
The Research Advances of Aptamers in Hematologic Malignancies

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Aptamers in Leukemia
2.1. Aptamers in AML
2.1.1. Cell Surface Protein Targeted Aptamers
2.1.2. Other Protein Targeted Aptamers
2.2. Aptamers in ALL
2.2.1. The Role of Aptamers in ALL Diagnoses
2.2.2. The Role of Aptamers in ALL Treatment
2.3. Aptamers in CML
2.4. Aptamers in CLL
3. Aptamers in Lymphoma

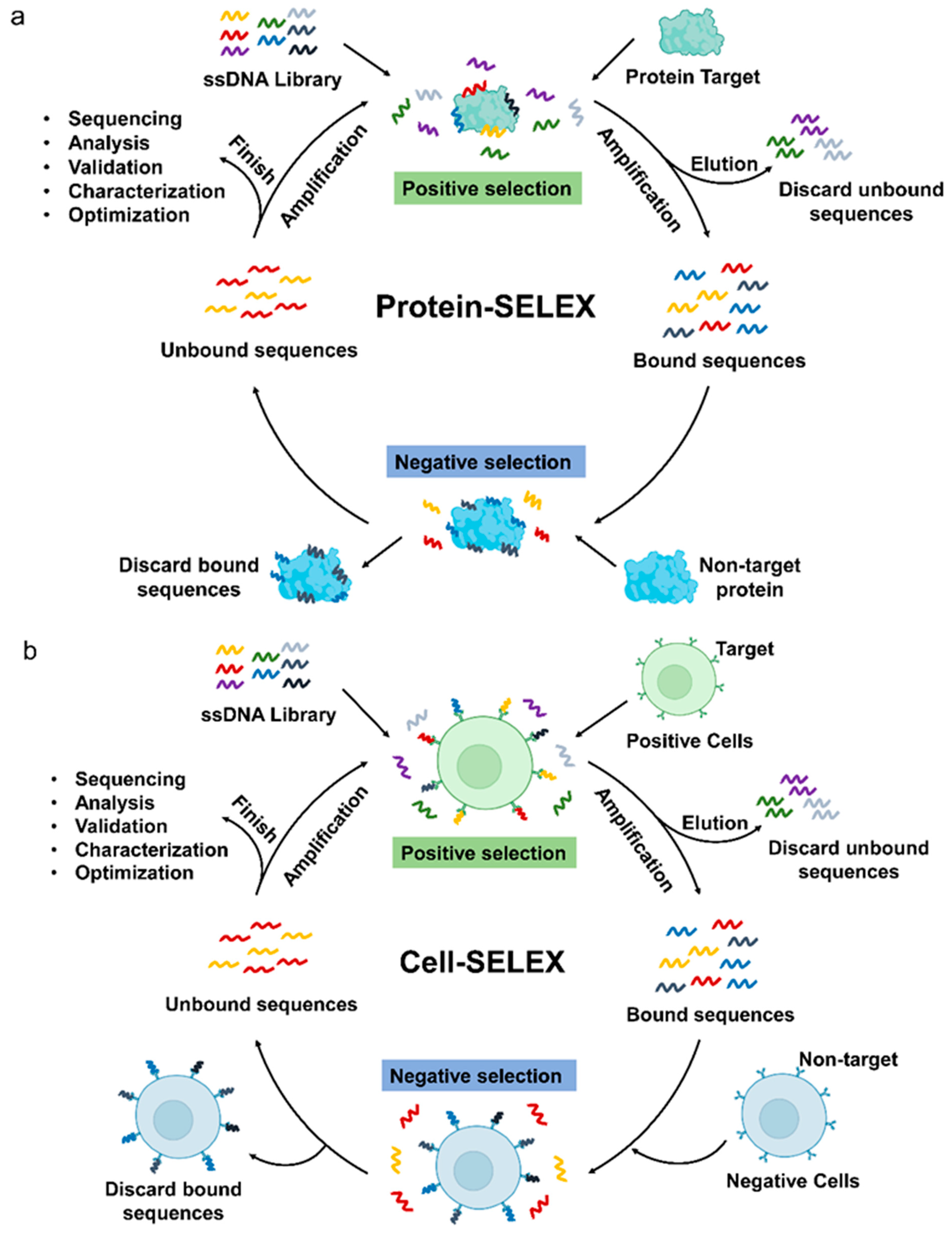{kind=link}
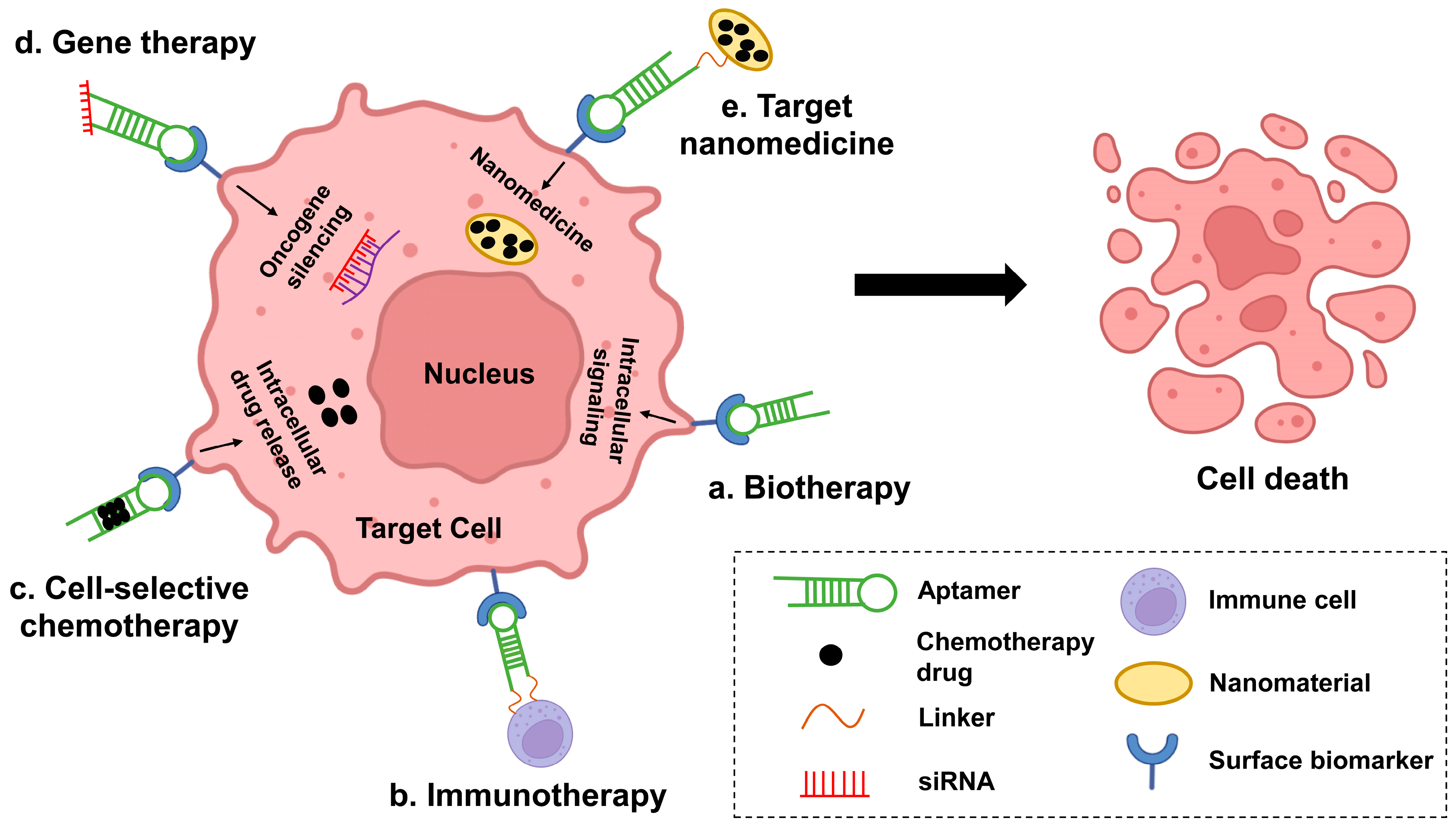{kind=link}
| Name | DNA/RNA | Type of SELEX | Target Cell | Target Molecule | Tested | Kd | Conjugated Molecule | Application | Reference |
|---|---|---|---|---|---|---|---|---|---|
| C2NP | DNA (31nt) | Hybrid SELEX | K299 | CD30 | In vitro | Growth inhibition, apoptosis induction | [127] | ||
| K299 | In vitro; In vivo | ALK siRNA, Dox | Combined cell-selective chemotherapy and oncogene-specific gene therapy. Apoptosis induction and growth inhibition in vitro and prolongation of survival in mice in vivo. | [131] | |||||
| K299, L428 | In vitro | PEG-PLGA NPs, Dox | Apoptosis induction and growth inhibition | [128] | |||||
| K299 | In vitro | S-DNT, T-DNT, Dox | Apoptosis induction and growth inhibition | [129] | |||||
| K299 | In vitro | NK cells | Growth inhibition, apoptosis induction. Immunotherapy | [146] | |||||
| RNA | K299 | In vitro | ALK siRNA | Growth arrest and apoptosis | [130] | ||||
| RNA | K299 | In vitro | dsDNA/Dox ALK siRNA | Combined cell-selective chemotherapy and oncogene-specific gene therapy. Growth inhibition | [132] | ||||
| PS1NP | DNA | Cell-SELEX | HDLM2, K299 | In vitro | 5 ± 0.5 nM | A carrier for in vivo targeted drug delivery | [147] | ||
| LC1 | DNA (59-mer) | Protein-SELEX | Ramos, Raji | CD19 | In vitro | 85.4 nM | Dox | Growth inhibition | [133] |
| TD05 | DNA | Cell-SELEX | Ramos | IGHM, mIgM | In vitro | Identification of potential markers | [134,137] | ||
| R1.2 | DNA | LIGS | BJAB | mIgM | In vitro | 35.5 ± 8.94 nM (4 °C), 65.6 ± 5.88 nM(37 °C) | Identification of potential markers | [144,145] | |
| R-1 | RNA | in vitro SELEX | Jeko-1 | BAFF-R | In vitro | 47 nM | STAT3 siRNA | Growth inhibition | [139] |
| C10.36 | DNA (36-mer) | Ramos | SSC | In vitro | Growth inhibition | [142] | |||
| AP-1 | DNA | Cell-SELEX | HEK293T -CD20 | CD20 | In vitro | 96.91 ± 4.5 nM | [142] | ||
| Raji | In vitro; In vivo | Ag-MOFs, PFK15, RBCm | Apoptosis induction and growth inhibition in vitro and prolongation of survival in mice in vivo. | [143] |
4. Aptamers in Multiple Myeloma
| Name | DNA/RNA | Type of SELEX | Target Cell | Target Molecule | Tested | Kd | Conjugated Molecule | Application | Reference |
|---|---|---|---|---|---|---|---|---|---|
| apt69.T | RNA (50-mer) | Cell-SELEX, Cell-internalizing SELEX | U266, H929 | BCMA | In vitro | 79.4 nM | miR-137, anti-miR-222 | Growth inhibition | [160] |
| wh6 | DNA (80 nt) | Protein-SELEX | MM.1R, MM.1S, ARP-1, ANBL-6 and RMPI-8226 cell | ANXA2 | In vitro; In vivo | 8.75 ± 1.26 nM | Adhesion and proliferation inhibition of MM cell lines in vitro and tumor targeting in vivo. | [166] | |
| TY04 | DNA | Cell-SELEX | MM.1S, NCI-H929, KM3, OPM2 | In vitro | inhibits the growth of multiple myeloma cells via cell cycle arrest | [166] | |||
| SL1 | DNA (50-mer) | Cell-SELEX | CD138+ cells (MM.1S, ARP-1) | c-met | In vitro; In vivo. Ex vivo | 135.6 nM (MM.1S), 237.1 nM (ARP-1) | Inhibition of MM cell growth, migration and adhesion in vitro and tumor targeting in vivo | [168] | |
| #1S | DNA | Hybrid SELEX (cell-based SELEX and protein-based SELEX) | CD38+cells (MM.1S, RPMI8226, MM1R, NCI-H929, Dox40) | CD38 | Dox | Growth inhibition and apoptosis induction in vitro and prolongation of survival in mice in vivo. | [169,170] |
5. Conclusions and Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SELEX | Systematic Evolution of Ligands by Exponential Enrichment |
| ssDNA | single–stranded DNA |
| AML | acute myeloid leukemia |
| ALL | acute lymphoblastic leukemia |
| MM | multiple myeloma |
| CML | chronic myeloid leukemia |
| CLL | Chronic Lymphocytic Leukemia |
| Kd | dissociation constant |
| Dox | doxorubicin |
| MTX | methotrexate |
| OFA/iLRP | Oncofetal antigen/immature laminin receptor protein |
| PTK7 | protein tyrosine kinase 7 |
| NPs | nanoparticles |
| FRET | fluorescence resonance energy transfer |
| MRD | Minimal residual disease |
| RCA | rolling cycle amplification |
| TPA | two-photon absorption |
| QCM | quartz crystal microbalance |
| AuNP | gold nanoparticle |
| PEC | photoelectrochemical |
| BP NS | black phosphorus nanosheets |
| Dau | Daunorubicin |
| PA | polyvalent aptamers |
| SWNTs | Single-walled carbon nanotubes |
| VCR | vincristine |
| TKI | tyrosine kinase inhibitors |
| HL | Hodgkin’s lymphoma |
| NHL | non-Hodgkin’s lymphoma |
| ALCL | anaplastic large cell lymphoma |
| ALK | anaplastic lymphoma kinase |
| BAFF-R | B-cell activating factor receptor |
| SSC | spliceosome complex |
| LIGS | LIgand-Guided-Selection |
| NK | natural killer |
| PC | plasma cells |
| BCMA | B-cell maturation antigen |
| CAM-DR | Cell adhesion-mediated drug resistance |
References
- Uribe-Herranz, M.; Klein-Gonzalez, N.; Rodriguez-Lobato, L.G.; Juan, M.; de Larrea, C.F. Gut Microbiota Influence in Hematological Malignancies: From Genesis to Cure. Int. J. Mol. Sci. 2021, 22, 1026. [Google Scholar] [CrossRef] [PubMed]
- Kansara, R.R.; Speziali, C. Immunotherapy in hematologic malignancies. Curr. Oncol. 2020, 27, S124–S131. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, K.; Rizvi, S.; Syed, B.A. Non-Hodgkin lymphoma therapy landscape. Nat. Rev. Drug Discov. 2019, 18, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.C.; Jones, J.J.; Woyach, J.A.; Johnson, A.J.; Flynn, J.M. Entering the era of targeted therapy for chronic lymphocytic leukemia: Impact on the practicing clinician. J. Clin. Oncol. 2014, 32, 3039–3047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Wang, M.; Wang, H.; Hu, D.; Zipfel, P.F.; Hu, Y. How Does Complement Affect Hematological Malignancies: From Basic Mechanisms to Clinical Application. Front. Immunol. 2020, 11, 593610. [Google Scholar] [CrossRef]
- Ye, M.; Hu, J.; Peng, M.; Liu, J.; Liu, J.; Liu, H.; Zhao, X.; Tan, W. Generating aptamers by cell-SELEX for applications in molecular medicine. Int. J. Mol. Sci. 2012, 13, 3341–3353. [Google Scholar] [CrossRef] [Green Version]
- Gold, L. The SELEX process: A surprising source of therapeutic and diagnostic compounds. Harvey Lect. 1995, 91, 47–57. [Google Scholar]
- Famulok, M.; Mayer, G.; Blind, M. Nucleic acid aptamers-from selection in vitro to applications in vivo. Acc. Chem. Res. 2000, 33, 591–599. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhao, J.; Li, J.; Liao, X.; Chen, F. Advances of aptamers screened by Cell-SELEX in selection procedure, cancer diagnostics and therapeutics. Anal. Biochem. 2020, 598, 113620. [Google Scholar] [CrossRef]
- Georges, J.F.; Liu, X.; Eschbacher, J.; Nichols, J.; Mooney, M.A.; Joy, A.; Spetzler, R.F.; Feuerstein, B.G.; Preul, M.C.; Anderson, T.; et al. Use of a conformational switching aptamer for rapid and specific ex vivo identification of central nervous system lymphoma in a xenograft model. PLoS ONE 2015, 10, e0123607. [Google Scholar] [CrossRef] [Green Version]
- Rahimizadeh, K.; AlShamaileh, H.; Fratini, M.; Chakravarthy, M.; Stephen, M.; Shigdar, S.; Veedu, R.N. Development of Cell-Specific Aptamers: Recent Advances and Insight into the Selection Procedures. Molecules 2017, 22, 2070. [Google Scholar] [CrossRef] [PubMed]
- Hoppe-Seyler, F.; Crnkovic-Mertens, I.; Tomai, E.; Butz, K. Peptide aptamers: Specific inhibitors of protein function. Curr. Mol. Med. 2004, 4, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Gelinas, A.D.; Davies, D.R.; Janjic, N. Embracing proteins: Structural themes in aptamer-protein complexes. Curr. Opin. Struct. Biol. 2016, 36, 122–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, G. The chemical biology of aptamers. Angew. Chem. Int. Ed. Engl. 2009, 48, 2672–2689. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Kim, J.; Easley, C.J. Quantifying Aptamer-Protein Binding via Thermofluorimetric Analysis. Anal. Methods 2015, 7, 7358–7362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Shangguan, D.; Wang, K.; Shi, H.; Sefah, K.; Mallikratchy, P.; Chen, H.W.; Li, Y.; Tan, W. Selection of aptamers for molecular recognition and characterization of cancer cells. Anal. Chem. 2007, 79, 4900–4907. [Google Scholar] [CrossRef]
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Rossi, J. Aptamers as targeted therapeutics: Current potential and challenges. Nat. Rev. Drug Discov. 2017, 16, 181–202. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Yigit, M.V.; Mazumdar, D.; Lu, Y. Molecular diagnostic and drug delivery agents based on aptamer-nanomaterial conjugates. Adv. Drug Deliv. Rev. 2010, 62, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Vorobyeva, M.A.; Davydova, A.S.; Vorobjev, P.E.; Venyaminova, A.G. Key Aspects of Nucleic Acid Library Design for in Vitro Selection. Int. J. Mol. Sci. 2018, 19, 470. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.; Yang, S. Replacing antibodies with aptamers in lateral flow immunoassay. Biosens. Bioelectron. 2015, 71, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Allemailem, K.S.; Almatroudi, A.; Alsahli, M.A.; Basfar, G.T.; Alrumaihi, F.; Rahmani, A.H.; Khan, A.A. Recent advances in understanding oligonucleotide aptamers and their applications as therapeutic agents. 3 Biotech 2020, 10, 551. [Google Scholar] [CrossRef] [PubMed]
- Drabik, A.; Ner-Kluza, J.; Hartman, K.; Mayer, G.; Silberring, J. Changes in Protein Glycosylation as a Result of Aptamer Interactions with Cancer Cells. Proteom. Clin. Appl. 2020, 14, e1800186. [Google Scholar] [CrossRef]
- Ma, H.; Liu, J.; Ali, M.M.; Mahmood, M.A.; Labanieh, L.; Lu, M.; Iqbal, S.M.; Zhang, Q.; Zhao, W.; Wan, Y. Nucleic acid aptamers in cancer research, diagnosis and therapy. Chem. Soc. Rev. 2015, 44, 1240–1256. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Yazdian-Robati, R.; Arab, A.; Ramezani, M.; Abnous, K.; Taghdisi, S.M. Application of aptamers in treatment and diagnosis of leukemia. Int. J. Pharm. 2017, 529, 44–54. [Google Scholar] [CrossRef]
- Fang, X.; Tan, W. Aptamers generated from cell-SELEX for molecular medicine: A chemical biology approach. Acc. Chem. Res. 2010, 43, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xu, S.; Yan, H.; Li, X.; Yazd, H.S.; Li, X.; Huang, T.; Cui, C.; Jiang, J.; Tan, W. Nucleic Acid Aptamers for Molecular Diagnostics and Therapeutics: Advances and Perspectives. Angew. Chem. Int. Ed. Engl. 2021, 60, 2221–2231. [Google Scholar] [CrossRef]
- Rodríguez-García, A.; García-Vicente, R.; Morales, M.L.; Ortiz-Ruiz, A.; Martínez-López, J.; Linares, M. Protein Carbonylation and Lipid Peroxidation in Hematological Malignancies. Antioxidants 2020, 9, 1212. [Google Scholar] [CrossRef]
- Laing, A.A.; Harrison, C.J.; Gibson, B.E.S.; Keeshan, K. Unlocking the potential of anti-CD33 therapy in adult and childhood acute myeloid leukemia. Exp. Hematol. 2017, 54, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Saultz, J.N.; Garzon, R. Acute Myeloid Leukemia: A Concise Review. J. Clin. Med. 2016, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Goolsby, C.; Yaseen, N.R. NUP98-HOXA9 induces long-term proliferation and blocks differentiation of primary human CD34+ hematopoietic cells. Cancer Res. 2006, 66, 6628–6637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmone, A.; Amorim, M.; Pontier, A.L.; Wang, S.; Jablonski, E.; Sipkins, D.A. Leukemic cells create bone marrow niches that disrupt the behavior of normal hematopoietic progenitor cells. Science 2008, 322, 1861–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disease, G.B.D.; Injury, I.; Prevalence, C. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar] [CrossRef] [Green Version]
- Short, N.J.; Ravandi, F. The safety and efficacy of vosaroxin in patients with first relapsed or refractory acute myeloid leukemia-a critical review. Expert Rev. Hematol. 2016, 9, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Ho, M.; Pastan, I. New immunotoxins targeting CD123, a stem cell antigen on acute myeloid leukemia cells. J. Immunother. 2007, 30, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Pulte, D.; Gondos, A.; Brenner, H. Expected long-term survival of patients diagnosed with acute myeloblastic leukemia during 2006–2010. Ann. Oncol. 2010, 21, 335–341. [Google Scholar] [CrossRef]
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Zhu, M.; Huang, Y.; Wei, S.; Xie, J.; Xiao, Y. CD123 and its potential clinical application in leukemias. Life Sci. 2015, 122, 59–64. [Google Scholar] [CrossRef]
- Mardiros, A.; Forman, S.J.; Budde, L.E. T cells expressing CD123 chimeric antigen receptors for treatment of acute myeloid leukemia. Curr. Opin. Hematol. 2015, 22, 484–488. [Google Scholar] [CrossRef]
- Li, B.; Zhao, W.; Zhang, X.; Wang, J.; Luo, X.; Baker, S.D.; Jordan, C.T.; Dong, Y. Design, synthesis and evaluation of anti-CD123 antibody drug conjugates. Bioorg. Med. Chem. 2016, 24, 5855–5860. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Gao, H.; Zhang, Q. The biomarkers of leukemia stem cells in acute myeloid leukemia. Stem. Cell Investig. 2017, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Wang, M.; Dai, B.; Zhang, Y.; Yang, Y.; Li, Q.; Duan, M.; Zhang, X.; Wang, X.; Li, A.; et al. Novel CD123-aptamer-originated targeted drug trains for selectively delivering cytotoxic agent to tumor cells in acute myeloid leukemia theranostics. Drug Deliv. 2017, 24, 1216–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wu, H.; Duan, M.; Yang, Y.; Wang, G.; Che, F.; Liu, B.; He, W.; Li, Q.; Zhang, L. SS30, a novel thioaptamer targeting CD123, inhibits the growth of acute myeloid leukemia cells. Life Sci. 2019, 232, 116663. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, L.; Zhu, Z.; Ding, C.; Chen, S.; Liu, R.; Fan, H.; Chen, Y.; Li, H. Novel CD123 polyaptamer hydrogel edited by Cas9/sgRNA for AML-targeted therapy. Drug Deliv. 2021, 28, 1166–1178. [Google Scholar] [CrossRef]
- Hans, C.P.; Finn, W.G.; Singleton, T.P.; Schnitzer, B.; Ross, C.W. Usefulness of anti-CD117 in the flow cytometric analysis of acute leukemia. Am. J. Clin. Pathol. 2002, 117, 301–305. [Google Scholar] [CrossRef]
- Doepfner, K.T.; Boller, D.; Arcaro, A. Targeting receptor tyrosine kinase signaling in acute myeloid leukemia. Crit. Rev. Oncol. Hematol. 2007, 63, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Stirewalt, D.L.; Meshinchi, S. Receptor tyrosine kinase alterations in AML-biology and therapy. Cancer Treat. Res. 2010, 145, 85–108. [Google Scholar] [CrossRef]
- Marcucci, G.; Haferlach, T.; Dohner, H. Molecular genetics of adult acute myeloid leukemia: Prognostic and therapeutic implications. J. Clin. Oncol. 2011, 29, 475–486. [Google Scholar] [CrossRef]
- Ashman, L.K.; Griffith, R. Therapeutic targeting of c-KIT in cancer. Expert Opin. Investig. Drugs. 2013, 22, 103–115. [Google Scholar] [CrossRef]
- Liang, J.; Wu, Y.L.; Chen, B.J.; Zhang, W.; Tanaka, Y.; Sugiyama, H. The C-kit receptor-mediated signal transduction and tumor-related diseases. Int. J. Biol. Sci. 2013, 9, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Pei, S.N.; Qi, J.; Zeng, Z.; Iyer, S.P.; Lin, P.; Tung, C.H.; Zu, Y. Oligonucleotide aptamer-drug conjugates for targeted therapy of acute myeloid leukemia. Biomaterials 2015, 67, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, S.; Wagner, A.; Kabelitz, D.; Marget, M.; Coggin, J., Jr.; Barsoum, A.; Rohrer, J.; Schmitz, N.; Zeis, M. Induction of cytotoxic T-cell responses against the oncofetal antigen-immature laminin receptor for the treatment of hematologic malignancies. Blood 2003, 102, 4416–4423. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Hu, Y.; Li, X.; Li, Z.; Duan, J.; Yang, X.D. Selection of a novel DNA aptamer against OFA/iLRP for targeted delivery of doxorubicin to AML cells. Sci. Rep. 2019, 9, 7343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godwin, C.D.; Gale, R.P.; Walter, R.B. Gemtuzumab ozogamicin in acute myeloid leukemia. Leukemia 2017, 31, 1855–1868. [Google Scholar] [CrossRef]
- Yang, C.; Wang, Y.; Ge, M.H.; Fu, Y.J.; Hao, R.; Islam, K.; Huang, P.; Chen, F.; Sun, J.; Hong, F.; et al. Rapid identification of specific DNA aptamers precisely targeting CD33 positive leukemia cells through a paired cell-based approach. Biomater. Sci. 2019, 7, 938–950. [Google Scholar] [CrossRef]
- Zaimy, M.A.; Jebali, A.; Bazrafshan, B.; Mehrtashfar, S.; Shabani, S.; Tavakoli, A.; Hekmatimoghaddam, S.H.; Sarli, A.; Azizi, H.; Izadi, P.; et al. Coinhibition of overexpressed genes in acute myeloid leukemia subtype M2 by gold nanoparticles functionalized with five antisense oligonucleotides and one anti-CD33(+)/CD34(+) aptamer. Cancer Gene Ther. 2016, 23, 315–320. [Google Scholar] [CrossRef]
- Earnest, K.G.; McConnell, E.M.; Hassan, E.M.; Wunderlich, M.; Hosseinpour, B.; Bono, B.S.; Chee, M.J.; Mulloy, J.C.; Willmore, W.G.; DeRosa, M.C.; et al. Development and characterization of a DNA aptamer for MLL-AF9 expressing acute myeloid leukemia cells using whole cell-SELEX. Sci. Rep. 2021, 11, 19174. [Google Scholar] [CrossRef]
- Soundararajan, S.; Wang, L.; Sridharan, V.; Chen, W.; Courtenay-Luck, N.; Jones, D.; Spicer, E.K.; Fernandes, D.J. Plasma membrane nucleolin is a receptor for the anticancer aptamer AS1411 in MV4-11 leukemia cells. Mol. Pharmacol. 2009, 76, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Mongelard, F.; Bouvet, P. AS-1411, a guanosine-rich oligonucleotide aptamer targeting nucleolin for the potential treatment of cancer, including acute myeloid leukemia. Curr. Opin. Mol. Ther. 2010, 12, 107–114. [Google Scholar]
- Deng, R.; Shen, N.; Yang, Y.; Yu, H.; Xu, S.; Yang, Y.W.; Liu, S.; Meguellati, K.; Yan, F. Targeting epigenetic pathway with gold nanoparticles for acute myeloid leukemia therapy. Biomaterials 2018, 167, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Hales, E.C.; Taub, J.W.; Matherly, L.H. New insights into Notch1 regulation of the PI3K-AKT-mTOR1 signaling axis: Targeted therapy of gamma-secretase inhibitor resistant T-cell acute lymphoblastic leukemia. Cell Signal. 2014, 26, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Samad, A.; Pombo-de-Oliveira, M.S.; Scelo, G.; Smith, M.T.; Feusner, J.; Wiemels, J.L.; Metayer, C. Global characteristics of childhood acute promyelocytic leukemia. Blood Rev. 2015, 29, 101–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robison, L.L.; Bhatia, S. Late-effects among survivors of leukaemia and lymphoma during childhood and adolescence. Br. J. Haematol. 2003, 122, 345–359. [Google Scholar] [CrossRef]
- Hirsch, L.R.; Stafford, R.J.; Bankson, J.A.; Sershen, S.R.; Rivera, B.; Price, R.E.; Hazle, J.D.; Halas, N.J.; West, J.L. Nanoshell-mediated near-infrared thermal therapy of tumors under magnetic resonance guidance. Proc. Natl. Acad. Sci. USA 2003, 100, 13549–13554. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wang, C.; Cheng, L.; He, W.; Cheng, Z.; Liu, Z. Protein modified upconversion nanoparticles for imaging-guided combined photothermal and photodynamic therapy. Biomaterials 2014, 35, 2915–2923. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, J.; Deng, L.; Ma, C.; Li, J.; Li, Y.; Yang, S.; Yang, J.; Wang, J.; Yang, R. Targeted Intracellular Controlled Drug Delivery and Tumor Therapy through in Situ Forming Ag Nanogates on Mesoporous Silica Nanocontainers. ACS Appl. Mater. Interfaces 2015, 7, 11930–11938. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, M.; Yue, B.; Yang, M.; Carter, C.; Al-Quran, S.Z.; Li, B.; Li, Y. PTK7: A new biomarker for immunophenotypic characterization of maturing T cells and T cell acute lymphoblastic leukemia. Leuk Res. 2012, 36, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Shangguan, D.; Cao, Z.; Fang, X.; Tan, W. Cell-specific internalization study of an aptamer from whole cell selection. Chemistry 2008, 14, 1769–1775. [Google Scholar] [CrossRef]
- Huang, Y.F.; Shangguan, D.; Liu, H.; Phillips, J.A.; Zhang, X.; Chen, Y.; Tan, W. Molecular assembly of an aptamer-drug conjugate for targeted drug delivery to tumor cells. Chembiochem 2009, 10, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Yang, N.; Zhong, L.; Luo, Y.; Wang, H.; Gong, W.; Zhou, S.; Li, Y.; He, J.; Cao, H.; et al. A novel label-free terbium(iii)-aptamer based aptasensor for ultrasensitive and highly specific detection of acute lymphoma leukemia cells. Analyst 2019, 144, 3843–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahreyni, A.; Yazdian-Robati, R.; Ramezani, M.; Rasouli, M.; Alinezhad Nameghi, M.; Alibolandi, M.; Abnous, K.; Taghdisi, S.M. Identification and imaging of leukemia cells using dual-aptamer-functionalized graphene oxide complex. J. Biomater. Appl. 2017, 32, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, H.; Xu, D. Silver decahedral nanoparticles-enhanced fluorescence resonance energy transfer sensor for specific cell imaging. Anal. Chem. 2015, 87, 3826–3833. [Google Scholar] [CrossRef] [PubMed]
- Grechkin, Y.A.; Grechkina, S.L.; Zaripov, E.A.; Fedorenko, S.V.; Mustafina, A.R.; Berezovski, M.V. Aptamer-Conjugated Tb(III)-Doped Silica Nanoparticles for Luminescent Detection of Leukemia Cells. Biomedicines 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, O.; Weiss, I.D.; Wang, L.; Wang, Z.; Yang, X.; Dewhurst, A.; Ma, Y.; Zhu, G.; Niu, G.; Kiesewetter, D.O.; et al. 18F-Labeled Single-Stranded DNA Aptamer for PET Imaging of Protein Tyrosine Kinase-7 Expression. J. Nucl. Med. 2015, 56, 1780–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, M.C.; O’Donoghue, M.B.; Chen, X.L.; Tan, W.H. Highly Fluorescent Dye-Doped Silica Nanoparticles Increase Flow Cytometry Sensitivity for Cancer Cell Monitoring. Nano Res. 2009, 2, 448–461. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.S.; Shi, H.; He, X.X.; Wang, K.M.; He, D.G.; Yan, L.A.; Xu, F.Z.; Lei, Y.L.; Tang, J.L.; Yu, Y.R. Iodide-Responsive Cu-Au Nanoparticle-Based Colorimetric Platform for Ultrasensitive Detection of Target Cancer Cells. Anal. Chem. 2015, 87, 7141–7147. [Google Scholar] [CrossRef]
- Khoshfetrat, S.M.; Mehrgardi, M.A. Amplified detection of leukemia cancer cells using an aptamer-conjugated gold-coated magnetic nanoparticles on a nitrogen-doped graphene modified electrode. Bioelectrochemistry 2017, 114, 24–32. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Du, Y.; Zhu, Z.; Zhang, M.; Lv, Z.; Wu, L.; Yang, Y.; Li, A.; Yang, L.; et al. Highly Sensitive Minimal Residual Disease Detection by Biomimetic Multivalent Aptamer Nanoclimber Functionalized Microfluidic Chip. Small 2020, 16, e2000949. [Google Scholar] [CrossRef]
- Li, X.; Zhou, B.; Zhao, Z.; Hu, Z.; Zhou, S.; Yang, N.; Huang, Y.; Zhang, Z.; Su, J.; Lan, D.; et al. A Smart Detection System Based on Specific Magnetic and Rolling Cycle Amplification Signal-Amplified Dual-Aptamers to Accurately Monitor Minimal Residual Diseases in Patients with T-ALL. J. Biomed. Nanotechnol. 2016, 12, 2151–2160. [Google Scholar] [CrossRef]
- Yan, H.; Ren, W.; Liu, S.; Yu, Y. Two-photon imaging of aptamer-functionalized Copolymer/TPdye fluorescent organic dots targeted to cancer cells. Anal. Chim. Acta 2020, 1106, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Guo, M.; Nie, Z.; Huang, Y.; Pan, C.; Zeng, K.; Zhang, Y.; Yao, S. Selective collection and detection of leukemia cells on a magnet-quartz crystal microbalance system using aptamer-conjugated magnetic beads. Biosens. Bioelectron. 2010, 25, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Pan, Y.; Fang, H.; Guo, M.; Nie, Z.; Huang, Y.; Yao, S. An aptamer-based quartz crystal microbalance biosensor for sensitive and selective detection of leukemia cells using silver-enhanced gold nanoparticle label. Talanta 2014, 126, 130–135. [Google Scholar] [CrossRef]
- Hu, X.; Wang, Y.; Zuping, X.; Song, P.; Wang, A.J.; Qian, Z.; Yuan, P.X.; Zhao, T.; Feng, J.J. Novel Aggregation-Enhanced PEC Photosensitizer Based on Electrostatic Linkage of Ionic Liquid with Protoporphyrin IX for Ultrasensitive Detection of Molt-4 Cells. Anal. Chem. 2022, 94, 3708–3717. [Google Scholar] [CrossRef] [PubMed]
- Mazloum-Ardakani, M.; Barazesh, B.; Moshtaghiun, S.M. A distinguished cancer-screening package containing a DNA sensor and an aptasensor for early and certain detection of acute lymphoblastic leukemia. Clin. Chim. Acta 2019, 497, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ma, W.; Li, Q.; Zhao, D.; Shao, X.; Huang, Q.; Hao, L.; Lin, Y. Aptamer-targeted DNA nanostructures with doxorubicin to treat protein tyrosine kinase 7-positive tumours. Cell Prolif. 2019, 52, e12511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ma, Y.; Xie, Y.; An, Y.; Huang, Y.; Zhu, Z.; Yang, C.J. A controllable aptamer-based self-assembled DNA dendrimer for high affinity targeting, bioimaging and drug delivery. Sci. Rep. 2015, 5, 10099. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.L.; Shiao, Y.S.; Huang, Y.F. Release of photoactivatable drugs from plasmonic nanoparticles for targeted cancer therapy. ACS Nano 2011, 5, 7796–7804. [Google Scholar] [CrossRef]
- Zong, S.; Wang, L.; Yang, Z.; Wang, H.; Wang, Z.; Cui, Y. Black Phosphorus-Based Drug Nanocarrier for Targeted and Synergetic Chemophotothermal Therapy of Acute Lymphoblastic Leukemia. ACS Appl. Mater. Interfaces 2019, 11, 5896–5902. [Google Scholar] [CrossRef]
- Taghdisi, S.M.; Abnous, K.; Mosaffa, F.; Behravan, J. Targeted delivery of daunorubicin to T-cell acute lymphoblastic leukemia by aptamer. J. Drug Target 2010, 18, 277–281. [Google Scholar] [CrossRef]
- Taghdisi, S.M.; Danesh, N.M.; Lavaee, P.; Emrani, A.S.; Hassanabad, K.Y.; Ramezani, M.; Abnous, K. Double targeting, controlled release and reversible delivery of daunorubicin to cancer cells by polyvalent aptamers-modified gold nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 61, 753–761. [Google Scholar] [CrossRef]
- Danesh, N.M.; Lavaee, P.; Ramezani, M.; Abnous, K.; Taghdisi, S.M. Targeted and controlled release delivery of daunorubicin to T-cell acute lymphoblastic leukemia by aptamer-modified gold nanoparticles. Int. J. Pharm. 2015, 489, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Taghdisi, S.M.; Lavaee, P.; Ramezani, M.; Abnous, K. Reversible targeting and controlled release delivery of daunorubicin to cancer cells by aptamer-wrapped carbon nanotubes. Eur. J. Pharm. Biopharm. 2011, 77, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Wang, X.; Sun, Y.; Fan, R.; Liu, Z.; Guo, R.; Xie, D. Sgc8 aptamer targeted glutathione-responsive nanoassemblies containing Ara-C prodrug for the treatment of acute lymphoblastic leukemia. Nanoscale 2019, 11, 23000–23012. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Yu, Y.; Lai, C.; Wang, D.; Wang, Y.; Xue, D.; Hu, Z.; Lu, X. Vincristine-Loaded and sgc8-Modified Liposome as a Potential Targeted Drug Delivery System for Treating Acute Lymphoblastic Leukemia. J. Biomed Nanotechnol. 2018, 14, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tang, C.; Hammink, R.; Nelissen, F.H.T.; Heus, H.A.; Kouwer, P.H.J. Multivalent Sgc8c-aptamer decorated polymer scaffolds for leukemia targeting. Chem. Commun. 2021, 57, 2744–2747. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Wang, D.; Long, C.Y.; Li, S.H.; Wang, X.Q.; Tan, W.H. Regulating the Anticancer Efficacy of Sgc8-Combretastatin A4 Conjugates: A Case of Recognizing the Significance of Linker Chemistry for the Design of Aptamer-Based Targeted Drug Delivery Strategies. J. Am. Chem. Soc. 2021, 143, 8559–8564. [Google Scholar] [CrossRef]
- Krenn, P.W.; Koschmieder, S.; Fassler, R. Kindlin-3 loss curbs chronic myeloid leukemia in mice by mobilizing leukemic stem cells from protective bone marrow niches. Proc. Natl. Acad. Sci. USA 2020, 117, 24326–24335. [Google Scholar] [CrossRef]
- Slupianek, A.; Dasgupta, Y.; Ren, S.Y.; Gurdek, E.; Donlin, M.; Nieborowska-Skorska, M.; Fleury, F.; Skorski, T. Targeting RAD51 phosphotyrosine-315 to prevent unfaithful recombination repair in BCR-ABL1 leukemia. Blood 2011, 118, 1062–1068. [Google Scholar] [CrossRef] [Green Version]
- Ping, J.; Shen, Z.H.; Wang, B.Q.; Zhao, N.; Li, R.; Li, M.; Pang, X.B.; Chen, C.B. [Effects of Aptamer-siRNA Nucleic Acid Compound on Growth and Apoptosis in Myeloid Leukemia Cell Line K562]. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2015, 23, 381–385. [Google Scholar] [CrossRef]
- Weisberg, E.L.; Sattler, M.; Azab, A.K.; Eulberg, D.; Kruschinski, A.; Manley, P.W.; Stone, R.; Griffin, J.D. Inhibition of SDF-1-induced migration of oncogene-driven myeloid leukemia by the L-RNA aptamer (Spiegelmer), NOX-A12, and potentiation of tyrosine kinase inhibition. Oncotarget 2017, 8, 109973–109984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallek, M. Chronic lymphocytic leukemia: 2017 update on diagnosis, risk stratification, and treatment. Am. J. Hematol. 2017, 92, 946–965. [Google Scholar] [CrossRef] [PubMed]
- Filip, A.A.; Grenda, A.; Popek, S.; Koczkodaj, D.; Michalak-Wojnowska, M.; Budzynski, M.; Wasik-Szczepanek, E.; Zmorzynski, S.; Karczmarczyk, A.; Giannopoulos, K. Expression of circulating miRNAs associated with lymphocyte differentiation and activation in CLL-another piece in the puzzle. Ann. Hematol. 2017, 96, 33–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenz, T.; Mertens, D.; Kuppers, R.; Dohner, H.; Stilgenbauer, S. From pathogenesis to treatment of chronic lymphocytic leukaemia. Nat. Rev. Cancer 2010, 10, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Kipps, T.J.; Stevenson, F.K.; Wu, C.J.; Croce, C.M.; Packham, G.; Wierda, W.G.; O’Brien, S.; Gribben, J.; Rai, K. Chronic lymphocytic leukaemia. Nat. Rev. Dis. Primers 2017, 3, 16096. [Google Scholar] [CrossRef] [Green Version]
- Rai, K.R.; Jain, P. Chronic lymphocytic leukemia (CLL)-Then and now. Am. J. Hematol. 2016, 91, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Stefaniuk, P.; Onyszczuk, J.; Szymczyk, A.; Podhorecka, M. Therapeutic Options for Patients with TP53 Deficient Chronic Lymphocytic Leukemia: Narrative Review. Cancer Manag. Res. 2021, 13, 1459–1476. [Google Scholar] [CrossRef]
- Rodríguez-Vicente, A.E.; Díaz, M.G.; Hernández-Rivas, J.M. Chronic lymphocytic leukemia: A clinical and molecular heterogenous disease. Cancer Genet. 2013, 206, 49–62. [Google Scholar] [CrossRef]
- Pekarsky, Y.; Zanesi, N.; Croce, C.M. Molecular basis of CLL. Semin. Cancer Biol. 2010, 20, 370–376. [Google Scholar] [CrossRef]
- Dapas, B.; Pozzato, G.; Zorzet, S.; Capolla, S.; Macor, P.; Scaggiante, B.; Coan, M.; Guerra, C.; Gnan, C.; Gattei, V.; et al. Effects of eEF1A1 targeting by aptamer/siRNA in chronic lymphocytic leukaemia cells. Int. J. Pharm. 2020, 574, 118895. [Google Scholar] [CrossRef]
- Scaggiante, B.; Dapas, B.; Bonin, S.; Grassi, M.; Zennaro, C.; Farra, R.; Cristiano, L.; Siracusano, S.; Zanconati, F.; Giansante, C.; et al. Dissecting the expression of EEF1A1/2 genes in human prostate cancer cells: The potential of EEF1A2 as a hallmark for prostate transformation and progression. Br. J. Cancer 2012, 106, 166–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scaggiante, B.; Dapas, B.; Pozzato, G.; Grassi, G. The more basic isoform of eEF1A relates to tumour cell phenotype and is modulated by hyper-proliferative/differentiating stimuli in normal lymphocytes and CCRF-CEM T-lymphoblasts. Hematol. Oncol. 2013, 31, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Scaggiante, B.; Farra, R.; Dapas, B.; Baj, G.; Pozzato, G.; Grassi, M.; Zanconati, F.; Grassi, G. Aptamer targeting of the elongation factor 1A impairs hepatocarcinoma cells viability and potentiates bortezomib and idarubicin effects. Int. J. Pharm. 2016, 506, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calissano, C.; Damle, R.N.; Marsilio, S.; Yan, X.J.; Yancopoulos, S.; Hayes, G.; Emson, C.; Murphy, E.J.; Hellerstein, M.K.; Sison, C.; et al. Intraclonal complexity in chronic lymphocytic leukemia: Fractions enriched in recently born/divided and older/quiescent cells. Mol. Med. 2011, 17, 1374–1382. [Google Scholar] [CrossRef]
- Steurer, M.; Montillo, M.; Scarfo, L.; Mauro, F.R.; Andel, J.; Wildner, S.; Trentin, L.; Janssens, A.; Burgstaller, S.; Fromming, A.; et al. Olaptesed pegol (NOX-A12) with bendamustine and rituximab: A phase IIa study in patients with relapsed/refractory chronic lymphocytic leukemia. Haematologica 2019, 104, 2053–2060. [Google Scholar] [CrossRef] [Green Version]
- Hoellenriegel, J.; Zboralski, D.; Maasch, C.; Rosin, N.Y.; Wierda, W.G.; Keating, M.J.; Kruschinski, A.; Burger, J.A. The Spiegelmer NOX-A12, a novel CXCL12 inhibitor, interferes with chronic lymphocytic leukemia cell motility and causes chemosensitization. Blood 2014, 123, 1032–1039. [Google Scholar] [CrossRef]
- Yang, M.; Jiang, G.; Li, W.; Qiu, K.; Zhang, M.; Carter, C.M.; Al-Quran, S.Z.; Li, Y. Developing aptamer probes for acute myelogenous leukemia detection and surface protein biomarker discovery. J. Hematol. Oncol. 2014, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Lu, L.; Kang, T.S.; Mergny, J.L.; Leung, C.H.; Ma, D.L. Interaction of an Iridium(III) Complex with G-Quadruplex DNA and Its Application in Luminescent Switch-On Detection of Siglec-5. Anal. Chem. 2016, 88, 10290–10295. [Google Scholar] [CrossRef]
- Sun, R.; Medeiros, L.J.; Young, K.H. Diagnostic and predictive biomarkers for lymphoma diagnosis and treatment in the era of precision medicine. Mod. Pathol. 2016, 29, 1118–1142. [Google Scholar] [CrossRef] [Green Version]
- Shankland, K.R.; Armitage, J.O.; Hancock, B.W. Non-Hodgkin lymphoma. Lancet 2012, 380, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.O.; Gascoyne, R.D.; Lunning, M.A.; Cavalli, F. Non-Hodgkin lymphoma. Lancet 2017, 390, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nowakowski, G.S.; Wang, M.L.; Ansell, S.M. Advances in CD30- and PD-1-targeted therapies for classical Hodgkin lymphoma. J. Hematol. Oncol. 2018, 11, 57. [Google Scholar] [CrossRef]
- de Leval, L.; Jaffe, E.S. Lymphoma Classification. Cancer J. 2020, 26, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Luchtel, R.A.; Dasari, S.; Oishi, N.; Pedersen, M.B.; Hu, G.; Rech, K.L.; Ketterling, R.P.; Sidhu, J.; Wang, X.; Katoh, R.; et al. Molecular profiling reveals immunogenic cues in anaplastic large cell lymphomas with DUSP22 rearrangements. Blood 2018, 132, 1386–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wu, Z.; Savin, A.; Yang, M.; Hsu, Y.R.; Jantuan, E.; Bacani, J.T.C.; Ingham, R.J. The c-Jun and JunB transcription factors facilitate the transit of classical Hodgkin lymphoma tumour cells through G1. Sci. Rep. 2018, 8, 16019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekh, P.; Kamble, S.; Zhao, N.; Zeng, Z.; Portier, B.P.; Zu, Y. Immunotherapy of CD30-expressing lymphoma using a highly stable ssDNA aptamer. Biomaterials 2013, 34, 8909–8917. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Yang, Y.; Kong, F.; Zhang, L.; Wei, K. CD30 aptamer-functionalized PEG-PLGA nanoparticles for the superior delivery of doxorubicin to anaplastic large cell lymphoma cells. Int. J. Pharm. 2019, 564, 340–349. [Google Scholar] [CrossRef]
- Chen, R.; Sun, P.; Chu, X.; Pu, X.; Yang, Y.; Zhang, N.; Zhao, Y. Synergistic Treatment of Tumor by Targeted Biotherapy and Chemotherapy via Site-Specific Anchoring of Aptamers on DNA Nanotubes. Int. J. Nanomed. 2020, 15, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Bagaria, H.G.; Wong, M.S.; Zu, Y. A nanocomplex that is both tumor cell-selective and cancer gene-specific for anaplastic large cell lymphoma. J. Nanobiotechnol. 2011, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Zeng, Z.; Zu, Y. Self-Assembled Aptamer-Nanomedicine for Targeted Chemotherapy and Gene Therapy. Small 2018, 14, 1702103. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Tung, C.H.; Zu, Y. Aptamer-Equipped Protamine Nanomedicine for Precision Lymphoma Therapy. Cancers 2020, 12, 780. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, X.; An, Y.; Duan, J.; Yang, X.D. Selection of a novel CD19 aptamer for targeted delivery of doxorubicin to lymphoma cells. Oncotarget 2018, 9, 26605–26615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallikaratchy, P.; Tang, Z.; Kwame, S.; Meng, L.; Shangguan, D.; Tan, W. Aptamer directly evolved from live cells recognizes membrane bound immunoglobin heavy mu chain in Burkitt’s lymphoma cells. Mol. Cell Proteom. 2007, 6, 2230–2238. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.D.; Srivastava, B.; Allman, D. Regulation of peripheral B cell maturation. Cell Immunol. 2006, 239, 92–102. [Google Scholar] [CrossRef]
- Cambier, J.C.; Campbell, K.S. Membrane immunoglobulin and its accomplices: New lessons from an old receptor. FASEB J. 1992, 6, 3207–3217. [Google Scholar] [CrossRef]
- Mallikaratchy, P.R.; Ruggiero, A.; Gardner, J.R.; Kuryavyi, V.; Maguire, W.F.; Heaney, M.L.; McDevitt, M.R.; Patel, D.J.; Scheinberg, D.A. A multivalent DNA aptamer specific for the B-cell receptor on human lymphoma and leukemia. Nucleic Acids Res. 2011, 39, 2458–2469. [Google Scholar] [CrossRef]
- Zhou, J.; Tiemann, K.; Chomchan, P.; Alluin, J.; Swiderski, P.; Burnett, J.; Zhang, X.; Forman, S.; Chen, R.; Rossi, J. Dual functional BAFF receptor aptamers inhibit ligand-induced proliferation and deliver siRNAs to NHL cells. Nucleic Acids Res. 2013, 41, 4266–4283. [Google Scholar] [CrossRef]
- Zhou, J.; Rossi, J.J.; Shum, K.T. Methods for assembling B-cell lymphoma specific and internalizing aptamer-siRNA nanoparticles via the sticky bridge. Methods Mol. Biol. 2015, 1297, 169–185. [Google Scholar] [CrossRef]
- Opazo, F.; Eiden, L.; Hansen, L.; Rohrbach, F.; Wengel, J.; Kjems, J.; Mayer, G. Modular Assembly of Cell-targeting Devices Based on an Uncommon G-quadruplex Aptamer. Mol. Ther. Nucleic Acids 2015, 4, e251. [Google Scholar] [CrossRef]
- Tonapi, S.S.; Pannu, V.; Duncan, J.E.; Rosenow, M.; Helmstetter, A.; Magee, D.; Zhang, Q.; Tinder, T.T.; Richards, M.; Halbert, D.D.; et al. Translocation of a Cell Surface Spliceosomal Complex Induces Alternative Splicing Events and Lymphoma Cell Necrosis. Cell Chem. Biol. 2019, 26, 756–764 e756. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.; Khanahmad, H.; Palizban, A. Selection and Characterization of Single-Stranded DNA Aptamers Binding Human B-Cell Surface Protein CD20 by Cell-SELEX. Molecules 2018, 23, 715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Li, J.; Wu, B.; Shang, Y.; Huang, X.; Dong, H.; Liu, H.; Chen, W.; Gui, R.; Nie, X. Smart Biomimetic Nanocomposites Mediate Mitochondrial Outcome through Aerobic Glycolysis Reprogramming: A Promising Treatment for Lymphoma. ACS Appl. Mater. Interfaces 2020, 12, 22687–22701. [Google Scholar] [CrossRef]
- Zümrüt, H.E.; Batool, S.; Van, N.; George, S.; Bhandari, S.; Mallikaratchy, P. Structural optimization of an aptamer generated from Ligand-Guided Selection (LIGS) resulted in high affinity variant toward mIgM expressed on Burkitt’s lymphoma cell lines. Biochim. Biophys. Acta. Gen. Subj. 2017, 1861, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Batool, S.; Argyropoulos, K.V.; Azad, R.; Okeoma, P.; Zumrut, H.; Bhandari, S.; Dekhang, R.; Mallikaratchy, P.R. Dimerization of an aptamer generated from Ligand-guided selection (LIGS) yields a high affinity scaffold against B-cells. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wen, J.; Li, H.; Xu, L.; Liu, Y.; Zhao, N.; Zeng, Z.; Qi, J.; Jiang, W.; Han, W.; et al. Aptamer-Engineered Natural Killer Cells for Cell-Specific Adaptive Immunotherapy. Small 2019, 15, e1900903. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.; Kamble, S.; Zhao, N.; Zeng, Z.; Wen, J.; Yuan, B.; Zu, Y. Biostable ssDNA aptamers specific for Hodgkin lymphoma. Sensors 2013, 13, 14543–14557. [Google Scholar] [CrossRef] [Green Version]
- Slyusarenko, M.; Shalaev, S.; Valitova, A.; Zabegina, L.; Nikiforova, N.; Nazarova, I.; Rudakovskaya, P.; Vorobiev, M.; Lezov, A.; Filatova, L.; et al. AuNP Aptasensor for Hodgkin Lymphoma Monitoring. Biosensors 2022, 12, 23. [Google Scholar] [CrossRef]
- Xi, D.; Li, Z.; Liu, L.; Ai, S.; Zhang, S. Ultrasensitive Detection of Cancer Cells Combining Enzymatic Signal Amplification with an Aerolysin Nanopore. Anal. Chem. 2018, 90, 1029–1034. [Google Scholar] [CrossRef]
- Fazlali, F.; Hashemi, P.; Khoshfetrat, S.M.; Halabian, R.; Baradaran, B.; Johari-Ahar, M.; Karami, P.; Hajian, A.; Bagheri, H. Electrochemiluminescent biosensor for ultrasensitive detection of lymphoma at the early stage using CD20 markers as B cell-specific antigens. Bioelectrochemistry 2021, 138, 107730. [Google Scholar] [CrossRef]
- Yi, Z.; Li, X.Y.; Gao, Q.; Tang, L.J.; Chu, X. Aptamer-aided target capturing with biocatalytic metal deposition: An electrochemical platform for sensitive detection of cancer cells. Analyst 2013, 138, 2032–2037. [Google Scholar] [CrossRef] [PubMed]
- Becker, N. Epidemiology of multiple myeloma. Recent Results Cancer Res. 2011, 183, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Caracciolo, D.; Montesano, M.; Altomare, E.; Scionti, F.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. The potential role of miRNAs in multiple myeloma therapy. Expert. Rev. Hematol. 2018, 11, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Morelli, E.; Biamonte, L.; Federico, C.; Amodio, N.; Di Martino, M.T.; Gallo Cantafio, M.E.; Manzoni, M.; Scionti, F.; Samur, M.K.; Gulla, A.; et al. Therapeutic vulnerability of multiple myeloma to MIR17PTi, a first-in-class inhibitor of pri-miR-17-92. Blood 2018, 132, 1050–1063. [Google Scholar] [CrossRef]
- Pharmacokinetics and Pharmacodynamics of a 13-mer LNA-inhibitor-miR-221 in Mice and Non-human Primates. Mol. Ther. Nucleic Acids 2016, 5, e336. [CrossRef]
- Sonneveld, P.; Broijl, A. Treatment of relapsed and refractory multiple myeloma. Haematologica 2016, 101, 396–406. [Google Scholar] [CrossRef]
- Watanabe, R.; Tokuhira, M.; Kizaki, M. Current approaches for the treatment of multiple myeloma. Int. J. Hematol. 2013, 97, 333–344. [Google Scholar] [CrossRef]
- Claudio, J.O.; Masih-Khan, E.; Tang, H.; Goncalves, J.; Voralia, M.; Li, Z.H.; Nadeem, V.; Cukerman, E.; Francisco-Pabalan, O.; Liew, C.C.; et al. A molecular compendium of genes expressed in multiple myeloma. Blood 2002, 100, 2175–2186. [Google Scholar] [CrossRef]
- Tai, Y.T.; Li, X.F.; Breitkreutz, I.; Song, W.; Neri, P.; Catley, L.; Podar, K.; Hideshima, T.; Chauhan, D.; Raje, N.; et al. Role of B-cell-activating factor in adhesion and growth of human multiple myeloma cells in the bone marrow microenvironment. Cancer Res. 2006, 66, 6675–6682. [Google Scholar] [CrossRef] [Green Version]
- Catuogno, S.; Di Martino, M.T.; Nuzzo, S.; Esposito, C.L.; Tassone, P.; de Franciscis, V. An Anti-BCMA RNA Aptamer for miRNA Intracellular Delivery. Mol. Ther. Nucleic Acids 2019, 18, 981–990. [Google Scholar] [CrossRef] [Green Version]
- El-Abd, N.; Fawzy, A.; Elbaz, T.; Hamdy, S. Evaluation of annexin A2 and as potential biomarkers for hepatocellular carcinoma. Tumour. Biol. 2016, 37, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Xiu, D.; Liu, L.; Qiao, F.; Yang, H.; Cui, L.; Liu, G. Annexin A2 Coordinates STAT3 to Regulate the Invasion and Migration of Colorectal Cancer Cells In Vitro. Gastroenterol. Res. Pract. 2016, 2016, 3521453. [Google Scholar] [CrossRef] [PubMed]
- Shetty, P.; Bargale, A.; Patil, B.R.; Mohan, R.; Dinesh, U.S.; Vishwanatha, J.K.; Gai, P.B.; Patil, V.S.; Amsavardani, T.S. Cell surface interaction of annexin A2 and galectin-3 modulates epidermal growth factor receptor signaling in Her-2 negative breast cancer cells. Mol. Cell Biochem. 2016, 411, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Seckinger, A.; Meissner, T.; Moreaux, J.; Depeweg, D.; Hillengass, J.; Hose, K.; Reme, T.; Rosen-Wolff, A.; Jauch, A.; Schnettler, R.; et al. Clinical and prognostic role of annexin A2 in multiple myeloma. Blood 2012, 120, 1087–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glavey, S.V.; Naba, A.; Manier, S.; Clauser, K.; Tahri, S.; Park, J.; Reagan, M.R.; Moschetta, M.; Mishima, Y.; Gambella, M.; et al. Proteomic characterization of human multiple myeloma bone marrow extracellular matrix. Leukemia 2017, 31, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zhang, Y.; Zeng, Y.; Peng, M.; Li, H.; Sun, S.; Ma, B.; Wang, Y.; Ye, M.; Liu, J. Screening and characterization of an Annexin A2 binding aptamer that inhibits the proliferation of myeloma cells. Biochimie 2018, 151, 150–158. [Google Scholar] [CrossRef]
- Ueki, R.; Sando, S. A DNA aptamer to c-Met inhibits cancer cell migration. Chem. Commun. 2014, 50, 13131–13134. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, H.; Zhou, W.; Sun, S.; Zeng, Y.; Zhang, H.; Liang, L.; Xiao, X.; Song, J.; Ye, M.; et al. Targeting c-met receptor tyrosine kinase by the DNA aptamer SL1 as a potential novel therapeutic option for myeloma. J. Cell Mol. Med. 2018, 22, 5978–5990. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Tao, W.; Hao, S.; Iyer, S.P.; Zu, Y. A unique aptamer-drug conjugate for targeted therapy of multiple myeloma. Leukemia 2016, 30, 987–991. [Google Scholar] [CrossRef]
- Bagalkot, V.; Farokhzad, O.C.; Langer, R.; Jon, S. An aptamer-doxorubicin physical conjugate as a novel targeted drug-delivery platform. Angew. Chem. Int. Ed. Engl. 2006, 45, 8149–8152. [Google Scholar] [CrossRef]
- Dai, H.; Ye, M.; Peng, M.; Zhou, W.; Bai, H.; Xiao, X.; Ma, B.; Zhou, J.; Tang, S.; Yao, S.; et al. Aptamer TY04 inhibits the growth of multiple myeloma cells via cell cycle arrest. Tumour. Biol. 2014, 35, 7561–7568. [Google Scholar] [CrossRef] [PubMed]
- Waldschmidt, J.M.; Simon, A.; Wider, D.; Muller, S.J.; Follo, M.; Ihorst, G.; Decker, S.; Lorenz, J.; Chatterjee, M.; Azab, A.K.; et al. CXCL12 and CXCR7 are relevant targets to reverse cell adhesion-mediated drug resistance in multiple myeloma. Br. J. Haematol. 2017, 179, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Gao, X.; Zhang, Z. Isolation and characterization of 2’-amino-modified RNA aptamers for human TNFalpha. Genom. Proteom. Bioinform. 2004, 2, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eulberg, D.; Klussmann, S. Spiegelmers: Biostable aptamers. Chembiochem 2003, 4, 979–983. [Google Scholar] [CrossRef]
- Sameiyan, E.; Bagheri, E.; Dehghani, S.; Ramezani, M.; Alibolandi, M.; Abnous, K.; Taghdisi, S.M. Aptamer-based ATP-responsive delivery systems for cancer diagnosis and treatment. Acta. Biomater. 2021, 123, 110–122. [Google Scholar] [CrossRef]
- Bukari, B.; Samarasinghe, R.M.; Noibanchong, J.; Shigdar, S.L. Non-Invasive Delivery of Therapeutics into the Brain: The Potential of Aptamers for Targeted Delivery. Biomedicines 2020, 8, 120. [Google Scholar] [CrossRef]
- Cesarini, V.; Scopa, C.; Silvestris, D.A.; Scafidi, A.; Petrera, V.; Del Baldo, G.; Gallo, A. Aptamer-Based In Vivo Therapeutic Targeting of Glioblastoma. Molecules 2020, 25, 4267. [Google Scholar] [CrossRef]
- Pasut, G.; Veronese, F.M. State of the art in PEGylation: The great versatility achieved after forty years of research. J. Control Release 2012, 161, 461–472. [Google Scholar] [CrossRef]
- Rusconi, C.P.; Roberts, J.D.; Pitoc, G.A.; Nimjee, S.M.; White, R.R.; Quick, G., Jr.; Scardino, E.; Fay, W.P.; Sullenger, B.A. Antidote-mediated control of an anticoagulant aptamer in vivo. Nat. Biotechnol. 2004, 22, 1423–1428. [Google Scholar] [CrossRef]
- Mercier, M.C.; Dontenwill, M.; Choulier, L. Selection of Nucleic Acid Aptamers Targeting Tumor Cell-Surface Protein Biomarkers. Cancers 2017, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Marshall, M.L.; Wagstaff, K.M. Internalized Functional DNA Aptamers as Alternative Cancer Therapies. Front. Pharmacol. 2020, 11, 1115. [Google Scholar] [CrossRef] [PubMed]
- Drabik, A.; Ner-Kluza, J.; Mielczarek, P.; Civit, L.; Mayer, G.; Silberring, J. Advances in the Study of Aptamer-Protein Target Identification Using the Chromatographic Approach. J. Proteome Res. 2018, 17, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Meyer, S.; Propson, N.E.; Nie, J.; Jiang, P.; Stewart, R.; Thomson, J.A. Characterization and target identification of a DNA aptamer that labels pluripotent stem cells. Cell Res. 2015, 25, 390–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryou, S.M.; Kim, J.M.; Yeom, J.H.; Hyun, S.; Kim, S.; Han, M.S.; Kim, S.W.; Bae, J.; Rhee, S.; Lee, K. Gold nanoparticle-assisted delivery of small, highly structured RNA into the nuclei of human cells. Biochem. Biophys. Res. Commun. 2011, 416, 178–183. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Sun, H.Y.; Lee, K.H.; Oh, B.H.; Cha, Y.J.; Kim, B.H.; Yoo, J.Y. 5′-Triphosphate-RNA-independent activation of RIG-I via RNA aptamer with enhanced antiviral activity. Nucleic Acids Res. 2012, 40, 2724–2733. [Google Scholar] [CrossRef]
- Choi, K.H.; Park, M.W.; Lee, S.Y.; Jeon, M.Y.; Kim, M.Y.; Lee, H.K.; Yu, J.; Kim, H.J.; Han, K.; Lee, H.; et al. Intracellular expression of the T-cell factor-1 RNA aptamer as an intramer. Mol. Cancer Ther. 2006, 5, 2428–2434. [Google Scholar] [CrossRef]


| Aptamers | Antibodies | |
|---|---|---|
| Composition | DNA or RNA (A, G, T, U, C) | Protein (Amino acid) |
| Size | Small (1–2 nm; <30 kDa) | Large (~15 nm; ~150 kDa) |
| Immunogenicity | Low | High |
| Binding affinity | High (nanomolar to picomolar range) | High (nanomolar to picomolar range) |
| Targets | Widely, from ions and small molecules to whole cells and live animals | Proteins mainly |
| Target Size | ≥600 Da | ≥60 Da |
| Synthesis time | Short (2–8 weeks) | Long (more than 6 months) |
| Reproducibility | None or little variation between batches | Wide variation between batches |
| Manufacturing Process | In vivo | In vitro |
| In vivo half-life | Short | Long |
| Cost | Cheap | Expensive |
| Modification | Easy | Hard |
| Stability | High (stable at a wide range of temperatures; reversible denaturation) | Low (susceptible to high temperatures and pH changes. irreversible denaturation) |
| Tissue penetration/kidney filtration | Fast | Slow |
| Toxicity | Low | High |
| Disease | Name | DNA/ RNA | Type of SELEX | Target Cell | Target Molecule | Tested | Kd | Conjugated Molecule | Application/Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| AML | ZW25; CY30 | DNA (66-mer) | Protein-SELEX | CD123+ cells | CD123 | In vitro; In vivo | 29.41 nM; 15.38 nM | Dox (By ZW25 mediated) | Inhibition of AML cells growth in vitro and prolongation of survival in mice in vivo | [43] |
| SS30 (thioaptamer from CY30) | DNA (66 bp) | CD123+ cells | In vitro; In vivo | 39.1 nM for CD123 peptide and 287.6 nM for CD123 AML cells | Selective inhibition of proliferation of AML cells in vitro and prolongation of survival in mice in vivo via JAK2/STAT5 signaling pathway | [44] | ||||
| sgRNA-targeting sequence | [45] | |||||||||
| #1 (#1-F) | DNA (77-mer) | Hybrid-SELEX | HEL cells | CD117 | In vitro | 4.24 nM | MTX | specifically inhibits AML cell growth and induces cell cycle arrest in the G1 phase | [52] | |
| K19 | DNA (50-mer) | Cell-SELEX | NB4 | Siglec-5 | In vitro | 12.37 nM | Detection of low concentrations of AML cells | [118] | ||
| two “lock” sequences and two G-rich sequences | Building a luminescence sensing platform to detect Siglec-5 | [119] | ||||||||
| AB3 | DNA (59nt) | Protein-SELEX | HL-60, Jurkat, Ramos | OFA/iLRP | In vitro | 101 nM | Dox | Selective delivery of Dox to OFA/iLRP-positive AML cells | [54] | |
| S30-T1 | DNA | Paired cell-based SELEX | CD33 transfected-HEK293T, HL-60 | CD33 | In vitro; In vivo | ~43 nM | Dox | Highly identifiable AML cells in vitro and in vivo. Specifically recognize and inhibit HL-60 cell proliferation by arresting the cell cycle at the G2 phase in vitro. | [56] | |
| AS1411 | DNA (26-mer) | NB4, Kas-1,HL60 | Nucleolin | In vitro; In vivo | AuNP, anti-221 | AS1411 and anti-221 synergistically suppress AML cell growth via targeting key molecules involved in NCL/miR-221/NFκB/DNMT1 pathway in vitro and in vivo | [61] | |||
| KGE02 | DNA (76-mer) | whole-cell SELEX | MLL-AF9 RAS (MA9Ras) AML cells | MLL-AF9 | In vitro | 37.5 ± 2.5 nM | A DNA aptamer specific to AML cells was developed and characterized for future drug-aptamer conjugates. | [58] | ||
| ALL | Sgc8 (Sgc8c) | DNA | Cell-SELEX | CCRF-CEM | PTK7 | In vitro, Ex vivo | 0.8 nM | Tb3+ | Cellular detection and early diagnosis of ALL | [71] |
| In vitro, Ex vivo | DEAS/PMMA-co-MAA | Precision targeting and imaging in ALL cells | [81] | |||||||
| In vitro | Ag 10 NPs | Cellular detection and imaging | [73] | |||||||
| In vitro, In vivo | 18F | Cellular detection and imaging | [75] | |||||||
| In vitro | silica | Cellular detection | [76] | |||||||
| In vitro | Cu-Au NPs | Cellular detection | [77] | |||||||
| In vitro | GMNPs | detection of leukemia cells | [78] | |||||||
| In vitro | MBs | Selective collection and detection of ALL cells | [82] | |||||||
| In vitro | APBA-AuNPs | Cellular detection | [83] | |||||||
| In vitro | BP NS, Dox, PEG | Growth inhibition, targeted and synergetic chemophotothermal therapy of ALL | [89] | |||||||
| In vitro | Dox | Growth inhibition of ALL cells | [70] | |||||||
| In vitro | Tetrahedron DNA, Dox | More cytotoxic to ALL cells | [86] | |||||||
| In vitro | Combretastatin A4 | More cytotoxic to ALL cells | [75,97] | |||||||
| In vitro; In vivo | PCL-ss-Ara, BSA | Inhibition of ALL cells growth in vitro and prolongation of survival in mice in vivo | [94] | |||||||
| In vitro | Au NP, Dox | Growth inhibition of ALL cells | [88] | |||||||
| In vitro | DNA dendrimer, Dox | Growth inhibition of ALL cells | [87] | |||||||
| In vitro | PIC | Inhibition of AML cells proliferation by G0/G1 phase arrest | [96] | |||||||
| In vitro | LIPO, VCR | More cytotoxic to ALL cells | [95] | |||||||
| Molt-4 | In vitro | Dau | Growth inhibition of ALL cells | [90] | ||||||
| In vitro | AuNPs, Dau, AS1411 | Growth inhibition of ALL cells | [91] | |||||||
| In vitro | AuNPs, Dau, | More cytotoxic to ALL cells | [92] | |||||||
| In vitro | SWNT, Dau | More cytotoxic to ALL cells | [93] | |||||||
| In vitro | ATP aptamer, DAFGO | Detection and early diagnosis by complexes being efficiently internalized into ALL cells and inducing intense fluorescence emission | [72] | |||||||
| In vitro | PPIX-[BMIm], Au NPs | Cellular detection | [84] | |||||||
| CML | K562 | In vitro | siRNA | The aptamer-siRNA compound can significantly induce K562 cell apoptosis | [100] | |||||
| NOX-A12 | RNA (45-mer) | BCR-ABL-positive leukemia cells | CXCL12 (SDF-1) | In vitro; In vivo | Combined use of targeted kinase inhibition and NOX-A12 for treatment. Cell migration | [101] | ||||
| CLL | CLL cells from peripheral blood samples | In vitro | NOX-A12 enhances the cytotoxicity of anticancer drugs by mobilizing leukemia cells to peripheral blood through inhibition of SDF-1. CLL migration and drug resistance | [117] | ||||||
| GT75 | MEC-1 | eEF1A1 | In vitro; In vivo | siRNA (siA1) | MEC-1 viability reduction/autophagy stimulation and in vivo tumor growth down-regulation. | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Xiong, S.; Ur Rehman, Z.; He, X.; Peng, H.; Liu, J.; Sun, S. The Research Advances of Aptamers in Hematologic Malignancies. Cancers 2023, 15, 300. https://doi.org/10.3390/cancers15010300
Liao Y, Xiong S, Ur Rehman Z, He X, Peng H, Liu J, Sun S. The Research Advances of Aptamers in Hematologic Malignancies. Cancers. 2023; 15(1):300. https://doi.org/10.3390/cancers15010300
Chicago/Turabian StyleLiao, Yongkang, Shijun Xiong, Zaid Ur Rehman, Xiaoli He, Hongling Peng, Jing Liu, and Shuming Sun. 2023. "The Research Advances of Aptamers in Hematologic Malignancies" Cancers 15, no. 1: 300. https://doi.org/10.3390/cancers15010300
APA StyleLiao, Y., Xiong, S., Ur Rehman, Z., He, X., Peng, H., Liu, J., & Sun, S. (2023). The Research Advances of Aptamers in Hematologic Malignancies. Cancers, 15(1), 300. https://doi.org/10.3390/cancers15010300