


c-Met and EPHA7 Receptor Tyrosine Kinases Are Related to Prognosis in Clear Cell Renal Cell Carcinoma: Focusing on the Association with Myoferlin Expression


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Receptor Tyrosine Kinases Involved in Clear Cell Renal Cell Carcinoma Using Proteomic Dataset
2.2. Immunohistochemical Staining for Receptor Tyrosine Kinases and Ligands
2.3. Clinical Information and International Metastatic RCC Database Consortium Risk Groups
2.4. Survival Analysis Using Immunohistochemical Expression
2.5. Network-Based Prioritization of Interacting Proteins and Functional Enrichment Analysis
2.6. Statistical Analysis
3. Results
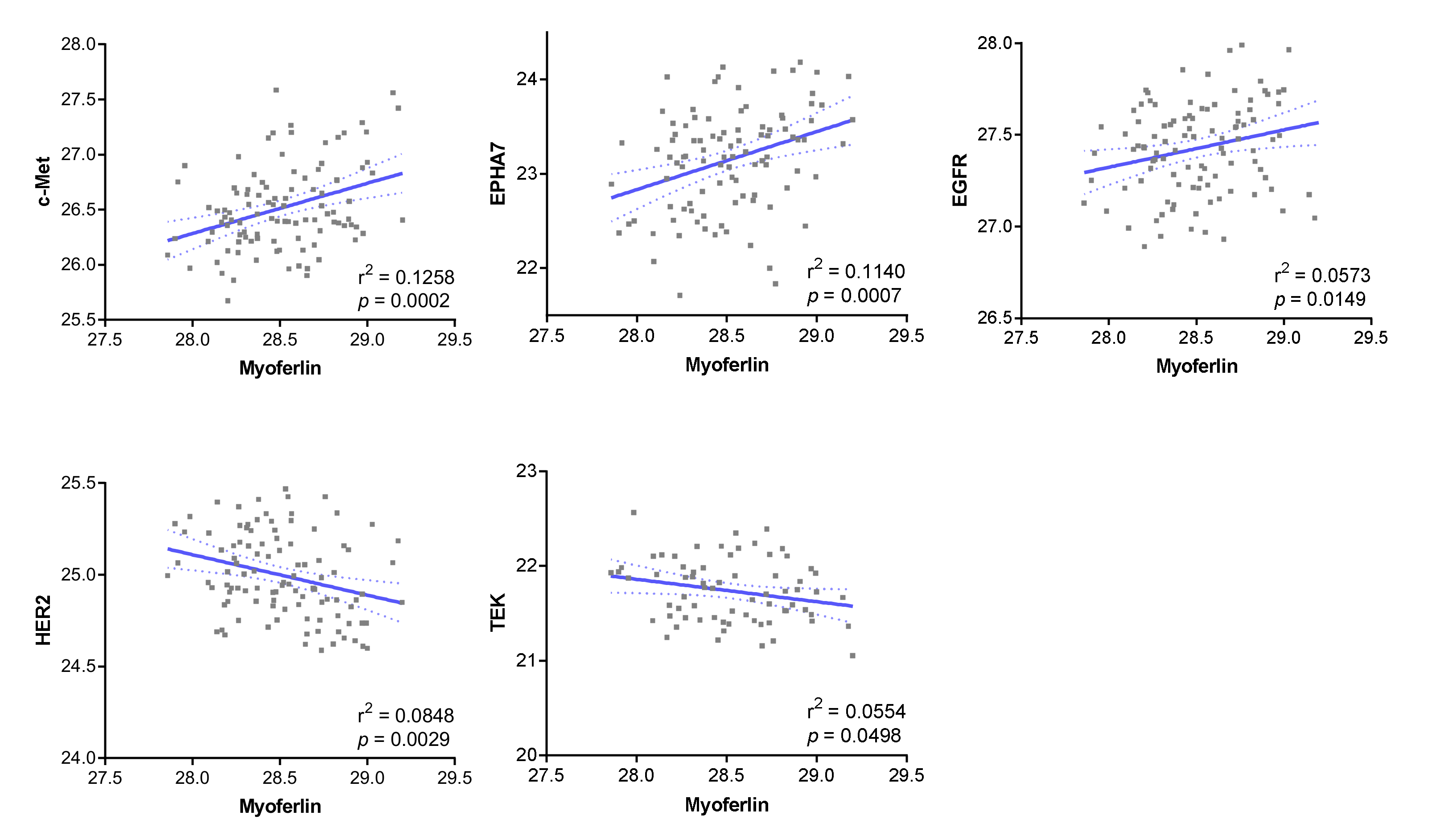
3.1. Receptor Tyrosine Kinases Correlated with Myoferlin in the Proteomic Dataset
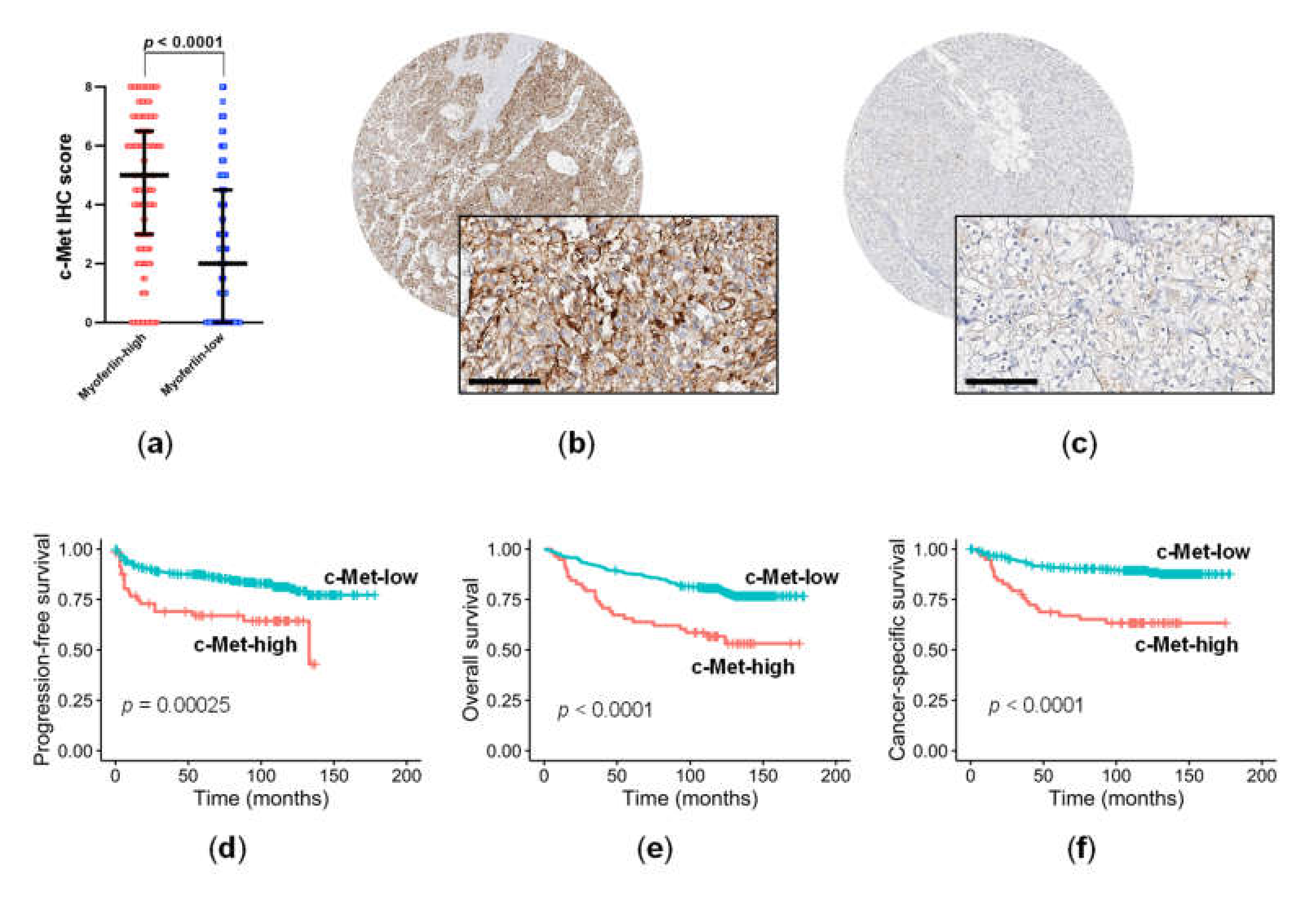
3.2. c-Met Expression Was Significantly Related to Myoferlin Expression and Pathological Parameters in ccRCC
3.3. High c-Met Expression Was an Independent Negative Prognostic Factor in ccRCC
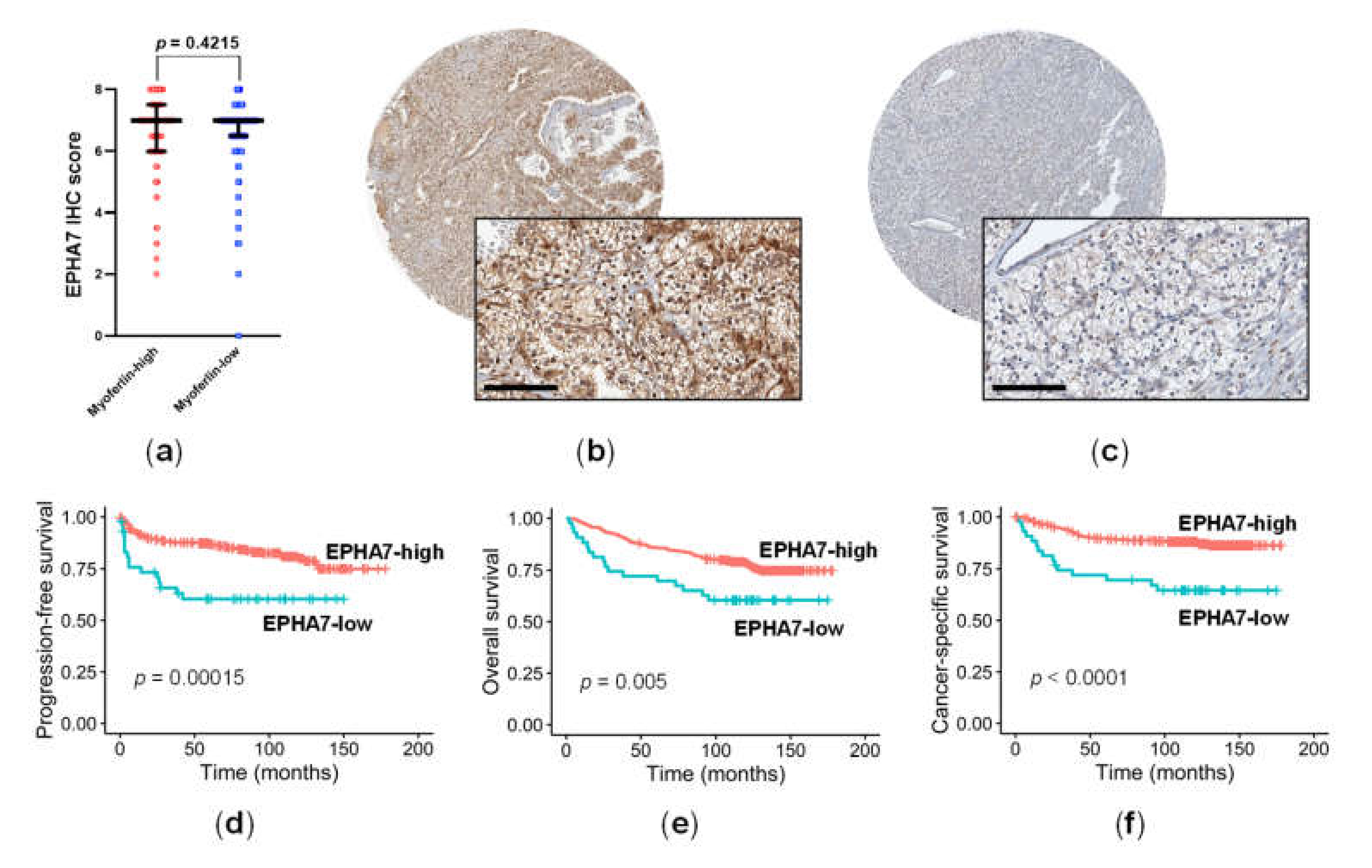
3.4. EPHA7 Expression Was Not Correlated with Myoferlin Expression, but Was Independently Associated with Prognosis in ccRCC
3.5. Network-Based Prioritization of Interacting Proteins for c-Met and Myoferlin
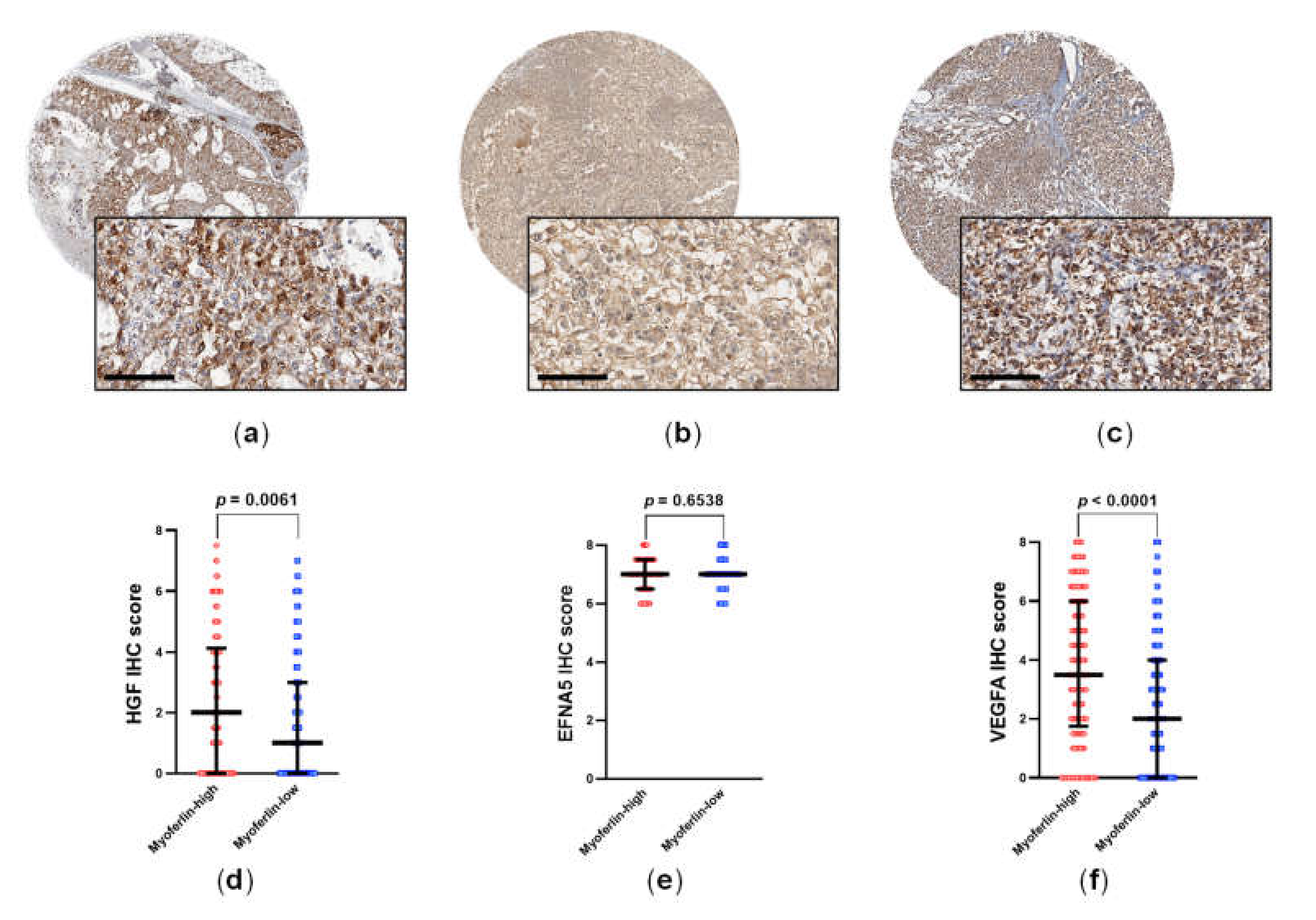
3.6. HGF and VEGFA Are RTK Ligands That Are Correlated to Myoferlin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Behbahani, T.E.; Thierse, C.; Baumann, C.; Holl, D.; Bastian, P.J.; von Ruecker, A.; Muller, S.C.; Ellinger, J.; Hauser, S. Tyrosine kinase expression profile in clear cell renal cell carcinoma. World J. Urol. 2012, 30, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gordoa, T.; Garcia-Bermejo, M.L.; Grande, E.; Garrido, P.; Carrato, A.; Molina-Cerrillo, J. Targeting tyrosine kinases in renal cell carcinoma: “New Bullets against Old Guys”. Int. J. Mol. Sci. 2019, 20, 1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motzer, R.J.; Jonasch, E.; Boyle, S.; Carlo, M.I.; Manley, B.; Agarwal, N.; Alva, A.; Beckermann, K.; Choueiri, T.K.; Costello, B.A.; et al. NCCN guidelines insights: Kidney cancer, version 1.2021. J. Natl. Compr. Cancer Netw. 2020, 18, 1160–1170. [Google Scholar] [CrossRef]
- Bernatchez, P.N.; Acevedo, L.; Fernandez-Hernando, C.; Murata, T.; Chalouni, C.; Kim, J.; Erdjument-Bromage, H.; Shah, V.; Gratton, J.P.; McNally, E.M.; et al. Myoferlin regulates vascular endothelial growth factor receptor-2 stability and function. J. Biol. Chem. 2007, 282, 30745–30753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahmy, K.; Gonzalez, A.; Arafa, M.; Peixoto, P.; Bellahcene, A.; Turtoi, A.; Delvenne, P.; Thiry, M.; Castronovo, V.; Peulen, O. Myoferlin plays a key role in VEGFA secretion and impacts tumor-associated angiogenesis in human pancreas cancer. Int. J. Cancer 2016, 138, 652–663. [Google Scholar] [CrossRef]
- Yu, C.; Sharma, A.; Trane, A.; Utokaparch, S.; Leung, C.; Bernatchez, P. Myoferlin gene silencing decreases Tie-2 expression in vitro and angiogenesis in vivo. Vascul. Pharmacol. 2011, 55, 26–33. [Google Scholar] [CrossRef]
- Demonbreun, A.R.; Posey, A.D.; Heretis, K.; Swaggart, K.A.; Earley, J.U.; Pytel, P.; McNally, E.M. Myoferlin is required for insulin-like growth factor response and muscle growth. FASEB J. 2010, 24, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Turtoi, A.; Blomme, A.; Bellahcene, A.; Gilles, C.; Hennequiere, V.; Peixoto, P.; Bianchi, E.; Noel, A.; De Pauw, E.; Lifrange, E.; et al. Myoferlin is a key regulator of EGFR activity in breast cancer. Cancer Res. 2013, 73, 5438–5448. [Google Scholar] [CrossRef] [Green Version]
- Hermanns, C.; Hampl, V.; Holzer, K.; Aigner, A.; Penkava, J.; Frank, N.; Martin, D.E.; Maier, K.C.; Waldburger, N.; Roessler, S.; et al. The novel MKL target gene myoferlin modulates expansion and senescence of hepatocellular carcinoma. Oncogene 2017, 36, 3464–3476. [Google Scholar] [CrossRef]
- Jung, M.; Lee, C.; Park, J.H.; Moon, K.C. Prognostic significance of immunohistochemical staining for myoferlin in clear cell renal cell carcinoma and its association with epidermal growth factor receptor expression. Urol. Oncol. 2019, 37, 812-e9. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Zhao, C.; Tolkach, Y.; Nettersheim, D.; Schmidt, D.; Kristiansen, G.; Mueller, S.C.; Ritter, M.; Hauser, S.; Ellinger, J. Knockdown of Myoferlin Suppresses Migration and Invasion in Clear-Cell Renal-Cell Carcinoma. Anticancer Res. 2020, 40, 3119–3128. [Google Scholar] [CrossRef]
- Lai, Y.; Zhao, Z.; Zeng, T.; Liang, X.; Chen, D.; Duan, X.; Zeng, G.; Wu, W. Crosstalk between VEGFR and other receptor tyrosine kinases for TKI therapy of metastatic renal cell carcinoma. Cancer Cell Int. 2018, 18, 31. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.J.; Dhanasekaran, S.M.; Petralia, F.; Pan, J.; Song, X.; Hu, Y.; da Veiga Leprevost, F.; Reva, B.; Lih, T.M.; Chang, H.Y.; et al. Integrated proteogenomic characterization of clear cell renal cell carcinoma. Cell 2019, 179, 964–983.e31. [Google Scholar] [CrossRef] [Green Version]
- Braschi, B.; Denny, P.; Gray, K.; Jones, T.; Seal, R.; Tweedie, S.; Yates, B.; Bruford, E. Genenames.org: The HGNC and VGNC resources in 2019. Nucleic Acids Res. 2019, 47, D786–D792. [Google Scholar] [CrossRef]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Ahmad, N.; Keehn, C.A.; Coppola, D. The expression of insulin-like growth factor-I receptor correlates with Fuhrman grading of renal cell carcinomas. Hum. Pathol. 2004, 35, 1132–1136. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.S.; Jung, J.G.; Oh, M.K.; Chung, T.H.; Kim, J. Korean alcohol guidelines for moderate drinking based on facial flushing. Korean J. Fam. Med. 2019, 40, 204–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, D.Y.; Xie, W.; Regan, M.M.; Warren, M.A.; Golshayan, A.R.; Sahi, C.; Eigl, B.J.; Ruether, J.D.; Cheng, T.; North, S.; et al. Prognostic factors for overall survival in patients with metastatic renal cell carcinoma treated with vascular endothelial growth factor-targeted agents: Results from a large, multicenter study. J. Clin. Oncol. 2009, 27, 5794–5799. [Google Scholar] [CrossRef]
- Schwartz, L.H.; Litiere, S.; de Vries, E.; Ford, R.; Gwyther, S.; Mandrekar, S.; Shankar, L.; Bogaerts, J.; Chen, A.; Dancey, J.; et al. RECIST 1.1-Update and clarification: From the RECIST committee. Eur. J. Cancer 2016, 62, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, B.; Zhao, C.; Ba, Z.; Xu, H.; Yan, X.; Liu, W.; Zhu, B.; Wang, L.; Ren, C. Myoferlin, a multifunctional protein in normal cells, has novel and key roles in various cancers. J. Cell Mol. Med. 2019, 23, 7180–7189. [Google Scholar] [CrossRef] [Green Version]
- Theodros, D.; Murter, B.M.; Sidhom, J.W.; Nirschl, T.R.; Clark, D.J.; Chen, L.; Tam, A.J.; Blosser, R.L.; Schwen, Z.R.; Johnson, M.H.; et al. High-dimensional cytometry (ExCYT) and mass spectrometry of myeloid infiltrate in clinically localized clear cell renal cell carcinoma identifies novel potential myeloid targets for immunotherapy. Mol. Cell Proteom. 2020, 19, 1850–1859. [Google Scholar] [CrossRef]
- Hornigold, N.; Dunn, K.R.; Craven, R.A.; Zougman, A.; Trainor, S.; Shreeve, R.; Brown, J.; Sewell, H.; Shires, M.; Knowles, M.; et al. Dysregulation at multiple points of the kynurenine pathway is a ubiquitous feature of renal cancer: Implications for tumour immune evasion. Br. J. Cancer 2020, 123, 137–147. [Google Scholar] [CrossRef]
- Orlando, E.; Aebersold, D.M.; Medova, M.; Zimmer, Y. Oncogene addiction as a foundation of targeted cancer therapy: The paradigm of the MET receptor tyrosine kinase. Cancer Lett. 2019, 443, 189–202. [Google Scholar] [CrossRef]
- Trusolino, L.; Bertotti, A.; Comoglio, P.M. MET signalling: Principles and functions in development, organ regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2010, 11, 834–848. [Google Scholar] [CrossRef]
- Harshman, L.C.; Choueiri, T.K. Targeting the hepatocyte growth factor/c-Met signaling pathway in renal cell carcinoma. Cancer J. 2013, 19, 316–323. [Google Scholar] [CrossRef]
- Gibney, G.T.; Aziz, S.A.; Camp, R.L.; Conrad, P.; Schwartz, B.E.; Chen, C.R.; Kelly, W.K.; Kluger, H.M. c-Met is a prognostic marker and potential therapeutic target in clear cell renal cell carcinoma. Ann. Oncol. 2013, 24, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Kanetake, H.; Kanda, S. Presence of phosphorylated hepatocyte growth factor receptor/c-Met is associated with tumor progression and survival in patients with conventional renal cell carcinoma. Clin. Cancer Res. 2006, 12, 4876–4881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choueiri, T.K.; Hessel, C.; Halabi, S.; Sanford, B.; Michaelson, M.D.; Hahn, O.; Walsh, M.; Olencki, T.; Picus, J.; Small, E.J.; et al. Cabozantinib versus sunitinib as initial therapy for metastatic renal cell carcinoma of intermediate or poor risk (Alliance A031203 CABOSUN randomised trial): Progression-free survival by independent review and overall survival update. Eur. J. Cancer 2018, 94, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, M.T.; Fan, C.M. c-MET regulates myoblast motility and myocyte fusion during adult skeletal muscle regeneration. PLoS ONE 2013, 8, e81757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duplaquet, L.; Kherrouche, Z.; Baldacci, S.; Jamme, P.; Cortot, A.B.; Copin, M.C.; Tulasne, D. The multiple paths towards MET receptor addiction in cancer. Oncogene 2018, 37, 3200–3215. [Google Scholar] [CrossRef]
- Makhov, P.; Joshi, S.; Ghatalia, P.; Kutikov, A.; Uzzo, R.G.; Kolenko, V.M. Resistance to systemic therapies in clear cell renal cell carcinoma: Mechanisms and management strategies. Mol. Cancer Ther. 2018, 17, 1355–1364. [Google Scholar] [CrossRef] [Green Version]
- Marona, P.; Gorka, J.; Kotlinowski, J.; Majka, M.; Jura, J.; Miekus, K. C-Met as a key factor responsible for sustaining undifferentiated phenotype and therapy resistance in renal carcinomas. Cells 2019, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Pasquale, E.B. Eph receptors and ephrins in cancer: Bidirectional signalling and beyond. Nat. Rev. Cancer 2010, 10, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Kou, C.J.; Kandpal, R.P. Differential expression patterns of Eph receptors and ephrin ligands in human cancers. Biomed. Res. Int. 2018, 2018, 7390104. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Characteristics Category | c-Met High (n = 58) | c-Met Low (n = 352) | p |
|---|---|---|---|---|
| Age | <58.5 | 26 (44.8%) | 179 (50.9%) | 0.4786 |
| >58.5 | 32 (55.2%) | 173 (49.1%) | ||
| Sex | Male | 45 (77.6%) | 257 (73.0%) | 0.5673 |
| Female | 13 (22.4%) | 95 (27.0%) | ||
| Surgery | Radical | 44 (75.9%) | 228 (64.8%) | 0.1321 |
| Partial | 14 (24.1%) | 124 (35.2%) | ||
| Body mass index 1 | <24.4 kg/m2 | 25 (49.0%) | 152 (49.2%) | 1.0000 |
| ≥24.4 kg/m2 | 26 (51.0%) | 157 (50.9%) | ||
| Smoking 1 | None | 43 (84.3%) | 250 (80.1%) | 0.7002 |
| Current or past | 8 (15.7%) | 59 (19.1%) | ||
| Alcohol 1 | Not heavy | 45 (88.2%) | 276 (89.3%) | 1.0000 |
| Heavy | 6 (11.8%) | 33 (10.7%) | ||
| IMDC risk group 2 | Good/intermediate | 36 (85.7%) | 246 (96.9%) | 0.0058 |
| Poor | 6 (14.3%) | 8 (3.1%) | ||
| TNM stage | I or II | 37 (63.8%) | 283 (80.4%) | 0.0078 |
| III or IV | 21 (36.2%) | 69 (19.6%) | ||
| WHO grade | 1 or 2 | 8 (13.8%) | 185 (52.6%) | <0.0001 |
| 3 or 4 | 50 (86.2%) | 167 (47.4%) | ||
| Myoferlin | Low | 22 (37.9%) | 265 (75.3%) | <0.0001 |
| High | 36 (62.1%) | 87 (24.7%) |
| Survival | Variables | Univariate HR 1 | p | Multivariate HR 1 | p |
|---|---|---|---|---|---|
| Progression-free survival | c-Met (high vs. low) | 2.516 (1.514–4.183) | 0.0004 | 1.538 (0.9102–2.597) | 0.1078 |
| TNM stage (III/IV vs. I/II) | 12.610 (7.896–20.140) | <0.0001 | 8.729 (5.376–14.174) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 4.990 (2.841–8.764) | <0.0001 | 2.440 (1.339–4.446) | 0.0036 | |
| Overall survival | c-Met (high vs. low) | 2.581 (1.652–4.032) | <0.0001 | 1.834 (1.153–2.919) | 0.0105 |
| TNM stage (III/IV vs. I/II) | 5.185 (3.551–7.571) | <0.0001 | 4.279 (2.840–6.449) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 2.515 (1.676–3.773) | <0.0001 | 1.674 (1.043–2.686) | 0.0327 | |
| Cancer-specific survival | c-Met (high vs. low) | 3.811 (2.239–6.485) | <0.0001 | 1.974 (1.150–3.389) | 0.0137 |
| TNM stage (III/IV vs. I/II) | 17.470 (9.719–31.410) | <0.0001 | 9.813 (5.383–17.886) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 15.570 (5.644–42.940) | <0.0001 | 6.200 (2.171–17.706) | <0.0001 |
| Survival | Variables | Univariate HR 1 | p | Multivariate HR 1 | p |
|---|---|---|---|---|---|
| Overall survival | c-Met (High vs. Low) | 3.674 (2.370–5.698) | <0.0001 | 2.381 (1.503–3.771) | 0.0002 |
| TNM stage (III/IV vs. I/II) | 5.892 (4.071–8.527) | <0.0001 | 4.492 (2.889–6.985) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 2.928 (2.007–4.271) | <0.0001 | 1.331 (0.840–2.111) | 0.2236 |
| Survival | Variables | Univariate HR 1 | p | Multivariate HR 1 | p |
|---|---|---|---|---|---|
| Progression-free survival | EPHA7 (Low vs. High) | 2.511 (1.358–4.644) | 0.0033 | 2.311 (1.237–4.319) | 0.0086 |
| TNM stage (III/IV vs. I/II) | 11.368 (7.118–18.156) | <0.0001 | 9.277 (5.710–15.074) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 4.585 (2.610–8.054) | <0.0001 | 2.485 (1.377–4.484) | 0.0025 | |
| Overall survival | EPHA7 (Low vs. High) | 1.814 (0.993–3.315) | 0.0528 | 1.655 (0.898–3.050) | 0.1063 |
| TNM stage (III/IV vs. I/II) | 5.342 (3.617–7.892) | <0.0001 | 4.527 (3.007–6.813) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 2.773 (1.795–4.286) | <0.0001 | 1.824 (1.151–2.891) | 0.0105 | |
| Cancer-specific survival | EPHA7 (Low vs. High) | 2.979 (1.548–5.733) | 0.0011 | 2.352 (1.214–4.558) | 0.0113 |
| TNM stage (III/IV vs. I/II) | 15.906 (8.846–28.601) | <0.0001 | 10.712 (5.879–19.516) | <0.0001 | |
| WHO grade (3/4 vs. 1/2) | 14.460 (5.242–39.889) | <0.0001 | 6.680 (2.362–18.888) | 0.0003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, M.; Lee, S.; Moon, K.C. c-Met and EPHA7 Receptor Tyrosine Kinases Are Related to Prognosis in Clear Cell Renal Cell Carcinoma: Focusing on the Association with Myoferlin Expression. Cancers 2022, 14, 1095. https://doi.org/10.3390/cancers14041095
Jung M, Lee S, Moon KC. c-Met and EPHA7 Receptor Tyrosine Kinases Are Related to Prognosis in Clear Cell Renal Cell Carcinoma: Focusing on the Association with Myoferlin Expression. Cancers. 2022; 14(4):1095. https://doi.org/10.3390/cancers14041095
Chicago/Turabian StyleJung, Minsun, Seokhyeon Lee, and Kyung Chul Moon. 2022. "c-Met and EPHA7 Receptor Tyrosine Kinases Are Related to Prognosis in Clear Cell Renal Cell Carcinoma: Focusing on the Association with Myoferlin Expression" Cancers 14, no. 4: 1095. https://doi.org/10.3390/cancers14041095
APA StyleJung, M., Lee, S., & Moon, K. C. (2022). c-Met and EPHA7 Receptor Tyrosine Kinases Are Related to Prognosis in Clear Cell Renal Cell Carcinoma: Focusing on the Association with Myoferlin Expression. Cancers, 14(4), 1095. https://doi.org/10.3390/cancers14041095