


Knockdown of UBQLN1 Functions as a Strategy to Inhibit CRC Progression through the ERK-c-Myc Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Plasmid Constructions
2.3. Establishment of Stable Expression Cell Lines
2.4. Cell Counting Kit-8 (CCK-8) Assay
2.5. Colony Formation Assay
2.6. Cell Invasion and Motility Assay
2.7. Western Blot
2.8. Nude Mice Xenograft and Metastasis Experiments
2.9. Statistical Analysis
3. Results
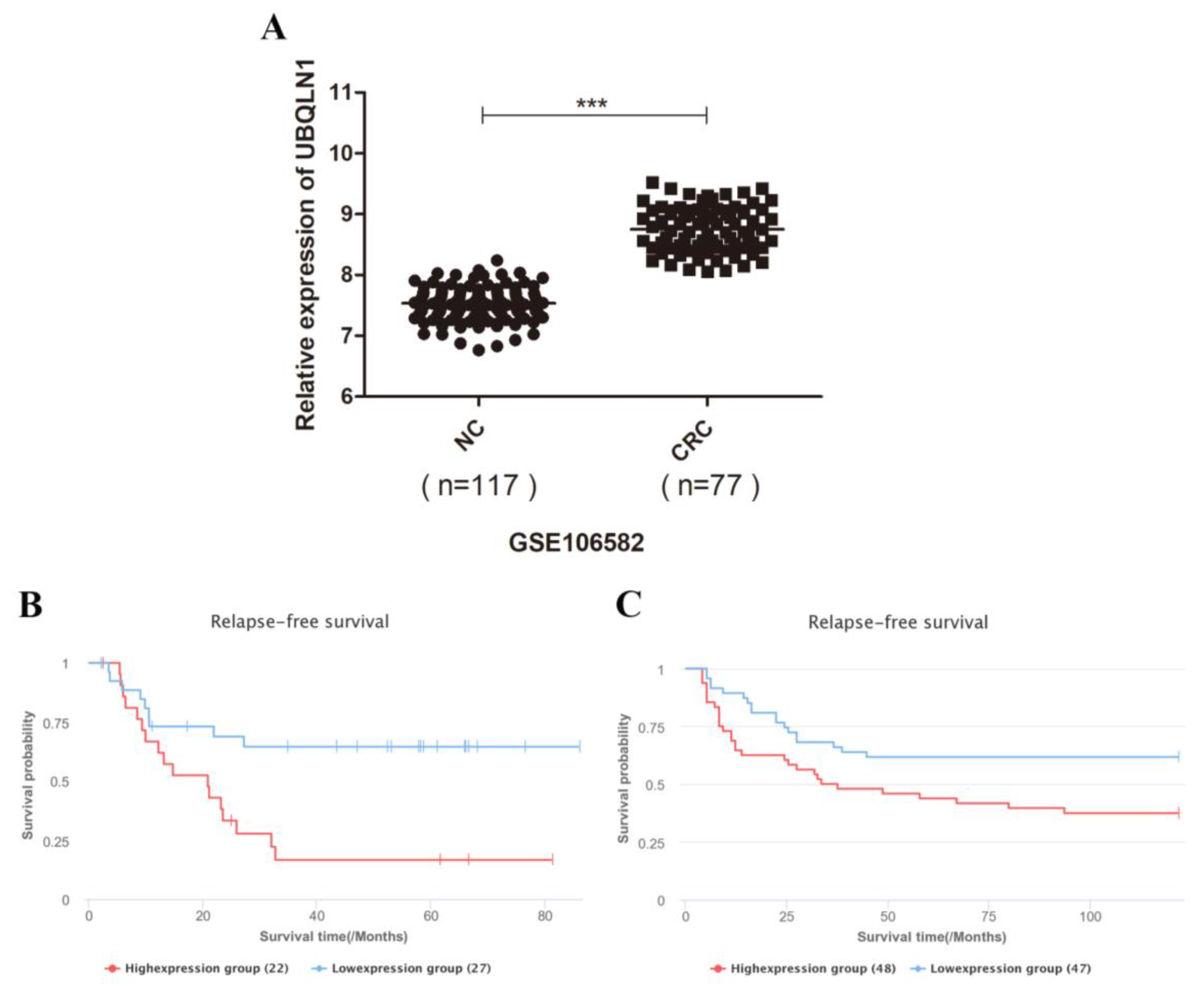
3.1. UBQLN1 Enhanced Expression in Colorectal Cancer Tissues and Is Correlated with Poor Prognosis
3.2. UBQLN1 Promoted CRC Cell Proliferation In Vitro
3.3. UBQLN1 Promoted CRC Cells’ EMT In Vitro
3.4. Reduced UBQLN1 Inhibited CRC Cells’ Tumorigenesis and Metastasis In Vivo
3.5. Knockdown of UBQLN1 Suppressed ERK-c-Myc Signaling Pathway in CRC
3.6. Knockdown of UBQLN1 Inhibited CRC Cells’ Malignant Progression through ERK-c-Myc Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biller, L.H.; Schrag, D. Diagnosis and Treatment of Metastatic Colorectal Cancer: A Review. JAMA 2021, 325, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wei, W.; Zeng, H.; Zheng, R.; Zhang, S.; An, L.; Chen, R.; Wang, S.; Sun, K.; Matsuda, T.; Bray, F.; et al. Cancer registration in China and its role in cancer prevention and control. Lancet Oncol. 2020, 21, e342–e349. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladabaum, U.; Dominitz, J.A.; Kahi, C.; Schoen, R.E. Strategies for Colorectal Cancer Screening. Gastroenterology 2020, 158, 418–432. [Google Scholar] [CrossRef]
- Marín, I. The ubiquilin gene family: Evolutionary patterns and functional insights. BMC Evol. Biol. 2014, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Kurlawala, Z.; Shah, P.P.; Shah, C.; Beverly, L.J. The STI and UBA Domains of UBQLN1 Are Critical Determinants of Substrate Interaction and Proteostasis. J. Cell. Biochem. 2017, 118, 2261–2270. [Google Scholar] [CrossRef]
- Chen, X.; Ebelle, D.L.; Wright, B.J.; Sridharan, V.; Hooper, E.; Walters, K.J. Structure of hRpn10 Bound to UBQLN2 UBL Illustrates Basis for Complementarity between Shuttle Factors and Substrates at the Proteasome. J. Mol. Biol. 2019, 431, 939–955. [Google Scholar] [CrossRef]
- Lipinszki, Z.; Kovács, L.; Deák, P.; Udvardy, A. Ubiquitylation of Drosophila p54/Rpn10/S5a regulates its interaction with the UBA-UBL polyubiquitin receptors. Biochemistry 2012, 51, 2461–2470. [Google Scholar] [CrossRef]
- Bertram, L.; Hiltunen, M.; Parkinson, M.; Ingelsson, M.; Lange, C.; Ramasamy, K.; Tanzi, R.E. Family-based association between Alzheimer’s disease and variants in UBQLN1. N. Engl. J. Med. 2005, 352, 884–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, N.J.; Lewis, J.; Clippinger, A.; Thomas, M.A.; Adamson, J.; Cruz, P.E.; Cannon, A.; Xu, G.; Golde, T.E.; Shaw, G.; et al. Unbiased screen reveals ubiquilin-1 and -2 highly associated with huntingtin inclusions. Brain Res. 2013, 1524, 62–73. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Cao, A.; Qin, T.; Zhang, Q.; Fan, S.; Wang, B.; Song, B.; Yu, X.; Li, L. Abnormally elevated ubiquilin-1 expression in breast cancer regulates metastasis and stemness via AKT signaling. Oncol. Rep. 2021, 46, 236. [Google Scholar] [CrossRef]
- Shah, P.P.; Lockwood, W.W.; Saurabh, K.; Kurlawala, Z.; Shannon, S.P.; Waigel, S.; Zacharias, W.; Beverly, L.J. Ubiquilin1 represses migration and epithelial-to-mesenchymal transition of human non-small cell lung cancer cells. Oncogene 2015, 34, 1709–1717. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Ji, L.; Ruan, Y.; Wan, Z.; Lin, Z.; Xia, S.; Tao, L.; Zheng, J.; Cai, L.; Wang, Y.; et al. UBQLN1 mediates sorafenib resistance through regulating mitochondrial biogenesis and ROS homeostasis by targeting PGC1β in hepatocellular carcinoma. Signal Transduct. Target. Ther. 2021, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Köhne, C.-H.; István Láng; Folprecht, G.; Nowacki, M.P.; Cascinu, S.; Shchepotin, I.; Maurel, J.; Cunningham, D.; Tejpar, S.; et al. Faculty Opinions recommendation of Cetuximab plus irinotecan, fluorouracil, and leucovorin as first-line treatment for metastatic colorectal cancer: Updated analysis of overall survival according to tumor KRAS and BRAF mutation status. J. Clin. Oncol. 2011, 29, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.M.; Der, C.J. KRAS: The Critical Driver and Therapeutic Target for Pancreatic Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a031435. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Hu, Q.; Ji, S.; Xu, J.; Dai, W.; Liu, W.; Xu, W.; Sun, Q.; Zhang, Z.; Ni, Q.; et al. Homeodomain-interacting protein kinase 2 suppresses proliferation and aerobic glycolysis via ERK/cMyc axis in pancreatic cancer. Cell Prolif. 2019, 52, e12603. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Pei, L.; Xia, H.; Tang, Q.; Bi, F. Role of oncogenic KRAS in the prognosis, diagnosis and treatment of colorectal cancer. Mol. Cancer 2021, 20, 143. [Google Scholar] [CrossRef]
- Hofmann, M.H.; Gerlach, D.; Misale, S.; Petronczki, M.; Kraut, N. Expanding the Reach of Precision Oncology by Drugging All KRAS Mutants. Cancer Discov. 2022, 12, 924–937. [Google Scholar] [CrossRef]
- Moore, A.R.; Rosenberg, S.C.; McCormick, F.; Malek, S. RAS-targeted therapies: Is the undruggable drugged? Nat. Rev. Drug Discov. 2020, 19, 533–552. [Google Scholar] [CrossRef] [PubMed]
- Drosten, M.; Barbacid, M. Targeting the MAPK Pathway in KRAS-Driven Tumors. Cancer Cell 2020, 37, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.J.; Stinchcombe, T.E. KRAS mutation: Should we test for it, and does it matter? J. Clin. Oncol. 2013, 31, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ding, C.; Chen, Y.; Hu, W.; Lu, Y.; Wu, W.; Zhang, Y.; Yang, B.; Wu, H.; Peng, C.; et al. ACSL4 promotes hepatocellular carcinoma progression via c-Myc stability mediated by ERK/FBW7/c-Myc axis. Oncogenesis 2020, 9, 42. [Google Scholar] [CrossRef]
- Satoh, J.; Tabunoki, H.; Ishida, T.; Saito, Y.; Arima, K. Ubiquilin-1 immunoreactivity is concentrated on Hirano bodies and dystrophic neurites in Alzheimer’s disease brains. Neuropathol. Appl. Neurobiol. 2013, 39, 817–830. [Google Scholar] [CrossRef]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Molina, J.R.; Adjei, A.A. The Ras/Raf/MAPK pathway. J. Thorac. Oncol. 2006, 1, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Roberts, P.J.; Der, C.J. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene 2007, 26, 3291–3310. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Yaeger, R.; Corcoran, R.B. Targeting Alterations in the RAF–MEK Pathway. Cancer Discov. 2019, 9, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Liu, Y.; Yang, S.; Wu, X.; Li, H.; Wang, Q. MEK inhibitors for the treatment of non-small cell lung cancer. J. Hematol. Oncol. 2021, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorley-Lawson, D.A.; Allday, M.J. The curious case of the tumour virus: 50 years of Burkitt’s lymphoma. Nat. Rev. Microbiol. 2008, 6, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. MYC on the path to cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yao, B.; Gui, T.; Guo, C.; Wu, X.; Li, J.; Ma, L.; Deng, Y.; Xu, P.; Wang, Y.; et al. PRMT5-dependent transcriptional repression of c-Myc target genes promotes gastric cancer progression. Theranostics 2020, 10, 4437–4452. [Google Scholar] [CrossRef]
- Figueiredo, J.C.; Knight, J.A.; Cho, S.; Savas, S.; Onay, U.V.; Briollais, L.; Goodwin, P.J.; McLaughlin, J.R.; Andrulis, I.L. Polymorphisms cMyc-N11S and p27-V109G and breast cancer risk and prognosis. BMC Cancer 2007, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Liang, X.; Liang, Z.; Zhao, L. Study on the effect and mechanism of NFKBIA on cervical cancer progress in vitro and in vivo. J. Obstet. Gynaecol. Res. 2021, 47, 3931–3942. [Google Scholar] [CrossRef]
- Martelli, A.M.; Evangelisti, C.; Paganelli, F.; Chiarini, F.; McCubrey, J.A. GSK-3: A multifaceted player in acute leukemias. Leukemia 2021, 35, 1829–1842. [Google Scholar] [CrossRef]
- Tsai, C.-C.; Su, Y.-C.; Bamodu, O.A.; Chen, B.-J.; Tsai, W.-C.; Cheng, W.-H.; Lee, C.-H.; Hsieh, S.-M.; Liu, M.-L.; Fang, C.-L.; et al. High-Grade B-Cell Lymphoma (HGBL) with MYC and BCL2 and/or BCL6 Rearrangements Is Predominantly BCL6-Rearranged and BCL6-Expressing in Taiwan. Cancers 2021, 13, 1620. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ni, R.; Jiang, J.; Zhao, M.; Huang, S.; Huang, C. Knockdown of UBQLN1 Functions as a Strategy to Inhibit CRC Progression through the ERK-c-Myc Pathway. Cancers 2023, 15, 3088. https://doi.org/10.3390/cancers15123088
Ni R, Jiang J, Zhao M, Huang S, Huang C. Knockdown of UBQLN1 Functions as a Strategy to Inhibit CRC Progression through the ERK-c-Myc Pathway. Cancers. 2023; 15(12):3088. https://doi.org/10.3390/cancers15123088
Chicago/Turabian StyleNi, Ruoxuan, Jianwei Jiang, Mei Zhao, Shengkai Huang, and Changzhi Huang. 2023. "Knockdown of UBQLN1 Functions as a Strategy to Inhibit CRC Progression through the ERK-c-Myc Pathway" Cancers 15, no. 12: 3088. https://doi.org/10.3390/cancers15123088
APA StyleNi, R., Jiang, J., Zhao, M., Huang, S., & Huang, C. (2023). Knockdown of UBQLN1 Functions as a Strategy to Inhibit CRC Progression through the ERK-c-Myc Pathway. Cancers, 15(12), 3088. https://doi.org/10.3390/cancers15123088