

Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon?




Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Diabetes
1.2. Gastrointestinal Cancer
1.3. Flavonoids Influence Diabetes and GI Cancer
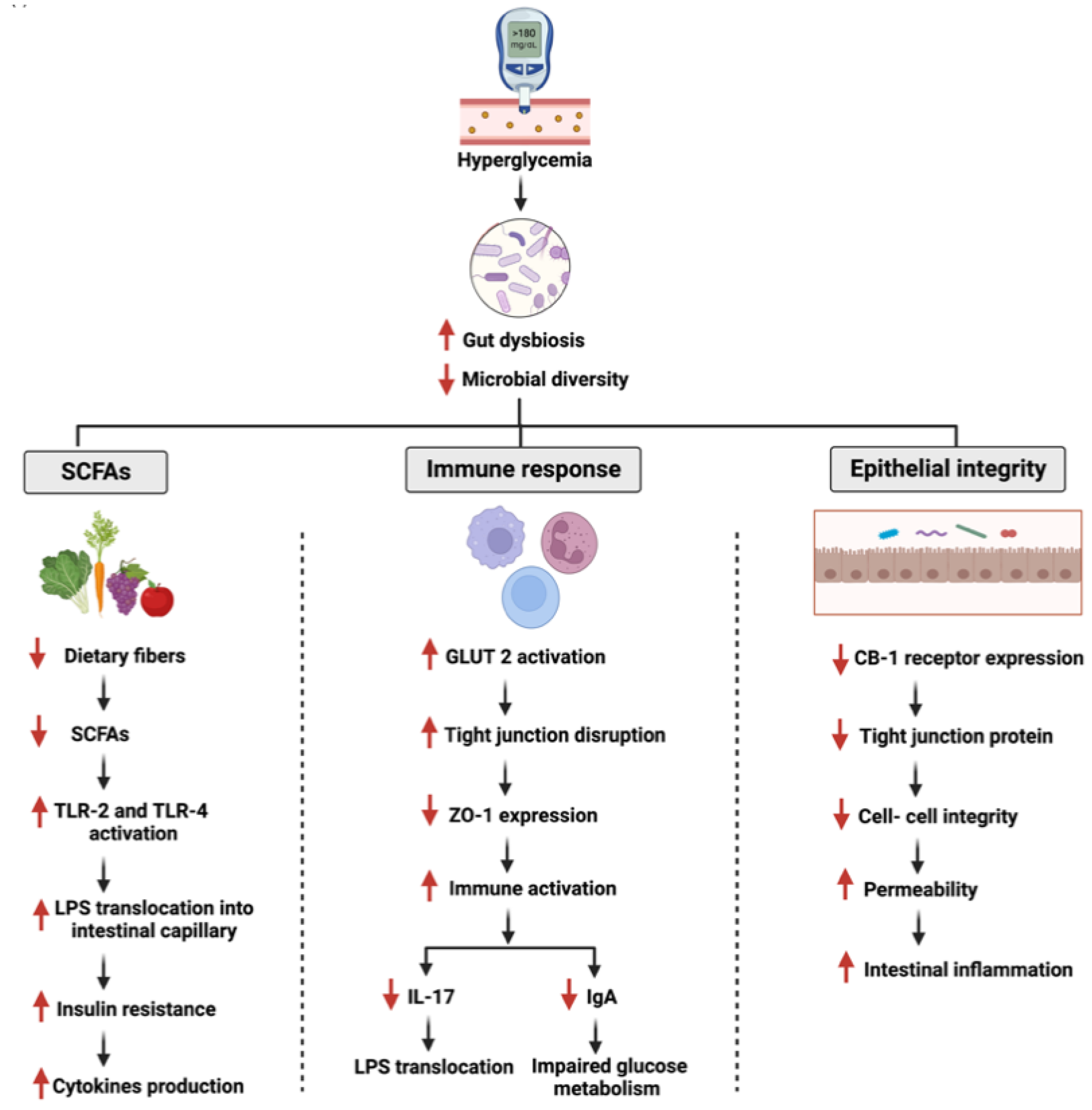
1.4. Gut Microbiota: The Role in Diabetes and GI Cancer
2. Search Strategy and Selection Criteria
3. Role of Flavonoids on Diabetic and GI Cancer Pathways
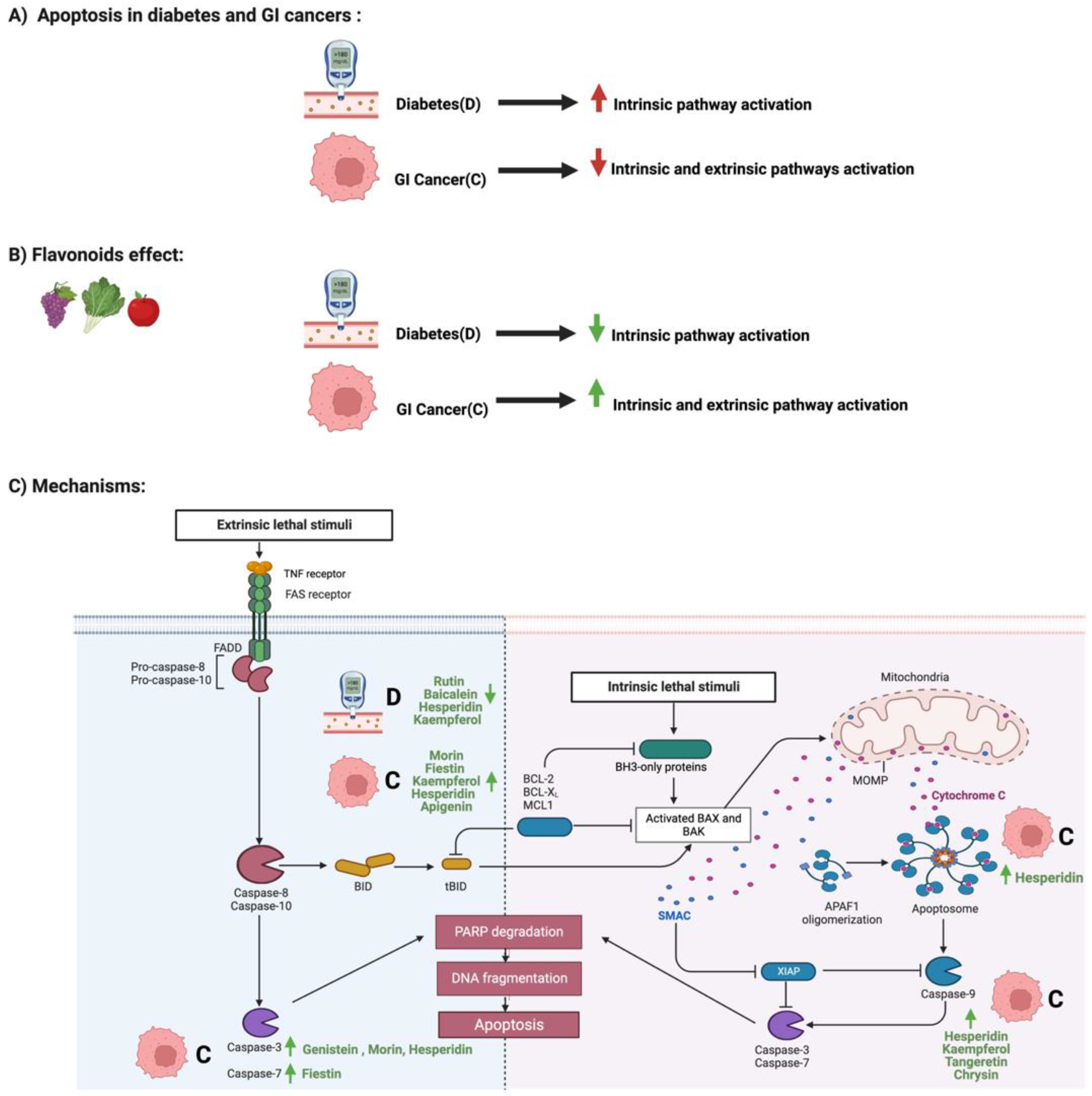
3.1. Flavonoids and Apoptosis in Diabetes and GI Cancers
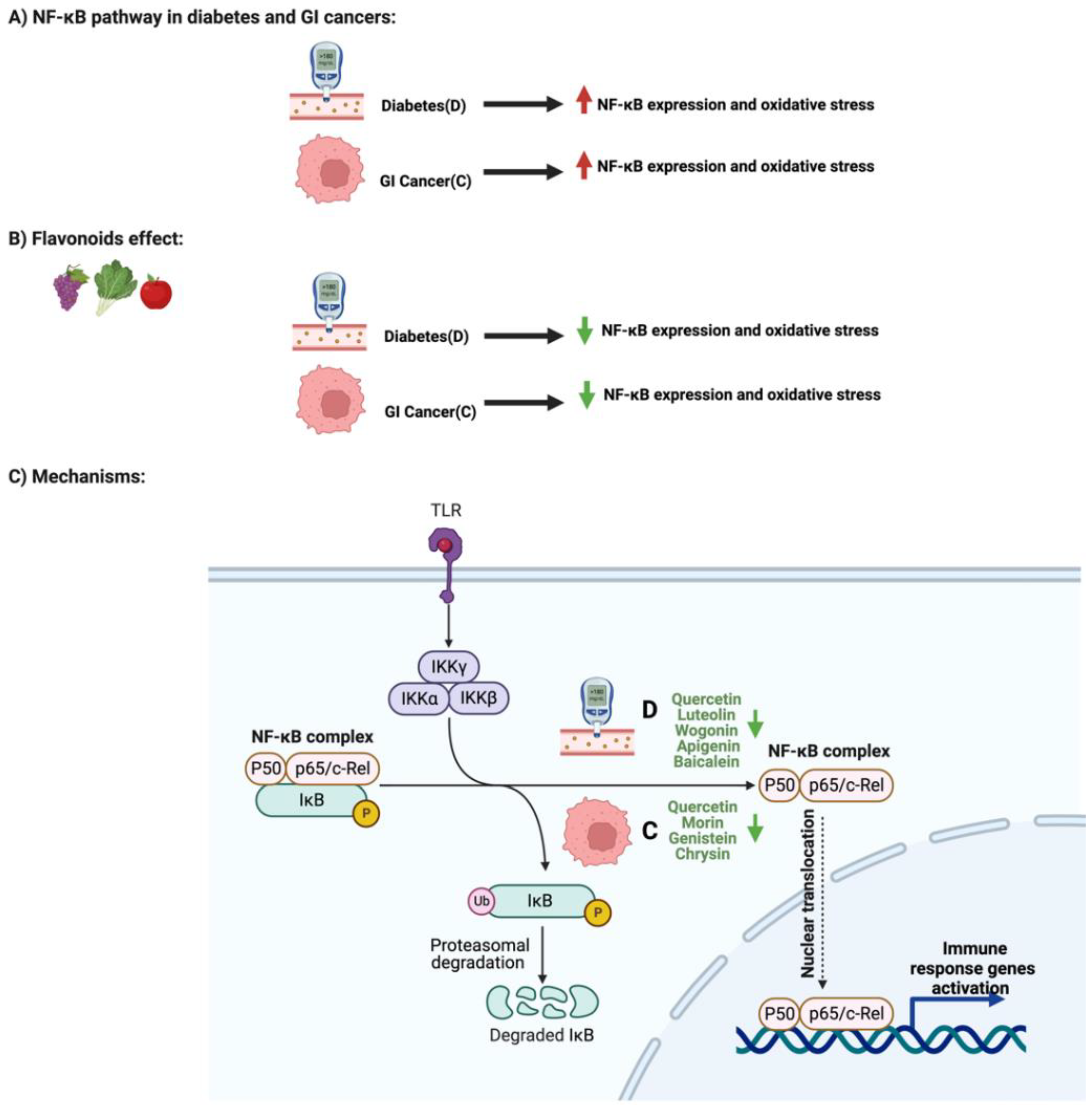
3.2. Flavonoids and NF-κB Pathway in Diabetes and GI Cancers
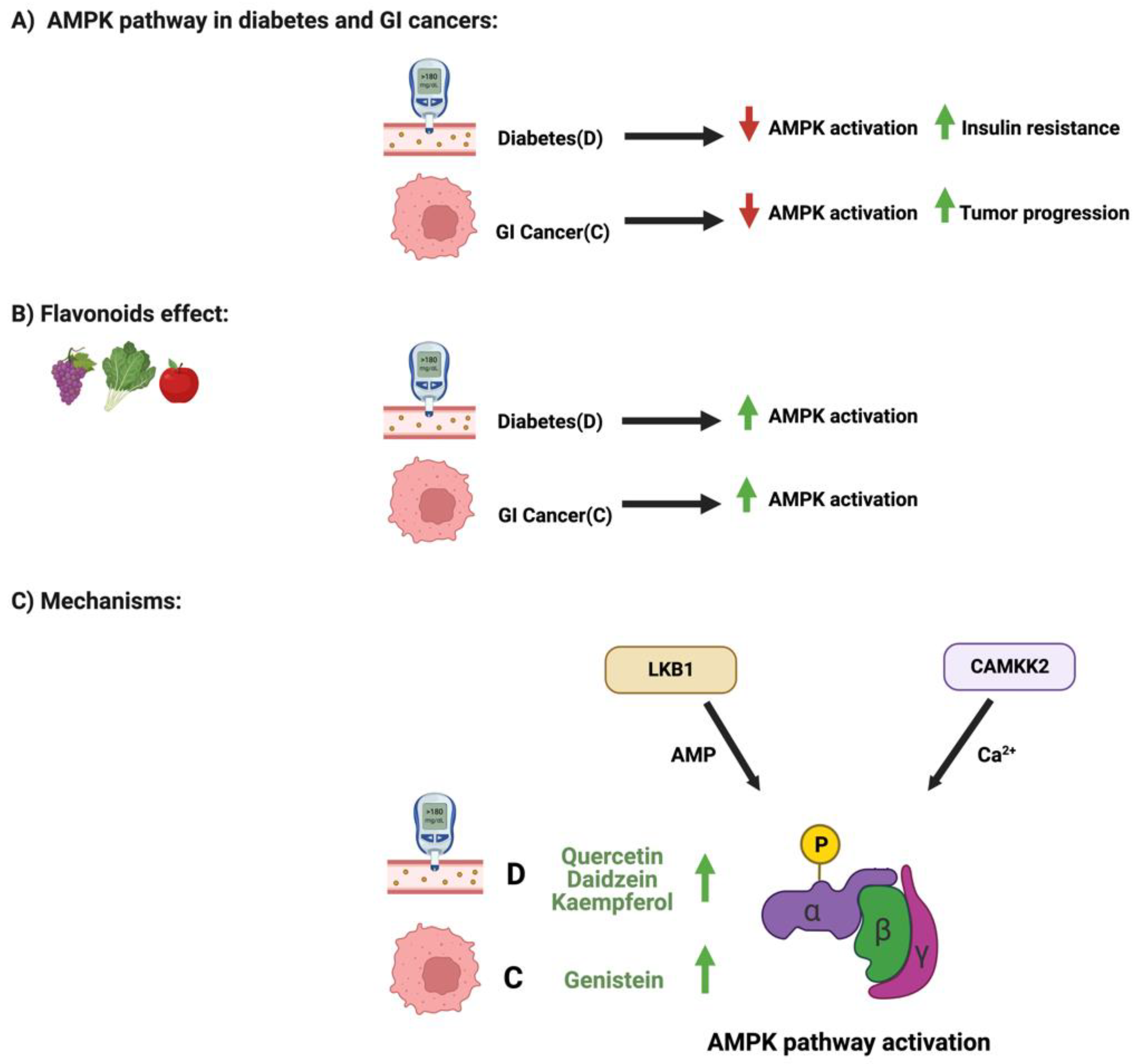
3.3. Flavonoids and AMPK Pathway in Diabetes and GI Cancers
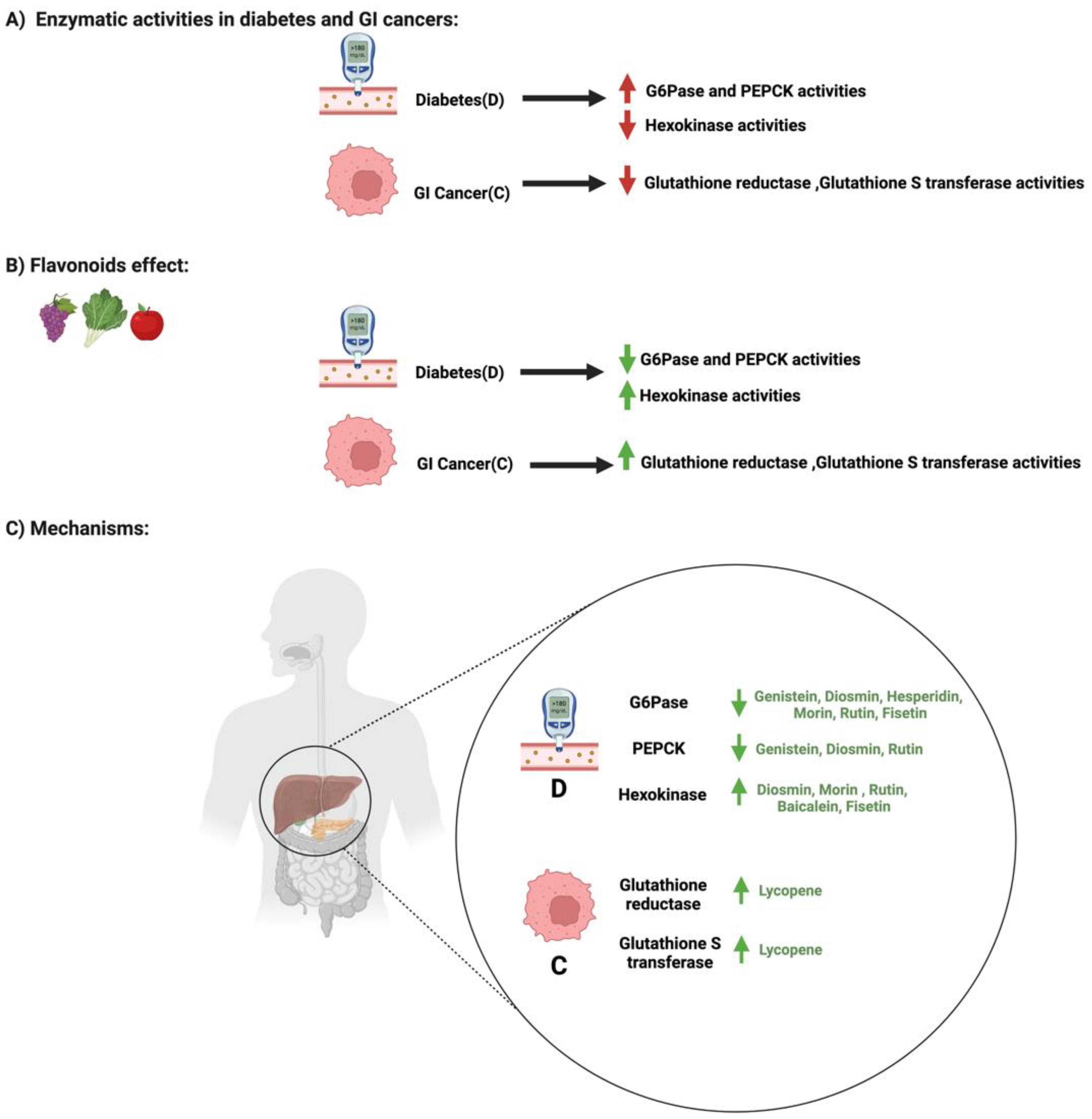
3.4. Flavonoids and Enzymatic Activities in Diabetes and GI Cancers
4. Possible Dual Effects of Flavonoids on Diabetes and GI Cancer
5. Discussion
5.1. Targeting Multiple Pathways with Multiple Flavonoids That Act Synergistically?
5.2. Gut Microbiome and Flavonoids’ Effects
5.3. Challenges with Studying the Field
5.4. GI Cancers and Diabetes: Their Relationship?
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GI | Gastrointestinal |
| CRC | Colorectal cancer |
| GC | Gastric cancer |
| IBD | Inflammatory bowel disease |
| SCFA | Short chain fatty acid |
| TLR | Toll-like receptor |
| LPS | Lipopolysaccharide |
| GLUT | Glucose transporter |
| AMPK | AMP-activated protein kinase |
| NF-κB | Nuclear factor kappa |
| ROS | Reactive oxygen species |
| EMT | Epithelial-mesenchymal transition |
| FISH | Fluorescence in situ hybridization |
| COX-2 | Cyclooxygenase-2 |
| G6Pase | Glucose 6-phosphatase |
| PEPCK | Phosphoenolpyruvate carboxykinase |
| PCR | Polymerase chain reaction |
| IP | Immunoprecipitation assay |
References
- Patel, D.K.; Kumar, R.; Laloo, D.; Hemalatha, S. Diabetes mellitus: An overview on its pharmacological aspects and reported medicinal plants having antidiabetic activity. Asian Pac. J. Trop. Biomed. 2012, 2, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of diabetes: An overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef]
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus-present and future perspectives. Nat. Rev. Endocrinol. 2011, 8, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.; Tripp-Reimer, T.; Parker, E.; Muller, B.; Laroche, H. Factors Influencing Diabetes Self-Management Among Medically Underserved Patients with Type II Diabetes. Glob. Qual. Nurs. Res. 2017, 4, 2333393617713097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and macrovascular complications in diabetes mellitus: Distinct or continuum? Indian J. Endocrinol. Metab. 2016, 20, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Shan, P.F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A. Know the signs and symptoms of diabetes. Indian J. Med. Res. 2014, 140, 579–581. [Google Scholar]
- Dhatariya, K. Diabetes: The place of new therapies. Adv. Endocrinol. Metab. 2019, 10, 2042018818807599. [Google Scholar] [CrossRef]
- Blaslov, K.; Naranda, F.S.; Kruljac, I.; Renar, I.P. Treatment approach to type 2 diabetes: Past, present and future. World J. Diabetes 2018, 9, 209–219. [Google Scholar] [CrossRef]
- Hassanzade, J.; Molavi, E.V.H.; Farahmand, M.; Rajaiifard, A.R. Incidence and Mortality Rate of Common Gastrointestinal Cancers in South of Iran, a Population Based Study. Iran. J. Cancer Prev. 2011, 4, 163–169. [Google Scholar] [PubMed]
- Bordry, N.; Astaras, C.; Ongaro, M.; Goossens, N.; Frossard, J.L.; Koessler, T. Recent advances in gastrointestinal cancers. World J. Gastroenterol. 2021, 27, 4493–4503. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Busselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zali, H.; Rezaei-Tavirani, M.; Azodi, M. Gastric cancer: Prevention, risk factors and treatment. Gastroenterol. Hepatol. Bed Bench. 2011, 4, 175–185. [Google Scholar]
- Han, C.J.; Reding, K.; Cooper, B.A.; Paul, S.M.; Conley, Y.P.; Hammer, M.; Miaskowski, C. Symptom Clusters in PatientsWith Gastrointestinal Cancers Using Different Dimensions of the Symptom Experience. J. Pain Symptom Manag. 2019, 58, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef]
- Correa, P. Gastric cancer: Overview. Gastroenterol. Clin. N. Am. 2020, 42, 211–217. [Google Scholar]
- Matsuoka, T.; Yashiro, M. Precision medicine for gastrointestinal cancer: Recent progress and future perspective. World J. Gastrointest Oncol. 2020, 12, 1–20. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Kozlowska, A.; Szostak-Wegierek, D. Flavonoids—food sources and health benefits. Rocz. Panstw. Zakl. Hig. 2014, 65, 79–85. [Google Scholar]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Juca, M.M.; Cysne Filho, F.M.S.; de Almeida, J.C.; Mesquita, D.D.S.; Barriga, J.R.M.; Dias, K.C.F.; Vasconcelos, S.M.M. Flavonoids: Biological activities and therapeutic potential. Nat. Prod. Res. 2020, 34, 692–705. [Google Scholar] [CrossRef] [PubMed]
- Sok Yen, F.; Shu Qin, C.; Tan Shi Xuan, S.; Jia Ying, P.; Yi Le, H.; Darmarajan, T.; Salvamani, S. Hypoglycemic Effects of Plant Flavonoids: A Review. Evid Based Complement. Altern. Med. 2021, 2057333. [Google Scholar] [CrossRef]
- Testa, R.; Bonfigli, A.R.; Genovese, S.; De Nigris, V.; Ceriello, A. The Possible Role of Flavonoids in the Prevention of Diabetic Complications. Nutrients 2016, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, C.; Sanchez-Quesada, C.; Gaforio, J.J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitelli Storelli, F.; Molina, A.J.; Zamora-Ros, R.; Fernandez-Villa, T.; Roussou, V.; Romaguera, D.; Martin, V. Flavonoids and the Risk of Gastric Cancer: An Exploratory Case-Control Study in the MCC-Spain Study. Nutrients 2019, 11, 967. [Google Scholar] [CrossRef] [Green Version]
- Bull, M.J.; Plummer, N.T. Part 1: The Human Gut Microbiome in Health and Disease. Integr. Med. (Encinitas) 2014, 13, 17–22. [Google Scholar]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Liskova, A.; Kubatka, P.; Busselberg, D. Enzymatic Metabolism of Flavonoids by Gut Microbiota and Its Impact on Gastrointestinal Cancer. Cancers 2021, 13, 3934. [Google Scholar] [CrossRef]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Pedersen, O. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Radjabzadeh, D.; Chen, L.; Kurilshikov, A.; Kavousi, M.; Ahmadizar, F.; Voortman, T. Association of Insulin Resistance and Type 2 Diabetes With Gut Microbial Diversity: A Microbiome-Wide Analysis From Population Studies. JAMA Netw. Open 2021, 4, e2118811. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.Y.; Wu, C.Y.; Yu, J. The role of gut microbiota in cancer treatment: Friend or foe? Gut 2020, 69, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Busselberg, D. Flavonoids and Their Antidiabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar]
- Al-Ishaq, R.K.; Overy, A.J.; Busselberg, D. Phytochemicals and Gastrointestinal Cancer: Cellular Mechanisms and Effects to Change Cancer Progression. Biomolecules 2020, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Aachoui, Y.; Sagulenko, V.; Miao, E.A.; Stacey, K.J. Inflammasome-mediated pyroptotic and apoptotic cell death, and defense against infection. Curr. Opin. Microbiol. 2013, 16, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Eid, A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed. Pharm. 2022, 146, 112442. [Google Scholar] [CrossRef]
- Naudi, A.; Jove, M.; Ayala, V.; Cassanye, A.; Serrano, J.; Gonzalo, H.; Pamplona, R. Cellular dysfunction in diabetes as maladaptive response to mitochondrial oxidative stress. Exp. Diabetes Res. 2012, 696215. [Google Scholar] [CrossRef] [Green Version]
- Pitocco, D.; Tesauro, M.; Alessandro, R.; Ghirlanda, G.; Cardillo, C. Oxidative stress in diabetes: Implications for vascular and other complications. Int. J. Mol. Sci. 2013, 14, 21525–21550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Mao, X.Y.; Li, X.L.; Ma, Y.; Qiao, Y.D.; Liu, Z.Q.; Cao, Y.G. Baicalin alleviates diabetes associated cognitive deficits via modulation of mitogen-activated protein kinase signaling, brain derived neurotrophic factor and apoptosis. Mol. Med. Rep. 2015, 12, 6377–6383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Xing, Y.; Sun, J.; Sun, W.; Xu, Y.; Quan, C. Baicalein Alleviates Liver Oxidative Stress and Apoptosis Induced by High-Level Glucose through the Activation of the PERK/Nrf2 Signaling Pathway. Molecules 2020, 25, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmar, M.S.; Syed, I.; Gray, J.P.; Ray, S.D. Curcumin, Hesperidin, and Rutin Selectively Interfere with Apoptosis Signaling and Attenuate Streptozotocin-Induced Oxidative Stress-Mediated Hyperglycemia. Curr. Neurovasc. Res. 2015, 12, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Han, Y.B.; Zhao, C.C.; Liu, L.; Zhang, F.L. Hesperidin alleviates insulin resistance by improving HG-induced oxidative stress and mitochondrial dysfunction by restoring miR-149. Diabetol. Metab. Syndr. 2021, 13, 50. [Google Scholar] [CrossRef]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharm. 2017, 96, 305–312. [Google Scholar] [CrossRef]
- Wang, J.; Wang, R.; Li, J.; Yao, Z. Rutin alleviates cardiomyocyte injury induced by high glucose through inhibiting apoptosis and endoplasmic reticulum stress. Exp. Med. 2021, 22, 944. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Maeda, Y.; Takahashi, H.; Nakai, N.; Yanagita, T.; Ando, N.; Okubo, T.; Takiguchi, S. Apigenin induces apoptosis by suppressing Bcl-xl and Mcl-1 simultaneously via signal transducer and activator of transcription 3 signaling in colon cancer. Int. J. Oncol. 2018, 52, 1661–1673. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Wang, C.; Hu, Z.; Chen, W.; Qi, W.; Li, A. Genistein induces apoptosis of colon cancer cells by reversal of epithelial-to-mesenchymal via a Notch1/NF-kappaB/slug/E-cadherin pathway. BMC Cancer 2017, 17, 813. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, S.Y.; Kim, M.; Cheon, C.; Ko, S.G. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis. 2018, 9, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahadori, M.; Baharara, J.; Amini, E. Anticancer Properties of Chrysin on Colon Cancer Cells, In vitro and In vivo with Modulation of Caspase-3, -9, Bax and Sall4. Iran. J. Biotechnol. 2016, 14, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sithara, T.; Arun, K.B.; Syama, H.P.; Reshmitha, T.R.; Nisha, P. Morin Inhibits Proliferation of SW480 Colorectal Cancer Cells by Inducing Apoptosis Mediated by Reactive Oxygen Species Formation and Uncoupling of Warburg Effect. Front. Pharm. 2017, 8, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wu, D.; Vikash, S.J.; Wang, J.; Yi, J.; Dong, W. Hesperetin Induces the Apoptosis of Gastric Cancer Cells via Activating Mitochondrial Pathway by Increasing Reactive Oxygen Species. Dig. Dis Sci. 2015, 60, 2985–2995. [Google Scholar] [CrossRef]
- Yang, C.; Song, J.; Hwang, S.; Choi, J.; Song, G.; Lim, W. Apigenin enhances apoptosis induction by 5-fluorouracil through regulation of thymidylate synthase in colorectal cancer cells. Redox Biol. 2021, 47, 102144. [Google Scholar] [CrossRef]
- Napetschnig, J.; Wu, H. Molecular basis of NF-kappaB signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef]
- Park, M.H.; Hong, J.T. Roles of NF-kappaB in Cancer and Inflammatory Diseases and Their Therapeutic Approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef]
- Paneni, F.; Beckman, J.A.; Creager, M.A.; Cosentino, F. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part I. Eur. Heart J. 2013, 34, 2436–2443. [Google Scholar] [CrossRef]
- Suryavanshi, S.V.; Kulkarni, Y.A. NF-kappabeta: A Potential Target in the Management of Vascular Complications of Diabetes. Front. Pharm. 2017, 8, 798. [Google Scholar] [CrossRef] [Green Version]
- Chaithongyot, S.; Jantaree, P.; Sokolova, O.; Naumann, M. NF-kappaB in Gastric Cancer Development and Therapy. Biomedicines 2021, 9, 870. [Google Scholar]
- Li, G.; Ding, K.; Qiao, Y.; Zhang, L.; Zheng, L.; Pan, T.; Zhang, L. Flavonoids Regulate Inflammation and Oxidative Stress in Cancer. Molecules 2020, 25, 5628. [Google Scholar] [CrossRef] [PubMed]
- Jubaidi, F.F.; Zainalabidin, S.; Taib, I.S.; Hamid, Z.A.; Budin, S.B. The Potential Role of Flavonoids in Ameliorating Diabetic Cardiomyopathy via Alleviation of Cardiac Oxidative Stress, Inflammation and Apoptosis. Int. J. Mol. Sci. 2021, 22, 5094. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Huang, L.; Ouyang, T.; Chen, L. Baicalein improves liver inflammation in diabetic db/db mice by regulating HMGB1/TLR4/NF-kappaB signaling pathway. Int. Immunopharmacol. 2018, 55, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Suchal, K.; Khan, S.I.; Bhatia, J.; Kishore, K.; Dinda, A.K.; Arya, D.S. Apigenin ameliorates streptozotocin-induced diabetic nephropathy in rats via MAPK-NF-kappaB-TNF-alpha and TGF-beta1-MAPK-fibronectin pathways. Am. J. Physiol. Ren. Physiol. 2017, 313, F414–F422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Luo, W.; Qian, Y.; Zhu, W.; Qian, J.; Li, J.; Liang, G. Luteolin protects against diabetic cardiomyopathy by inhibiting NF-kappaB-mediated inflammation and activating the Nrf2-mediated antioxidant responses. Phytomedicine 2019, 59, 152774. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.F.; Hassan, N.A.; El Bassossy, H.M.; Fahmy, A. Quercetin protects against diabetes-induced exaggerated vasoconstriction in rats: Effect on low grade inflammation. PLoS ONE 2013, 8, e63784. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lian, S.; Khoi, P.N.; Yoon, H.J.; Han, J.Y.; Chay, K.O.; Jung, Y.D. Chrysin inhibits cell invasion by inhibition of Recepteur d’origine Nantais via suppressing early growth response-1 and NF-kappaB transcription factor activities in gastric cancer cells. Int. J. Oncol. 2015, 46, 1835–1843. [Google Scholar] [CrossRef]
- Chen, R.; Zhang, L. Morin inhibits colorectal tumor growth through inhibition of NF-kappaB signaling pathway. Immunopharmacol. Immunotoxicol. 2019, 41, 622–629. [Google Scholar] [CrossRef]
- Li, Y.S.; Wu, L.P.; Li, K.H.; Liu, Y.P.; Xiang, R.; Zhang, S.B.; Zhang, L.Y. Involvement of nuclear factor kappaB (NF-kappaB) in the downregulation of cyclooxygenase-2 (COX-2) by genistein in gastric cancer cells. J. Int. Med. Res. 2011, 39, 2141–2150. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Chen, C. Quercetin Has Antimetastatic Effects on Gastric Cancer Cells via the Interruption of uPA/uPAR Function by Modulating NF-kappab, PKC-delta, ERK1/2, and AMPKalpha. Integr. Cancer 2018, 17, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Carling, D.; Gamblin, S.J. AMP-activated protein kinase: Also regulated by ADP? Trends Biochem. Sci. 2011, 36, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Song, P.; Zou, M.H. AMP-activated protein kinase, stress responses and cardiovascular diseases. Clin. Sci. 2012, 122, 555–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coughlan, K.A.; Valentine, R.J.; Ruderman, N.B.; Saha, A.K. AMPK activation: A therapeutic target for type 2 diabetes? Diabetes Metab. Syndr. Obes. 2014, 7, 241–253. [Google Scholar]
- Li, W.; Saud, S.M.; Young, M.R.; Chen, G.; Hua, B. Targeting AMPK for cancer prevention and treatment. Oncotarget 2015, 6, 7365–7378. [Google Scholar] [CrossRef] [Green Version]
- Dhanya, R.; Arya, A.D.; Nisha, P.; Jayamurthy, P. Quercetin, a Lead Compound against Type 2 Diabetes Ameliorates Glucose Uptake via AMPK Pathway in Skeletal Muscle Cell Line. Front. Pharm. 2017, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. J. Nutr. Biochem. 2014, 25, 136–143. [Google Scholar] [CrossRef]
- Varshney, R.; Gupta, S.; Roy, P. Cytoprotective effect of Kaempferol against palmitic acid-induced pancreatic beta-cell death through modulation of autophagy via AMPK/mTOR signaling pathway. Mol. Cell Endocrinol. 2017, 448, 1–20. [Google Scholar] [CrossRef]
- Talib, W.H.; Awajan, D.; Hamed, R.A.; Azzam, A.O.; Mahmod, A.I.; Al-Yasari, I.H. Combination Anticancer Therapies Using Selected Phytochemicals. Molecules 2022, 27, 5452. [Google Scholar] [CrossRef]
- Zallot, R.; Oberg, N.; Gerlt, J.A. Discovery of new enzymatic functions and metabolic pathways using genomic enzymology web tools. Curr. Opin. Biotechnol. 2021, 69, 77–90. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress-A concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, A.; Simon, M.C. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Thornalley, P.J. Hexokinase-2 Glycolytic Overload in Diabetes and Ischemia-Reperfusion Injury. Trends Endocrinol. Metab. 2019, 30, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.S.; Jung, U.J.; Yeo, J.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res. Rev. 2008, 24, 74–81. [Google Scholar] [CrossRef]
- Pari, L.; Srinivasan, S. Antihyperglycemic effect of diosmin on hepatic key enzymes of carbohydrate metabolism in streptozotocin-nicotinamide-induced diabetic rats. Biomed. Pharm. 2010, 64, 477–481. [Google Scholar] [CrossRef]
- Patel, S.S.; Shah, R.S.; Goyal, R.K. Antihyperglycemic, antihyperlipidemic and antioxidant effects of Dihar, a polyherbal ayurvedic formulation in streptozotocin induced diabetic rats. Indian J. Exp. Biol. 2009, 47, 564–570. [Google Scholar]
- Vanitha, P.; Uma, C.; Suganya, N.; Bhakkiyalakshmi, E.; Suriyanarayanan, S.; Gunasekaran, P.; Ramkumar, K.M. Modulatory effects of morin on hyperglycemia by attenuating the hepatic key enzymes of carbohydrate metabolism and beta-cell function in streptozotocin-induced diabetic rats. Environ. Toxicol. Pharm. 2014, 37, 326–335. [Google Scholar] [CrossRef]
- Hanschmann, E.M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, glutaredoxins, and peroxiredoxins--molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling. Antioxid Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef]
- Kennedy, L.; Sandhu, J.K.; Harper, M.E.; Cuperlovic-Culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Marzocco, S.; Singla, R.K.; Capasso, A. Multifaceted Effects of Lycopene: A Boulevard to the Multitarget-Based Treatment for Cancer. Molecules 2021, 26, 5333. [Google Scholar] [CrossRef] [PubMed]
- JZ, A.L.; BinMowyna, M.N.; AlFaris, N.A.; Alagal, R.I.; El-Kott, A.F.; Al-Farga, A.M. Fisetin protects against streptozotocin-induced diabetic cardiomyopathy in rats by suppressing fatty acid oxidation and inhibiting protein kinase R. Saudi Pharm. J. 2021, 29, 27–42. [Google Scholar]
- Akiyama, S.; Katsumata, S.; Suzuki, K.; Ishimi, Y.; Wu, J.; Uehara, M. Dietary hesperidin exerts hypoglycemic and hypolipidemic effects in streptozotocin-induced marginal type 1 diabetic rats. J. Clin. Biochem. Nutr. 2010, 46, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.T.; Wu, X.D.; Davey, A.K.; Wang, J. Antihyperglycemic effects of baicalin on streptozotocin-nicotinamide induced diabetic rats. Phytother. Res. 2011, 25, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.C.; Zhu, W.; Lei, L.; Liu, X.Q.; Wu, Y.G. Wogonin Ameliorates Renal Inflammation and Fibrosis by Inhibiting NF-kappaB and TGF-beta1/Smad3 Signaling Pathways in Diabetic Nephropathy. Drug Des. Devel. 2020, 14, 4135–4148. [Google Scholar] [CrossRef]
- Zhang, F.; Feng, J.; Zhang, J.; Kang, X.; Qian, D. Quercetin modulates AMPK/SIRT1/NF-kappaB signaling to inhibit inflammatory/oxidative stress responses in diabetic high fat diet-induced atherosclerosis in the rat carotid artery. Exp. Med. 2020, 20, 280. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Antihyperglycemic effect of equol, a daidzein derivative, in cultured L6 myocytes and ob/ob mice. Mol. Nutr. Food Res. 2014, 58, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, B.; Bhuvaneswari, V.; Nagini, S. Antiperoxidative effects of lycopene during N-methyl-N′-nitro-N-nitrosoguanidine-induced gastric carcinogenesis. Fitoterapia 2002, 73, 604–611. [Google Scholar] [CrossRef]
- Hwang, J.T.; Ha, J.; Park, O.J. Combination of 5-fluorouracil and genistein induces apoptosis synergistically in chemo-resistant cancer cells through the modulation of AMPK and COX-2 signaling pathways. Biochem. Biophys. Res. Commun 2005, 332, 433–440. [Google Scholar] [CrossRef]
- Fan, Q.; Wang, X.; Chinnathambi, A.; Alharbi, S.; Wang, Q. Fisetin suppresses 1,2-dimethylhydrazine-induced colon tumorigenesis in Wistar rats. J. King Saud Univ. Sci. 2020, 32, 1959–1964. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, M.J.; Ha, E.; Chung, J.H. Apoptotic effect of hesperidin through caspase3 activation in human colon cancer cells, SNU-C4. Phytomedicine 2008, 15, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Cho, H.J.; Yu, R.; Lee, K.W.; Chun, H.S.; Park, J.H. Mechanisms underlying apoptosis-inducing effects of Kaempferol in HT-29 human colon cancer cells. Int. J. Mol. Sci. 2014, 15, 2722–2737. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Cao, A.; Shi, J.; Yin, P.; Wang, L.; Ji, G.; Wu, D. Tangeretin, a citrus polymethoxyflavonoid, induces apoptosis of human gastric cancer AGS cells through extrinsic and intrinsic signaling pathways. Oncol. Rep. 2014, 31, 1788–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanchang, W.; Khamchan, A.; Wongmanee, N.; Seedadee, C. Hesperidin ameliorates pancreatic beta-cell dysfunction and apoptosis in streptozotocin-induced diabetic rat model. Life Sci. 2019, 235, 116858. [Google Scholar] [CrossRef] [PubMed]
- Riahi-Chebbi, I.; Souid, S.; Othman, H.; Haoues, M.; Karoui, H.; Morel, A.; Essafi-Benkhadir, K. The Phenolic compound Kaempferol overcomes 5-fluorouracil resistance in human resistant LS174 colon cancer cells. Sci. Rep. 2019, 9, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, D. Flavonol kaempferol improves chronic hyperglycemia-impaired pancreatic beta-cell viability and insulin secretory function. Eur. J. Pharm. 2011, 670, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.A.; Zhang, S.; Yin, Q.; Zhang, J. Quercetin induces human colon cancer cells apoptosis by inhibiting the nuclear factor-kappa B Pathway. Pharm. Mag. 2015, 11, 404–409. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gao, M.; Kang, G.; Huang, H. The Potential Role of Phytonutrients Flavonoids Influencing Gut Microbiota in the Prophylaxis and Treatment of Inflammatory Bowel Disease. Front. Nutr. 2021, 8, 798038. [Google Scholar] [CrossRef]
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159. [Google Scholar] [CrossRef]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid metabolism: The interaction of metabolites and gut microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Microbiota and diabetes: An evolving relationship. Gut 2014, 63, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Z.; Stirling, K.; Yang, J.J.; Zhang, L. Gut microbiota and diabetes: From correlation to causality and mechanism. World J. Diabetes 2020, 11, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.K. Potential Role of the Gut Microbiome In Colorectal Cancer Progression. Front. Immunol. 2021, 12, 807648. [Google Scholar] [CrossRef]
- Liu, Y.; Baba, Y.; Ishimoto, T.; Gu, X.; Zhang, J.; Nomoto, D.; Qiu, P. Gut microbiome in gastrointestinal cancer: A friend or foe? Int. J. Biol Sci. 2022, 18, 4101–4117. [Google Scholar] [CrossRef]
- Tremmel, M.; Paetz, C.; Heilmann, J. In Vitro Liver Metabolism of Six Flavonoid C-Glycosides. Molecules 2021, 26, 6632. [Google Scholar] [CrossRef]
- Wang, L.; Huang, G.; Hou, R.; Qi, D.; Wu, Q.; Nie, Y.; Wei, F. Multi-omics reveals the positive leverage of plant secondary metabolites on the gut microbiota in a non-model mammal. Microbiome 2021, 9, 192. [Google Scholar] [CrossRef]
- Sampson, L.; Rimm, E.; Hollman, P.C.; de Vries, J.H.; Katan, M.B. Flavonol and flavone intakes in US health professionals. J. Am. Diet. Assoc. 2002, 102, 1414–1420. [Google Scholar] [CrossRef]
- Boker, K.L.; Van der Schouw, Y.T.; De Kleijn, M.J.; Jacques, P.F.; Grobbee, D.E.; Peeters, P.H. Intake of Dietary Phytoestrogens by Dutch Women. J. Nutr. 2002, 132, 1319. [Google Scholar] [CrossRef] [Green Version]
- Arts, I.C.; Hollman, P.C.; Feskens, E.J.; Bueno de Mesquita, H.B.; Kromhout, D. Catechin intake and associated dietary and lifestyle factors in a representative sample of Dutch men and women. Eur. J. Clin. Nutr. 2001, 55, 76–81. [Google Scholar] [CrossRef]
- Rodriguez De Luna, S.L.; Ramirez-Garza, R.E.; Serna Saldivar, S.O. Environmentally Friendly Methods for Flavonoid Extraction from Plant Material: Impact of Their Operating Conditions on Yield and Antioxidant Properties. Sci. World J. 2020, 6792069. [Google Scholar] [CrossRef]
- Thilakarathna, S.H.; Rupasinghe, H.P. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Loosen, S.H.; Hoyer, L.; Luedde, T.; Kostev, K. Prevalence of diabetes mellitus among 80,193 gastrointestinal cancer patients in five European and three Asian countries. J. Cancer Res. Clin. Oncol. 2022, 148, 1057–1062. [Google Scholar] [CrossRef]
- Tseng, C.H. The Relationship between Diabetes Mellitus and Gastric Cancer and the Potential Benefits of Metformin: An Extensive Review of the Literature. Biomolecules 2021, 11, 1022. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.H.; Tseng, F.H. Diabetes and gastric cancer: The potential links. World J. Gastroenterol. 2014, 20, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Samec, M.; Liskova, A.; Koklesova, L.; Mersakova, S.; Strnadel, J.; Kajo, K.; Kubatka, P. Flavonoids Targeting HIF-1: Implications on Cancer Metabolism. Cancers 2021, 13, 559–587. [Google Scholar] [CrossRef]
- Kubatka, P.; Mazurakova, A.; Samec, M.; Koklesova, L.; Zhai, K.; Al-Ishaq, R.; Golubnitschaja, O. Flavonoids against non-physiologic inflammation attributed to cancer initiation, development, and progression-3PM pathways. EPMA J. 2021, 12, 559–587. [Google Scholar] [CrossRef]
- Zhang, S.; Jin, S.; Zhang, S.; Li, Y.Y.; Wang, H.; Chen, Y.; Lu, H. Vitexin protects against high glucose-induced endothelial cell apoptosis and oxidative stress via Wnt/beta-catenin and Nrf2 signalling pathway. Arch. Physiol. Biochem. 2022, 1–10. [Google Scholar] [CrossRef]
- Ling, J.Y.; Wang, Q.L.; Liang, H.N.; Liu, Q.B.; Yin, D.H.; Lin, L. Flavonoid-Rich Extract of Oldenlandia diffusa (Willd.) Roxb. Inhibits Gastric Cancer by Activation of Caspase-Dependent Mitochondrial Apoptosis. Chin. J. Integr. Med. 2022, 1–11. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Pathway | Name of Flavonoid | Flavonoid Subclass | Metabolites Produced by Gut Microbiota | Disease(s) | Influence of Flavonoids on Disease | Methods of Testing | Model Used | References | |
|---|---|---|---|---|---|---|---|---|---|
| In vivo | In vitro | ||||||||
| Enzymatic Modification | Rutin | Flavonol | Quercetin-3-O-glucoside Quercetin | Diabetes | * Enhanced the activity of liver hexokinase * Decreased the activities of glucose-6-phosphatase (G6Pase) | Oral glucose tolerance test (OGTT) Histology RNA extraction Reverse transcription PCR | * White male albino rats | * Tissues from the albino rats | [87] |
| Fisetin | Flavonol | No available data | Diabetes | * Inhibited hepatic gluconeogenesis * Decreased the activities of glucose-6-phosphatase (G6Pase) * Decreased the activities of Phosphoenolpyruvate carboxykinase (PEPCK) | Western blot analysis ELISA RNA extraction Reverse transcription PCR | * Male albino Wistar rats | * Liver tissues | [92] | |
| Morin | Flavonol | Morin glucuronides Morin sulfates | Diabetes | * Significantly reduced blood glucose * Morin treatment significantly reduced the G6Pase activities * Increased insulin levels and hexokinase activities | Intraperitoneal glucose tolerance test (IPGTT) Histopathological examination Immunohistochemical staining | * Male Albino Wistar rats | * Pancreatic tissue | [88] | |
| Hesperidin | Flavanones | Hesperetin | Diabetes | * Increased insulin levels * Hesperidin treatment normalized the G6Pase activities | Blood glucose meter Liquid chromatography ELISA | * Male Wistar rats | [93] | ||
| Genistein | Isoflavones | Dihydrogenistein 6-hydroxy-O-desmethylangolensin 2-(4-hydroxyphenyl) propionic acid | Diabetes | * Decreased the activities of glucose-6-phosphatase (G6Pase) * Decreased the activities of Phosphoenolpyruvate carboxykinase (PEPCK) | Intraperitoneal glucose tolerance test (IPGTT) Radioimmunoassay Immunohistochemistry | * Female NOD mice | * Pancreatic tissue *Liver tissues | [85] | |
| Baicalein | Flavones | Baicalein | Diabetes | * Hepatic hexokinase was significantly increased after the administration of 25, 50 or 10 mg/kg of baicalein | Histopathological examination ELISA | * Male Wistar rats | * Pancreatic tissue * Liver tissues | [94] | |
| Diosmin | Flavones | Diosmetin | Diabetes | * Decreased the activities of glucose-6-phosphatase (G6Pase) * Decreased the activities of Phosphoenolpyruvate carboxykinase (PEPCK) * Enhanced hexokinase activities | ELISA | * Male albino Wistar | [86] | ||
| NF-κB Pathway | Baicalein | Flavones | Baicalein | Diabetes | * Baicalein remarkably supressed the inflammatory cascade such as Myd88 and NF-κB thus inhibiting the production of interleukin (IL)-1β and IL-6 | Reverse transcription PCR Flow cytometry Western blot analysis | * C57BL/KsJ-db/db mice | * MPC-5 cells * HepG2 cells | [63] |
| Luteolin | Flavones | No available data | Diabetes | * Luteolin significantly inhibited the activation of nuclear factor-kappa B (NF-κB) pathway | Reverse transcription PCR Tissue histology Western blot analysis Cell viability | * Male C57BL/6 mice | * H9C2 cells | [65] | |
| Apigenin | Flavones | Apigenin 3-(4-hydroxyphenyl) propionic acid | Diabetes | * The intragastric administration of apigenin-SLNP supressed the expression of nuclear factor-kappa B (NF-κB) pathway * Protective effect of apigenin against diabetic nephropathy | Superoxide Dismutase (SOD) Reverse transcription PCR Histopathological Changes Western blot analysis | * Rats | * Kidney tissue | [64] | |
| Wogonin | Flavones | Wogonin | Diabetes | * The intragastric administration of wogonin (10, 20, and 40 mg/kg) for 13 days attenuated histological damages and regulated inflammation through nuclear factor-kappa B (NF-κB) pathway | Immunohistochemistry Assay RNA Isolation Reverse transcription PCR ELISA | * C57/BL male mice | * HK-2 cells | [95] | |
| Quercetin | Flavonol | 3,4-dihydroxyphenylacetic acid 3-(3-hydroxyphenyl) propionic acid 3,4-dihydroxybenzoic 4-hydroxybenzoic acid | Diabetes | * Daily administration of quercetin for six weeks showed inhibitory effects on inflammatory NF-κB signaling pathway | Immunohistochemistry Histopathological Examination | * Male albino rats | [66] | ||
| AMPK Pathway | Kaempferol | Flavonol | Kaempferol -3-O-glucoside p-coumaric acid kaempferol 3-(4 hydroxyphenyl) propionic acid 3-phenylpropionic acid | Diabetes | * Kaempferol illustrated cytoprotective effects during diabetes through the upregulation of AMPK phosphorylation * Kaempferol administration enhanced the metabolism of lipid through the activation of AMPK pathway | Cytotoxicity assays DNA ladder assay Electron microscopy Flow cytometry Reverse transcription PCR | * Female Sprague-Dawley rats | * RIN-5F cells | [78] |
| Quercetin | Flavonol | 3,4-dihydroxyphenylacetic acid 3-(3-hydroxyphenyl) propionic acid 3,4-dihydroxybenzoic 4-hydroxybenzoic acid | Diabetes | * Quercetin administration enhanced the AMPK activation and lipid metabolism | PCR Western blot analysis Immunofluorescence Assay | * Male Wistar rats | * L6 myoblast | [96] | |
| Daidzein | Isoflavones | Dihydrodaidzein O-desmethylangolensin S- equol | Diabetes | * Administration of daidzein markedly improved the AMPK phosphorylation | Protein isolation Western blot analysis PCR ELISA | * Male KK-Ay/Ta Jcl mice * C57BL/6J Jcl mice | * L6 myoblasts | [77,97] | |
| Apoptosis | Baicalein | Flavones | Baicalein | Diabetes | * The administration of baicalein significantly reduced oxidative stress and apoptosis in cells stimulated with glucose | Fluorescence microscopy Flow cytometry Cell viability assays Immunoblotting Quantitative PCR | * Male Wistar rats | * HL-7702 cells | [42,43] |
| Rutin | Flavonol | Quercetin-3-O-glucoside Quercetin | Diabetes | * Rutin administration to myoplastic cells indicated that rutin can, in a dose-dependent manner, reduce the level of endoplasmic reticulum stress and apoptosis | TUNEL assay Cell counting kit-8 assay Western blot analysis | * H9C2 myoblast cells | [47] | ||
| Hesperidin | Flavanones | Hesperetin | Diabetes | * Induced hyperglycemic mice administered with hesperidin downregulated antiapoptotic proteins such as Bcl-2 and Bcl-XL and upregulated the pro-apoptotic proteins | Western blot analysis Glutathione Assay Kit Liver histopathology | * Female ICR mice | [44] | ||
| Kaempferol | Flavonol | Kaempferol-3-O-glucoside p-coumaric acid kaempferol 3-(4 hydroxyphenyl) propionic acid 3-phenylpropionic acid | Diabetes | * Kaempferol treatment enhanced cellular viability and antiapoptotic activitites which suggested a cytoprotective role | Cytotoxicity assays Western blot analysis Flow cytometry | * Female Sprague-Dawley rats | * RIN-5F cells | [78] | |
| Targeted Pathway | Name of Flavonoid | Flavonoid Subclass | Metabolites Produced by Gut Microbiota | Disease(s) | Influence of Flavonoids on Disease | Methods of Testing | Model Used | References | |
|---|---|---|---|---|---|---|---|---|---|
| In vivo | In vitro | ||||||||
| Enzymatic Modification | Lycopene | Carotenoid | No available data | Cancer | * Lycopene administration could suppress gastric cancer through the reduction of lipid peroxidation and the enhancement of GSH-dependent enzymes such as glutathione reductase and glutathione-s-transferase | Histopathological examination | * Male Wistar rats | [98] | |
| NF-κB Pathway | Genistein | Isoflavones | Dihydrogenistein 6-hydroxy-O-desmethylangolensin 2-(4-hydroxyphenyl) propionic acid | Cancer | * Genistein treatment significantly inhibited the activities of nuclear factor-kappa B (NF-κB) signaling pathway which resulted in a reduced COX-2 protein level | ELISA Flow cytometry Western blot analysis Morphological determination | * Human gastric cancer BGC-823 cells | [69] | |
| Morin | Flavonol | Morin glucuronides Morin sulfates | Cancer | * Morin inhibited the activation of nuclear factor-kappa B (NF-κB) pathway, thus reducing the production of IL-6 and IL-8 | Reverse transcription PCR Apoptosis assay ELISA Western blot analysis Cell viability Immunofluorescent staining assay | * Athymic nude mice | * HT-29 cells * HCT-116 cells | [68] | |
| Chrysin | Flavones | No available data | Cancer | * Chrysin downregulated the activities of nuclear factor-kappa B (NF-κB) which reduced the expression of recepteur d’origine nantais (RON), a c-Met factor receptor critical for gastric cancer invasion and metastasis | Reverse transcription PCR ELISA Western blot analysis Cell viability | * AGS human gastric cancer cells | [67] | ||
| Quercetin | Flavonol | 3,4-dihydroxyphenylacetic acid 3-(3-hydroxyphenyl) propionic acid 3,4-dihydroxybenzoic 4-hydroxybenzoic acid | Cancer | * Administration of quercetin showed an antimetastatic effect against gastric cancer through the inhibition of nuclear factor-kappa B (NF-κB) | uPA activity assay Western blot analysis Cell viability | * MGC803 cells * GC7901 cells * AGS human gastric cancer cells | [70] | ||
| AMPK Pathway | Genistein | Isoflavones | Dihydrogenistein 6-hydroxy-O-desmethylangolensin 2-(4-hydroxyphenyl) propionic acid | Cancer | * Combination of 5-Fluorouracil (5-FU) and genistein in colon cancer cells activated AMPK activities and upregulated p53, p21, and Bax signals | Reverse transcription PCR ELISA Western blot analysis Cell viability | * HT-29 cells | [99] | |
| Apoptosis | Morin | Flavonol | Morin glucuronides Morin sulfates | Cancer | * The administration of morin induced apoptosis through the increased level of ROS | Electron microscopy Flow cytometry Cell viability assays Immunoblotting Quantitative PCR Flow cytometry | * SW480 cells | [53] | |
| Fisetin | Flavonol | No available data | Cancer | * Fisetin treatment significantly induced the expression of pro-apoptotic Caspase-9 and Caspase-3 and inhibited the anti-apoptotic Bcl-2 | Histopathological analysis ELISA Estimation of lipid peroxidation | * Male Wistar rats | [100] | ||
| Hesperidin | Flavanones | Hesperetin | Cancer | * Cells treated with hesperidin showed a decreased expression of B-cell CLL/lymphoma 2 (BCL2) mRNA, and an increased expression of BCL2-associated X protein (BAX). * Hesperidin induced the expression of pro-apoptotic Caspase-3 | Reverse transcription PCR Caspase3 activity assay Western blot analysis Cell viability | * SNU-C4 cells | [101] | ||
| Kaempferol | Flavonol | Kaempferol-3-O-glucoside p-coumaric acid kaempferol 3-(4 hydroxyphenyl) propionic acid 3-phenylpropionic acid | Cancer | * Kaempferol treatment increased the number of early apoptotic cells * Treatment with Kaempferol increased the levels of cleaved caspase-9, caspase-3, and caspase-7 | LDH assays Cell viability Immunoprecipitation (IP) assay Reverse transcription PCR Western blot analysis Caspase-8 assays | * AGS cells * SNU-216 cells * NCI-N87 cells * HT-29 cells | [51,102] | ||
| Apigenin | Flavones | Apigenin 3-(4-hydroxyphenyl) propionic acid | Cancer | * Apigenin treatment reduced cellular proliferation and induced apoptosis through the suppression of the antiapoptotic proteins Bcl-xL and Mcl-1 * Apigenin reported to induce apoptosis through the inhibition of STAT3 phosphorylation | Immunohistochemistry Cell proliferation assay Western blot analysis | * HT29 cells | [49] | ||
| Genistein | Isoflavones | Dihydrogenistein 6-hydroxy-O-desmethylangolensin 2-(4-hydroxyphenyl) propionic acid | Cancer | * Genistein induced apoptosis by promoting the expression of Bax/Bcl-2 and caspase-3 | Quantitative PCR Flow cytometry Immunofluorescence imaging Protein extraction Immunoblot | * HT-29 cells | [50] | ||
| Chrysin | Flavones | No available data | Cancer | * Chrysin reduced cellular proliferation and induced apoptosis in colon cancer cells through caspase-3 and caspase-9 | Caspase-3 and Caspase-9 Asctivity Morphological Assessments Reverse transcription PCR Flow cytometry | * Male BALB/c mice | * CT26 cells | [52] | |
| Tangeretin | Flavones | Tangeretin-O-glucuronides | Cancer | * Tangeretin administration induced apoptosis by upregulating the activities of caspase 3, 8, and 9 * Tangeretin treatment decreased the membrane potential of mitochondria (MMP) | Caspase-3 and Caspase-9 Asctivity Western blot analysis MTT assay for cell viability Flow cytometry | * AGS cells | [103] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL-Ishaq, R.K.; Mazurakova, A.; Kubatka, P.; Büsselberg, D. Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon? Cancers 2022, 14, 6073. https://doi.org/10.3390/cancers14246073
AL-Ishaq RK, Mazurakova A, Kubatka P, Büsselberg D. Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon? Cancers. 2022; 14(24):6073. https://doi.org/10.3390/cancers14246073
Chicago/Turabian StyleAL-Ishaq, Raghad Khalid, Alena Mazurakova, Peter Kubatka, and Dietrich Büsselberg. 2022. "Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon?" Cancers 14, no. 24: 6073. https://doi.org/10.3390/cancers14246073
APA StyleAL-Ishaq, R. K., Mazurakova, A., Kubatka, P., & Büsselberg, D. (2022). Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon? Cancers, 14(24), 6073. https://doi.org/10.3390/cancers14246073