


Insights into the Role of Sialylation in Cancer Metastasis, Immunity, and Therapeutic Opportunity
Abstract
:Simple Summary
Abstract
1. Introduction
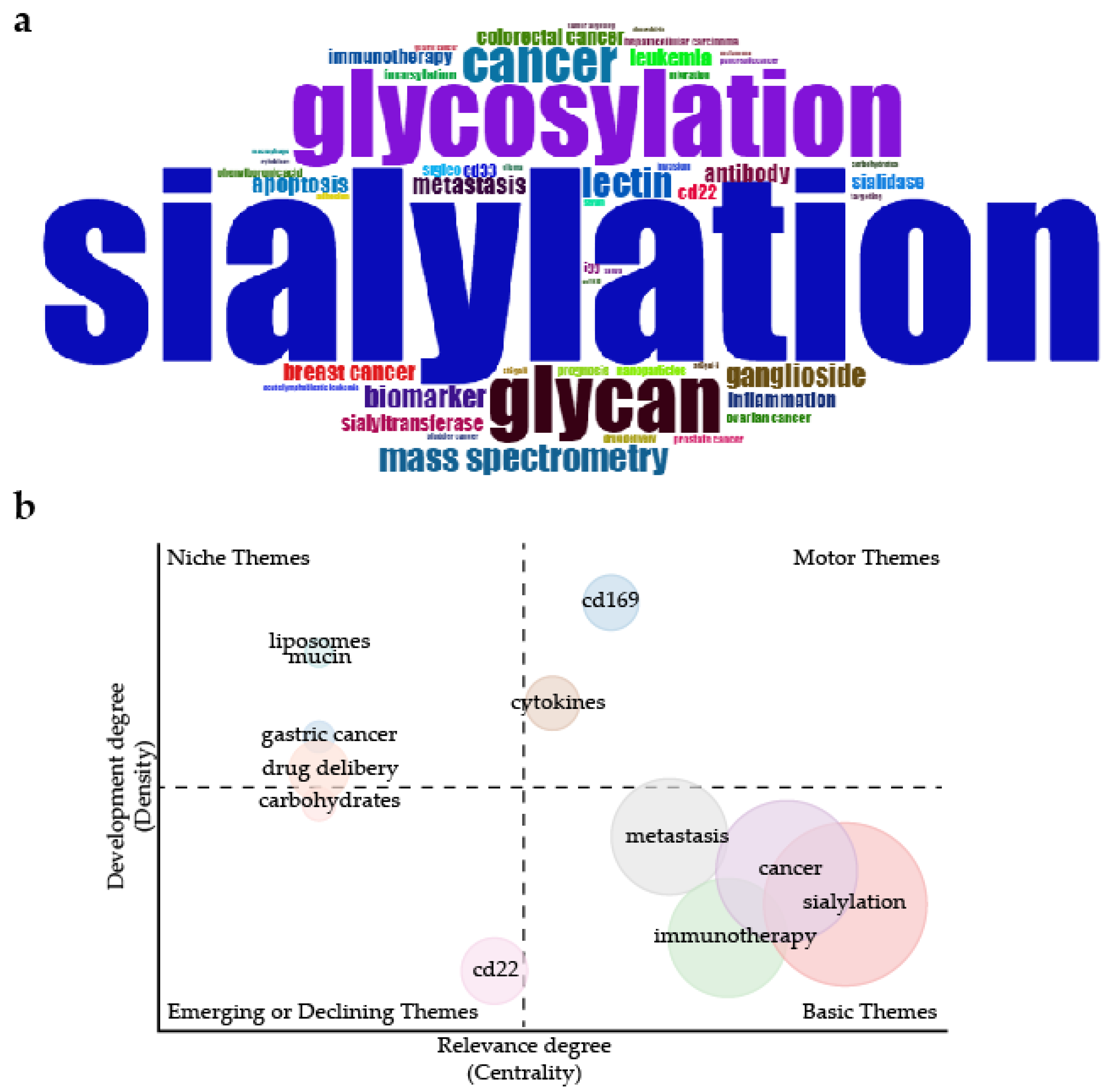
2. Overview of Articles on Sialylation in Cancer
3. Structural Basis of Sialylation
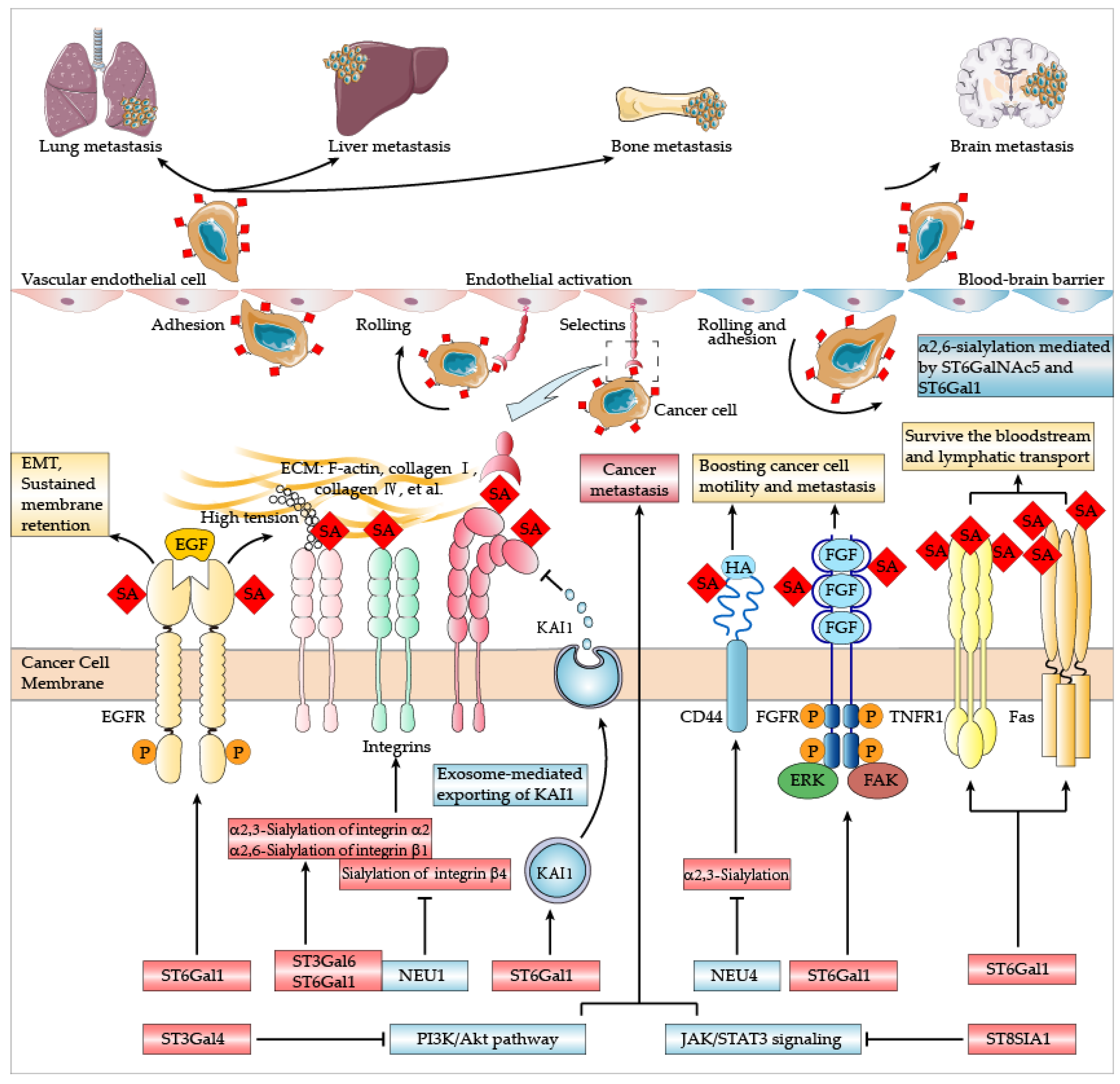
4. Sialylation and Cancer Metastasis
4.1. Lung Metastasis
4.2. Liver Metastasis
4.3. Bone Metastasis
4.4. Brain Metastasis
4.5. Controversial Roles in Metastasis
5. Sialylation and Cancer Immunity
5.1. Immune Recognition
5.2. Anti-Cancer Immunity Modulation
6. Sialylation and Anti-Cancer Therapy
6.1. Blocking Sialic Acid Metabolism
6.2. Interfering with Ligand-Receptor Interaction
6.3. Antibody-Sialidase Conjugates
6.4. Vaccines Development
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Li, F.; Ding, J. Sialylation is involved in cell fate decision during development, reprogramming and cancer progression. Protein Cell 2018, 10, 550–565. [Google Scholar] [CrossRef] [PubMed]
- Pietrobono, S.; Stecca, B. Aberrant Sialylation in Cancer: Biomarker and Potential Target for Therapeutic Intervention? Cancers 2021, 13, 2014. [Google Scholar] [CrossRef] [PubMed]
- Bull, C.; Stoel, M.A.; den Brok, M.H.; Adema, G.J. Sialic acids sweeten a tumor’s life. Cancer Res. 2014, 74, 3199–3204. [Google Scholar] [CrossRef] [Green Version]
- Munkley, J. Aberrant Sialylation in Cancer: Therapeutic Opportunities. Cancers 2022, 14, 4248. [Google Scholar] [CrossRef]
- Macbeth, R.A.L.; Bekesi, J.G. Plasma glycoproteins in various disease states including carcinoma. Cancer Res. 1962, 22, 1170–1176. [Google Scholar]
- Pearce, O.M.T.; Läubli, H. Sialic acids in cancer biology and immunity. Glycobiology 2016, 26, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Ip, C.; Yin, J.; Ng, P.; Lin, S.-Y.; Mills, G. Genomic-Glycosylation Aberrations in Tumor Initiation, Progression and Management. AIMS Med. Sci. 2016, 3, 386–416. [Google Scholar] [CrossRef]
- Wang, W.Y.; Cao, Y.X.; Zhou, X.; Wei, B.; Zhan, L.; Sun, S.Y. Stimulative role of ST6GALNAC1 in proliferation, migration and invasion of ovarian cancer stem cells via the Akt signaling pathway. Cancer Cell Int. 2019, 19, 86. [Google Scholar] [CrossRef] [Green Version]
- Wichert, B.; Milde-Langosch, K.; Galatenko, V.; Schmalfeldt, B.; Oliveira-Ferrer, L. Prognostic role of the sialyltransferase ST6GAL1 in ovarian cancer. Glycobiology 2018, 28, 898–903. [Google Scholar] [CrossRef] [Green Version]
- Swindall, A.F.; Londoño-Joshi, A.I.; Schultz, M.J.; Fineberg, N.; Buchsbaum, D.J.; Bellis, S.L. ST6Gal-I protein expression is upregulated in human epithelial tumors and correlates with stem cell markers in normal tissues and colon cancer cell lines. Cancer Res. 2013, 73, 2368–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büll, C.; Boltje, T.J.; Wassink, M.; de Graaf, A.M.; van Delft, F.L.; den Brok, M.H.; Adema, G.J. Targeting aberrant sialylation in cancer cells using a fluorinated sialic acid analog impairs adhesion, migration, and in vivo tumor growth. Mol. Cancer Ther. 2013, 12, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Jandus, C.; Boligan, K.; Chijioke, O.; Liu, H.; Dahlhaus, M.; Démoulins, T.; Schneider, C.; Wehrli, M.; Hunger, R.; Baerlocher, G.; et al. Interactions between Siglec-7/9 receptors and ligands influence NK cell-dependent tumor immunosurveillance. J. Clin. Investig. 2014, 124, 1810–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, R.B.; Appelbaum, F.R.; Estey, E.H.; Bernstein, I.D. Acute myeloid leukemia stem cells and CD33-targeted immunotherapy. Blood 2012, 119, 6198–6208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiding, K.R.; Blank, D.; Kuijper, D.M.; Deelder, A.M.; Wuhrer, M. High-throughput profiling of protein N-glycosylation by MALDI-TOF-MS employing linkage-specific sialic acid esterification. Anal. Chem. 2014, 86, 5784–5793. [Google Scholar] [CrossRef] [PubMed]
- Alley, W.R., Jr.; Vasseur, J.A.; Goetz, J.A.; Svoboda, M.; Mann, B.F.; Matei, D.E.; Menning, N.; Hussein, A.; Mechref, Y.; Novotny, M.V. N-linked glycan structures and their expressions change in the blood sera of ovarian cancer patients. J. Proteome Res. 2012, 11, 2282–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.J.; Yi, J.Y.; Jin, Y.B.; Lee, Y.J.; Lee, J.S.; Lee, Y.S.; Ko, Y.G.; Lee, M. Sialylation of epidermal growth factor receptor regulates receptor activity and chemosensitivity to gefitinib in colon cancer cells. Biochem. Pharmacol. 2012, 83, 849–857. [Google Scholar] [CrossRef]
- Schultz, M.J.; Swindall, A.F.; Wright, J.W.; Sztul, E.S.; Landen, C.N.; Bellis, S.L. ST6Gal-I sialyltransferase confers cisplatin resistance in ovarian tumor cells. J. Ovarian. Res. 2013, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Deshayes, S.; Cabral, H.; Ishii, T.; Miura, Y.; Kobayashi, S.; Yamashita, T.; Matsumoto, A.; Miyahara, Y.; Nishiyama, N.; Kataoka, K. Phenylboronic acid-installed polymeric micelles for targeting sialylated epitopes in solid tumors. J. Am. Chem. Soc. 2013, 135, 15501–15507. [Google Scholar] [CrossRef] [PubMed]
- Haso, W.; Lee, D.W.; Shah, N.N.; Stetler-Stevenson, M.; Yuan, C.M.; Pastan, I.H.; Dimitrov, D.S.; Morgan, R.A.; FitzGerald, D.J.; Barrett, D.M.; et al. Anti-CD22-chimeric antigen receptors targeting B-cell precursor acute lymphoblastic leukemia. Blood 2013, 121, 1165–1174. [Google Scholar] [CrossRef] [Green Version]
- Mosimann, S.C.; Gilbert, M.; Dombroswki, D.; To, R.; Wakarchuk, W.; Strynadka, N.C. Structure of a sialic acid-activating synthetase, CMP-acylneuraminate synthetase in the presence and absence of CDP. J. Biol. Chem. 2001, 276, 8190–8196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, M.M.; McArthur, J.B.; Li, Y.; Yu, H.; Chen, X.; Fisher, A.J. Catalytic Cycle of Neisseria meningitidis CMP-Sialic Acid Synthetase Illustrated by High-Resolution Protein Crystallography. Biochemistry 2020, 59, 3157–3168. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.E.; Thomas, P. Inhibition of CMP-sialic acid transport in human liver and colorectal cancer cell lines by a sialic acid nucleoside conjugate (KI-8110). Biochem. Biophys. Res. Commun. 1993, 190, 571–575. [Google Scholar] [CrossRef]
- Nji, E.; Gulati, A.; Qureshi, A.A.; Coincon, M.; Drew, D. Structural basis for the delivery of activated sialic acid into Golgi for sialyation. Nat. Struct. Mol. Biol. 2019, 26, 415–423. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, T.L.; Vertel, B.M.; Colley, K.J. The CMP-sialic acid transporter is localized in the medial-trans Golgi and possesses two specific endoplasmic reticulum export motifs in its carboxyl-terminal cytoplasmic tail. J. Biol. Chem. 2006, 281, 31106–31118. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Mukhopadhyay, S. A three-pocket model for substrate coordination and selectivity by the nucleotide sugar transporters SLC35A1 and SLC35A2. J. Biol. Chem. 2021, 297, 101069. [Google Scholar] [CrossRef]
- Schultz, M.J.; Swindall, A.F.; Bellis, S.L. Regulation of the metastatic cell phenotype by sialylated glycans. Cancer Metastasis Rev. 2012, 31, 501–518. [Google Scholar] [CrossRef] [Green Version]
- Marcelo, F.; Garcia-Martin, F.; Matsushita, T.; Sardinha, J.; Coelho, H.; Oude-Vrielink, A.; Koller, C.; Andre, S.; Cabrita, E.; Gabius, H.; et al. Delineating binding modes of Gal/GalNAc and structural elements of the molecular recognition of tumor-associated mucin glycopeptides by the human macrophage galactose-type lectin. Chemistry 2014, 20, 16147–16155. [Google Scholar] [CrossRef]
- Gabba, A.; Bogucka, A.; Luz, J.; Diniz, A.; Coelho, H.; Corzana, F.; Canada, F.; Marcelo, F.; Murphy, P.; Birrane, G. Crystal Structure of the Carbohydrate Recognition Domain of the Human Macrophage Galactose C-Type Lectin Bound to GalNAc and the Tumor-Associated Tn Antigen. Biochemistry 2021, 60, 1327–1336. [Google Scholar] [CrossRef]
- Jegouzo, S.A.; Quintero-Martinez, A.; Ouyang, X.; dos Santos, A.; Taylor, M.E.; Drickamer, K. Organization of the extracellular portion of the macrophage galactose receptor: A trimeric cluster of simple binding sites for N-acetylgalactosamine. Glycobiology 2013, 23, 853–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatson, R.; Maurstad, G.; Picco, G.; Arulappu, A.; Coleman, J.; Wandell, H.H.; Clausen, H.; Mandel, U.; Taylor-Papadimitriou, J.; Sletmoen, M.; et al. The Breast Cancer-Associated Glycoforms of MUC1, MUC1-Tn and sialyl-Tn, Are Expressed in COSMC Wild-Type Cells and Bind the C-Type Lectin MGL. PLoS ONE 2015, 10, e0125994. [Google Scholar] [CrossRef]
- Zhuo, Y.; Chammas, R.; Bellis, S.L. Sialylation of beta1 integrins blocks cell adhesion to galectin-3 and protects cells against galectin-3-induced apoptosis. J. Biol. Chem. 2008, 283, 22177–22185. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.E.; O’Sullivan, J.M.; Moran, A.B.; Spencer, D.I.R.; Gardner, R.A.; Sharma, J.; Fazavana, J.; Monopoli, M.; McKinnon, T.A.J.; Chion, A.; et al. Sialylation on O-linked glycans protects von Willebrand factor from macrophage galactose lectin-mediated clearance. Haematologica 2022, 107, 668–679. [Google Scholar] [CrossRef]
- Blaum, B.S.; Hannan, J.P.; Herbert, A.P.; Kavanagh, D.; Uhrin, D.; Stehle, T. Structural basis for sialic acid-mediated self-recognition by complement factor H. Nat. Chem. Biol. 2015, 11, 77–82. [Google Scholar] [CrossRef]
- Thiesler, H.; Kucukerden, M.; Gretenkort, L.; Rockle, I.; Hildebrandt, H. News and Views on Polysialic Acid: From Tumor Progression and Brain Development to Psychiatric Disorders, Neurodegeneration, Myelin Repair and Immunomodulation. Front. Cell Dev. Biol. 2022, 10, 871757. [Google Scholar] [CrossRef]
- Villanueva-Cabello, T.M.; Gutierrez-Valenzuela, L.D.; Salinas-Marin, R.; Lopez-Guerrero, D.V.; Martinez-Duncker, I. Polysialic Acid in the Immune System. Front. Immunol. 2021, 12, 823637. [Google Scholar] [CrossRef]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- van Zijl, F.; Krupitza, G.; Mikulits, W. Initial steps of metastasis: Cell invasion and endothelial transmigration. Mutat. Res. 2011, 728, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Seyfried, T.N.; Huysentruyt, L.C. On the origin of cancer metastasis. Crit. Rev. Oncog. 2013, 18, 43–73. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Joon, A.; Kumari, M.; Singh, T.; Bal, A.; Maan, P.; Ghosh, S. Role of a Disease-associated ST3Gal-4 in Non-small Cell Lung Cancer. Cell Biochem. Biophys. 2022, 80, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Britain, C.M.; Bhalerao, N.; Silva, A.D.; Chakraborty, A.; Buchsbaum, D.J.; Crowley, M.R.; Crossman, D.K.; Edwards, Y.J.K.; Bellis, S.L. Glycosyltransferase ST6Gal-I promotes the epithelial to mesenchymal transition in pancreatic cancer cells. J. Biol. Chem. 2021, 296, 100034. [Google Scholar] [CrossRef] [PubMed]
- Rao, T.C.; Ma, V.P.; Blanchard, A.; Urner, T.M.; Grandhi, S.; Salaita, K.; Mattheyses, A.L. EGFR activation attenuates the mechanical threshold for integrin tension and focal adhesion formation. J. Cell Sci. 2020, 133, jcs238840. [Google Scholar] [CrossRef]
- Rao, T.C.; Beggs, R.R.; Ankenbauer, K.E.; Hwang, J.; Ma, V.P.; Salaita, K.; Bellis, S.L.; Mattheyses, A.L. ST6Gal-I-mediated sialylation of the epidermal growth factor receptor modulates cell mechanics and enhances invasion. J. Biol. Chem. 2022, 298, 101726. [Google Scholar] [CrossRef] [PubMed]
- Christie, D.R.; Shaikh, F.M.; Lucas, J.A.t.; Lucas, J.A., 3rd; Bellis, S.L. ST6Gal-I expression in ovarian cancer cells promotes an invasive phenotype by altering integrin glycosylation and function. J. Ovarian Res. 2008, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Semel, A.C.; Seales, E.C.; Singhal, A.; Eklund, E.A.; Colley, K.J.; Bellis, S.L. Hyposialylation of integrins stimulates the activity of myeloid fibronectin receptors. J. Biol. Chem. 2002, 277, 32830–32836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seales, E.C.; Jurado, G.A.; Singhal, A.; Bellis, S.L. Ras oncogene directs expression of a differentially sialylated, functionally altered beta1 integrin. Oncogene 2003, 22, 7137–7145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dou, P.; Akhtar, M.L.; Liu, F.; Hu, X.; Yang, L.; Yang, D.; Zhang, X.; Li, Y.; Qiao, S.; et al. NEU4 inhibits motility of HCC cells by cleaving sialic acids on CD44. Oncogene 2021, 40, 5427–5440. [Google Scholar] [CrossRef]
- Ou, L.; He, X.; Liu, N.; Song, Y.; Li, J.; Gao, L.; Huang, X.; Deng, Z.; Wang, X.; Lin, S. Sialylation of FGFR1 by ST6GalI overexpression contributes to ovarian cancer cell migration and chemoresistance. Mol. Med. Rep. 2020, 21, 1449–1460. [Google Scholar] [CrossRef] [Green Version]
- Cazet, A.; Julien, S.; Bobowski, M.; Krzewinski-Recchi, M.A.; Harduin-Lepers, A.; Groux-Degroote, S.; Delannoy, P. Consequences of the expression of sialylated antigens in breast cancer. Carbohydr. Res. 2010, 345, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, H.; Matsuzaki, H.; Ando, H.; Kaji, H.; Nakanishi, H.; Ikehara, Y.; Narimatsu, H. Enhancement of metastatic ability by ectopic expression of ST6GalNAcI on a gastric cancer cell line in a mouse model. Clin. Exp. Metastasis 2012, 29, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Elkashef, S.M.; Allison, S.J.; Sadiq, M.; Basheer, H.A.; Ribeiro Morais, G.; Loadman, P.M.; Pors, K.; Falconer, R.A. Polysialic acid sustains cancer cell survival and migratory capacity in a hypoxic environment. Sci. Rep. 2016, 6, 33026. [Google Scholar] [CrossRef] [Green Version]
- Rosa, P.; Scibetta, S.; Pepe, G.; Mangino, G.; Capocci, L.; Moons, S.J.; Boltje, T.J.; Fazi, F.; Petrozza, V.; Di Pardo, A.; et al. Polysialic Acid Sustains the Hypoxia-Induced Migration and Undifferentiated State of Human Glioblastoma Cells. Int. J. Mol. Sci. 2022, 23, 9563. [Google Scholar] [CrossRef]
- Bindeman, W.E.; Fingleton, B. Glycosylation as a regulator of site-specific metastasis. Cancer Metastasis Rev. 2022, 41, 107–129. [Google Scholar] [CrossRef]
- Uemura, T.; Shiozaki, K.; Yamaguchi, K.; Miyazaki, S.; Satomi, S.; Kato, K.; Sakuraba, H.; Miyagi, T. Contribution of sialidase NEU1 to suppression of metastasis of human colon cancer cells through desialylation of integrin beta4. Oncogene 2009, 28, 1218–1229. [Google Scholar] [CrossRef] [Green Version]
- Van Slambrouck, S.; Groux-Degroote, S.; Krzewinski-Recchi, M.A.; Cazet, A.; Delannoy, P.; Steelant, W.F. Carbohydrate-to-carbohydrate interactions between α2,3-linked sialic acids on α2 integrin subunits and asialo-GM1 underlie the bone metastatic behaviour of LNCAP-derivative C4-2B prostate cancer cells. Biosci. Rep. 2014, 34, 546–557. [Google Scholar] [CrossRef]
- Jung, Y.R.; Park, J.J.; Jin, Y.B.; Cao, Y.J.; Park, M.J.; Kim, E.J.; Lee, M. Silencing of ST6Gal I enhances colorectal cancer metastasis by down-regulating KAI1 via exosome-mediated exportation and thereby rescues integrin signaling. Carcinogenesis 2016, 37, 1089–1097. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Hu, J.; Ma, J.; Qi, X.; Zhou, H.; Miao, X.; Zheng, W.; Jia, L. MiR-193a-3p and miR-224 mediate renal cell carcinoma progression by targeting alpha-2,3-sialyltransferase IV and the phosphatidylinositol 3 kinase/Akt pathway. Mol. Carcinog. 2018, 57, 1067–1077. [Google Scholar] [CrossRef]
- Yu, S.; Wang, S.; Sun, X.; Wu, Y.; Zhao, J.; Liu, J.; Yang, D.; Jiang, Y. ST8SIA1 inhibits the proliferation, migration and invasion of bladder cancer cells by blocking the JAK/STAT signaling pathway. Oncol. Lett. 2021, 22, 736. [Google Scholar] [CrossRef] [PubMed]
- Laubli, H.; Borsig, L. Selectins as mediators of lung metastasis. Cancer Microenviron. 2010, 3, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiratsuka, S.; Goel, S.; Kamoun, W.S.; Maru, Y.; Fukumura, D.; Duda, D.G.; Jain, R.K. Endothelial focal adhesion kinase mediates cancer cell homing to discrete regions of the lungs via E-selectin up-regulation. Proc. Natl. Acad. Sci. USA 2011, 108, 3725–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laubli, H.; Spanaus, K.S.; Borsig, L. Selectin-mediated activation of endothelial cells induces expression of CCL5 and promotes metastasis through recruitment of monocytes. Blood 2009, 114, 4583–4591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnoub, A.E.; Dash, A.B.; Vo, A.P.; Sullivan, A.; Brooks, M.W.; Bell, G.W.; Richardson, A.L.; Polyak, K.; Tubo, R.; Weinberg, R.A. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature 2007, 449, 557–563. [Google Scholar] [CrossRef]
- Lange, T.; Valentiner, U.; Wicklein, D.; Maar, H.; Labitzky, V.; Ahlers, A.K.; Starzonek, S.; Genduso, S.; Staffeldt, L.; Pahlow, C.; et al. Tumor cell E-selectin ligands determine partialefficacy of bortezomib on spontaneous lung metastasis formation of solid human tumors in vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 1536–1552. [Google Scholar] [CrossRef]
- Ghirardello, M.; Shyam, R.; Galan, M.C. Reengineering of cancer cell surface charges can modulate cell migration. Chem. Commun. 2022, 58, 5522–5525. [Google Scholar] [CrossRef]
- Büll, C.; Boltje, T.J.; van Dinther, E.A.; Peters, T.; de Graaf, A.M.; Leusen, J.H.; Kreutz, M.; Figdor, C.G.; den Brok, M.H.; Adema, G.J. Targeted delivery of a sialic acid-blocking glycomimetic to cancer cells inhibits metastatic spread. ACS Nano 2015, 9, 733–745. [Google Scholar] [CrossRef]
- Kaur, S.; Kumar, S.; Momi, N.; Sasson, A.R.; Batra, S.K. Mucins in pancreatic cancer and its microenvironment. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Lakshmanan, I.; Chaudhary, S.; Vengoji, R.; Seshacharyulu, P.; Rachagani, S.; Carmicheal, J.; Jahan, R.; Atri, P.; Chirravuri-Venkata, R.; Gupta, R.; et al. ST6GalNAc-I promotes lung cancer metastasis by altering MUC5AC sialylation. Mol. Oncol. 2021, 15, 1866–1881. [Google Scholar] [CrossRef]
- Chen, S.H.; Dallas, M.R.; Balzer, E.M.; Konstantopoulos, K. Mucin 16 is a functional selectin ligand on pancreatic cancer cells. FASEB J. 2012, 26, 1349–1359. [Google Scholar] [CrossRef]
- Zhao, Q.; Barclay, M.; Hilkens, J.; Guo, X.; Barrow, H.; Rhodes, J.M.; Yu, L.G. Interaction between circulating galectin-3 and cancer-associated MUC1 enhances tumour cell homotypic aggregation and prevents anoikis. Mol. Cancer 2010, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Gupta, B.K.; Maher, D.M.; Ebeling, M.C.; Sundram, V.; Koch, M.D.; Lynch, D.W.; Bohlmeyer, T.; Watanabe, A.; Aburatani, H.; Puumala, S.E.; et al. Increased expression and aberrant localization of mucin 13 in metastatic colon cancer. J. Histochem. Cytochem. 2012, 60, 822–831. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, P.L.; Auger, F.A.; Huot, J. Regulation of transendothelial migration of colon cancer cells by E-selectin-mediated activation of p38 and ERK MAP kinases. Oncogene 2006, 25, 6563–6573. [Google Scholar] [CrossRef] [Green Version]
- Brodt, P.; Fallavollita, L.; Bresalier, R.S.; Meterissian, S.; Norton, C.R.; Wolitzky, B.A. Liver endothelial E-selectin mediates carcinoma cell adhesion and promotes liver metastasis. Int. J. Cancer 1997, 71, 612–619. [Google Scholar] [CrossRef]
- Esposito, M.; Mondal, N.; Greco, T.M.; Wei, Y.; Spadazzi, C.; Lin, S.C.; Zheng, H.; Cheung, C.; Magnani, J.L.; Lin, S.H.; et al. Bone vascular niche E-selectin induces mesenchymal-epithelial transition and Wnt activation in cancer cells to promote bone metastasis. Nat. Cell Biol. 2019, 21, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Aoki, M.; Kannagi, R. Transcription factors c-Myc and CDX2 mediate E-selectin ligand expression in colon cancer cells undergoing EGF/bFGF-induced epithelial-mesenchymal transition. Proc. Natl. Acad. Sci. USA 2012, 109, 7776–7781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, T.T.; Burness, M.L.; Sivan, A.; Warner, M.J.; Cheng, R.; Lee, C.H.; Olivere, L.; Comatas, K.; Magnani, J.; Kim Lyerly, H.; et al. Dormant breast cancer micrometastases reside in specific bone marrow niches that regulate their transit to and from bone. Sci. Transl. Med. 2016, 8, 340ra373. [Google Scholar] [CrossRef] [PubMed]
- Julien, S.; Ivetic, A.; Grigoriadis, A.; QiZe, D.; Burford, B.; Sproviero, D.; Picco, G.; Gillett, C.; Papp, S.L.; Schaffer, L.; et al. Selectin ligand sialyl-Lewis x antigen drives metastasis of hormone-dependent breast cancers. Cancer Res. 2011, 71, 7683–7693. [Google Scholar] [CrossRef] [PubMed]
- Acikgoz, E.; Duzagac, F.; Guven, U.; Yigitturk, G.; Kose, T.; Oktem, G. “Double hit” strategy: Removal of sialic acid from the dendritic cell surface and loading with CD44+/CD24-/low cell lysate inhibits tumor growth and metastasis by targeting breast cancer stem cells. Int. Immunopharmacol. 2022, 107, 108684. [Google Scholar] [CrossRef]
- Glavey, S.V.; Manier, S.; Natoni, A.; Sacco, A.; Moschetta, M.; Reagan, M.R.; Murillo, L.S.; Sahin, I.; Wu, P.; Mishima, Y.; et al. The sialyltransferase ST3GAL6 influences homing and survival in multiple myeloma. Blood 2014, 124, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Goli, M.; Mirzaei, P.; Mechref, Y. Revealing the Biological Attributes of N-Glycan Isomers in Breast Cancer Brain Metastasis Using Porous Graphitic Carbon (PGC) Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS). J. Proteome Res. 2019, 18, 3731–3740. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Mirzaei, P.; Zhu, R.; Zhou, S.; Mechref, Y. Comparative Membrane N-Glycomics of Different Breast Cancer Cell Lines To Understand Breast Cancer Brain Metastasis. J. Proteome Res. 2020, 19, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Zhu, R.; Zhou, S.; Mirzaei, P.; Mechref, Y. Integrated Transcriptomics, Proteomics, and Glycomics Reveals the Association between Up-regulation of Sialylated N-glycans/Integrin and Breast Cancer Brain Metastasis. Sci. Rep. 2019, 9, 17361. [Google Scholar] [CrossRef]
- Bos, P.D.; Zhang, X.H.; Nadal, C.; Shu, W.; Gomis, R.R.; Nguyen, D.X.; Minn, A.J.; van de Vijver, M.J.; Gerald, W.L.; Foekens, J.A.; et al. Genes that mediate breast cancer metastasis to the brain. Nature 2009, 459, 1005–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finke, J.M.; Ayres, K.R.; Brisbin, R.P.; Hill, H.A.; Wing, E.E.; Banks, W.A. Antibody blood-brain barrier efflux is modulated by glycan modification. Biochim. Biophys. Acta. Gen. Subj. 2017, 1861, 2228–2239. [Google Scholar] [CrossRef]
- Julien, S.; Adriaenssens, E.; Ottenberg, K.; Furlan, A.; Courtand, G.; Vercoutter-Edouart, A.S.; Hanisch, F.G.; Delannoy, P.; Le Bourhis, X. ST6GalNAc I expression in MDA-MB-231 breast cancer cells greatly modifies their O-glycosylation pattern and enhances their tumourigenicity. Glycobiology 2006, 16, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julien, S.; Lagadec, C.; Krzewinski-Recchi, M.A.; Courtand, G.; Le Bourhis, X.; Delannoy, P. Stable expression of sialyl-Tn antigen in T47-D cells induces a decrease of cell adhesion and an increase of cell migration. Breast Cancer Res. Treat. 2005, 90, 77–84. [Google Scholar] [CrossRef]
- Sewell, R.; Backstrom, M.; Dalziel, M.; Gschmeissner, S.; Karlsson, H.; Noll, T.; Gatgens, J.; Clausen, H.; Hansson, G.C.; Burchell, J.; et al. The ST6GalNAc-I sialyltransferase localizes throughout the Golgi and is responsible for the synthesis of the tumor-associated sialyl-Tn O-glycan in human breast cancer. J. Biol. Chem. 2006, 281, 3586–3594. [Google Scholar] [CrossRef] [Green Version]
- Murugaesu, N.; Iravani, M.; van Weverwijk, A.; Ivetic, A.; Johnson, D.A.; Antonopoulos, A.; Fearns, A.; Jamal-Hanjani, M.; Sims, D.; Fenwick, K.; et al. An in vivo functional screen identifies ST6GalNAc2 sialyltransferase as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, C.M.; Reginato, M.J. Sticking to sugars at the metastatic site: Sialyltransferase ST6GalNAc2 acts as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 275–277. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, J.; Mi, W.; Yang, J.; Han, F.; Lu, X.; Yu, W. Silencing of GM3 synthase suppresses lung metastasis of murine breast cancer cells. Breast Cancer Res. 2008, 10, R1. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Yamashita, S.; Endoh, M.; Yamato, T.; Hoshi, S.; Ohyama, C.; Watanabe, R.; Ito, A.; Satoh, M.; Wada, T.; et al. Clinical significance of ST3Gal IV expression in human renal cell carcinoma. Oncol. Rep. 2002, 9, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Kroes, R.A.; He, H.; Emmett, M.R.; Nilsson, C.L.; Leach, F.E., 3rd; Amster, I.J.; Marshall, A.G.; Moskal, J.R. Overexpression of ST6GalNAcV, a ganglioside-specific alpha2,6-sialyltransferase, inhibits glioma growth in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 12646–12651. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, D.X.; Zhang, X.W.; Jin, B.; Zhang, Z.; Xie, B.S.; Wu, C.L.; Gong, K.; Mao, Z.B. CSTP1, a novel protein phosphatase, blocks cell cycle, promotes cell apoptosis, and suppresses tumor growth of bladder cancer by directly dephosphorylating Akt at Ser473 site. PLoS ONE 2013, 8, e65679. [Google Scholar] [CrossRef]
- Risso, G.; Blaustein, M.; Pozzi, B.; Mammi, P.; Srebrow, A. Akt/PKB: One kinase, many modifications. Biochem. J. 2015, 468, 203–214. [Google Scholar] [CrossRef]
- Wang, S.; Huang, X.; Sun, D.; Xin, X.; Pan, Q.; Peng, S.; Liang, Z.; Luo, C.; Yang, Y.; Jiang, H.; et al. Extensive crosstalk between O-GlcNAcylation and phosphorylation regulates Akt signaling. PLoS ONE 2012, 7, e37427. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.S.; Han, D.; Park, J.; Kwak, T.K.; Oh, M.A.; Lee, S.A.; Choi, S.; Park, Z.Y.; Kim, Y.; Lee, J.W. O-GlcNAc modulation at Akt1 Ser473 correlates with apoptosis of murine pancreatic beta cells. Exp. Cell Res. 2008, 314, 2238–2248. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.L.; Luo, A.; Grill, A.; Lao, T.; Zou, Y.; Chen, Y. Fluorescent Detection of O-GlcNAc via Tandem Glycan Labeling. Bioconjugate Chem. 2020, 31, 2098–2102. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Tatge, T.J.; Grill, A.E.; Zou, Y. Detecting and Imaging O-GlcNAc Sites Using Glycosyltransferases: A Systematic Approach to Study O-GlcNAc. Cell Chem. Biol. 2018, 25, 1428–1435.e1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biwi, J.; Clarisse, C.; Biot, C.; Kozak, R.P.; Madunic, K.; Mortuaire, M.; Wuhrer, M.; Spencer, D.I.; Schulz, C.; Guerardel, Y.; et al. OGT Controls the Expression and the Glycosylation of E-cadherin, and Affects Glycosphingolipid Structures in Human Colon Cell Lines. Proteomics 2019, 19, e1800452. [Google Scholar] [CrossRef]
- Crespo, H.J.; Lau, J.T.; Videira, P.A. Dendritic cells: A spot on sialic Acid. Front. Immunol. 2013, 4, 491. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Gagneux, P. Multifarious roles of sialic acids in immunity. Ann. N. Y. Acad. Sci. 2012, 1253, 16–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varki, A. Since there are PAMPs and DAMPs, there must be SAMPs? Glycan “self-associated molecular patterns” dampen innate immunity, but pathogens can mimic them. Glycobiology 2011, 21, 1121–1124. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.; Stanczak, M.; Mantuano, N.; Xiao, H.; Pijnenborg, J.; Malaker, S.; Miller, C.; Weidenbacher, P.; Tanzo, J.; Ahn, G.; et al. Targeted glycan degradation potentiates the anticancer immune response in vivo. Nat. Chem. Biol. 2020, 16, 1376–1384. [Google Scholar] [CrossRef]
- van de Wall, S.; Santegoets, K.; van Houtum, E.; Bull, C.; Adema, G. Sialoglycans and Siglecs Can Shape the Tumor Immune Microenvironment. Trends Immunol. 2020, 41, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Sanford, B. An alteration in tumor histocompatibility induced by neuraminidase. Transplantation 1967, 5, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Rosato, F.E.; Miller, E.; Rosato, E.F.; Brown, A.; Wallack, M.K.; Johnson, J.; Moskowitz, A. Active specific immunotherapy of human solid tumors. Ann. N. Y. Acad. Sci. 1976, 277, 332–338. [Google Scholar] [CrossRef]
- Napoletano, C.; Steentoff, C.; Battisti, F.; Ye, Z.; Rahimi, H.; Zizzari, I.; Dionisi, M.; Cerbelli, B.; Tomao, F.; French, D.; et al. Investigating Patterns of Immune Interaction in Ovarian Cancer: Probing the O-glycoproteome by the Macrophage Galactose-Like C-type Lectin (MGL). Cancers 2020, 12, 2841. [Google Scholar] [CrossRef]
- Läubli, H.; Varki, A. Sialic acid-binding immunoglobulin-like lectins (Siglecs) detect self-associated molecular patterns to regulate immune responses. Cell Mol. Life Sci. 2020, 77, 593–605. [Google Scholar] [CrossRef]
- Kim, C.-H. CD33 and CD33-Related Siglecs in Pathogen Recognition and Endocytosis of DC in the Innate Immune System. In Glycobiology of Innate Immunology; Kim, C.-H., Ed.; Springer: Singapore, 2022; pp. 631–656. [Google Scholar] [CrossRef]
- Hiltbold, E.M.; Vlad, A.M.; Ciborowski, P.; Watkins, S.C.; Finn, O.J. The mechanism of unresponsiveness to circulating tumor antigen MUC1 is a block in intracellular sorting and processing by dendritic cells. J. Immunol. 2000, 165, 3730–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Housseau, F.; Moorthy, A.; Langer, D.A.; Robbins, P.F.; Gonzales, M.I.; Topalian, S.L. N-linked carbohydrates in tyrosinase are required for its recognition by human MHC class II-restricted CD4+ T cells. Eur. J. Immunol. 2001, 31, 2690–2701. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. USA 2012, 109, 261–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, B.; Piksa, M.; Beley, I.; Patoux, A.; Besson, T.; Cordier, V.; Voedisch, B.; Schindler, P.; Stollner, D.; Perrot, L.; et al. Therapeutic antibody glycosylation impacts antigen recognition and immunogenicity. Immunology 2022, 166, 380–407. [Google Scholar] [CrossRef]
- Boligan, K.F.; Mesa, C.; Fernandez, L.E.; von Gunten, S. Cancer intelligence acquired (CIA): Tumor glycosylation and sialylation codes dismantling antitumor defense. Cell Mol. Life Sci. 2015, 72, 1231–1248. [Google Scholar] [CrossRef]
- Cohen, M.; Elkabets, M.; Perlmutter, M.; Porgador, A.; Voronov, E.; Apte, R.N.; Lichtenstein, R.G. Sialylation of 3-methylcholanthrene-induced fibrosarcoma determines antitumor immune responses during immunoediting. J. Immunol. 2010, 185, 5869–5878. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Geng, Z.; Shao, W.; Liu, E.; Zhang, J.; Tang, J.; Wang, P.; Sun, X.; Xiao, L.; Xu, W.; et al. Cancer-derived sialylated IgG promotes tumor immune escape by binding to Siglecs on effector T cells. Cell. Mol. Immunol. 2020, 17, 1148–1162. [Google Scholar] [CrossRef]
- Li, M.; Zheng, H.; Duan, Z.; Liu, H.; Hu, D.; Bode, A.; Dong, Z.; Cao, Y. Promotion of cell proliferation and inhibition of ADCC by cancerous immunoglobulin expressed in cancer cell lines. Cell. Mol. Immunol. 2012, 9, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Carrascal, M.A.; Severino, P.F.; Guadalupe Cabral, M.; Silva, M.; Ferreira, J.A.; Calais, F.; Quinto, H.; Pen, C.; Ligeiro, D.; Santos, L.L.; et al. Sialyl Tn-expressing bladder cancer cells induce a tolerogenic phenotype in innate and adaptive immune cells. Mol. Oncol. 2014, 8, 753–765. [Google Scholar] [CrossRef]
- Perdicchio, M.; Cornelissen, L.A.; Streng-Ouwehand, I.; Engels, S.; Verstege, M.I.; Boon, L.; Geerts, D.; van Kooyk, Y.; Unger, W.W. Tumor sialylation impedes T cell mediated anti-tumor responses while promoting tumor associated-regulatory T cells. Oncotarget 2016, 7, 8771–8782. [Google Scholar] [CrossRef]
- Caldwell, S.; Heitger, A.; Shen, W.; Liu, Y.; Taylor, B.; Ladisch, S. Mechanisms of ganglioside inhibition of APC function. J. Immunol. 2003, 171, 1676–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manni, M.; Laubli, H. Targeting glyco-immune checkpoints for cancer therapy. Expert Opin. Biol. Ther. 2021, 21, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Bartish, M.; Rincón, S.V.d.; Rudd, C.E.; Saragovi, H.U. Aiming for the Sweet Spot: Glyco-Immune Checkpoints and γδ T Cells in Targeted Immunotherapy. Front. Immunol. 2020, 11, 564499. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.; Stanczak, M.A.; Xiao, H.; Pijnenborg, J.F.; Mantuano, N.R.; Malaker, S.A.; Weidenbacher, P.A.; Miller, C.L.; Tanzo, J.T.; Ahn, G.; et al. Targeted Desialylation Overcomes Glyco-Immune Checkpoints and Potentiates the Anticancer Immune Response in Vivo. ChemRxiv. Camb. Camb. Open Engag. 2019, preprint. [Google Scholar]
- Bärenwaldt, A.; Läubli, H. The sialoglycan-Siglec glyco-immune checkpoint-a target for improving innate and adaptive anti-cancer immunity. Expert Opin. Ther. Targets 2019, 23, 839–853. [Google Scholar] [CrossRef]
- Rodriguez, E.; Boelaars, K.; Brown, K.; Eveline Li, R.J.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic acids in pancreatic cancer cells drive tumour-associated macrophage differentiation via the Siglec receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1270. [Google Scholar] [CrossRef]
- Beatson, R.; Graham, R.; Grundland Freile, F.; Cozzetto, D.; Kannambath, S.; Pfeifer, E.; Woodman, N.; Owen, J.; Nuamah, R.; Mandel, U.; et al. Cancer-associated hypersialylated MUC1 drives the differentiation of human monocytes into macrophages with a pathogenic phenotype. Commun. Biol. 2020, 3, 644. [Google Scholar] [CrossRef]
- Rosenstock, P.; Kaufmann, T. Sialic Acids and Their Influence on Human NK Cell Function. Cells 2021, 10, 263. [Google Scholar] [CrossRef]
- Hudak, J.E.; Canham, S.M.; Bertozzi, C.R. Glycocalyx engineering reveals a Siglec-based mechanism for NK cell immunoevasion. Nat. Chem. Biol. 2014, 10, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Daly, J.; Sarkar, S.; Natoni, A.; Stark, J.C.; Riley, N.M.; Bertozzi, C.R.; Carlsten, M.; O’Dwyer, M.E. Targeting hypersialylation in multiple myeloma represents a novel approach to enhance NK cell-mediated tumor responses. Blood. Adv. 2022, 6, 3352–3366. [Google Scholar] [CrossRef] [PubMed]
- Haas, Q.; Boligan, K.; Jandus, C.; Schneider, C.; Simillion, C.; Stanczak, M.; Haubitz, M.; Jafari, S.; Zippelius, A.; Baerlocher, G.; et al. Siglec-9 regulates an effector memory CD8+ T-cell subset that congregates in the melanoma tumor microenvironment. Cancer Immunol. Res. 2019, 7, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Ho, M.; Adeniji, O.; Giron, L.; Bordoloi, D.; Kulkarni, A.; Puchalt, A.; Abdel-Mohsen, M.; Muthumani, K. Development of Siglec-9 Blocking Antibody to Enhance Anti-Tumor Immunity. Front. Oncol. 2021, 11, 778989. [Google Scholar] [CrossRef] [PubMed]
- Vuchkovska, A.; Glanville, D.G.; Scurti, G.M.; Nishimura, M.I.; White, P.; Ulijasz, A.T.; Iwashima, M. Siglec-5 is an inhibitory immune checkpoint molecule for human T cells. Immunology 2022, 166, 238–248. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; et al. Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nat. Med. 2019, 25, 656–666. [Google Scholar] [CrossRef]
- Stanczak, M.A.; Siddiqui, S.S.; Trefny, M.P.; Thommen, D.S.; Boligan, K.F.; von Gunten, S.; Tzankov, A.; Tietze, L.; Lardinois, D.; Heinzelmann-Schwarz, V.; et al. Self-associated molecular patterns mediate cancer immune evasion by engaging Siglecs on T cells. J. Clin. Investig. 2018, 128, 4912–4923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.E.; de Haas, A.; Rodriguez, E.; Kalay, H.; Zaal, A.; Jimenez, C.R.; Piersma, S.R.; Pham, T.V.; Henneman, A.A.; de Goeij-de Haas, R.R.; et al. Quantitative Phosphoproteomic Analysis Reveals Dendritic Cell- Specific STAT Signaling After alpha2-3-Linked Sialic Acid Ligand Binding. Front. Immunol. 2021, 12, 673454. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Guo, Z.; Liu, Y.; Li, X.; Zhang, Q.; Xu, X.; Gu, Y.; Zhang, Y.; Zhao, D.; Cao, X. The lectin Siglec-G inhibits dendritic cell cross-presentation by impairing MHC class I-peptide complex formation. Nat. Immunol. 2016, 17, 1167–1175. [Google Scholar] [CrossRef]
- Prager, M.D.; Baechtel, F.S.; Ribble, R.J.; Ludden, C.M.; Mehta, J.M. Immunological stimulation with modified lymphoma cells in a minimally responsive tumor-host system. Cancer Res. 1974, 34, 3203–3209. [Google Scholar]
- Rios, A.; Simmons, R. Active specific immunotherapy of minimal residual tumor: Excision plus neuraminidase-treated tumor cells. Int. J. Cancer 1974, 13, 71–81. [Google Scholar] [CrossRef]
- Watkins, E., Jr.; Fau-Gray, B.N.; Gray Bn Fau-Anderson, L.L.; Anderson Ll Fau-Baralt, O.L.; Baralt Ol Fau-Nebril, L.R.; Nebril Lr Fau-Waters, M.F.; Waters Mf Fau-Connery, C.K.; Connery, C.K. Neuraminidase-mediated augmentation of in vitro immune response of patients with solid tumors. Int. J. Cancer 1974, 14, 799–807. [Google Scholar] [CrossRef]
- Rios, A.; Simmons, R. Experimental cancer immunotherapy: Modification of tumor cells to increase immunogenicity. Ann. New York Acad. Sci. 1976, 276, 45–60. [Google Scholar] [CrossRef]
- Hakim, A.A. Modification of the immunologic properties of the cell surface. Immunol. Commun. 1978, 7, 25–39. [Google Scholar] [CrossRef]
- Bowles, W.H.D.; Gloster, T.M. Sialidase and Sialyltransferase Inhibitors: Targeting Pathogenicity and Disease. Front. Mol. Biosci. 2021, 8, 705133. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Wu, L.; Sun, X.L. Sialyltransferase inhibition and recent advances. Biochim. Biophys. Acta 2016, 1864, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Moons, S.J.; Rossing, E.; Janssen, M.; Heise, T.; Bull, C.; Adema, G.J.; Boltje, T.J. Structure-Activity Relationship of Metabolic Sialic Acid Inhibitors and Labeling Reagents. ACS Chem. Biol. 2022, 17, 590–597. [Google Scholar] [CrossRef]
- Büll, C.; Boltje, T.; Balneger, N.; Weischer, S.; Wassink, M.; van Gemst, J.; Bloemendal, V.; Boon, L.; van der Vlag, J.; Heise, T.; et al. Sialic acid blockade suppresses tumor growth by enhancing T cell-mediated tumor immunity. Cancer Res. 2018, 78, 3574–3588. [Google Scholar] [CrossRef] [Green Version]
- Heise, T.; Pijnenborg, J.F.A.; Bull, C.; van Hilten, N.; Kers-Rebel, E.D.; Balneger, N.; Elferink, H.; Adema, G.J.; Boltje, T.J. Potent Metabolic Sialylation Inhibitors Based on C-5-Modified Fluorinated Sialic Acids. J. Med. Chem. 2019, 62, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Macauley, M.S.; Arlian, B.M.; Rillahan, C.D.; Pang, P.C.; Bortell, N.; Marcondes, M.C.; Haslam, S.M.; Dell, A.; Paulson, J.C. Systemic blockade of sialylation in mice with a global inhibitor of sialyltransferases. J. Biol. Chem. 2014, 289, 35149–35158. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.C.; Lin, T.W.; Chang, W.W.; Wu, C.Y.; Lo, W.H.; Wang, P.H.; Tsai, Y.C. Soyasaponin-I-modified invasive behavior of cancer by changing cell surface sialic acids. Gynecol. Oncol. 2005, 96, 415–422. [Google Scholar] [CrossRef]
- Hunter, C.; Gao, Z.; Chen, H.-M.; Thompson, N.; Wakarchuk, W.; Nitz, M.; Withers, S.; Willis, L. Attenuation of polysialic acid biosynthesis in cells by the small molecule inhibitor 8-keto-sialic acid. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ahuja, S.; Cahill, J.; Hartfield, K.; Whorton, M.R. Inhibition of CMP-sialic acid transport by endogenous 5-methyl CMP. PLoS ONE 2021, 16, e0249905. [Google Scholar] [CrossRef] [PubMed]
- Muz, B.; Abdelghafer, A.; Markovic, M.; Yavner, J.; Melam, A.; Salama, N.N.; Azab, A.K. Targeting E-selectin to Tackle Cancer Using Uproleselan. Cancers 2021, 13, 335. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Chen, M.L.; Huang, H.L.; Lai, C.J.; Liu, C.H.; Chuu, C.P.; Lin, Y.H. Active Targeting of P-Selectin by Fucoidan Modulates the Molecular Profiling of Metastasis in Docetaxel-Resistant Prostate Cancer. Mar. Drugs 2022, 20, 542. [Google Scholar] [CrossRef]
- Cai, Z.; Yan, Y.; Zhou, J.; Yang, Y.; Zhang, Y.; Chen, J. Multifunctionalized Brush-Like Glycopolymers with High Affinity to P-Selectin and Antitumor Metastasis Activity. Biomacromolecules 2021, 22, 1177–1185. [Google Scholar] [CrossRef]
- Ibarlucea-Benitez, I.; Weitzenfeld, P.; Smith, P.; Ravetch, J.V. Siglecs-7/9 function as inhibitory immune checkpoints in vivo and can be targeted to enhance therapeutic antitumor immunity. Proc. Natl. Acad. Sci. USA 2021, 118, e2107424118. [Google Scholar] [CrossRef]
- He, F.; Wang, N.; Li, J.; He, L.; Yang, Z.; Lu, J.; Xiong, G.; Yu, C.; Wang, S. High affinity monoclonal antibody targeting Siglec-15 for cancer immunotherapy. J. Clin. Transl. Res. 2021, 7, 739–749. [Google Scholar]
- Zhou, Z.; Wang, X.; Jiang, L.; Li, D.; Qian, R. Sialidase-Conjugated “NanoNiche” for Efficient Immune Checkpoint Blockade Therapy. ACS Appl. Bio Mater. 2021, 4, 5735–5741. [Google Scholar] [CrossRef]
- Sharma, M.; Lathers, D.; Johnson, M.; Luke, J.; Puzanov, I.; Curti, B.; Chen, C.; El-Khoueiry, A.; Henick, B.; Callahan, M.; et al. 772 A phase 1/2 dose escalation/expansion study evaluating the safety, pharmacokinetics, pharmacodynamics, and antitumor activity of E-602, a bi-sialidase fusion protein, in advanced cancer (GLIMMER-01). J. Immuno Ther. Cancer 2022, 10, A802. [Google Scholar] [CrossRef]
- Li, C.; Qiu, Q.; Gao, X.; Yan, X.; Fan, C.; Luo, X.; Liu, X.; Wang, S.; Lai, X.; Song, Y.; et al. Sialic acid conjugate-modified liposomal platform modulates immunosuppressive tumor microenvironment in multiple ways for improved immune checkpoint blockade therapy. J. Control Release 2021, 337, 393–406. [Google Scholar] [CrossRef]
- Meng, X.; Wang, M.; Zhang, K.; Sui, D.; Chen, M.; Xu, Z.; Guo, T.; Liu, X.; Deng, Y.; Song, Y. An Application of Tumor-Associated Macrophages as Immunotherapy Targets: Sialic Acid-Modified EPI-Loaded Liposomes Inhibit Breast Cancer Metastasis. AAPS PharmSciTech 2022, 23, 285. [Google Scholar] [CrossRef]
- Mantuano, N.; Natoli, M.; Zippelius, A.; Läubli, H. Tumor-associated carbohydrates and immunomodulatory lectins as targets for cancer immunotherapy. J. Immunother. Cancer 2020, 8, e001222. [Google Scholar] [CrossRef]
- Monzavi-Karbassi, B.; Pashov, A.; Kieber-Emmons, T. Tumor-Associated Glycans and Immune Surveillance. Vaccines 2013, 1, 174–203. [Google Scholar] [CrossRef] [Green Version]
- Cadena, A.; Cushman, T.; Welsh, J. Glycosylation and Antitumor Immunity. Int. Rev. Cell Mol. Biol. 2019, 343, 111–127. [Google Scholar] [CrossRef]
- O’Cearbhaill, R.E.; Ragupathi, G.; Zhu, J.; Wan, Q.; Mironov, S.; Yang, G.; Spassova, M.K.; Iasonos, A.; Kravetz, S.; Ouerfelli, O.; et al. A Phase I Study of Unimolecular Pentavalent (Globo-H-GM2-sTn-TF-Tn) Immunization of Patients with Epithelial Ovarian, Fallopian Tube, or Peritoneal Cancer in First Remission. Cancers 2016, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini, P.J.; Ragupathi, G.; Hood, C.; Aghajanian, C.A.; Juretzka, M.; Iasonos, A.; Hensley, M.L.; Spassova, M.K.; Ouerfelli, O.; Spriggs, D.R.; et al. Pilot study of a heptavalent vaccine-keyhole limpet hemocyanin conjugate plus QS21 in patients with epithelial ovarian, fallopian tube, or peritoneal cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 4170–4177. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Wang, Q.; Zhang, J.; Wu, Q.; Guo, Z. Synthesis and Evaluation of Protein Conjugates of GM3 Derivatives Carrying Modified Sialic Acids as Highly Immunogenic Cancer Vaccine Candidates. Medchemcomm 2011, 2, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Guo, Z. Improving the Antigenicity of sTn Antigen by Modification of Its Sialic Acid Residue for Development of Glycoconjugate Cancer Vaccines. Bioconjugate Chem. 2006, 17, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Miles, D.; Papazisis, K. Rationale for the clinical development of STn-KLH (Theratope) and anti-MUC-1 vaccines in breast cancer. Clin. Breast Cancer 2003, 3 (Suppl. S4), S134–S138. [Google Scholar] [CrossRef]
- Huang, J.; Li, M.; Mei, B.; Li, J.; Zhu, Y.; Guo, Q.; Huang, J.; Zhang, G. Whole-cell tumor vaccines desialylated to uncover tumor antigenic Gal/GalNAc epitopes elicit anti-tumor immunity. J. Transl. Med. 2022, 20, 496. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Title | First Author (Year) | Source | Citations * |
|---|---|---|---|---|
| 1 | ST6Gal-I protein expression is upregulated in human epithelial tumors and correlates with stem cell markers in normal tissues and colon cancer cell lines [10] | Swindall AF et al. (2013) | Cancer Research | 52 |
| 2 | Sialylation of epidermal growth factor receptor regulates receptor activity and chemosensitivity to gefitinib in colon cancer cells [19] | Park JJ et al. (2012) | Biochemical Pharmacology | 49 |
| 3 | Phenylboronic acid-installed polymeric micelles for targeting sialylated epitopes in solid tumors [21] | Deshayes S et al. (2013) | Journal of the American Chemical Society | 47 |
| 4 | Interactions between Siglec-7/9 receptors and ligands influence NK cell-dependent tumor immunosurveillance [15] | Jandus C et al. (2014) | The Journal of Clinical Investigation | 39 |
| 5 | High-throughput profiling of protein N-glycosylation by MALDI-TOF-MS employing linkage-specific sialic acid esterification [17] | Reiding KR et al. (2014) | Analytical Chemistry | 37 |
| 6 | Targeting aberrant sialylation in cancer cells using a fluorinated sialic acid analog impairs adhesion, migration, and in vivo tumor growth [11] | Bull C et al. (2013) | Molecular Cancer Therapeutics | 34 |
| 7 | ST6Gal-I sialyltransferase confers cisplatin resistance in ovarian tumor cells [20] | Schultz MJ et al. (2013) | Journal of Ovarian Research | 32 |
| 8 | Acute myeloid leukemia stem cells and CD33-targeted immunotherapy [16] | Walter RB et al. (2012) | Blood | 31 |
| 9 | Anti-CD22-chimeric antigen receptors targeting B-cell precursor acute lymphoblastic leukemia [22] | Haso W et al. (2013) | Blood | 30 |
| 10 | N-linked glycan structures and their expressions change in the blood sera of ovarian cancer patients [18] | Alley WR et al. (2012) | Journal of Proteome Research | 29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Huang, J.; Zhang, G. Insights into the Role of Sialylation in Cancer Metastasis, Immunity, and Therapeutic Opportunity. Cancers 2022, 14, 5840. https://doi.org/10.3390/cancers14235840
Huang J, Huang J, Zhang G. Insights into the Role of Sialylation in Cancer Metastasis, Immunity, and Therapeutic Opportunity. Cancers. 2022; 14(23):5840. https://doi.org/10.3390/cancers14235840
Chicago/Turabian StyleHuang, Jianmei, Jianming Huang, and Guonan Zhang. 2022. "Insights into the Role of Sialylation in Cancer Metastasis, Immunity, and Therapeutic Opportunity" Cancers 14, no. 23: 5840. https://doi.org/10.3390/cancers14235840
APA StyleHuang, J., Huang, J., & Zhang, G. (2022). Insights into the Role of Sialylation in Cancer Metastasis, Immunity, and Therapeutic Opportunity. Cancers, 14(23), 5840. https://doi.org/10.3390/cancers14235840