
RBM24 in the Post-Transcriptional Regulation of Cancer Progression: Anti-Tumor or Pro-Tumor Activity?


Abstract
:Simple Summary
Abstract
1. Introduction
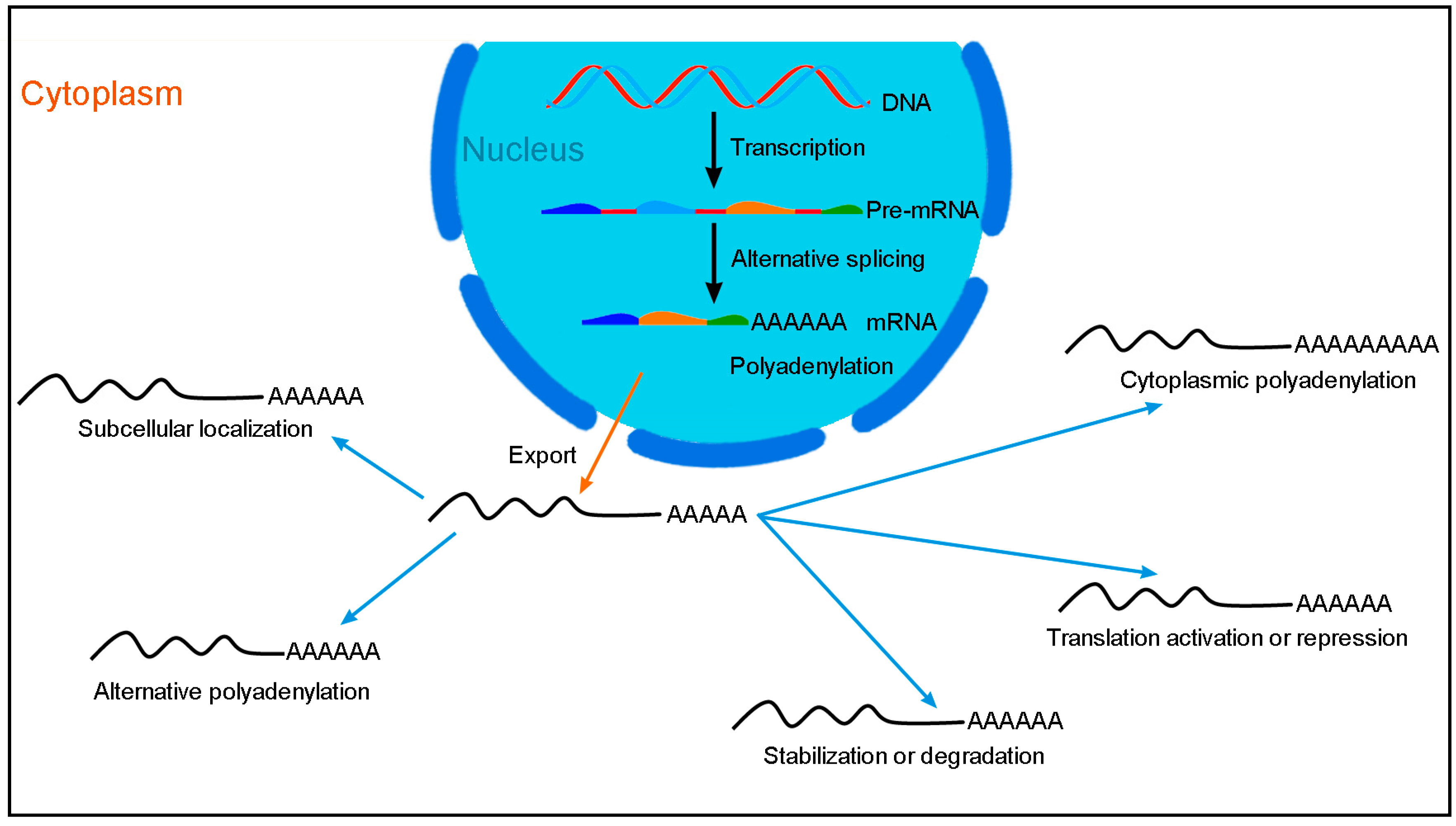
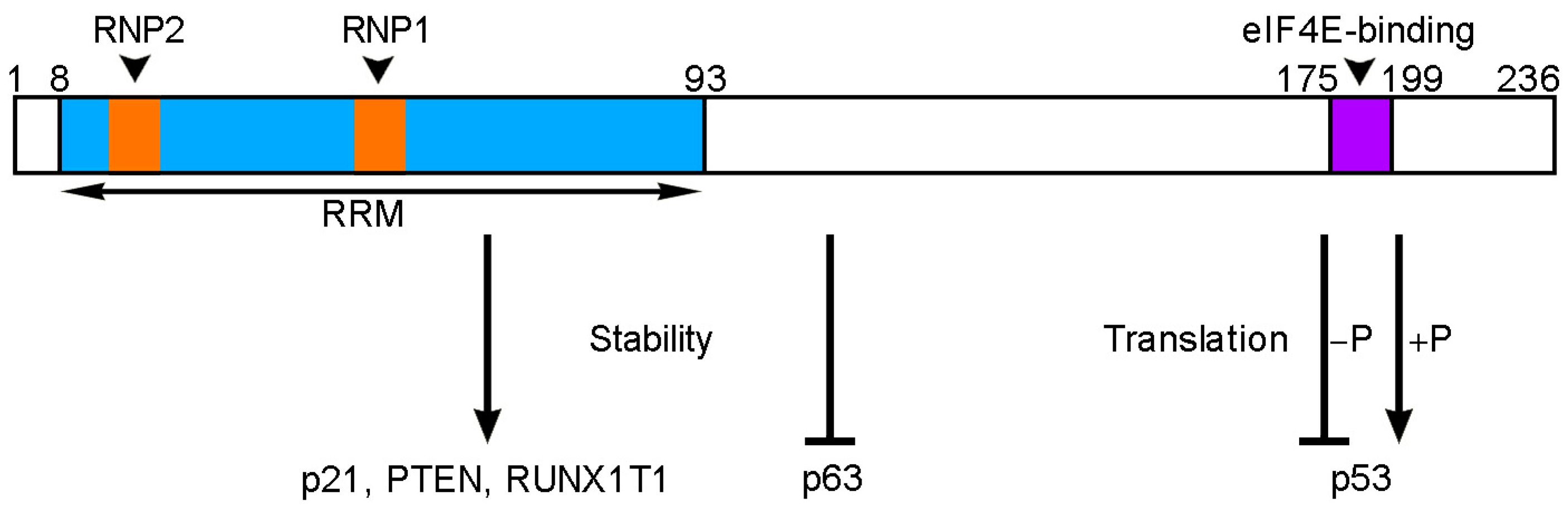
2. RBM24-Mediated Post-Transcriptional Regulation of Gene Expression in Cancer Cells
3. RBM24 in Cancer Development
3.1. RBM24 in Hepatocellular Carcinoma
3.2. RBM24 in Gastrointestinal Cancers
3.3. Tumor Suppressor Function of RBM24 in Nasopharyngeal Carcinoma
3.4. RBM24 Expression and Function in Lung Cancer
3.5. Pro-Tumor Activity of RBM24 in Bladder Cancer
3.6. RBM24 in Other Cancers
3.7. Possible Roles of RBM24 in the Progression of Other Diseases
4. Regulation of RBM24 Expression and Activity in Cancers
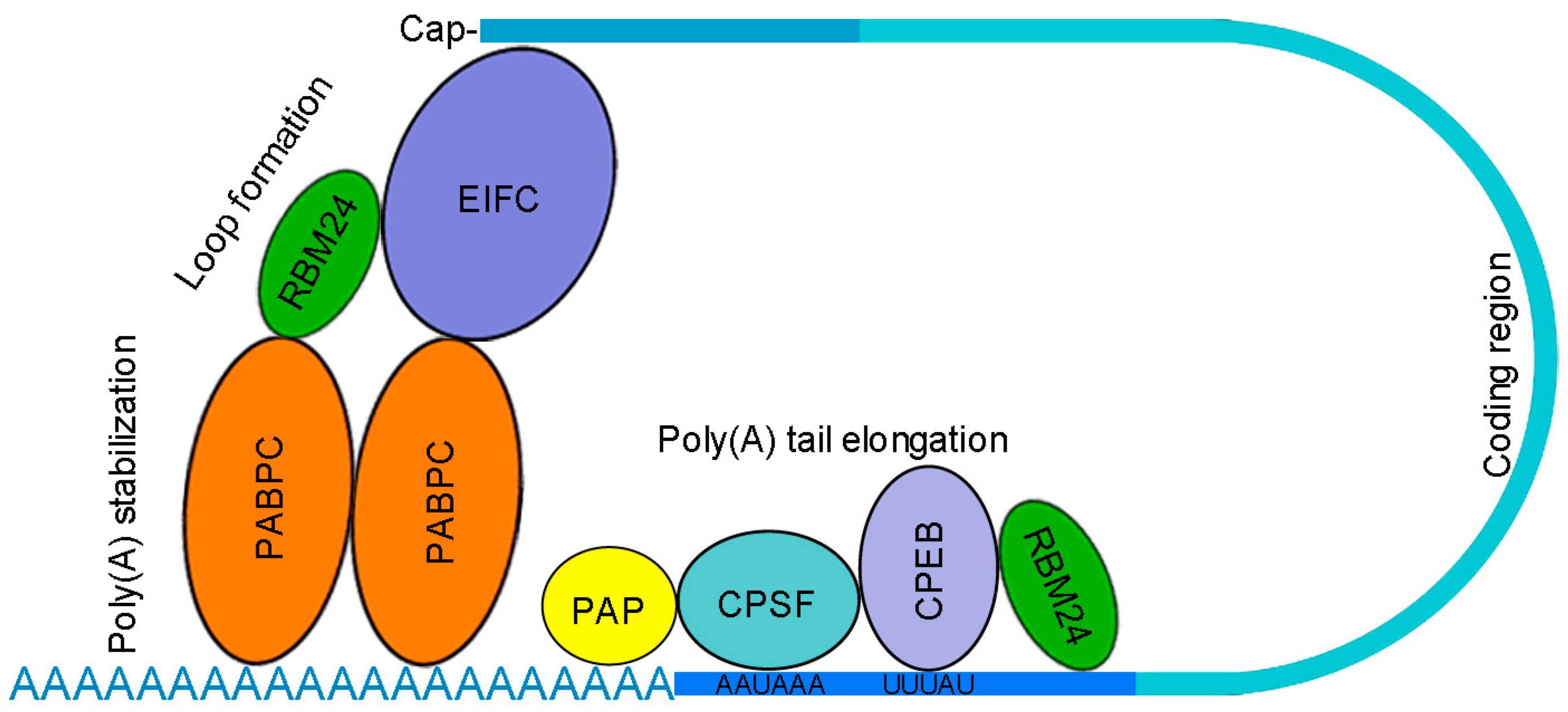
5. RBM24-Interacting Proteins
6. Discussion
7. Conclusions
Funding
Conflicts of Interest
References
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukong, K.E.; Chang, K.W.; Khandjian, E.W.; Richard, S. RNA-binding proteins in human genetic disease. Trends Genet. 2008, 24, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Brinegar, A.E.; Cooper, T.A. Roles for RNA-binding proteins in development and disease. Brain Res. 2016, 1647, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.L.; Grifone, R. RNA-binding proteins in the post-transcriptional control of skeletal muscle development, regeneration and disease. Front. Cell Dev. Biol. 2021, 9, 738978. [Google Scholar] [CrossRef] [PubMed]
- Marchese, D.; de Groot, N.S.; Lorenzo Gotor, N.; Livi, C.M.; Tartaglia, G.G. Advances in the characterization of RNA-binding proteins. Wiley Interdiscip. Rev. RNA 2016, 7, 793–810. [Google Scholar] [CrossRef]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef]
- Sebestyén, E.; Singh, B.; Miñana, B.; Pagès, A.; Mateo, F.; Pujana, M.A.; Valcárcel, J.; Eyras, E. Large-scale analysis of genome and transcriptome alterations in multiple tumors unveils novel cancer-relevant splicing networks. Genome Res. 2016, 26, 732–744. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.L.; Li, B.; Luo, Y.X.; Lin, Q.; Liu, S.R.; Zhang, X.Q.; Zhou, H.; Yang, J.H.; Qu, L.H. Comprehensive genomic characterization of RNA-binding proteins across human cancers. Cell Rep. 2018, 22, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Ni, H.; Liu, Y.; Yuan, Y.; Xi, T.; Li, X.; Zheng, L. RNA-binding proteins in tumor progression. J. Hematol. Oncol. 2020, 13, 90. [Google Scholar] [CrossRef]
- Zhang, B.; Babu, K.R.; Lim, C.Y.; Kwok, Z.H.; Li, J.; Zhou, S.; Yang, H.; Tay, Y. A comprehensive expression landscape of RNA-binding proteins (RBPs) across 16 human cancer types. RNA Biol. 2020, 17, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Neelamraju, Y.; Gonzalez-Perez, A.; Bhat-Nakshatri, P.; Nakshatri, H.; Janga, S.C. Mutational landscape of RNA-binding proteins in human cancers. RNA Biol. 2018, 15, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.; Billaud, M.; Almeida, R. RNA-binding proteins in cancer: Old players and new actors. Trends Cancer 2017, 3, 506–528. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mir, C.; Garcia-Mayea, Y.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. RNA-binding proteins: Underestimated contributors in tumorigenesis. In Seminars in Cancer Biology; Academic Press: New York, NY, USA, 2022. [Google Scholar] [CrossRef]
- Hong, S. RNA binding protein as an emerging therapeutic target for cancer prevention and treatment. J. Cancer Prev. 2017, 22, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, C.W.; Preet, R.; Dhir, T.; Dixon, D.A.; Brody, J.R. Understanding and targeting the disease-related RNA binding protein human antigen R (HuR). Wiley Interdiscip. Rev. RNA 2020, 11, e1581. [Google Scholar] [CrossRef]
- Kang, D.; Lee, Y.; Lee, J.S. RNA-binding proteins in cancer: Functional and therapeutic perspectives. Cancers 2020, 12, 2699. [Google Scholar] [CrossRef]
- Mohibi, S.; Chen, X.; Zhang, J. Cancer the‘RBP’eutics-RNA-binding proteins as therapeutic targets for cancer. Pharmacol. Ther. 2019, 203, 107390. [Google Scholar] [CrossRef]
- Grifone, R.; Xie, X.; Bourgeois, A.; Saquet, A.; Duprez, D.; Shi, D.L. The RNA-binding protein Rbm24 is transiently expressed in myoblasts and is required for myogenic differentiation during vertebrate development. Mech. Dev. 2014, 134, 1–15. [Google Scholar] [CrossRef]
- Grifone, R.; Saquet, A.; Xu, Z.G.; Shi, D.L. Expression patterns of Rbm24 in lens, nasal epithelium, and inner ear during mouse embryonic development. Dev. Dyn. 2018, 247, 1160–1169. [Google Scholar] [CrossRef] [Green Version]
- Grifone, R.; Shao, M.; Saquet, A.; Shi, D.L. RNA-binding protein Rbm24 as a multifaceted post-transcriptional regulator of embryonic lineage differentiation and cellular homeostasis. Cells 2020, 9, 1891. [Google Scholar] [CrossRef]
- Jin, D.; Hidaka, K.; Shirai, M.; Morisaki, T. RNA-binding motif protein 24 regulates myogenin expression and promotes myogenic differentiation. Genes Cells 2010, 15, 1158–1167. [Google Scholar] [CrossRef]
- Yang, J.; Hung, L.H.; Licht, T.; Kostin, S.; Looso, M.; Khrameeva, E.; Bindereif, A.; Schneider, A.; Braun, T. RBM24 is a major regulator of muscle-specific alternative splicing. Dev. Cell 2014, 31, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhang, M.; Qian, Y.; Xu, E.; Zhang, J.; Chen, X. Rbm24, an RNA-binding protein and a target of p53, regulates p21 expression via mRNA stability. J. Biol. Chem. 2014, 289, 3164–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; de Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130. [Google Scholar] [CrossRef] [Green Version]
- Shao, M.; Lu, T.; Zhang, C.; Zhang, Y.Z.; Kong, S.H.; Shi, D.L. Rbm24 controls poly(A) tail length and translation efficiency of crystallin mRNAs in the lens via cytoplasmic polyadenylation. Proc. Natl. Acad. Sci. USA 2020, 117, 7245–7254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Zhang, J.J.; Shi, D.L. Loss of Rbm24a causes defective hair cell development in the zebrafish inner ear and neuromasts. J. Genet. Genomics 2020, 47, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Grifone, R.; Saquet, A.; Desgres, M.; Sangiorgi, C.; Gargano, C.; Li, Z.; Coletti, D.; Shi, D.L. Rbm24 displays dynamic functions required for myogenic differentiation during muscle regeneration. Sci. Rep. 2021, 11, 9423. [Google Scholar] [CrossRef]
- Hua, W.F.; Zhong, Q.; Xia, T.L.; Chen, Q.; Zhang, M.Y.; Zhou, A.J.; Tu, Z.W.; Qu, C.; Li, M.Z.; Xia, Y.F.; et al. RBM24 suppresses cancer progression by upregulating miR-25 to target MALAT1 in nasopharyngeal carcinoma. Cell Death Dis. 2016, 7, e2352. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Ma, Y.; Ma, Z.; Liu, S.; Sun, L.; Li, J.; Zhao, F.; Li, Y.; Zhang, J.; Li, S.; et al. Circular RNA SMARCA5 suppressed non-small cell lung cancer progression by regulating miR-670-5p/RBM24 axis. Acta Biochim. Biophys. Sin. 2020, 52, 1071–1080. [Google Scholar] [CrossRef]
- Moon, S.U.; Kim, J.H.; Woo, H.G. Tumor suppressor RBM24 inhibits nuclear translocation of CTNNB1 and TP63 expression in liver cancer cells. Oncol. Lett. 2021, 22, 674. [Google Scholar] [CrossRef]
- Choi, J.H.; Kwon, S.M.; Moon, S.U.; Yoon, S.; Shah, M.; Lee, B.G.; Yang, J.; Park, Y.N.; Wang, H.J.; Woo, H.G. TPRG1-AS1 induces RBM24 expression and inhibits liver cancer progression by sponging miR-4691-5p and miR-3659. Liver Int. 2021, 41, 2788–2800. [Google Scholar] [CrossRef]
- Xia, R.M.; Liu, T.; Li, W.G.; Xu, X.Q. RNA-binding protein RBM24 represses colorectal tumourigenesis by stabilising PTEN mRNA. Clin. Transl. Med. 2021, 11, e383. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Mao, F.F.; Guo, L.; Guo, W.X. TRIM56 suppresses the malignant development of hepatocellular carcinoma via targeting RBM24 and inactivating the Wnt signaling. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Teng, L.; Fu, S.; Wang, G.; Li, Z.; Ding, C.; Wang, H.; Bi, L. Highly heterogeneous-related genes of triple-negative breast cancer: Potential diagnostic and prognostic biomarkers. BMC Cancer 2021, 21, 644. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Wu, C.; Sun, W.; Qiu, Y.; Li, J.; Liu, Z.; Jing, T.; Wang, H.; Liao, Y. miR-383 increases the cisplatin sensitivity of lung adenocarcinoma cells through inhibition of the RBM24-mediated NF-kappaB signaling pathway. Int. J. Oncol. 2021, 59, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.W.; Liu, K.L.; Lu, B.S.; Li, W.; Niu, Y.L.; Zhao, C.M.; Yang, Z.; Guo, P.Y.; Qi, J.C. RBM24 exacerbates bladder cancer progression by forming a Runx1t1/TCF4/miR-625-5p feedback loop. Exp. Mol. Med. 2021, 53, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.; Suh, Y.S.; Park, S.; Lee, J.; Kim, J.I.; Lee, S.; Lee, W.P.; Anczukow, O.; Yang, H.K.; Lee, C. Comprehensive analysis of alternative splicing in gastric cancer identifies epithelial-mesenchymal transition subtypes associated with survival. Cancer Res. 2022, 82, 543–555. [Google Scholar] [CrossRef]
- Ray, D.; Kazan, H.; Chan, E.T.; Peña-Castillo, L.; Chaudhry, S.; Talukder, S.; Blencowe, B.J.; Morris, Q.; Hughes, T.R. Rapid and systematic analysis of the RNA recognition specificities of RNA-binding proteins. Nat. Biotechnol. 2009, 27, 667–670. [Google Scholar] [CrossRef]
- Lucchesi, C.A.; Zhang, J.; Ma, B.; Chen, M.; Chen, X. Disruption of the Rbm38-eIF4E complex with a synthetic peptide pep8 increases p53 expression. Cancer Res. 2019, 79, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Zhang, J.; Chen, X.; Cho, S.J.; Chen, X. Glycogen synthase kinase 3 promotes p53 mRNA translation via phosphorylation of RNPC1. Genes Dev. 2013, 27, 2246–2258. [Google Scholar] [CrossRef] [Green Version]
- Xu, E.; Zhang, J.; Zhang, M.; Jiang, Y.; Cho, S.J.; Chen, X. RNA-binding protein RBM24 regulates p63 expression via mRNA stability. Mol. Cancer Res. 2014, 12, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Melino, G. p63 is a suppressor of tumorigenesis and metastasis interacting with mutant p53. Cell Death Differ. 2011, 18, 1487–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kouwenhove, M.; Kedde, M.; Agami, R. MicroRNA regulation by RNA-binding proteins and its implications for cancer. Nat. Rev. Cancer 2011, 11, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.A.; Galardi, S. microRNAs and RNA-binding proteins: A complex network of interactions and reciprocal regulations in cancer. RNA Biol. 2013, 10, 935–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucarull, B.; Tutusaus, A.; Rider, P.; Hernáez-Alsina, T.; Cuño, C.; García de Frutos, P.; Colell, A.; Marí, M.; Morales, A. Hepatocellular carcinoma: Molecular pathogenesis and therapeutic advances. Cancers 2022, 14, 621. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Cao, H.; Zhao, K.; Yao, Y.; Guo, J.; Gao, X.; Yang, Q.; Guo, M.; Zhu, W.; Wang, Y.; Wu, C.; et al. RNA binding protein 24 regulates the translation and replication of hepatitis C virus. Protein Cell 2018, 9, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, B.; Cao, H.; Zhao, K.; Yuan, Y.; Chen, Y.; Zhang, Z.; Wang, Y.; Pei, R.; Chen, J.; et al. RBM24 stabilizes hepatitis B virus pregenomic RNA but inhibits core protein translation by targeting the terminal redundancy sequence. Emerg. Microbes Infect. 2018, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Yang, B.; Chen, Y.; Wang, H.; Hu, X.; Zhou, Y.; Gao, X.; Lu, M.; Niu, J.; Wen, Z.; et al. RNA-binding motif protein 24 (RBM24) is involved in pregenomic RNA packaging by mediating interaction between hepatitis B virus polymerase and the epsilon element. J. Virol. 2019, 93, e02161-18. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global burden of 5 major types of gastrointestinal cancer. Gastroenterology 2020, 159, 335–349.e15. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caramel, J.; Ligier, M.; Puisieux, A. Pleiotropic roles for ZEB1 in cancer. Cancer Res. 2018, 78, 30–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non-small cell lung cancer: Epidemiology, screening, diagnosis, and treatment. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, M.Q.; Ward, C.; Muller, H.K.; Sohal, S.S.; Walters, E.H. Epithelial mesenchymal transition (EMT) and non-small cell lung cancer (NSCLC): A mutual association with airway disease. Med. Oncol. 2017, 34, 45. [Google Scholar] [CrossRef]
- Larsen, J.E.; Nathan, V.; Osborne, J.K.; Farrow, R.K.; Deb, D.; Sullivan, J.P.; Dospoy, P.D.; Augustyn, A.; Hight, S.K.; Sato, M.; et al. ZEB1 drives epithelial-to-mesenchymal transition in lung cancer. J. Clin. Investig. 2016, 126, 3219–3235. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Xia, C.; Zhang, C.; Yang, D.; Ma, K. Prognostic significance of SNCA and its methylation in bladder cancer. BMC Cancer 2022, 22, 330. [Google Scholar] [CrossRef]
- Jin, Y.; Qin, X. Comprehensive analysis of the roles and prognostic value of RNA-binding proteins in head and neck squamous cell carcinoma. DNA Cell Biol. 2020, 39, 1789–1798. [Google Scholar] [CrossRef]
- Yang, R.H.; Liang, B.; Li, J.H.; Pi, X.B.; Yu, K.; Xiang, S.J.; Gu, N.; Chen, X.D.; Zhou, S.T. Identification of a novel tumour microenvironment-based prognostic biomarker in skin cutaneous melanoma. J. Cell. Mol. Med. 2021, 25, 10990–11001. [Google Scholar] [CrossRef]
- Gershanov, S.; Madiwale, S.; Feinberg-Gorenshtein, G.; Vainer, I.; Nehushtan, T.; Michowiz, S.; Goldenberg-Cohen, N.; Birger, Y.; Toledano, H.; Salmon-Divon, M. Classifying medulloblastoma subgroups based on small, clinically achievable gene sets. Front. Oncol. 2021, 11, 637482. [Google Scholar] [CrossRef]
- Li, H.Y.; Bourdelas, A.; Carron, C.; Shi, D.L. The RNA-binding protein Seb4/RBM24 is a direct target of MyoD and is required for myogenesis during Xenopus early development. Mech. Dev. 2010, 127, 281–291. [Google Scholar] [CrossRef]
- Maragh, S.; Miller, R.A.; Bessling, S.L.; McGaughey, D.M.; Wessels, M.W.; de Graaf, B.; Stone, E.A.; Bertoli-Avella, A.M.; Gearhart, J.D.; Fisher, S.; et al. Identification of RNA binding motif proteins essential for cardiovascular development. BMC Dev. Biol. 2011, 11, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, K.L.; Tan, K.T.; Wei, Y.Y.; Ng, C.P.; Colman, A.; Korzh, V.; Xu, X.Q. RNA-binding protein RBM24 is required for sarcomere assembly and heart contractility. Cardiovasc. Res. 2012, 94, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Kong, X.; Zhang, M.; Yang, X.; Xu, X.Q. RNA binding protein 24 deletion disrupts global alternative splicing and causes dilated cardiomyopathy. Protein Cell 2019, 10, 405–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arandel, L.; Polay Espinoza, M.; Matloka, M.; Bazinet, A.; De Dea Diniz, D.; Naouar, N.; Rau, F.; Jollet, A.; Edom-Vovard, F.; Mamchaoui, K.; et al. Immortalized human myotonic dystrophy muscle cell lines to assess therapeutic compounds. Dis. Model Mech. 2017, 10, 487–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.T.; Treacy, D.; Eichinger, K.; Struck, A.; Estabrook, J.; Olafson, H.; Wang, T.T.; Bhatt, K.; Westbrook, T.; Sedehizadeh, S.; et al. Transcriptome alterations in myotonic dystrophy skeletal muscle and heart. Hum. Mol. Genet. 2019, 28, 1312–1321. [Google Scholar] [CrossRef]
- Chamberlain, J.S.; Metzger, J.; Reyes, M.; Townsend, D.; Faulkner, J.A. Dystrophin-deficient mdx mice display a reduced life span and are susceptible to spontaneous rhabdomyosarcoma. FASEB J. 2007, 21, 2195–2204. [Google Scholar] [CrossRef] [Green Version]
- Chandler, E.; Rawson, L.; Debski, R.; McGowan, K.; Lakhotia, A. Rhabdomyosarcoma in a patient with Duchenne muscular dystrophy: A possible association. Child Neurol. Open. 2021, 8, 2329048X211041471. [Google Scholar] [CrossRef]
- Boscolo Sesillo, F.; Fox, D.; Sacco, A. Muscle stem cells give rise to rhabdomyosarcomas in a severe mouse model of Duchenne muscular dystrophy. Cell Rep. 2019, 26, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Kong, X.; Lee, Y.M.; Zhang, M.K.; Guo, L.Y.; Lin, Y.; Lim, T.K.; Lin, Q.; Xu, X.Q. Stk38 modulates Rbm24 protein stability to regulate sarcomere assembly in cardiomyocytes. Sci. Rep. 2017, 7, 44870. [Google Scholar] [CrossRef]
- Cao, Q.; Anyansi, C.; Hu, X.; Xu, L.; Xiong, L.; Tang, W.; Mok, M.T.S.; Cheng, C.; Fan, X.; Gerstein, M.; et al. Reconstruction of enhancer-target networks in 935 samples of human primary cells, tissues and cell lines. Nat. Genet. 2017, 49, 1428–1436. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, B.; Chen, Y.; Huang, D.; Liu, C.; Sun, H.; Hu, X.; Zhou, Y.; Wang, Y.; Chen, J.; et al. RNA-Binding motif protein 38 (RBM38) mediates HBV pgRNA packaging into the nucleocapsid. Antiviral Res. 2022, 198, 105249. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Miranda, G.; Méndez, R. The CPEB-family of proteins, translational control in senescence and cancer. Ageing Res. Rev. 2012, 11, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tsai, Y.H.; Tseng, S.H. Regulation of the expression of cytoplasmic polyadenylation element binding proteins for the treatment of cancer. Anticancer Res. 2016, 36, 5673–5680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaoka, K.; Fujii, K.; Zhang, H.; Usuda, K.; Watanabe, G.; Ivshina, M.; Richter, J.D. CPEB1 mediates epithelial-to-mesenchyme transition and breast cancer metastasis. Oncogene 2016, 35, 2893–2901. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Guijarro, E.; Karras, P.; Cifdaloz, M.; Martínez-Herranz, R.; Cañón, E.; Graña, O.; Horcajada-Reales, C.; Alonso-Curbelo, D.; Calvo, T.G.; Gómez-López, G.; et al. Lineage-specific roles of the cytoplasmic polyadenylation factor CPEB4 in the regulation of melanoma drivers. Nat. Commun. 2016, 7, 13418. [Google Scholar] [CrossRef]
- Shu, L.; Yan, W.; Chen, X. RNPC1, an RNA-binding protein and a target of the p53 family, is required for maintaining the stability of the basal and stress-induced p21 transcript. Genes Dev. 2006, 20, 2961–2972. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Cho, S.J.; Shu, L.; Yan, W.; Guerrero, T.; Kent, M.S.; Skorupski, K.; Chen, H.; Chen, X. Translational repression of p53 by RNPC1, a p53 target overexpressed in lymphomas. Genes Dev. 2011, 25, 1528–1543. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xu, E.; Ren, C.; Yan, W.; Zhang, M.; Chen, M.; Cardiff, R.D.; Imai, D.M.; Wisner, E.; Chen, X. Mice deficient in Rbm38, a target of the p53 family, are susceptible to accelerated aging and spontaneous tumors. Proc. Natl. Acad. Sci. USA 2014, 111, 18637–18642. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xu, E.; Ren, C.; Yang, H.J.; Zhang, Y.; Sun, W.; Kong, X.; Zhang, W.; Chen, M.; Huang, E.; et al. Genetic ablation of Rbm38 promotes lymphomagenesis in the context of mutant p53 by downregulating PTEN. Cancer Res. 2018, 78, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Wan, Y.; He, L.; Zheng, J.H.; Mei, Y.; Shi, J.; Zhang, M.; Dong, Z.; Zhang, D. RBM38 in cancer: Role and mechanism. Cell. Mol. Life Sci. 2021, 78, 117–128. [Google Scholar] [CrossRef]
- Amrane, S.; Rebora, K.; Zniber, I.; Dupuy, D.; Mackereth, C.D. Backbone-independent nucleic acid binding by splicing factor SUP-12 reveals key aspects of molecular recognition. Nat. Commun. 2014, 5, 4595. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cancers | Expression | Activity or Outcome | Target Genes | Pro- or Anti-Tumor | Reference |
|---|---|---|---|---|---|
| HCC | Low | Suppress cell proliferation | β-catenin, p63 | Anti-tumor | [30] |
| Low | Inhibit cancer progression | Downstream of TPRG1-AS1 | Anti-tumor | [31] | |
| High | Promote cell proliferation | Downstream of TRIM56 | Pro-tumor | [33] | |
| LUAD | Low | Inhibit tumor growth and induce apoptosis | Downstream of SMARCA5 | Anti-tumor | [29] |
| High | Decrease chemosensitivity | Unknown | Pro-tumor | [35] | |
| CRC | Low | Repress tumor progression | PTEN | Anti-tumor | [32] |
| NPC | Low | Inhibit NPC growth and viability | miR-25 | Anti-tumor | [28] |
| Bladder cancer | High | Increase tumor size | RUNX1T1 | Pro-tumor | [36] |
| TNBC | High in DFI | Poor prognosis | Unknown | Pro-tumor | [34] |
| HNSCC | High | Low overall survival | Unknown | Pro-tumor | [58] |
| SKCM | High | Low overall survival | Unknown | Pro-tumor | [59] |
| MB (group 4) | High | Biomarker | Unknown | Pro-tumor | [60] |
| RBM24 Partners | RBM24 Regions Involved | Functions | References |
|---|---|---|---|
| eIF4E | eIF4E-binding motif | Prevent the assembly of translation initiation complex | [24] |
| GSK3β | eIF4E-binding motif | Phosphorylate the serine residue within the eIF4E-binding motif | [24,40] |
| Stk38 | N- and C-terminal regions | Reduce RBM24 stability by phosphorylation | [70] |
| RBM38 | C-terminal region | Regulate hepatitis B virus replication | [72] |
| CPEB4 | C-terminal region | Cytoplasmic polyadenylation | [25] |
| PABPC1 | C-terminal region | Cytoplasmic polyadenylation | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, D.-L. RBM24 in the Post-Transcriptional Regulation of Cancer Progression: Anti-Tumor or Pro-Tumor Activity? Cancers 2022, 14, 1843. https://doi.org/10.3390/cancers14071843
Shi D-L. RBM24 in the Post-Transcriptional Regulation of Cancer Progression: Anti-Tumor or Pro-Tumor Activity? Cancers. 2022; 14(7):1843. https://doi.org/10.3390/cancers14071843
Chicago/Turabian StyleShi, De-Li. 2022. "RBM24 in the Post-Transcriptional Regulation of Cancer Progression: Anti-Tumor or Pro-Tumor Activity?" Cancers 14, no. 7: 1843. https://doi.org/10.3390/cancers14071843
APA StyleShi, D. -L. (2022). RBM24 in the Post-Transcriptional Regulation of Cancer Progression: Anti-Tumor or Pro-Tumor Activity? Cancers, 14(7), 1843. https://doi.org/10.3390/cancers14071843