramazzina, I.; Bonacini, M.; Tardito, S.; Gatti, R.; Uggeri, J.; Bettuzzi, S. Polyphenon E®, a standardized green tea extract, induces endoplasmic reticulum stress, leading to death of immortalized PNT1a cells by anoikis and tumorigenic PC3 by necroptosis. Carcinogenesis 2014, 35, 828–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmer, T.G.; Ward, M.D.; Hait, W.N. Effects of rottlerin, an inhibitor of calmodulin-dependent protein kinase III, on cellular proliferation, viability, and cell cycle distribution in malignant glioma cells. Cell Growth Differ. 1997, 8, 327–334. [Google Scholar] [PubMed]
- Bove, K.; Lincoln, D.W.; Tsan, M.-F. Effect of resveratrol on growth of 4T1 breast cancer cells in vitro and in vivo. Biochem. Bioph. Res. Commun. 2002, 291, 1001–1005. [Google Scholar] [CrossRef]
- Asensi, M.; Medina, I.; Ortega, A.; Carretero, J.; Baño, M.C.; Obrador, E.; Estrela, J.M. Inhibition of cancer growth by resveratrol is related to its low bioavailability. Free Radical. Biol. Med. 2002, 33, 387–398. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.C.; Tseng, S.H. Tetrandrine suppresses tumor growth and angiogenesis of gliomas in rats. Int. J. Cancer 2009, 124, 2260–2269. [Google Scholar] [CrossRef]
- Zhuang, W.; Long, L.; Zheng, B.; Ji, W.; Yang, N.; Zhang, Q.; Liang, Z. Curcumin promotes differentiation of glioma-initiating cells by inducing autophagy. Cancer Sci. 2012, 103, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.-Y.; Zhang, L.; Lugea, A.; Moro, A.; Edderkaoui, M.; Eibl, G.; Pandol, S.J.; Go, V.-L.W. Determination of rottlerin, a natural protein kinases C inhibitor, in pancreatic cancer cells and mouse xenografts by RP-HPLC method. Int. J. Chromatogr. Sep. Tech. 2013, 4, 100062. [Google Scholar]
- El-Magd, M.A.; Khamis, A.; Eldeen, S.K.N.; Ibrahim, W.M.; Salama, A.F. Trehalose enhances the antitumor potential of methotrexate against mice bearing Ehrlich ascites carcinoma. Biomed. Pharmacother. 2017, 92, 870–878. [Google Scholar] [CrossRef]
- Guéritte, F.; Fahy, J. The Vinca Alkaloids. In Anticancer Agents from Natural Products; CRC Press: Boca Raton, FL, USA, 2005; Volume 10. [Google Scholar]
- Young, S.D.; Whissell, M.; Noble, J.C.; Cano, P.O.; Lopez, P.G.; Germond, C.J. Phase II clinical trial results involving treatment with low-dose daily oral cyclophosphamide, weekly vinblastine, and rofecoxib in patients with advanced solid tumors. Clin. Cancer Res. 2006, 12, 3092–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galsky, M.D.; Dritselis, A.; Kirkpatrick, P.; Oh, W.K. Cabazitaxel. Nat. Rev. Drug Discov. 2010, 9, 677–678. [Google Scholar] [CrossRef] [PubMed]
- Mita, A.C.; Denis, L.J.; Rowinsky, E.K.; DeBono, J.S.; Goetz, A.D.; Ochoa, L.; Forouzesh, B.; Beeram, M.; Patnaik, A.; Molpus, K. Phase I and pharmacokinetic study of XRP6258 (RPR 116258A), a novel taxane, administered as a 1-hour infusion every 3 weeks in patients with advanced solid tumors. Clin. Cancer Res. 2009, 15, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivot, X.; Koralewski, P.; Hidalgo, J.; Chan, A.; Goncalves, A.; Schwartsmann, G.; Assadourian, S.; Lotz, J. A multicenter phase II study of XRP6258 administered as a 1-h iv infusion every 3 weeks in taxane-resistant metastatic breast cancer patients. Ann. Oncol. 2008, 19, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- De Bono, J.S.; Oudard, S.; Ozguroglu, M.; Hansen, S.; Machiels, J.-P.; Kocak, I.; Gravis, G.; Bodrogi, I.; Mackenzie, M.J.; Shen, L. Prednisone plus cabazitaxel or mitoxantrone for metastatic castration-resistant prostate cancer progressing after docetaxel treatment: A randomised open-label trial. Lancet 2010, 376, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Abidi, A. Cabazitaxel: A novel taxane for metastatic castration-resistant prostate cancer-current implications and future prospects. J. Pharmacol. Pharmacother. 2013, 4, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Tsao, C.-K.; Cutting, E.; Martin, J.; Oh, W.K. The role of cabazitaxel in the treatment of metastatic castration-resistant prostate cancer. Ther. Adv. Urol. 2014, 6, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, D.P.; Emerson, D.L.; Qian, Y.; Anwar, T.; Liewehr, D.J.; Steinberg, S.M.; Silberman, S.; Palmieri, D.; Steeg, P.S. TPI-287, a new taxane family member, reduces the brain metastatic colonization of breast cancer cells. Mol. Cancer Ther. 2012, 11, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuade, J.L.; Posada, L.P.; Lecagoonporn, S.; Cain, S.; Bassett, R.L., Jr.; Patel, S.P.; Hwu, W.-J.; Hwu, P.; Davies, M.A.; Bedikian, A.Y. A phase I study of TPI 287 in combination with temozolomide for patients with metastatic melanoma. Melanoma. Res. 2016, 26, 604–608. [Google Scholar] [CrossRef] [Green Version]
- Hendrickson, A.E.W.; Menefee, M.E.; Hartmann, L.C.; Long, H.J.; Northfelt, D.W.; Reid, J.M.; Boakye-Agyeman, F.; Kayode, O.; Flatten, K.S.; Harrell, M.I. A phase I clinical trial of the poly (ADP-ribose) polymerase inhibitor veliparib and weekly topotecan in patients with solid tumors. Clin. Cancer Res. 2018, 24, 744–752. [Google Scholar] [CrossRef]
- Ernani, V.; Jahan, R.; Smith, L.M.; Marr, A.S.; Kimbrough, S.E.; Kos, M.E.; Tijerina, J.; Pivovar, S.; Lakshmanan, I.; Ketcham, M.; et al. A phase I study of weekly doxorubicin and oral topotecan for patients with relapsed or refractory small cell lung cancer (SCLC): A Fred and Pamela Buffet Cancer Center Clinical Trials Network study. Cancer Treat. Res. Commun. 2020, 22, 100162. [Google Scholar] [CrossRef]
- Zhu, A.X.; Ready, N.; Clark, J.W.; Safran, H.; Amato, A.; Salem, N.; Pace, S.; He, X.; Zvereva, N.; Lynch, T.J. Phase I and pharmacokinetic study of gimatecan given orally once a week for 3 of 4 weeks in patients with advanced solid tumors. Clin. Cancer Res. 2009, 15, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.F.; Sang, C.Y.; Xu, X.H.; Zhang, L.L.; Yang, X.; Hui, L.; Zhang, J.B.; Chen, S.W. Synthesis and cytotoxic activity on human cancer cells of carbamate derivatives of 4β-(1,2,3-triazol-1-yl)podophyllotoxin. Eur. J. Med. Chem. 2013, 64, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Grossman, S.A.; Carson, K.A.; Phuphanich, S.; Batchelor, T.; Peereboom, D.; Nabors, L.B.; Lesser, G.; Hausheer, F.; Supko, J.G. Phase I and pharmacokinetic study of karenitecin in patients with recurrent malignant gliomas. Neuro-Oncol. 2008, 10, 608–616. [Google Scholar] [CrossRef]
- Abigerges, D.; Chabot, G.G.; Armand, J.-P.; Hérait, P.; Gouyette, A.; Gandia, D. Phase I and pharmacologic studies of the camptothecin analog irinotecan administered every 3 weeks in cancer patients. J. Clin. Oncol. 1995, 13, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Vanhoefer, U.; Harstrick, A.; Achterrath, W.; Cao, S.; Seeber, S.; Rustum, Y.M. Irinotecan in the treatment of colorectal cancer: Clinical overview. J. Clin. Oncol. 2001, 19, 1501–1518. [Google Scholar] [CrossRef] [PubMed]
- Raymond, E.; Fabbro, M.; Boige, V.; Rixe, O.; Frenay, M.; Vassal, G.; Faivre, S.; Sicard, E.; Germa, C.; Rodier, J. Multicentre phase II study and pharmacokinetic analysis of irinotecan in chemotherapy-naive patients with glioblastoma. Ann. Oncol. 2003, 14, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Batchelor, T.T.; Gilbert, M.R.; Supko, J.G.; Carson, K.A.; Nabors, L.B.; Grossman, S.A.; Lesser, G.J.; Mikkelsen, T.; Phuphanich, S.; NABTT CNS Consortium. Phase 2 study of weekly irinotecan in adults with recurrent malignant glioma: Final report of NABTT 97–11. Neuro-Oncol. 2004, 6, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg-Lerner, A.; Bialik, S.; Simon, H.U.; Kimchi, A. Life and death partners: Apoptosis, autophagy and the cross-talk between them. Cell Death Differ. 2009, 16, 966–975. [Google Scholar] [CrossRef]
- Huo, R.; Wang, L.; Liu, P.; Zhao, Y.; Zhang, C.; Bai, B.; Liu, X.; Shi, C.; Wei, S.; Zhang, H. Cabazitaxel-induced autophagy via the PI3K/Akt/mTOR pathway contributes to A549 cell death. Mol. Med. Rep. 2016, 14, 3013–3020. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell. Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Nasiri, J.; Naghavi, M.R.; Motamedi, E.; Alizadeh, H.; Moghadam, M.R.F.; Nabizadeh, M.; Mashouf, A. Carbonaceous sorbents alongside an optimized magnetic solid phase extraction (MSPE) towards enrichment of crude Paclitaxel extracts from callus cultures of Taxus baccata. J. Chromatogr. B 2017, 1043, 96–106. [Google Scholar] [CrossRef]
- Nasiri, J.; Naghavi, M.R.; Alizadeh, H.; Fattahi Moghadam, M.R.; Mashouf, A.; Nabizadeh, M. Modified AHP-based decision-making model toward accurate selection of eligible maintenance media for production of taxanes in Taxus baccata callus culture. Acta. Physiol. Plant 2015, 37, 110. [Google Scholar] [CrossRef]
- Malik, S.; Cusidó, R.M.; Mirjalili, M.H.; Moyano, E.; Palazón, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Process Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Marimpietri, D.; Brignole, C.; Nico, B.; Pastorino, F.; Pezzolo, A.; Piccardi, F.; Cilli, M.; Di Paolo, D.; Pagnan, G.; Longo, L. Combined therapeutic effects of vinblastine and rapamycin on human neuroblastoma growth, apoptosis, and angiogenesis. Clin. Cancer Res. 2007, 13, 3977–3988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcea, G.; Berry, D.P.; Jones, D.J.; Singh, R.; Dennison, A.R.; Farmer, P.B.; Sharma, R.A.; Steward, W.P.; Gescher, A.J. Consumption of the putative chemopreventive agent curcumin by cancer patients: Assessment of curcumin levels in the colorectum and their pharmacodynamic consequences. Cancer Epidemiol. Biomark. Prev. 2005, 14, 120–125. [Google Scholar] [CrossRef]
- Kim, H.G.; Lee, J.H.; Lee, S.J.; Oh, J.-H.; Shin, E.; Jang, Y.P.; Lee, Y.-J. The increased cellular uptake and biliary excretion of curcumin by quercetin: A possible role of albumin binding interaction. Drug Metabo. Dispos. 2012, 40, 1452–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Kulkarni, K.; Zhu, W.; Hu, M. Bioavailability and pharmacokinetics of genistein: Mechanistic studies on its ADME. Anti-Cancer Agents Med. Chem. 2012, 12, 1264–1280. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Ferrucci, L.; Bartali, B.; Urpí-Sarda, M.; Zamora-Ros, R.; Sun, K.; Cherubini, A.; Bandinelli, S.; Andres-Lacueva, C. Resveratrol levels and all-cause mortality in older community-dwelling adults. JAMA Intern. Med. 2014, 174, 1077–1084. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Sang, S. Metabolism and pharmacokinetics of resveratrol and pterostilbene. BioFactors 2018, 44, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. NY Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Venditto, V.J.; Simanek, E.E. Cancer therapies utilizing the camptothecins: A review of the in vivo literature. Mol. Pharm. 2010, 7, 307–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, E.; Mehrpour, M.; Botti, J.; Dupont, N.; Hamai, A.; Nascimbeni, A.C.; Codogno, P. Autophagy: A druggable process. Annu. Rev. Pharmacol. 2017, 57, 375–398. [Google Scholar] [CrossRef] [PubMed]






 : Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right VD24OB6Q8E, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right VD24OB6Q8E, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right VD24OB6Q8E, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right VD24OB6Q8E, 21 November 2022.

 : Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right AI24OB6U1K, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right AI24OB6U1K, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right AI24OB6U1K, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right AI24OB6U1K, 21 November 2022.

 : Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right OG24OB731X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right OG24OB731X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right OG24OB731X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right OG24OB731X, 21 November 2022.
 : Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right IP24OB76U3, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right IP24OB76U3, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right IP24OB76U3, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. − and +: Indicate the effect of the proteins on different pathways (− means negative effect and + means positive effect). The figure was created with BioRender.com. Licensing Right IP24OB76U3, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right RD24OB7C3X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right RD24OB7C3X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right RD24OB7C3X, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right RD24OB7C3X, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PC24OB7FPB, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PC24OB7FPB, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PC24OB7FPB, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PC24OB7FPB, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right MV24OB7IC0, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right MV24OB7IC0, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right MV24OB7IC0, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right MV24OB7IC0, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right VE24OB7LHH, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right VE24OB7LHH, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right VE24OB7LHH, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right VE24OB7LHH, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PA24OB7OP8, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PA24OB7OP8, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PA24OB7OP8, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right PA24OB7OP8, 21 November 2022. : Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right IB24OB7ST2, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right IB24OB7ST2, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right IB24OB7ST2, 21 November 2022.
: Indicates deactivation or a decrease in the mentioned protein expression. +: means positive effect of the proteins on different pathways. The figure was created with BioRender.com. Licensing Right IB24OB7ST2, 21 November 2022.


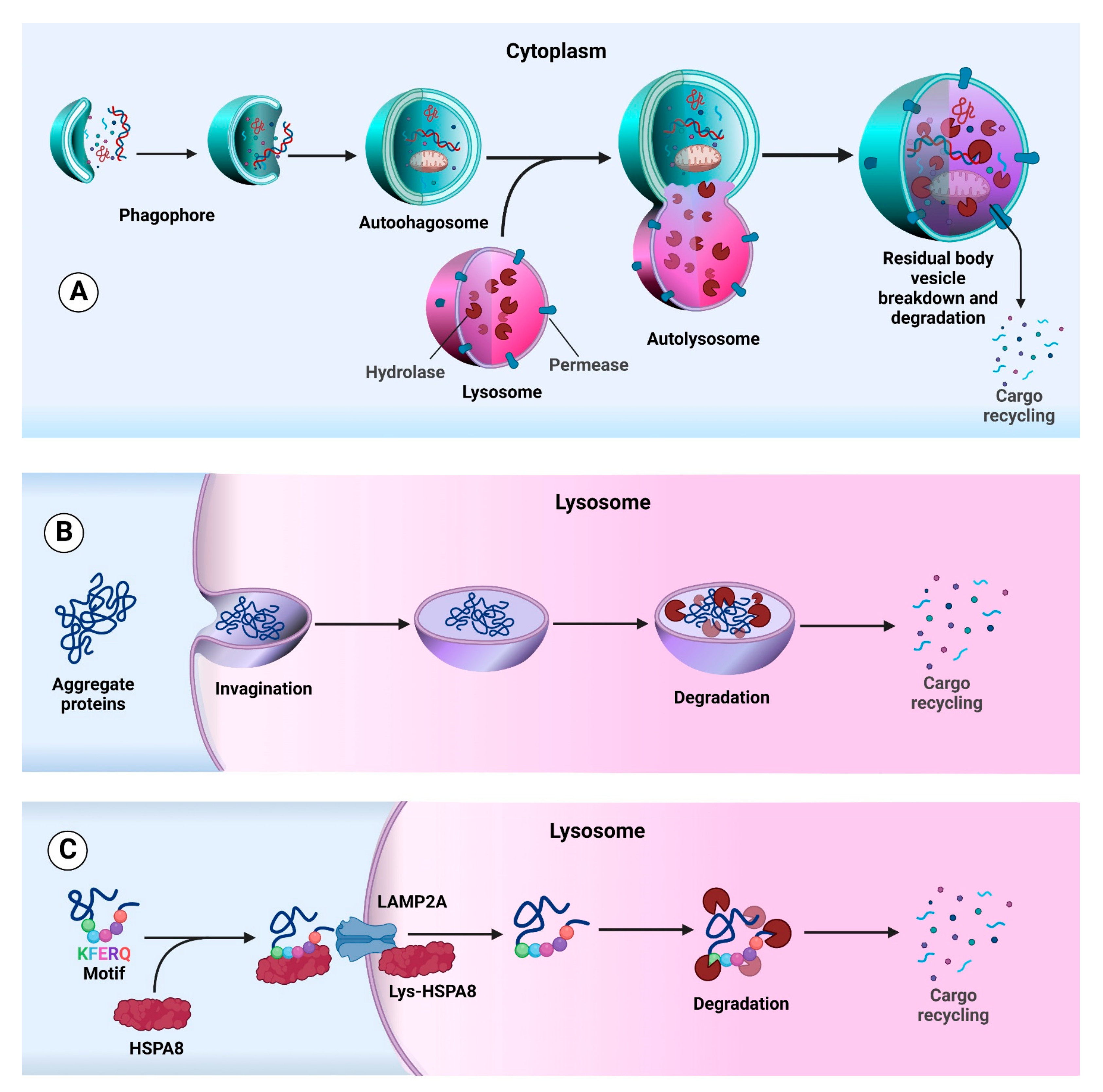{kind=link}
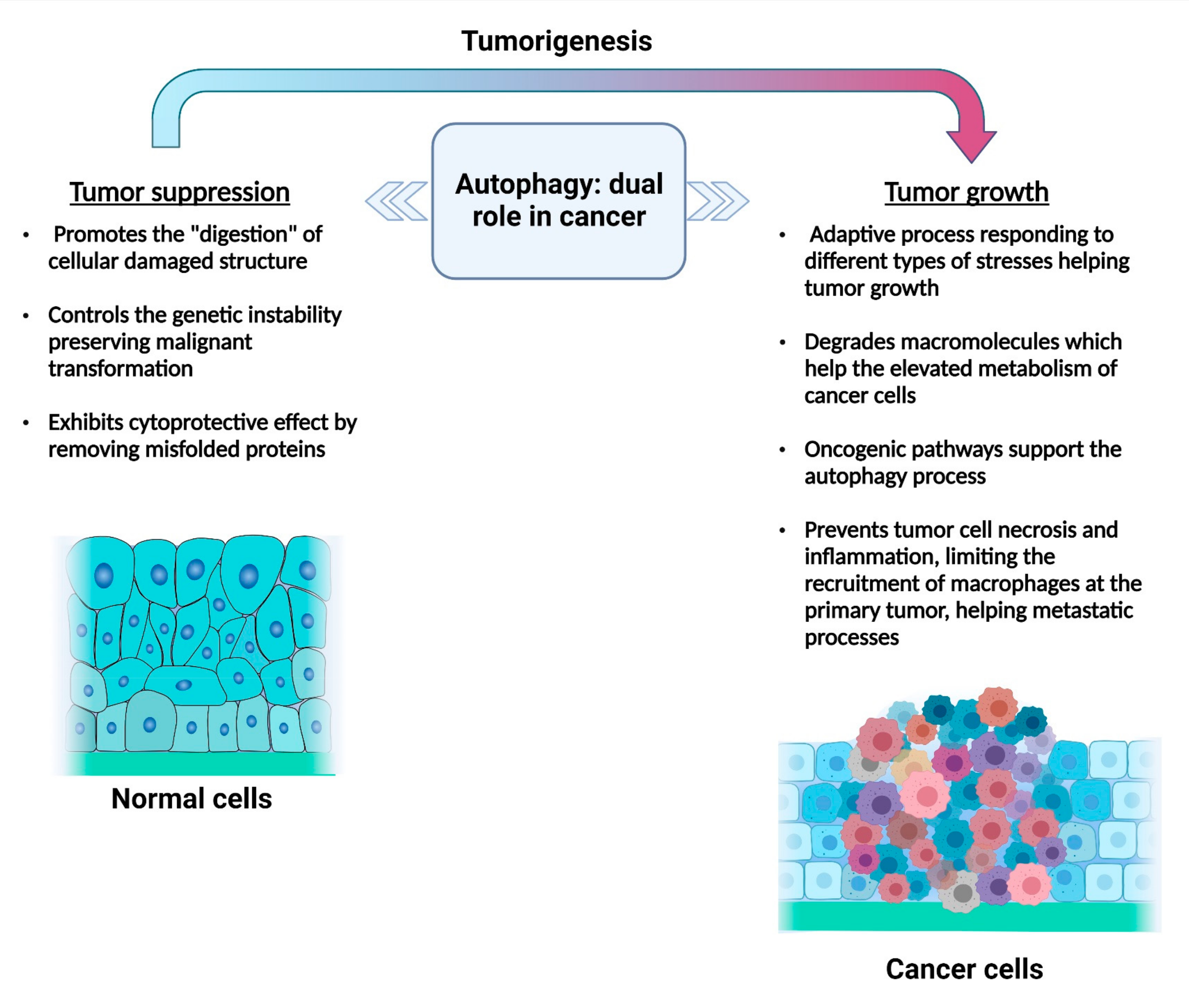{kind=link}
{kind=link}
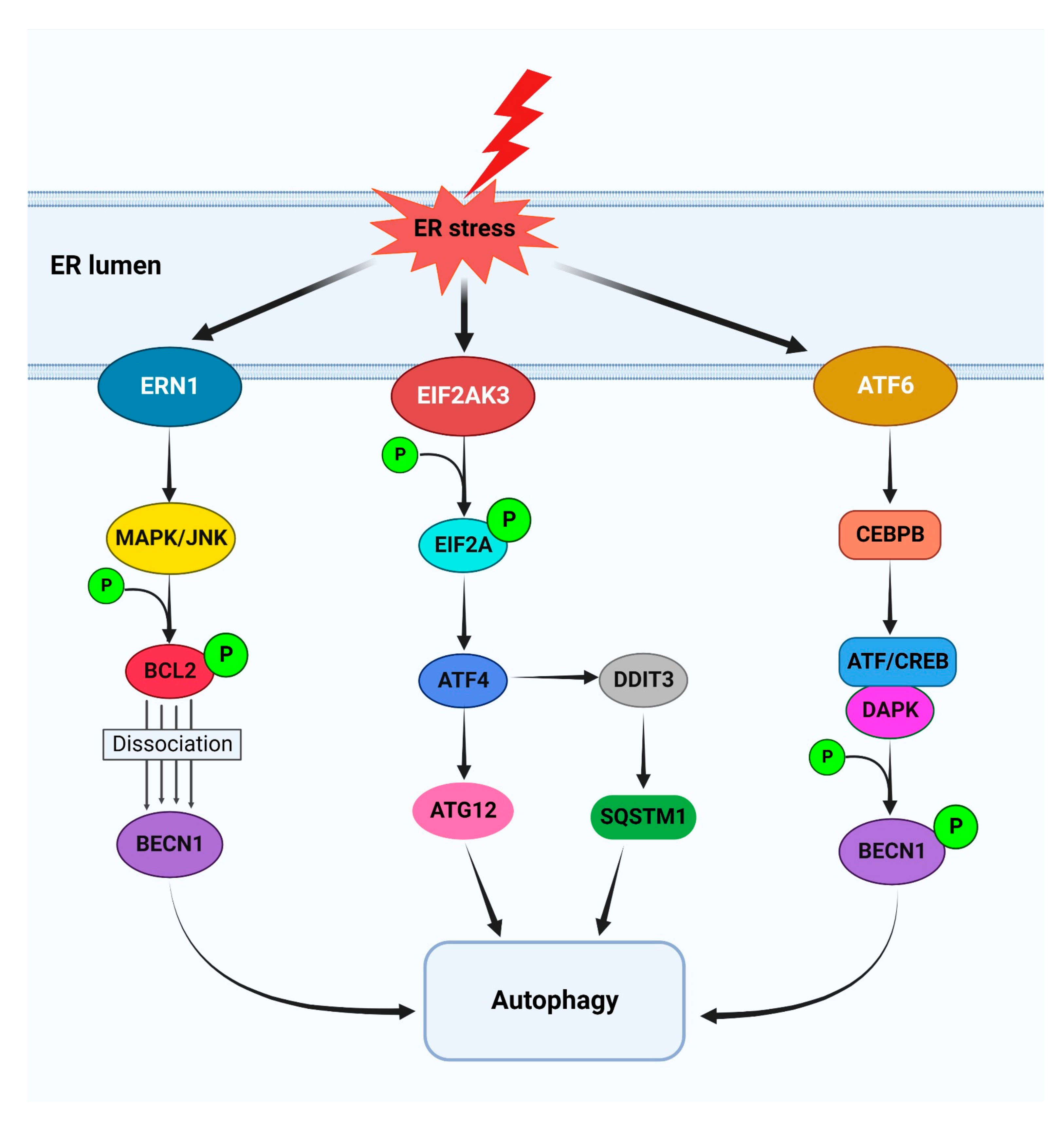{kind=link}
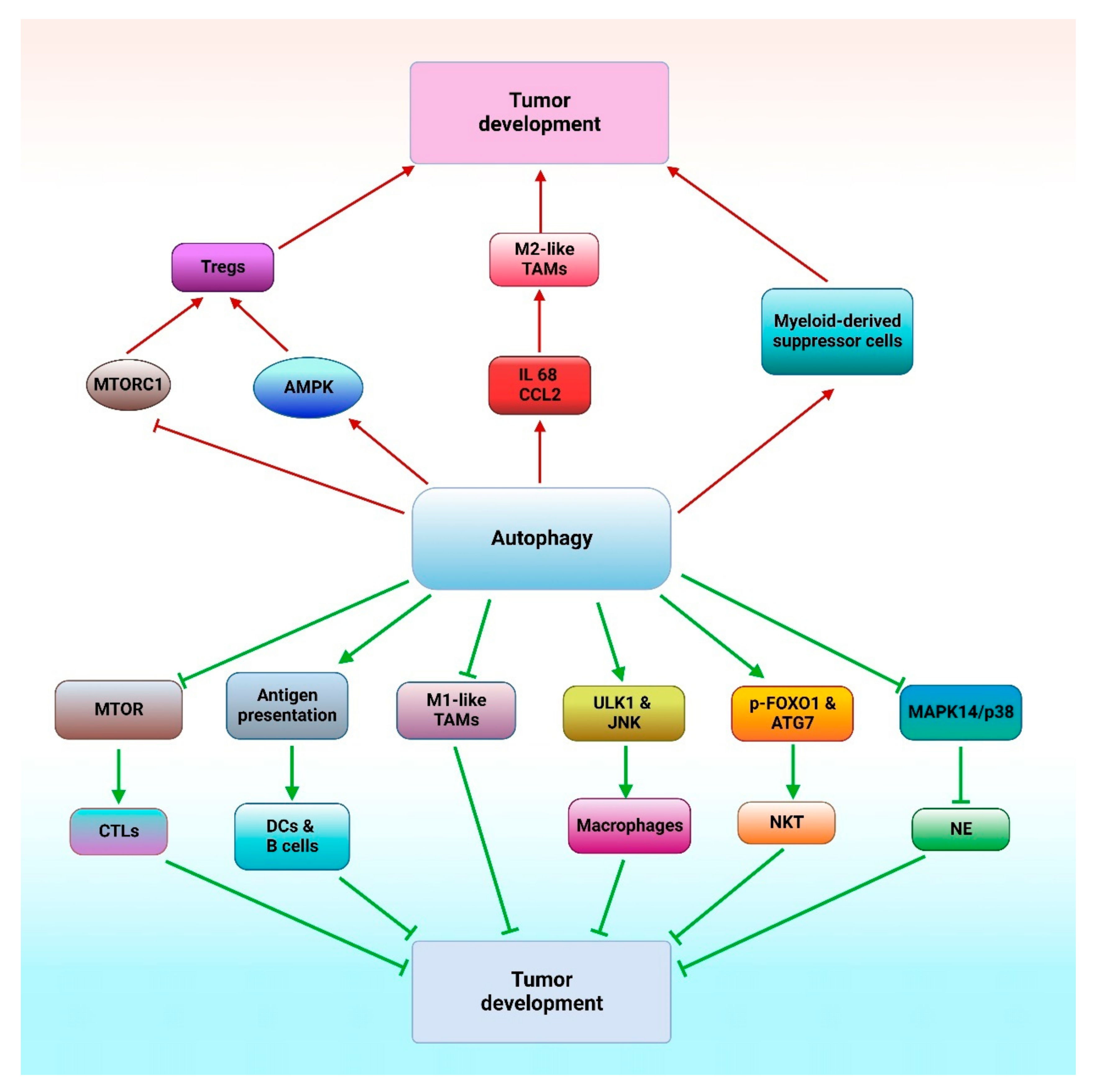{kind=link}
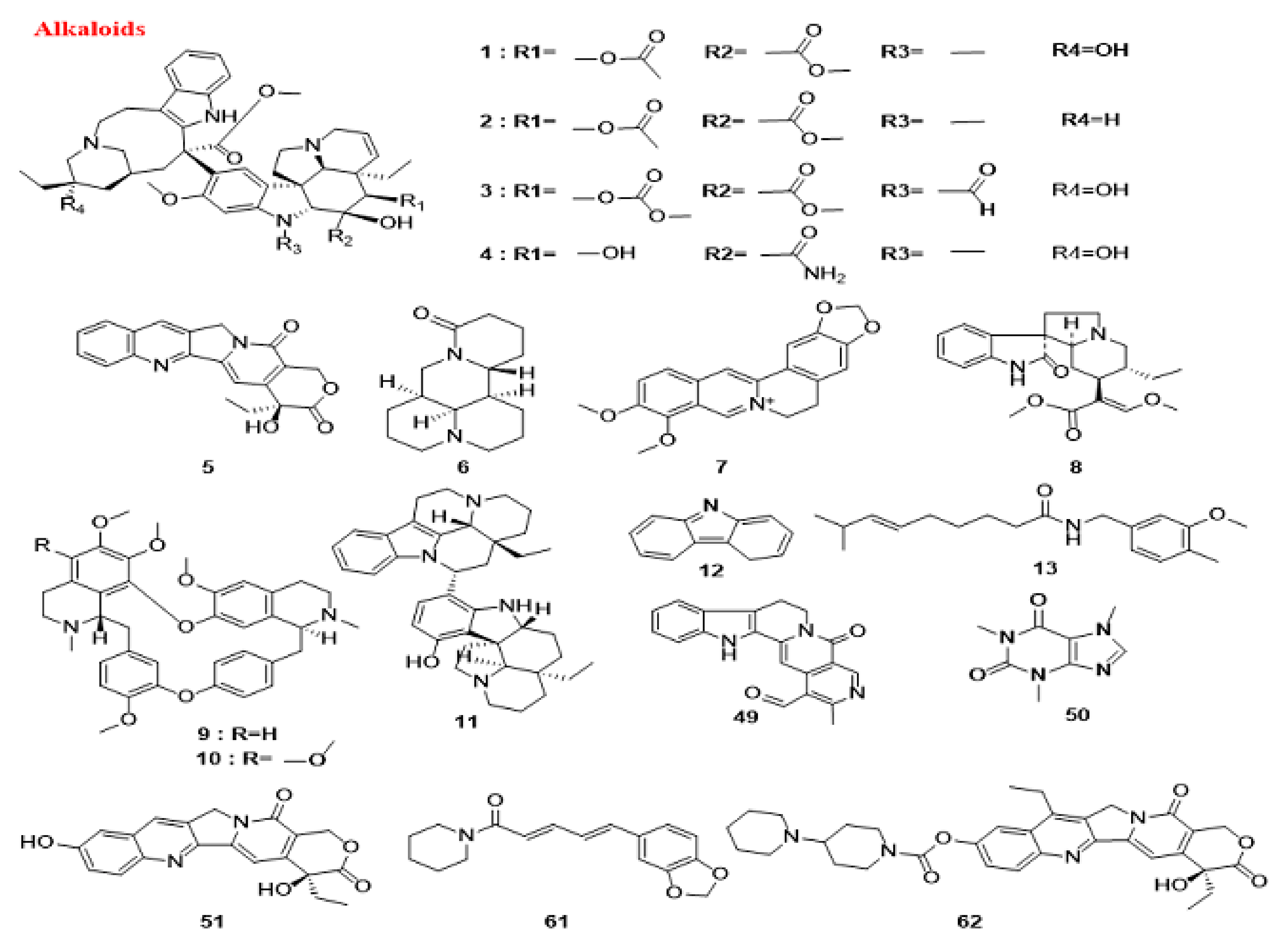{kind=link}
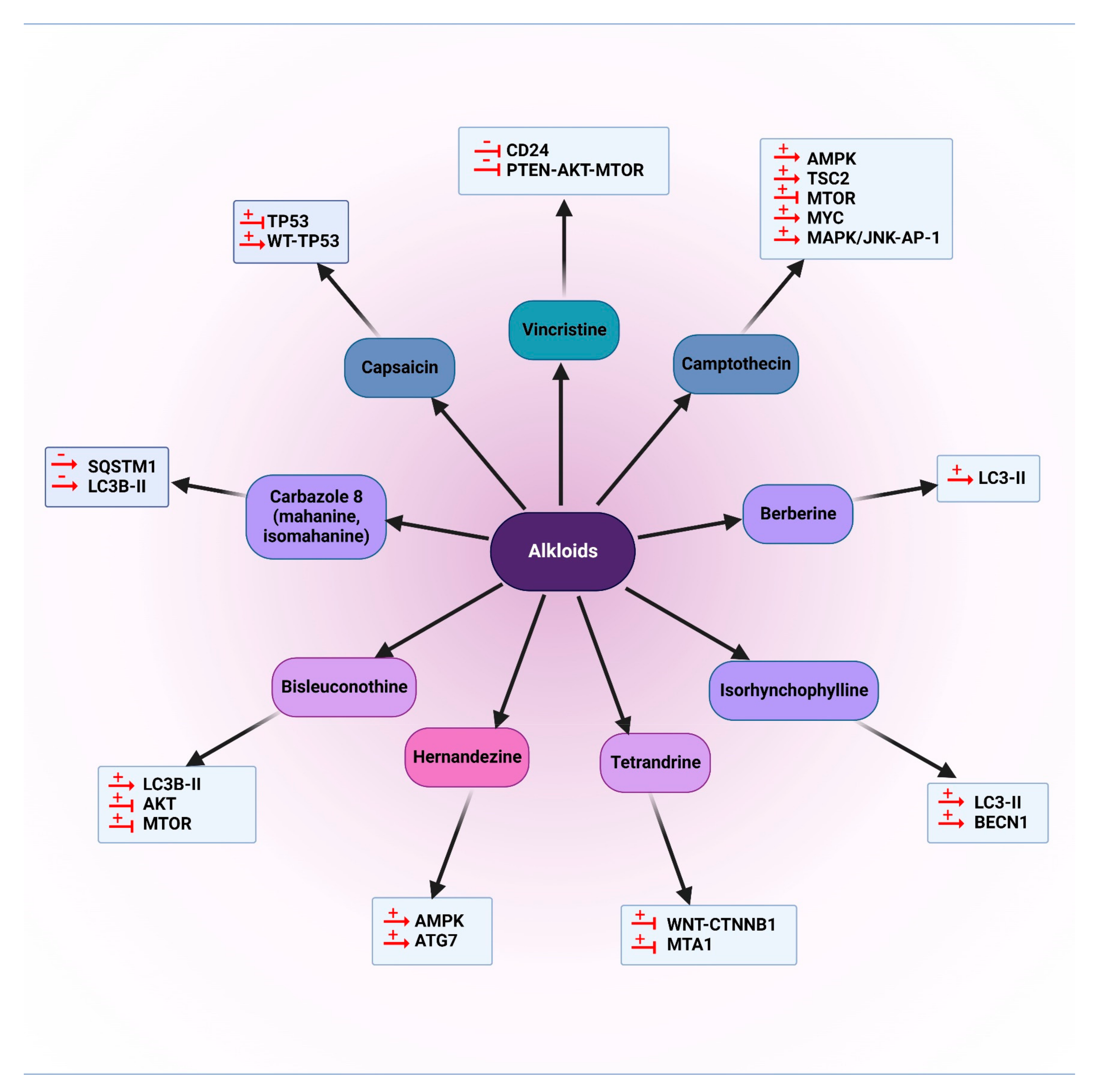{kind=link}
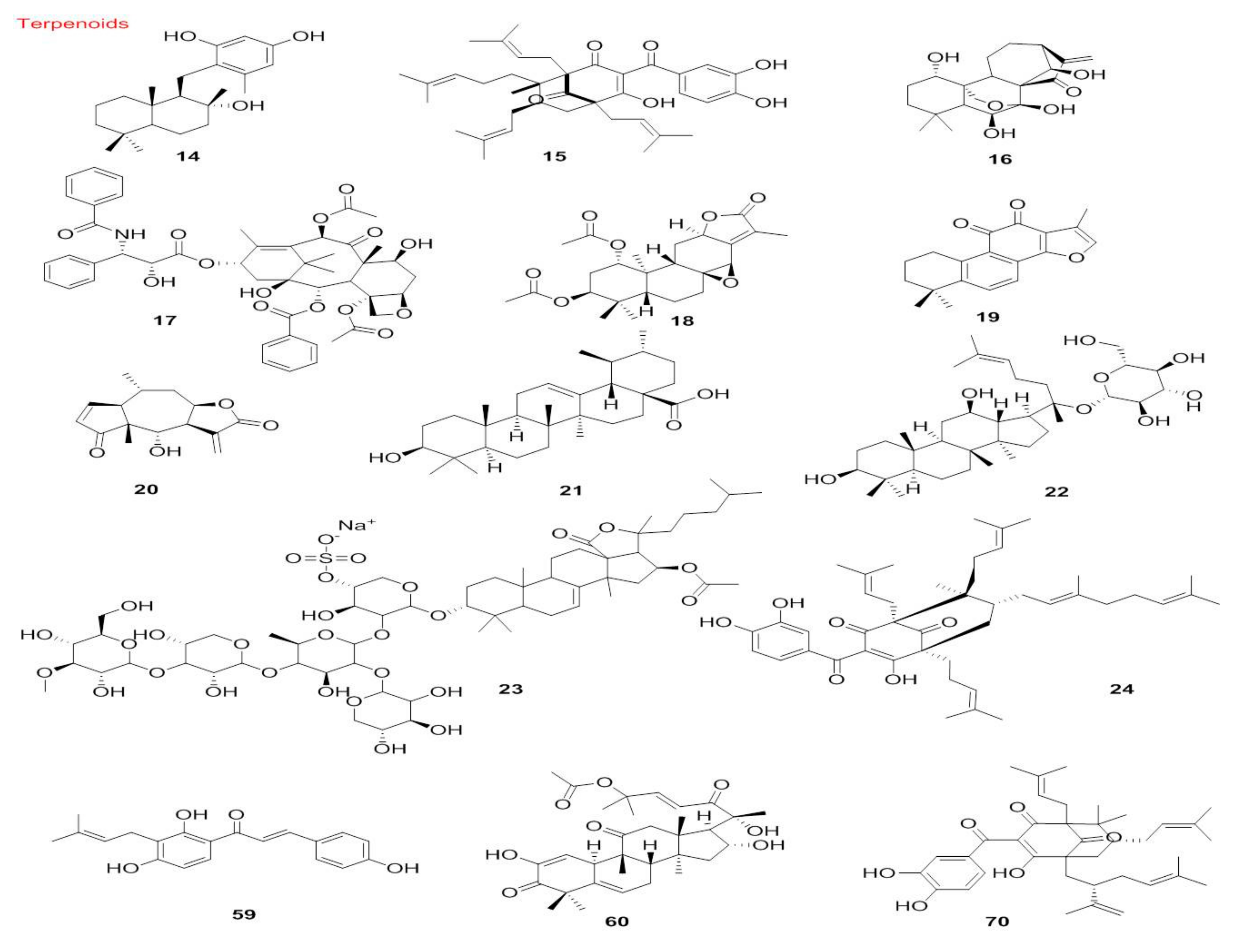{kind=link}
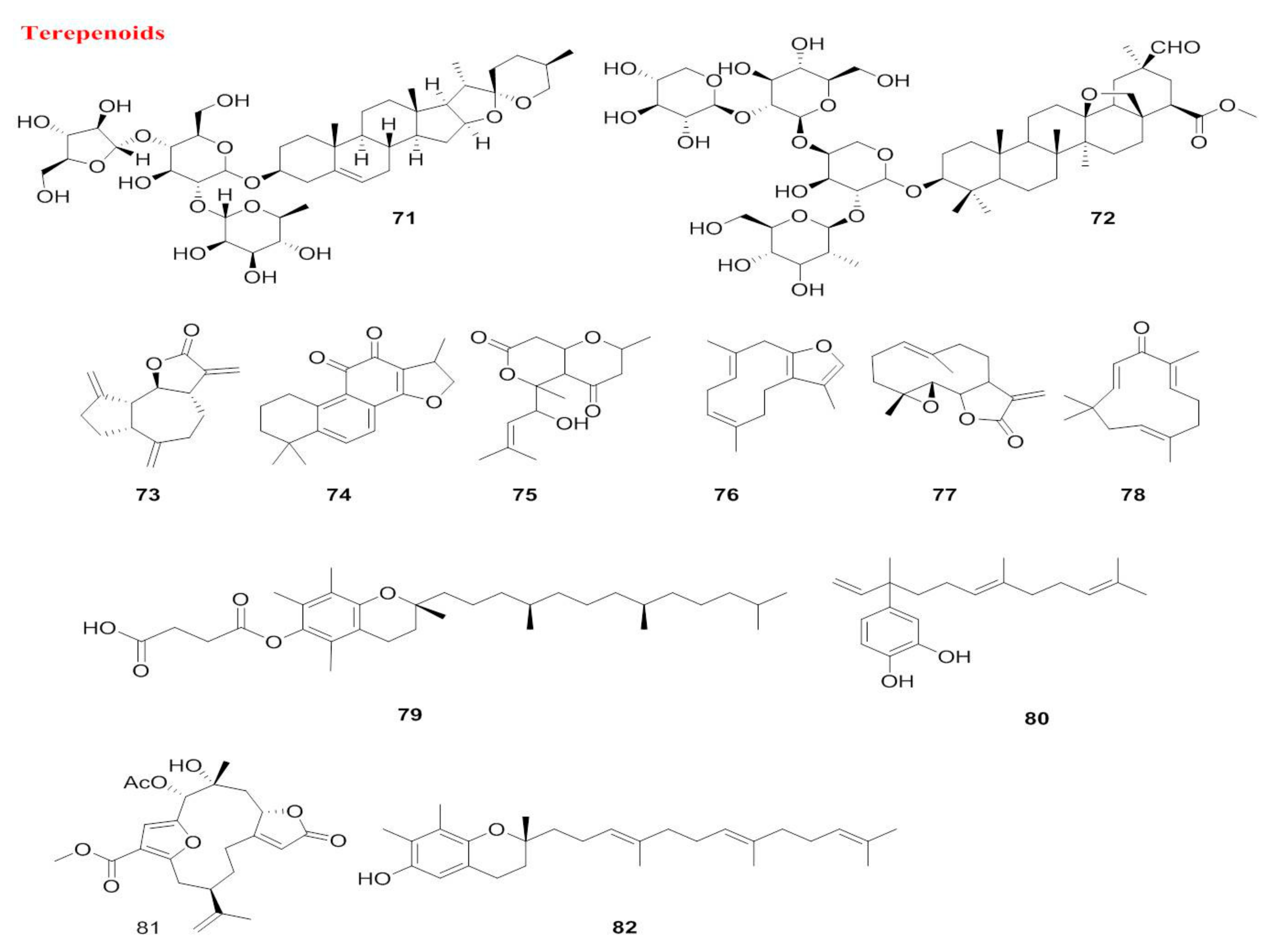{kind=link}
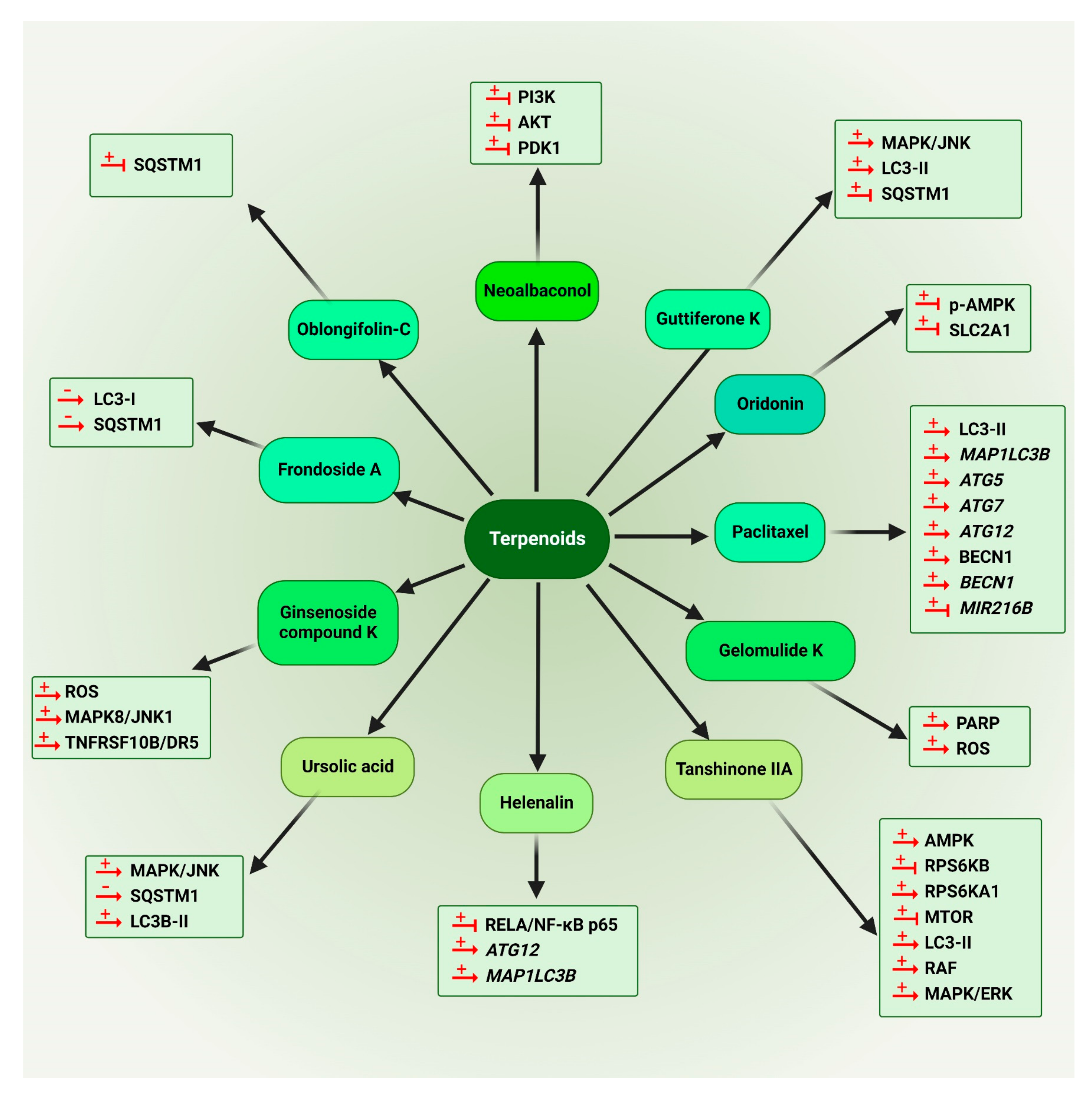{kind=link}
{kind=link}
{kind=link}
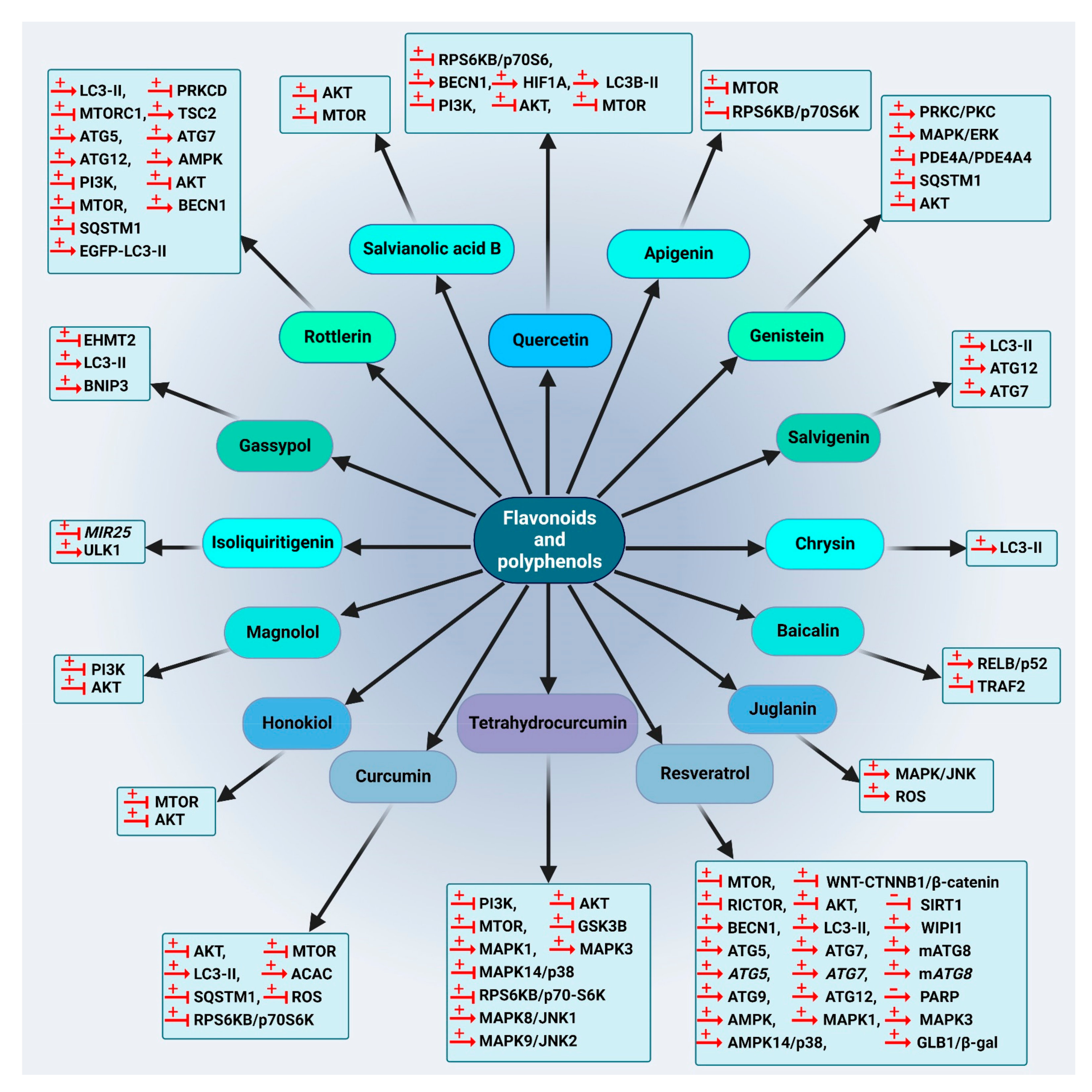{kind=link}
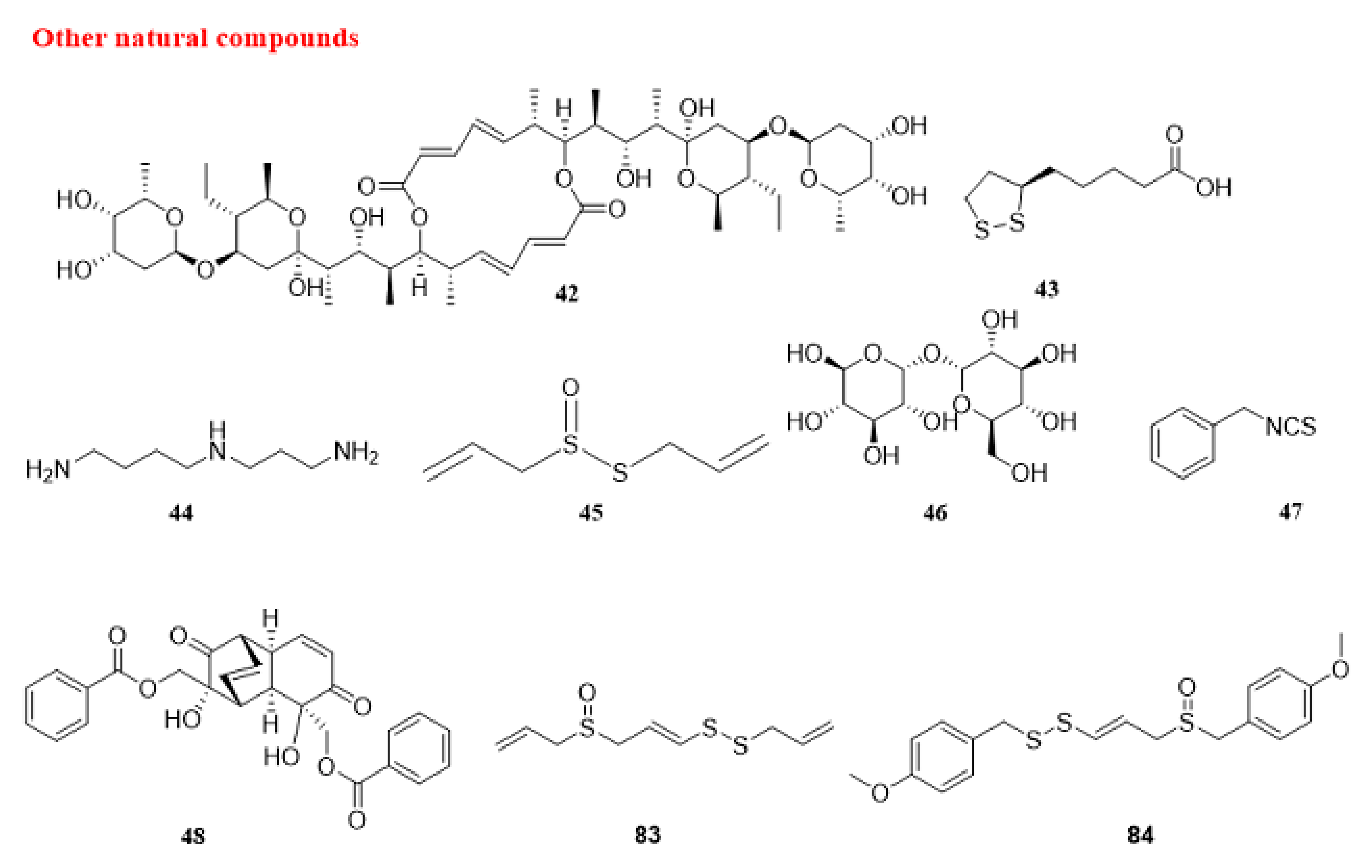{kind=link}
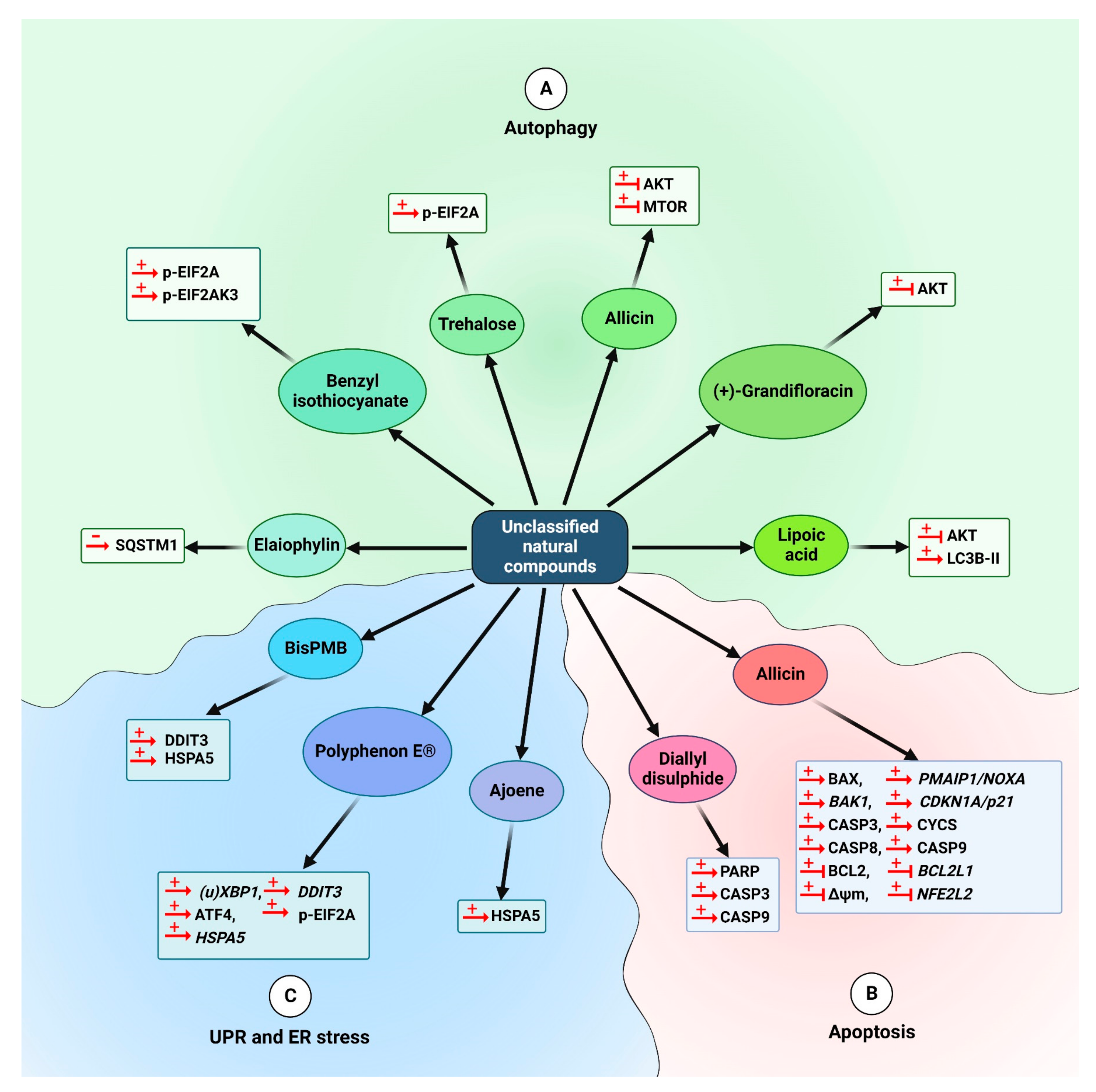{kind=link}
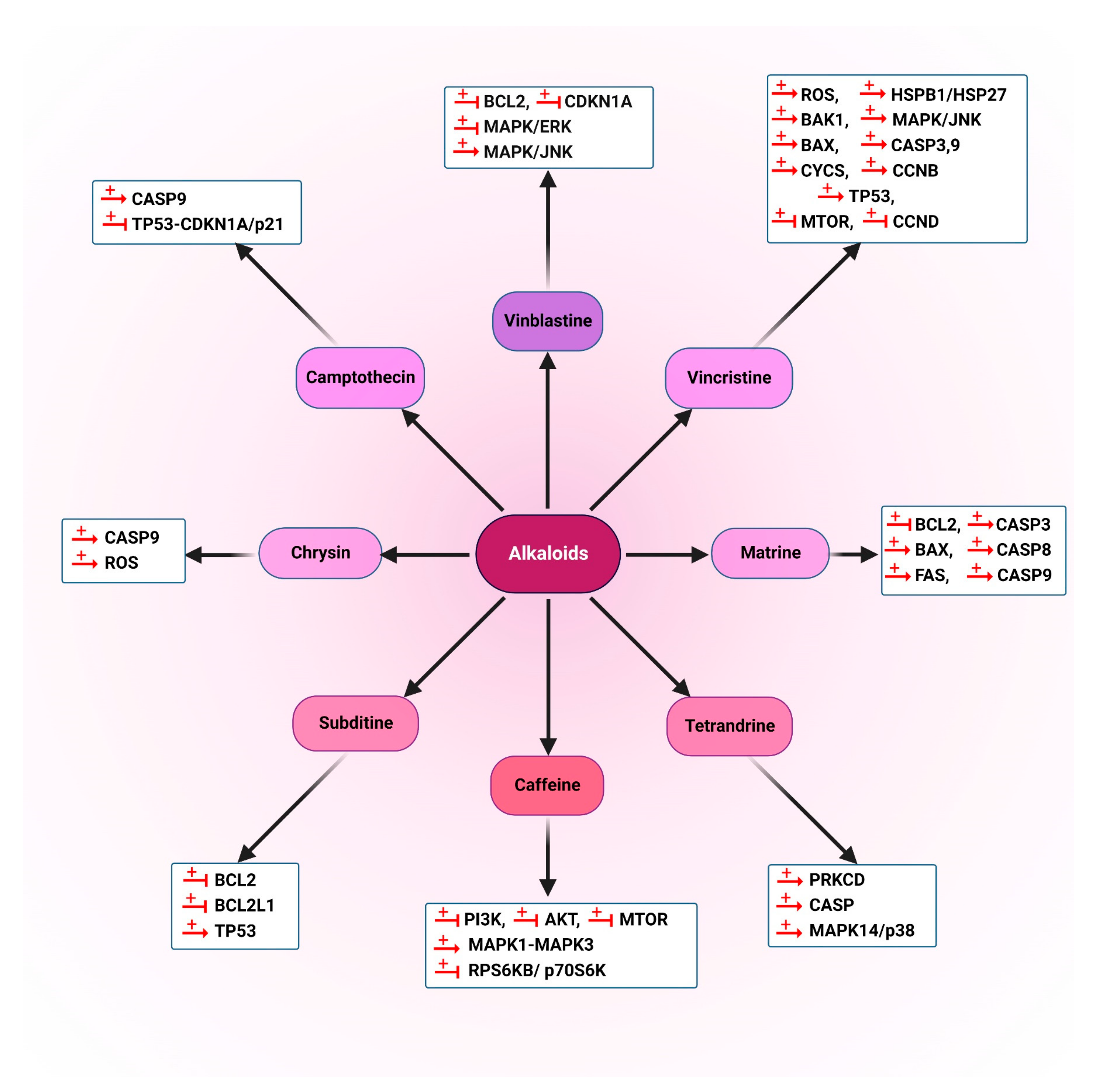{kind=link}
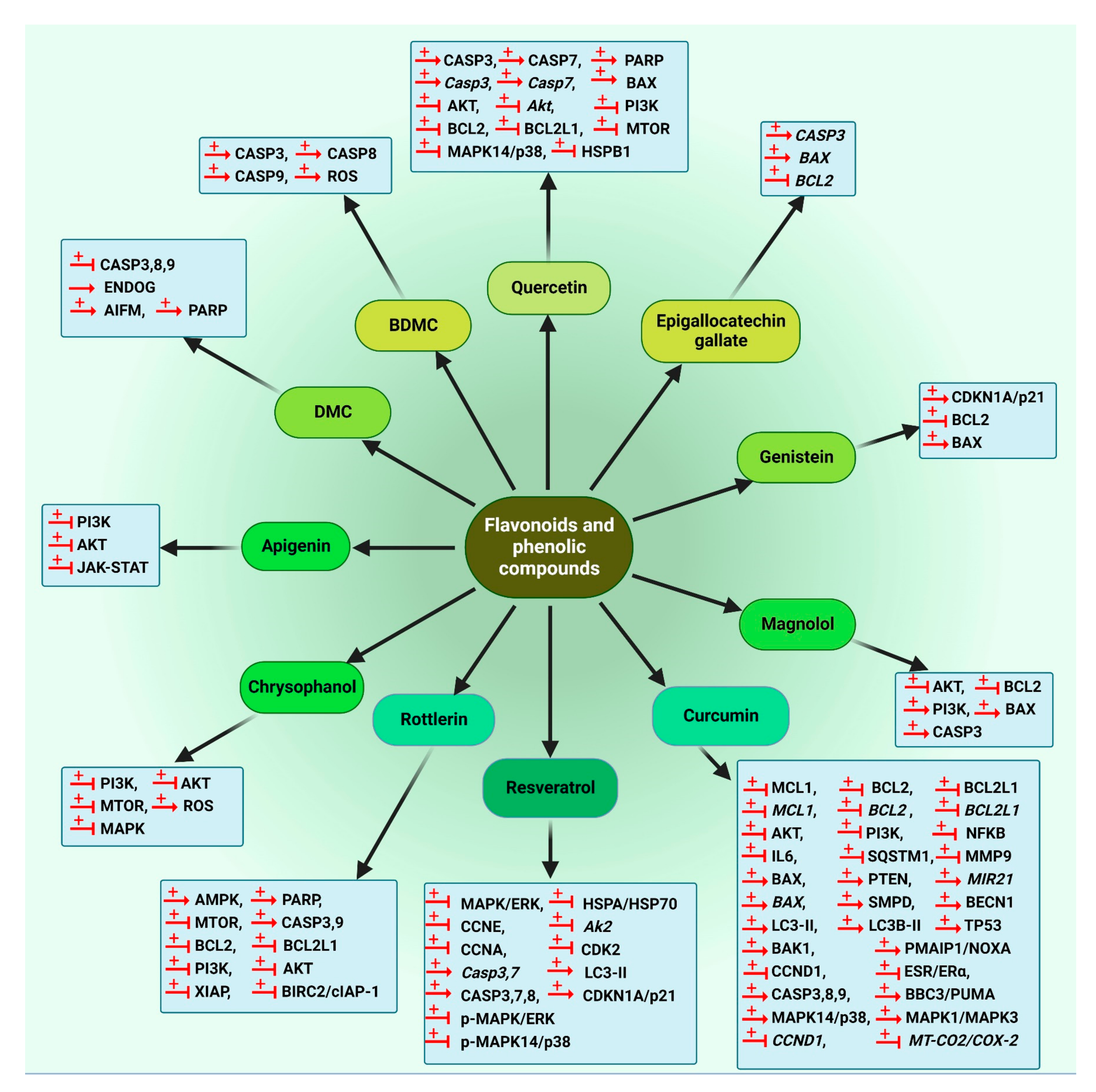{kind=link}
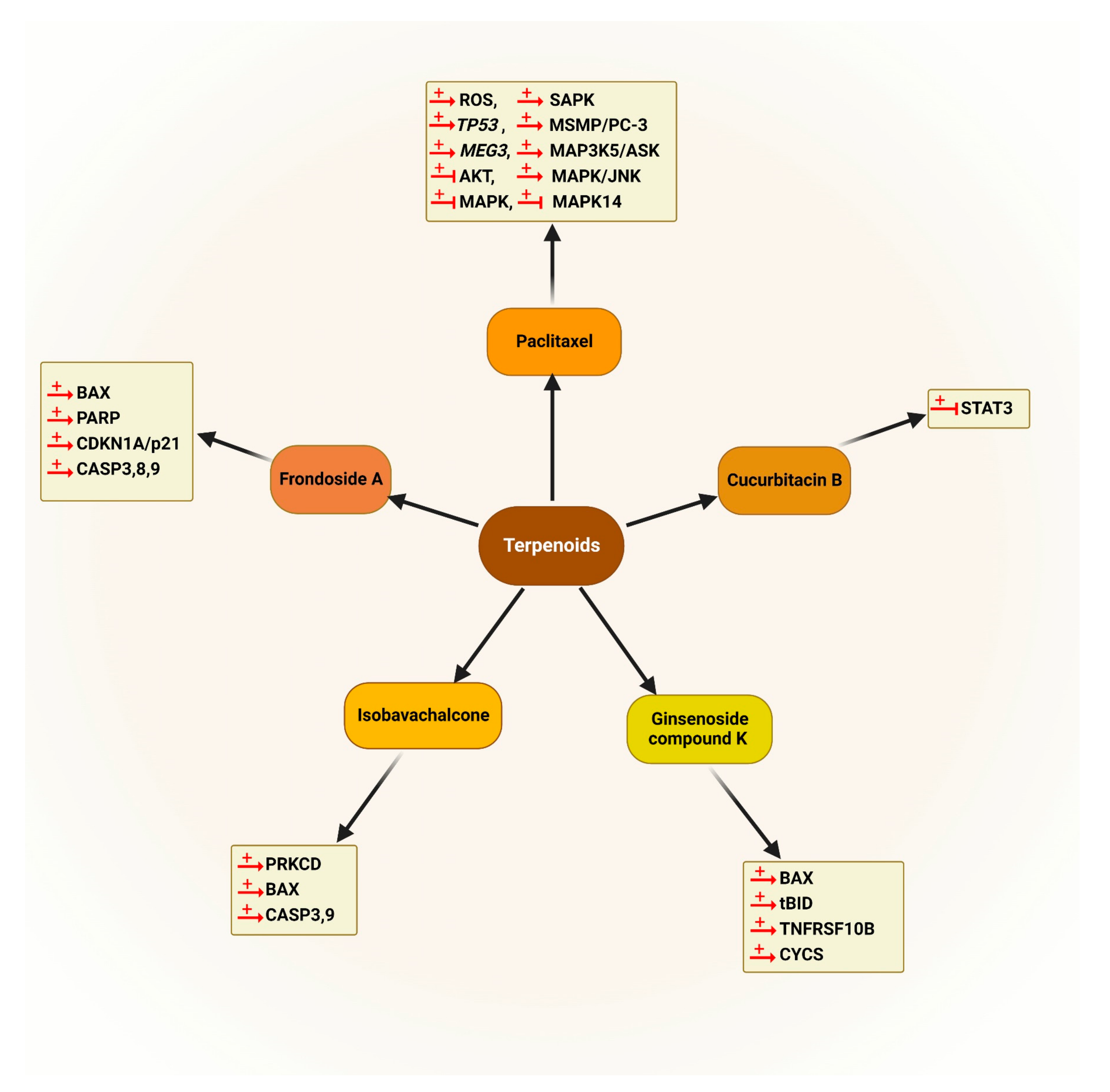{kind=link}
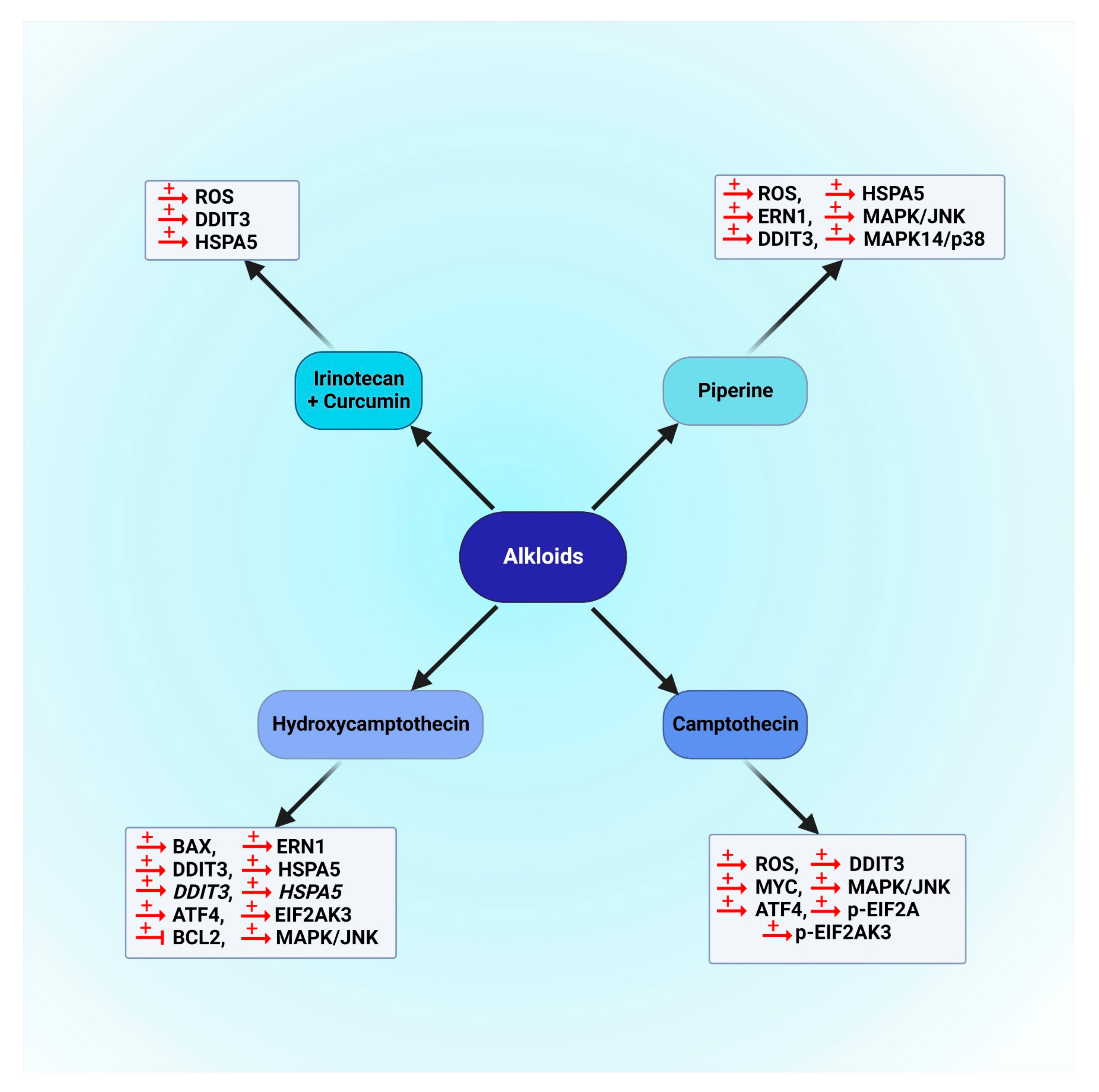{kind=link}
{kind=link}
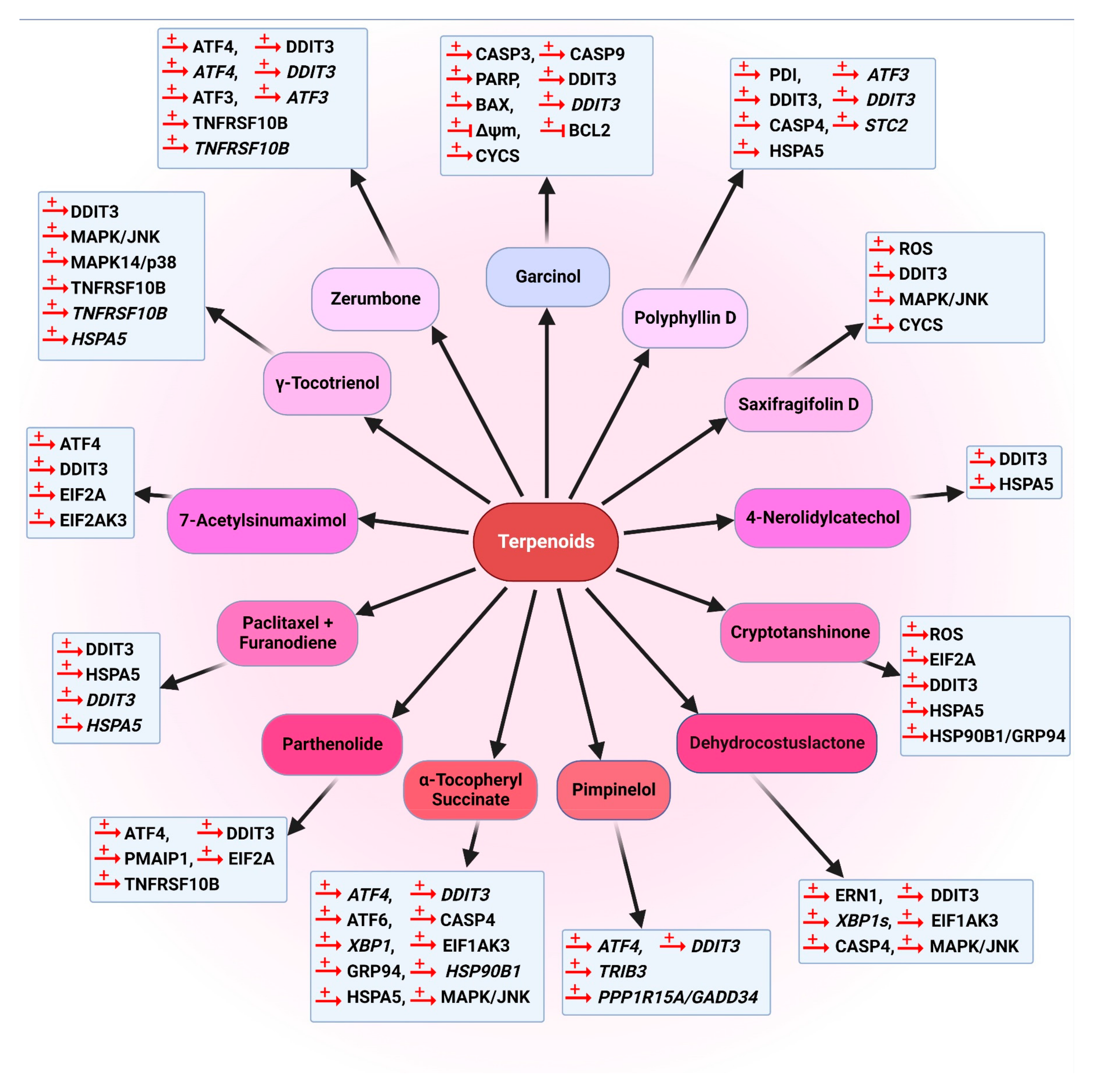{kind=link}
{kind=link}
| Cancer Type | Compound | Cell Line | Target | Reference |
|---|---|---|---|---|
| Human colon cancer | Synthesized derivatives of berberine (7) | HCT116, SW613-B3 | ↑ LC3-II | [149] |
| Oridonin (16) | SW480 | ↑ p-AMPK, ↓ SLC2A1/GLUT1 | [161] | |
| Vinblastine (1) | LS174T | ↑ LC3-II, ↑ autophagic vacuole, | [134] | |
| Camptothecin (5) | HCT116, RKO | AMPK-TSC2-MTOR | [144] | |
| B-group soyasaponins | HCT-15 | ↑LC3-II, ↑ autophagic vacuoles | [157] | |
| Helenalin (20) | RKO | ↓ RELA, ↑ LC3B, ATG12 and caspases | [170] | |
| Ursolic acid (21) | HCT15 | MAPK/JNK, ↑ LC3, SQSTM1 | [172] | |
| Ginsenoside compound K (22) | HCT15 | ROS-MAPK/JNK, ↑ TNFRSF10A | [173] | |
| Quercetin (25) | DLD-1, HT-29 | ↑ EGFP-LC3 | [180] | |
| Curcumin (33) | HCT116 | ↑ ROS-MAPK1-MAPK3-MAPK14/p38 | [220] | |
| ↓ AKT-MTOR, ↑TFEB | [226] | |||
| Bis-dehydroxycurcumin (36) | HCT116 | ER stress | [231] | |
| Salvianolic acid B (40) | HCT116, HT-29 | ↓ AKT-MTOR | [245] | |
| Lipoic acid (43) | HCT116 | ↓ MGMT, AKT ↑ LC3-II | [248] | |
| Resveratrol (32) and spermidine (44) | HCT116 | AMPK-MTOR independent, Change phosphoproteome and acetylproteome | [250] | |
| Neuroblastoma | Salvigenin (28) | SH-SY5Y | ↑ LC3-II, ATG7, ATG12 | [191] |
| Hepatocellular carcinoma | Tetrandrine (9) | Huh7, HCCLM9, Hep3B | WNT-CTNNB1 pathway and ↓ MTA1 | [152] |
| Resveratrol (32) | Huh-7 | ↑ ATG5, ATG7, ATG9, ATG12 | [209] | |
| Baicalin (30) | MHCC97L | ↑ RELB/p52, ↓ TRAF2 | [194] | |
| Thyroid cancer | Allicin (45) | SW1736, HTh-7 | ↓ AKT-MTOR | [253] |
| Oral squamous cell carcinoma | Carbazole alkaloids | CLS-354 | ↑ SQSTM1, LC3B-II, cleaved CASP3 | [155] |
| Breast cancer | Resveratrol (32) | MCF-7, SUM159 | ↓ WNT-CTNNB1 ↑ LC3-II, BECN1, ATG7 | [198] |
| MCF-7 | ↓ AKT-MTOR | [205] | ||
| MCF-7 | WIPI1, ↑ LC3-II, ATG7, ATG5 | [208] | ||
| Bisleuconothine A (11) | MCF-7 | AMPK-MTOR, ↑ LC3-II, | [154] | |
| Helenalin (20) | MCF-7 | ↓ RELA, ↑ LC3B, ATG12 and caspases | [170] | |
| Qauercetin (25) | MCF-7 | Triggering macroautophagy, ↓ MTOR | [182] | |
| Rottlerin (38) | MCF-7 | ↑ TSC2,MTORC1, ↑ EGFP-LC3-II, EGFP | [238] | |
| ↑ LC3-II, ↓ SQSTM1 | [239] | |||
| MTORC1 inhibition | [241] | |||
| ↑ LC3, BECN1, ATG12, AMPK | [240] | |||
| Isoliquiritigenin (41) | MCF-7 | ↓ MIR25, ↑ ULK1 | [246] | |
| Juglanin (31) | MCF-7, SKBR3, | ↑ MAPK/JNK, ROS | [195] | |
| chrysin (29) | MCF-7 | ↑ LC3-II | [192] | |
| Lung cancer | Hernandezine (10) | A549, H1299 | ↑ AMPK, ↑ ATG7 | [153] |
| Curcumin (33) | A549 | ↑ LC3-II, p-AMPK, p-ACAC ↓ SQSTM1, AMPK | [222] | |
| Resveratrol (32) | HFL1 | SIRT1, PARP | [213] | |
| Bisleuconothine A (11) | A549 | AKT-MTOR, ↑LC3-II | [154] | |
| Camptothecin (5) | H1299 | ↑ Autophagosome formation | [146] | |
| Paclitaxel (17) | A549, Calu-3 | ↑ BECN1, ↓ MIR216B | [166] | |
| Capsaicin (13) | H1299 | ↑ WT TP53 | [156] | |
| Benzyl isothiocyanate (47) | A549 | ↑ EIF2AK3, EIF2A, LC3-II-ATG5, AVOs | [255]. | |
| Osteosarcoma | Curcumin (33) | MG63 | ↑ MAPK/JNK | [171] |
| Human fibrosarcoma | Rottlerin (38) | HT1080 | ↑ LC3-II | [236] |
| Gastric cancer (GC) | Matrine (6) | SGC-7901 | blocking autophagic degradation | [147] |
| Quercetin (25) | AGS, BGC-823, SGC-7901, MKN-28 | PI3K-AKT-MTOR, HIF1A | [181] | |
| Curcumin (33) | SGC-7901, BGC-823 | ↓ PI3K-AKT-MTOR ↑ TP53, CDKN1A/p21 | [230] | |
| MKN-28 | PI3K-AKT-MTOR | [255] | ||
| Magnolol (35) | SGC-7901 | ↓ PI3K-AKT | [234] | |
| Uterine leiomyosarcoma | Curcumin (33) | SKN, SK-UT-1 | ↑ MAPK1, MAPK3 | [223] |
| Pancreatic cancer | Curcumin (33) | PANC-1, BxPC3 | ↑ LC3-II, ↓ MTOR | [229] |
| (+)-Grandifloracin (48) | PANC-1 | ↓ AKT | [256] | |
| Rottlerin (38) | MDA-Panc28 | ↓ PRKCD, TGM2 | [237] | |
| Gossypol (39) | PANC-1 | ↑ LC3-II, BNIP3 ↓ EHMT2 | [244] | |
| Nasopharyngeal carcinoma | Neoalbaconol (14) | C666, HK1, CNE1 | PI3K-AKT | [158] |
| prostate cancer | Resveratrol (32) | PC-3, DU145 | ↑ AMPK ↓ STIM1, AKT, MTOR | [201] |
| Camptothecin (5) | LNCaP | ↑ EIF2AK3, EIF2A, ATF4, DDIT3, MYC, ROS, MAPK/JNK | [145] | |
| Rottlerin (38) | Cancer stem cells | ↓ PI3K-AKT-MTOR ↑ LC3-II, ATG5, ATG7, ATG12, BECN1 | [242] | |
| leukemia | Tanshinone IIA (19) | KBM-5 | ↑ AMPK, LC3-II, MAPK/ERK, RAF, RPS6KA1/p90RSK ↓ MTOR, RPS6KB/p70S6K | [169] |
| Resveratrol (32) | K562 | MAPK/JNK ↑ SQSTM1, AMPK | [204] | |
| Tetrahydrocurcumin (37) | HL-60 | ↓ PI3K-AKT-MTOR-RPS6KB/p70S6K, GSK3B-MAPK14/p38, ↑ MAPK1-MAPK3,MAPK8/JNK1-MAPK9/JNK2 | [232] | |
| Apigenin (26) | TF1 | ↓ MTOR, RPS6KB/p70S6K, JAK-STAT | [187] | |
| Cervical cancer | Guttiferone K (15) | Hela | ↑ MAPK/JNK, LC3-II ↓ AKT-MTOR, SQSTM1 | [159] |
| Paclitaxel (17) | SiHa | ↑ LC3-II, ATG7 | [167] | |
| HeLa-R | ↑ ATG genes (ATG7, ATG12–ATG5), BECN1, MAP1LC3B, HIF1A | [165] | ||
| Resveratrol (32) | SiHa, Hela | ↑ LC3-II, autophagosomes | [214] | |
| Hela | ↑ Lysosomal permeability | [199] | ||
| Oral squamous cell carcinoma | Curcumin (33) | YD10B | ↑ LC3-II, AVOs | [221] |
| Glioma | Resveratrol (32) | U373 | ↑ MAPK14/p38-MAPK1-MAPK3 | [203] |
| U-87, U-251, U-138 | ↑ LC3-II, ATG5, BECN1 | [211] | ||
| Curcumin (33) | U87-MG, U373-MG | ↓ AKT-MTOR-RPS6KB/p70S6K, ↑ MAPK1-MAPK3 | [219] | |
| Ovarian | Helenalin (20) | A2780 | ↓ RELA, ↑ LC3B, ATG12 and caspases | [170] |
| Quercetin (25) | OVCAR3 | ↑ BECN1, LC3B-II | [183] | |
| Resveratrol (32) | A2780, CaOV3 | ↑ BCL2L1, BCL2 | [197] | |
| SKOV3, CaOV3 | ↓ AKT-MTOR | [202] | ||
| Genistein (27) | A2780, CaOV3, ES2 | ↓ p-AKT | [189] | |
| CHO | ↓ PDE4A4-SQSTM1 | [190] | ||
| Elaiophylin (42) | SKOV3, A2780 | ↑ SQSTM1 | [247] | |
| Epidermoid carcinoma | Resveratrol (32) | A431 | ↓ RICTOR, RHOA GTP ↑ GLB1/β-gal | [206] |
| Urothelial carcinoma | Frondoside A (23) | RT112 | ↑ LC3-I/II, SQSTM1 | [175] |
| Skin cancer (melanoma) | Resveratrol (32) | B16 | ↑ Ceramide, ↓ AKT-MTOR pathway | [200] |
| Honokiol (34) | B16-F10 | ↓ NOTCH1, AKT-MTOR | [233] | |
| Human melanoma | Curcumin (33) | A375,C8161 | ↓ AKT-MTOR | [228] |
| Cancer Type | Compound | Cell Line | Target | Reference |
|---|---|---|---|---|
| Pancreatic cancer | Matrine (6) | BxPC-3, PANC-1 | ↑ BCL2-BAX, FAS, CASP3, CASP8, CASP9 | [266] |
| Rottrelin (38) | stem cells (CD44+ CD24+ ESA+) | PI3K-AKT-MTOR inhibition | [317] | |
| Cervical cancer | Allicin (45) | SiHa | ↑ CASP3, CASP8, CASP9 | [334] |
| Leukemia | Tetrandrine (9) | U937 | ↑ Caspases, PRKCD | [151] |
| Diallyl disulfide | HL-60 | ↑ CASP3, ↓ PARP | [340] | |
| Murine colon cancer | Tetrandrine (9) | CT-26 | ↑ MAPK14/p38 | [267] |
| Human prostate cancer | Subditine (49) | LNCaP, PC-3 | ↓ BCL2-BCL2L1 | [268] |
| Curcumin (33) | LNCaP | NFKB, NFKBIA/IκBα | [302] | |
| PC-3 | ↓ p-AKT and NFKB | [303] | ||
| PC-3 | ↓ BCL2, BCL2L1 ↑ BAX, BAK1, BBC3/PUMA, PMAIP1/NOXA, CASP3, CASP9 | [291] | ||
| Quercetin (25) | DU-145, LNCaP | ↓ AKT, ↑ Activation | [275] | |
| LNCaP | ↓ BCL2L1 ↑ BAX, caspases | [277] | ||
| Resveratrol (32) | DU-145 | ↑ CASP3 activity by HSPA8/HSP70 involvement | [316] | |
| Rottlerin (38) | Cancer stem cells | ↓ PI3K-AKT-MTOR ↑ AMPK | [242] | |
| Caffeine (50) | PC12D, Hela, SH-SY5Y | ↓ PI3K-AKT-MTOR-RPS6KB/p70S6K ↑ MAPK1-MAPK3 | [270] | |
| Human lung cancer | Camptothecin (5) | H1299, H460 | ↑ CASP9 | [146] |
| Bisdemethoxycurcumin (BDMC) (56) | NCI-H460 | ↑ CASP3, CASP8 and CASP9 | [310] | |
| Demethoxycurcumin (DMC) (55) | NCI-H460 | ↑ CASP3, CASP8 and CASP9, AIFM, ENDOG, PARP | [311] | |
| Curcumin analog (WZ35) (57) | H1975 | ROS, ER stress, mitochondrial dysfunction | [312] | |
| Cucurbitacins (A, B, I and Q) | A549 | Inhibition of STAT3 | [321] | |
| Human laryngeal cancer | Cucurbitacin B (60) | Hep-2 | ↓ p-STAT3 | [325] |
| Human renal cancer | Curcumin (33) | Caki | ↑ ROS, TNFRSF10B | [304] |
| Human tenon’s capsule fibroblasts | Hydroxycamptothecin(51) | HTCFs | ↑ EIF2AK3/PERK | [271] |
| Human colon cancer | Camptothecin (5) | HCT116, RKO | ↑ BECN1, TP53 | [144] |
| Curcumin (33) | HCT116 | ↓ NFKB | [283] | |
| Resveratrol (32) | HT-29, COLO 201 | ROS, CASP3 and CASP8 activation | [314] | |
| Allicin (45) | HCT116 | ↑ CYCS, BAX ↓ BCL2 | [335] | |
| Genistein (27) | HT-29 | ↑ CDKN1A, BAX-BCL2 | [281] | |
| Ginsenoside compound K (22) | HT-29, HCT 116 | ↑ BAX, tBID, CYCS | [173] | |
| Diallyl disulfide | HT-29 | ↑ CASP9, CASP3 | [342] | |
| Urothelial carcinoma | Frondoside A (23) | RT112 RT4,HT-1197, TCC-SUP, T-24 | ↑ CASP3, CASP8 and CASP9, PARP, BAX, CDKN1A, DNA fragmentation | [175] |
| Glioma | Resveratrol (32) | U251 | Inhibiting autophagy | [315] |
| Ovarian cancer | Curcumin (33) | SKOV3 ES-2 | Activation of intrinsic and extrinsic pathways | [305] |
| Curcumin (33) | HEY OVCA429OCC1, SKOV3 | AKT signaling. ↑ CASP3, CASP8 and CASP9, CYCS, BID, ↓ BCL2 | [307] | |
| Human pancreatic cancer | Curcumin (33) | PANC1,BxPC3 | ↑ BAX, ↓ BCL2 | [229] |
| Human hepatocellular carcinoma | Resveratrol (32) | Huh-7 | ↑ CDKN1A, ↓ CCNE, CCNA, CDK2, p-MAPK/ERK, p-MAPK14/p38 | [209] |
| Brain cancer | Triazolyl curcumins (53, 54) | CRT-MG | Increased cytotoxicity and apoptosis | [306] |
| Gastric cancer | Curcumin (33) | MKN-28 | ↓ BCL2, ↑ BAX, CASP3 and CASP9 activation | [255] |
| Curcumin (33) + quercetin (25) | MGC-803 | ↑ CYCS, ↓ phosphorylation of AKT and MAPK/ERK | [309] | |
| Quercetin (25) | BGC-82 | ↓ BCL2:BAX ratio, ↑ CASP3 | [278] | |
| Magnolol (35) | SGC-7901 | ↓ PI3K-AKT, ↑ BAX- BCL2 and CASP3 | [234] | |
| Allicin (45) | SGC-7901 | ↑ FAS, BAX, CYTC, CASP3, CASP8, CASP9 | [338] | |
| Curcumin (33) | SGC-7901 | ↓ NFKB, BCL2, BCL2L1 | [284] | |
| Osteosarcoma | Curcumin (33) | MG63 | MAPK/JNK pathway, inhibition of autophagy | [171] |
| Human breast cancer | Allicin (45) | MCF-7, HCC-70 | ↑ CASP3, CASP8 and CASP9, CDKN1A/p21 PMAIP1/NOXA, BAK1 ↓ ∆Ψm, BCL2L1 | [339] |
| Chrysophanol (58) | MCF-7, BT-474 | PI3K-AKT and MAPK | [318] | |
| Curcumin (33) | MCF-7 | ↑ TP53, BAX | [288] | |
| MCF-7 | ↓ MIR21, AKT ↑ PTEN | [286] | ||
| T-47D | ↓ TP53, ESR1/ERα | [290] | ||
| MCF-10F | ↑ ROS | [308] | ||
| Quercetin (25) | Cancer stem cells (CSCs) | ↑ BAX, ↓ BCL2, PI3K-AKT-MTOR, | [279] | |
| Quercetin (25) | BT-20, MCF-7 | ↑ PARP cleavage, CASP3 and CASP7 | [276] | |
| Chrysin (29) | MCF-7 | ↑ ROS, CASP3 | [192] | |
| Rottlerin (38) | MCF-7 | ↑ CASP9, CASP3, PARP cleavage | [239] | |
| cancer stem cells (CSCs) | AKT-MTOR, AMPK, ↓ BCL2, BCL2L1, XIAP and BIRC2/cIAP-1 | [240] | ||
| Human fibrosarcoma | Rottlerin (38) | HT1080 | PRKCD-independent pathway | [236] |
| Triple-negative breast cancer | Curcumin (33) | MDA-MB-231 | ↓ p-MAPK1-MAPK3, EGFR inhibition | [297] |
| Epigallocatechin gallate (52) | Hs578T | ↓ BCL2 ↑ CASP3, BAX | [280] | |
| Human multiple myeloma | Curcumin (33) | MM RPMI 8226 | ↑ TP53, BAX | [289] |
| Head and neck squamous cell cancer | Curcumin (33) | MDA 686LN | ↓ NFKB, BCL2, CCND1, IL6, MT-CO2, MMP9 | [292] |
| Oral squamous cancer | Quercetin (25) | Cancer stem cell (CSCs) | ↓ MAPK14/p38-HSPB1 | [278] |
| Human leukemia | Curcumin (33) | HL60 | ↑ SMPD, ceramide generation | [293] |
| Ceramide generation, CASP8 and inhibition of BCL2L1 | [294] | |||
| K562 | ↑ BECN1, LC3-II, promoting apoptosis by autophagy | [295] | ||
| Apigenin (26) | HL60 | ↓ PI3K-AKT, JAK-STAT | [187] | |
| Multiple myeloma | Isobavachalcone (59) | H929 | Activation of PRKCD | [320] |
| Leiomyosarcoma | Curcumin (33) | SKN, SK-UT-1 | ↑ MAPK1-MAPK3, LC3B-II, ↓ SQSTM1 | [223] |
| Cancer Type | Compound | Cell Line | Target | Reference |
|---|---|---|---|---|
| Lung cancer | Polyphyllin D (71) | NCI-H460 | ↑ DDIT3, HSPA5, PDI, CASP4, ATF3, DDIT3, STC2 | [381] |
| Dehydrocostuslactone (73) | NCI-H460 A549 | ↑ EIF2AK3, DDIT3, ERN1, MAPK/JNK, ROS, XBP1s | [383] | |
| Resveratrol (32) | NCI-H460 | ↑ DDIT3, HSPA5, RECK, MIR200C | [370] | |
| Bisdemethoxycurcumin (BDMC) (56) | NCI-H460 | ↑ HSPA5; ERN1-ERN2; DDIT3; ATF6; ATF6B; CASP4 | [310] | |
| Demethoxycurcumin (DMC) (55) | NCI-H460 | ↑ HSPA5, ERN2, DDIT3, ATF6, ATF6B, CASP4, ROS ↓ ΔΨm | [311] | |
| WZ35 (57) | HI975 | ↑ p-EIF2A; ATF4; DDIT3 | [312] | |
| Resveratrol (32) + arsenic trioxide | A549 | ↑ HSPA5, DDIT3, CASP12 | [372] | |
| TMS Resveratrol (32) analog | A579; H1975 | ↑ EIF2A, EIF2AK3, DDIT3, Ca2+, AMPK | [373] | |
| Furanodiene (76) | A549, 95-D | ↑ DDIT3, HSPA5, DDIT3 | [386] | |
| Parthenolide (77) | A549, Calu-1, H1299, H1792 | ↑ ATF4, DDIT3, EIF2A, | [387] | |
| Human multiple myeloma | Resveratrol (32) | ANBL-6 | ↑ ERN1, EIF2AK3, ATF6, DDIT3, MAPK/JNK, PPP1R15A/GADD34 ↓ XBP1s, VEGFA | [367] |
| Breast cancer | Cryptotanshinone (74) | MCF-7 | ↑ EIF2A, HSP90B1/GRP94, HSPA5, DDIT3, ROS | [384] |
| Pimpinelol (75) | MCF-7 | ↑ ATF4, DDIT3, PPP1R15A, TRIB3 | [385] | |
| Ajoene (83) | MDA-MB-231 | ↑ HSPA5 | [396] | |
| Chrysophanol (58) | MCF-7; BT-474 | ↑ ROS, EIF2AK3, EIF2A, DDIT3, ERN1 | [318] | |
| Saxifragifolin D (72) | MDA-MB-231, MCF-7 | ↑ MAPK/JNK, Ca2+, ROS, DDIT3 | [382] | |
| Ampelopsin (64) | MDA-MB-231, MCF-7 | ↑ HSPA5, p-EIF2AK3, p-EIF2A, cleaved ATF6, DDIT3, ROS | [360] | |
| γ-tocotrienol (82) | MDA-MB-231, MCF-7 | ↑ DDIT3, HSPA5, XBP1, TNFRSF10B, TNFRSF10B, MAPK/JNK¸CASP3, CASP8 | [394] | |
| Colon cancer | Curcumin (33) | AGS, HT-29 | ↑ DDIT3, MAPK/JNK-FADD | [359] |
| Curcumin (33)+ sildenafil | HCT116, HT-29 | ↑ p-EIF2A; DDIT3 | [363] | |
| Curcumin (33)+ irinotecan (62) | LoVo, HT-29 | ↑ DDIT3, PDI, HSPA5 | [357] | |
| Piperine (61) | HT-29 | ↑ ERN1, DDIT3, HSPA5, ROS, MAPK/JNK | [355] | |
| Zerumbone (78) | HCT116-p53null, SW480 | ↑ ATF4, DDIT3, ATF3, ATF4, DDIT3, ATF3, EIF2AK3, EIF2A, TNFRSF10B, TNFRSF10B | [389] | |
| Flavokawain B (66) | HCT116 | ↑ DDIT3, DDIT3, ROS, BCL2 | [365] | |
| Gastric cancer (GC) | α-Tocopheryl succinate (79) | SGC-7901 | ↑ DDIT3, MAPK/JNK, EIF2AK3, ATF6, XBP1, ATF4 HSP90B1/GRP94, HSPA5, CASP4 | [390] |
| 7-Acetylsinumaximol B (81) | NCI-N87 | ↑ EIF2AK3, EIF2A, ATF4, DDIT3 | [393] | |
| α-Tocopheryl succinate (79) | SGC-7901 | ↑ DDIT3, MAPK/JNK, HSPA5, ROS, CASP4 | [391] | |
| Honokiol (34) | MKN-45 SCM-1, AGS, N87 | ↑ DDIT3, CAPN2, PARP and HSP90B1/GRP94, cleavage | [379] | |
| Casticin (63) | BGC-823 | ↑ DDIT3, p-EIF2A, EIF2A, HSPA5 | [359] | |
| Casticin (63) | SGC-7901 MGC-803 | ↑ DDIT3, TNFRSF10B, ROS | [359] | |
| Malignant mesothelioma | Epigallocatechin gallate (52) | MM98 | ↑ HSPA5; DDIT3; ATF4; EDEM; XBP1 | [358] |
| Liver cancer | Curcumin (33)+ sildenafil | HEPG2 | ↑ EIF2A, BECN1; ↓ MTORC1 and MTORC2 activity | [363] |
| Murine Myeloma | Curcumin (33) | WEHI-3 | ↑ ATF6; DDIT3; ERN1; CASP12; BCL2, ROS, Ca2+, ↓ ΔΨm | [361] |
| Human melanoma | 4-nerolidylcatechol (80) | SK-MEL-28; BRAi/MEKi | ↑ HSPA5, DDIT3 | [392]. |
| Vincristine (3) | B16 | ↑ AMPK, TP53 ↓MTORC1 | [264] | |
| Resveratrol (32) | A375SM | ↑ p-EIF2A, DDIT3, ROS, MAPK14/p38, TP53, BAX, ↓ BCL2 | [368] | |
| Pheochromocytoma | α-Mangostin (67) | PC12 | ↓ ATP2A/Ca2+ ATPase activity ↑ MAPK/JNK, MAPK/SAPK | [376] |
| Human ovarian cancer | Garcinone E (68) | HEY; A2780, A2780/Taxol cells | ↑ HSPA5, ERN1, XBP1, DDIT3, CASP12 | [377] |
| B19 (65) | HO8910 | ↑ PDI, HSPA5, DDIT3, ATF6, XBP1, CASP3 | [359] | |
| Human hepatoblastoma | Resveratrol (32) + palmitate | HepG2 | ↑ XBP1s, DDIT3, ↓ ROS | [369] |
| RES006 Resveratrol (32) analog | HepG2 | ↑ ROS | [374] | |
| Human nasopharyngeal cancer | Resveratrol (32) | NPC-TW076; NPC-TW039 | ↑ ERN1, DDIT3, ATF6, p-EIF2AK3 | [371] |
| Human prostate cancer | Gartanin (69) | LNCaP; 22RV1 | ↑ DDIT3, HSPA5, ERN1, EIF2AK3 | [378]. |
| Polyphenon E | PNT1a | ↑ DDIT3, HSPA5 p-EIF2A and ATF4 | [398] | |
| PC3 | ↑ DDIT3, (u)XBP1 p-EIF2A and ATF4 | [398] | ||
| Garlic extract | DU-145 DU145, U2OS,67NR | ↑ HSPA5 | [395] | |
| Curcumin (33) | PC3 | ↑ ERN1; ROS; PDI; CALR | [362] | |
| Human hepatocellular carcinoma | Garcinol (70) | Hep3B | ↑ DDIT3, DDIT3, ROS, BAX, CASP3, CASP9, PARP; ↓ ΔΨm, BCL2 | [380] |
| Human esophageal cancer | Ajoene (83) | WHCO1 | ↑ HSPA5 | [396] |
| BisPMB (84) | WHCO1 | ↑ HSPA5, DDIT3 | [397] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martelli, A.; Omrani, M.; Zarghooni, M.; Citi, V.; Brogi, S.; Calderone, V.; Sureda, A.; Lorzadeh, S.; da Silva Rosa, S.C.; Grabarek, B.O.; et al. New Visions on Natural Products and Cancer Therapy: Autophagy and Related Regulatory Pathways. Cancers 2022, 14, 5839. https://doi.org/10.3390/cancers14235839
Martelli A, Omrani M, Zarghooni M, Citi V, Brogi S, Calderone V, Sureda A, Lorzadeh S, da Silva Rosa SC, Grabarek BO, et al. New Visions on Natural Products and Cancer Therapy: Autophagy and Related Regulatory Pathways. Cancers. 2022; 14(23):5839. https://doi.org/10.3390/cancers14235839
Chicago/Turabian StyleMartelli, Alma, Marzieh Omrani, Maryam Zarghooni, Valentina Citi, Simone Brogi, Vincenzo Calderone, Antoni Sureda, Shahrokh Lorzadeh, Simone C. da Silva Rosa, Beniamin Oscar Grabarek, and et al. 2022. "New Visions on Natural Products and Cancer Therapy: Autophagy and Related Regulatory Pathways" Cancers 14, no. 23: 5839. https://doi.org/10.3390/cancers14235839
APA StyleMartelli, A., Omrani, M., Zarghooni, M., Citi, V., Brogi, S., Calderone, V., Sureda, A., Lorzadeh, S., da Silva Rosa, S. C., Grabarek, B. O., Staszkiewicz, R., Los, M. J., Nabavi, S. F., Nabavi, S. M., Mehrbod, P., Klionsky, D. J., & Ghavami, S. (2022). New Visions on Natural Products and Cancer Therapy: Autophagy and Related Regulatory Pathways. Cancers, 14(23), 5839. https://doi.org/10.3390/cancers14235839