
Differential Gene Expression of Checkpoint Markers and Cancer Markers in Mouse Models of Spontaneous Chronic Colitis

 , ,
, ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Tissues
2.2. RNA Extraction
2.3. Library Preparation and RNA Sequencing
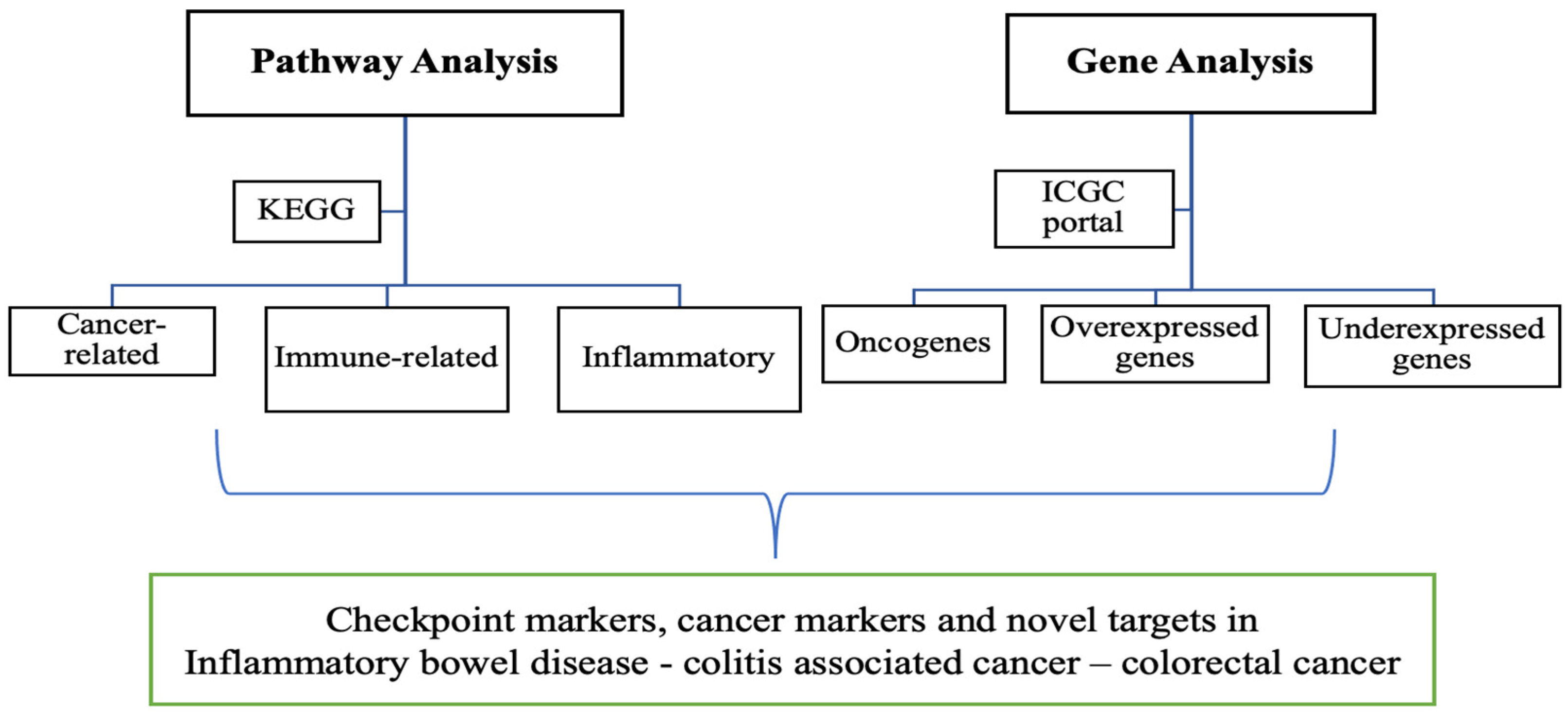
2.4. Data Analysis
3. Results and Discussion
3.1. Pathway Analysis
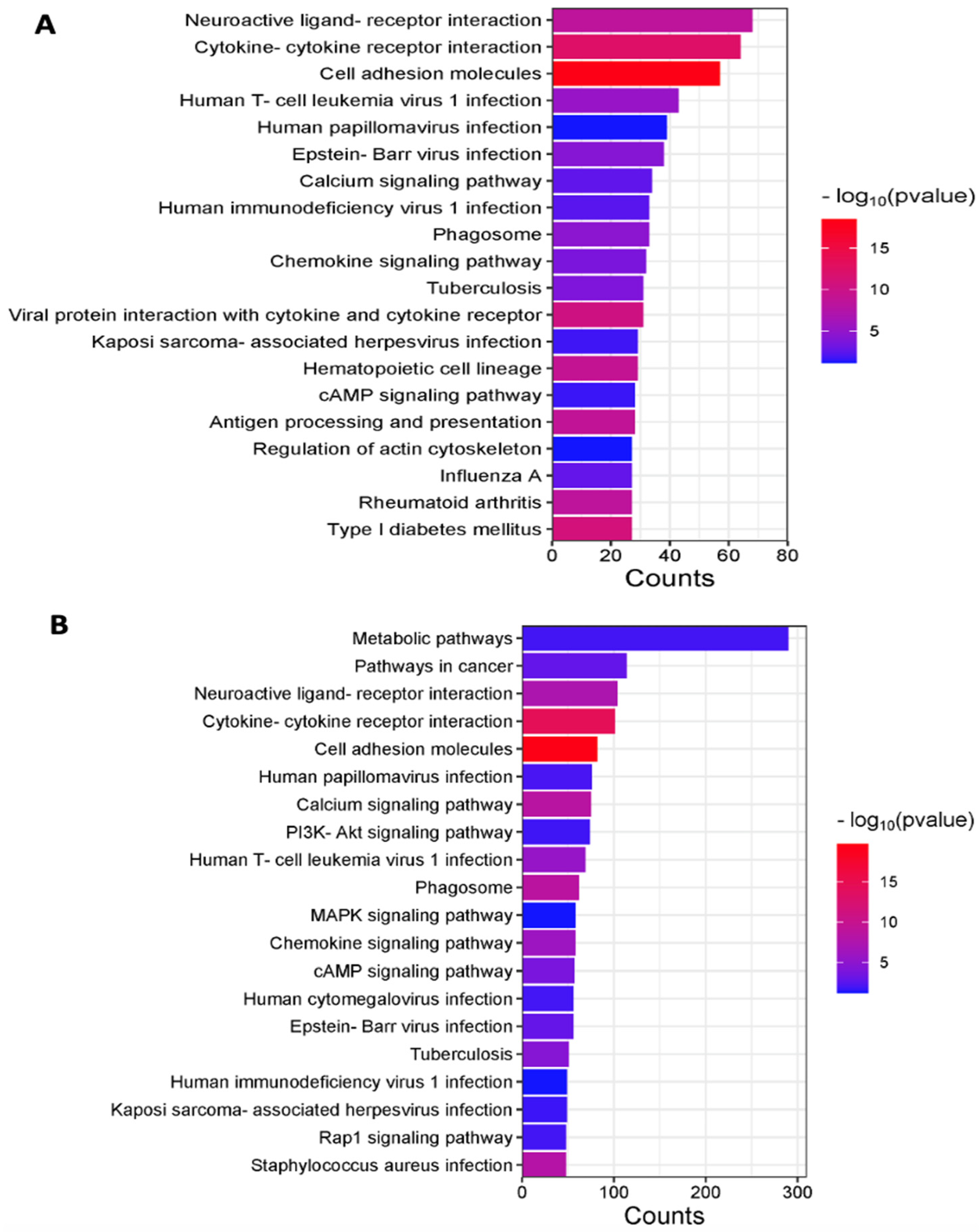
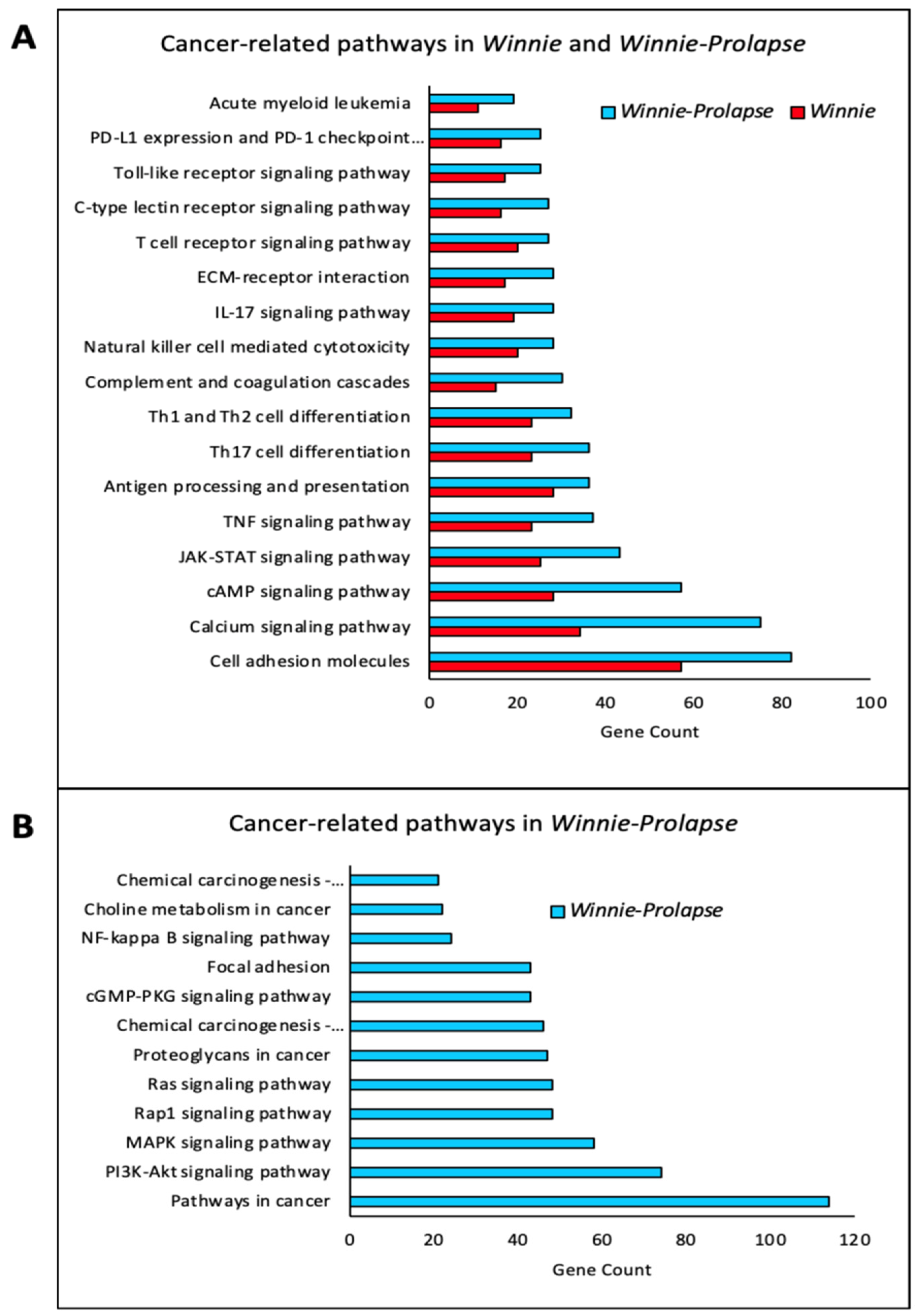
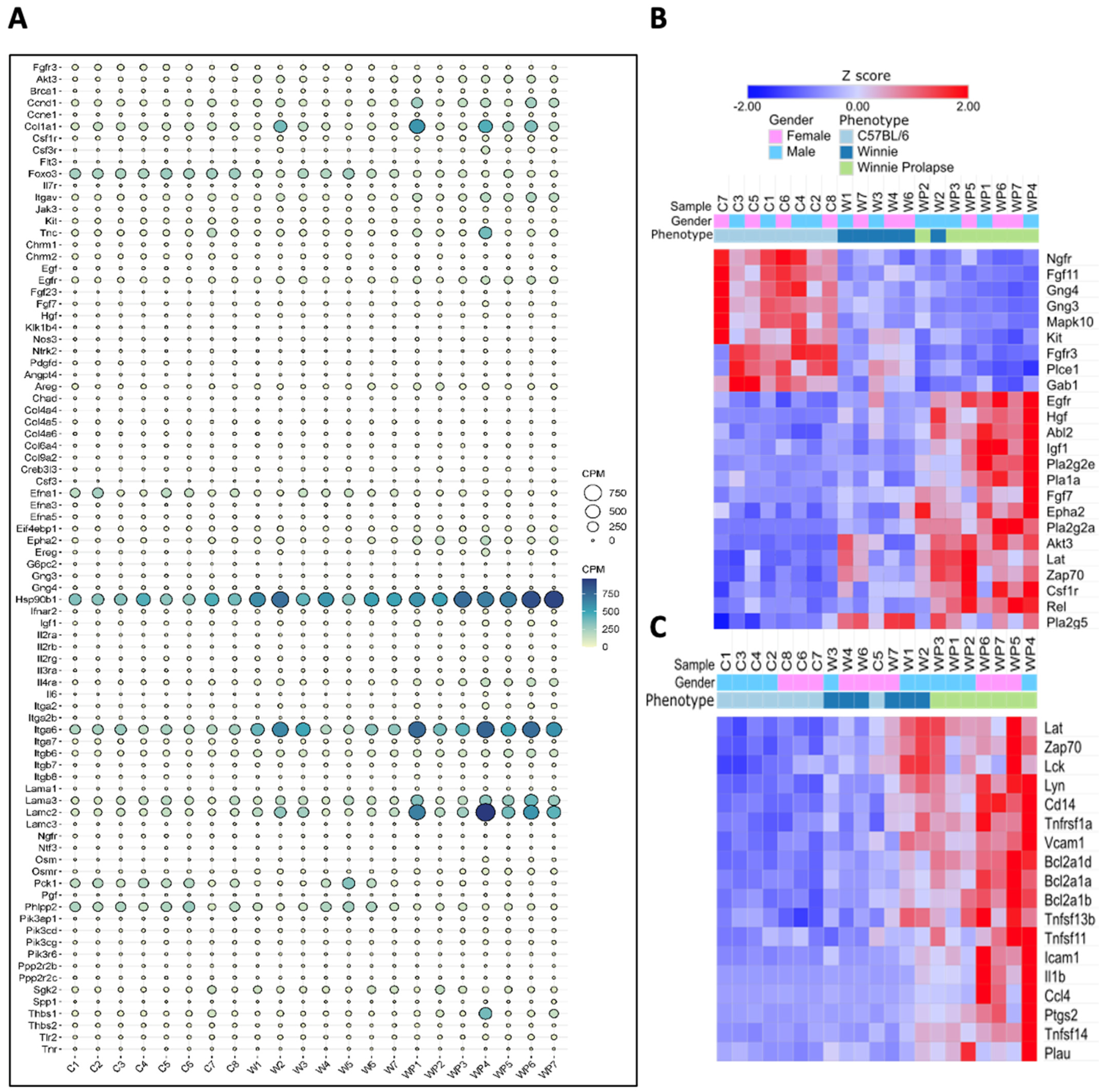
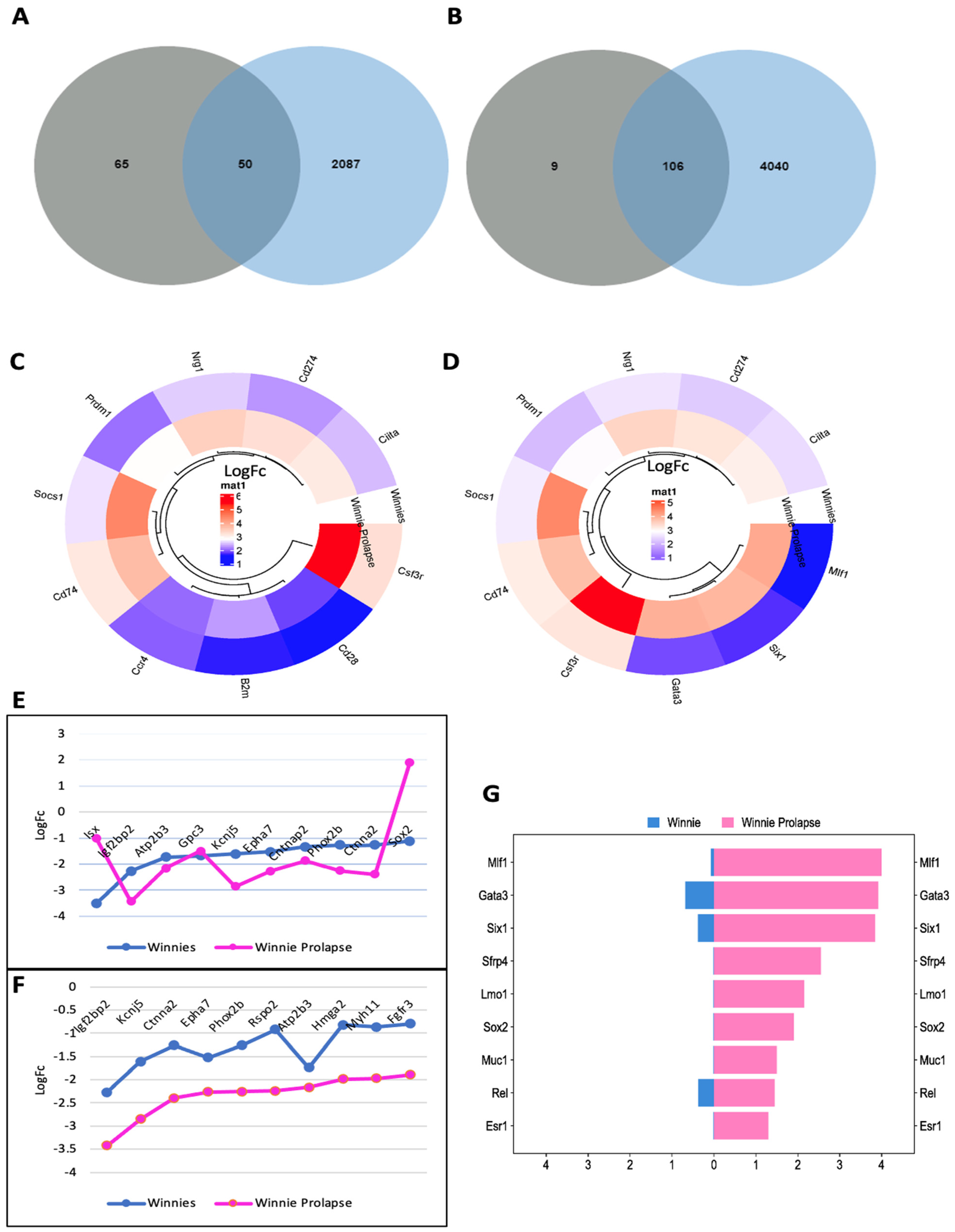
3.1.1. Inflammatory and Oncogenic Pathways Are Upregulated in Winnie (Colitis—Inflammation) Mice with Further Upregulation in Winnie-Prolapse (Severe Colitis—Severe Inflammation) Mice
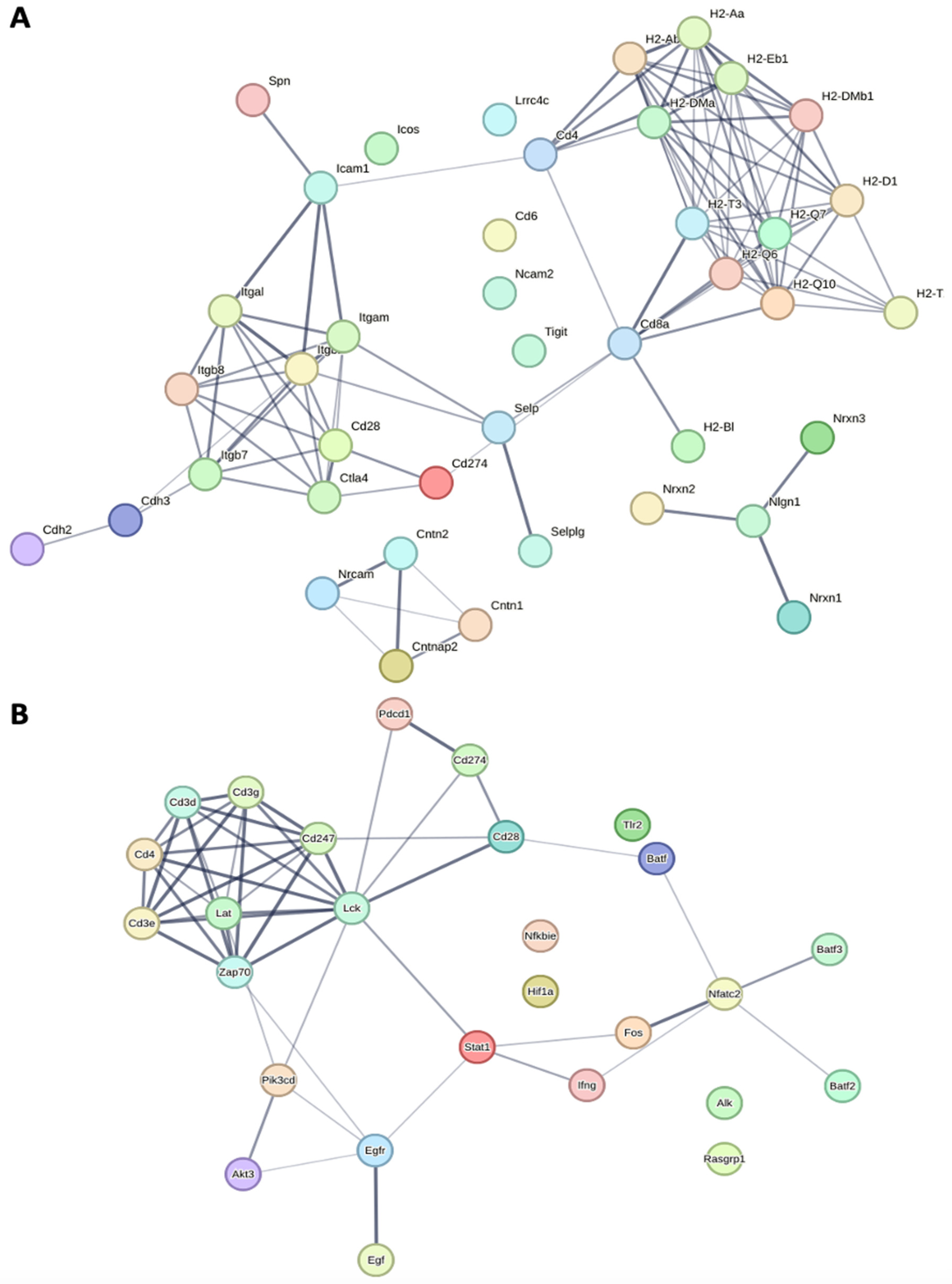
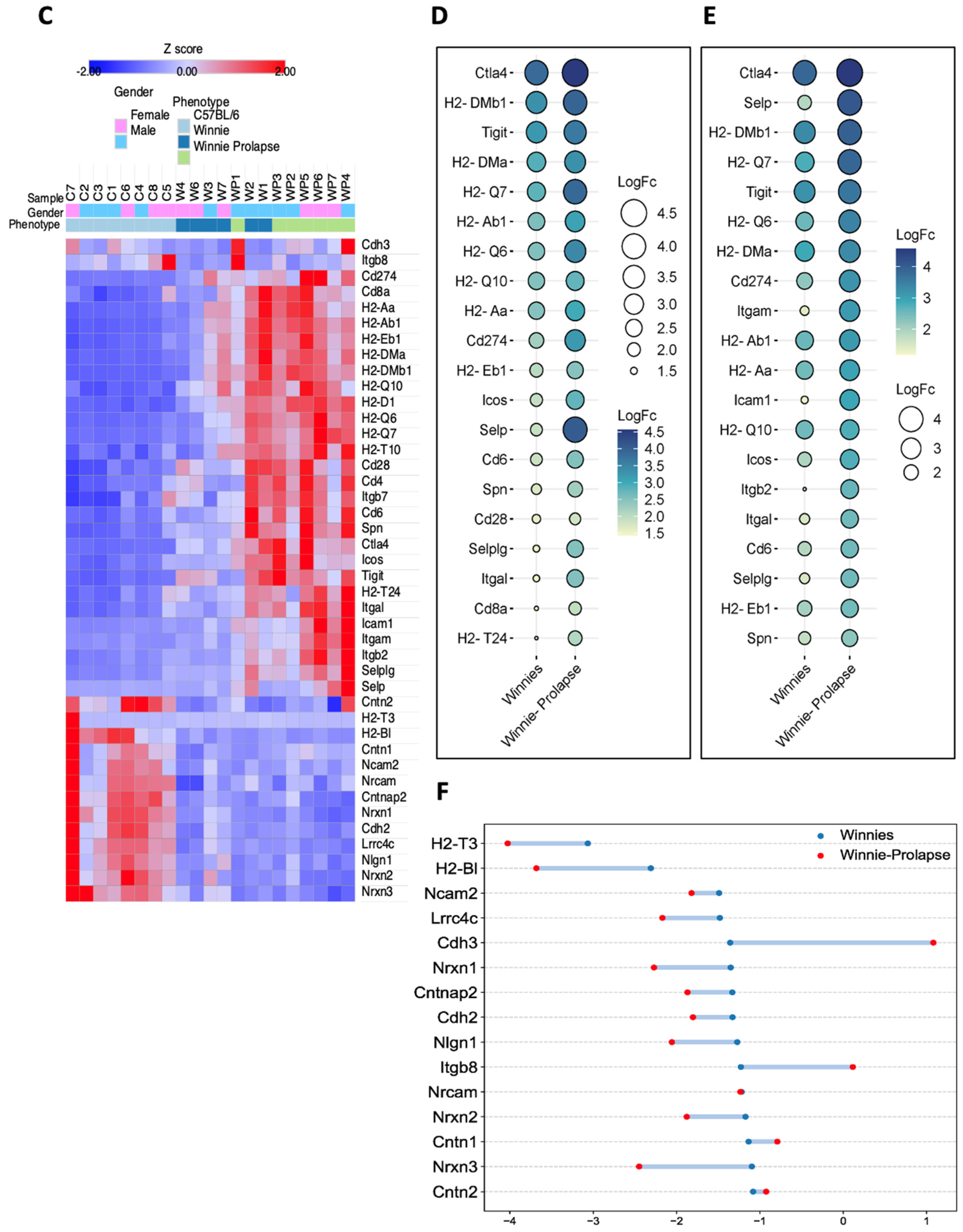
3.1.2. Upregulation of Key Cellular Adhesion Molecules Which Act as Checkpoint Markers in Winnie Mice
3.1.3. Transcriptomes of Winnie-Prolapse Mice Are Enriched in Cancer-Related Pathways That Are Absent in Winnie Mice
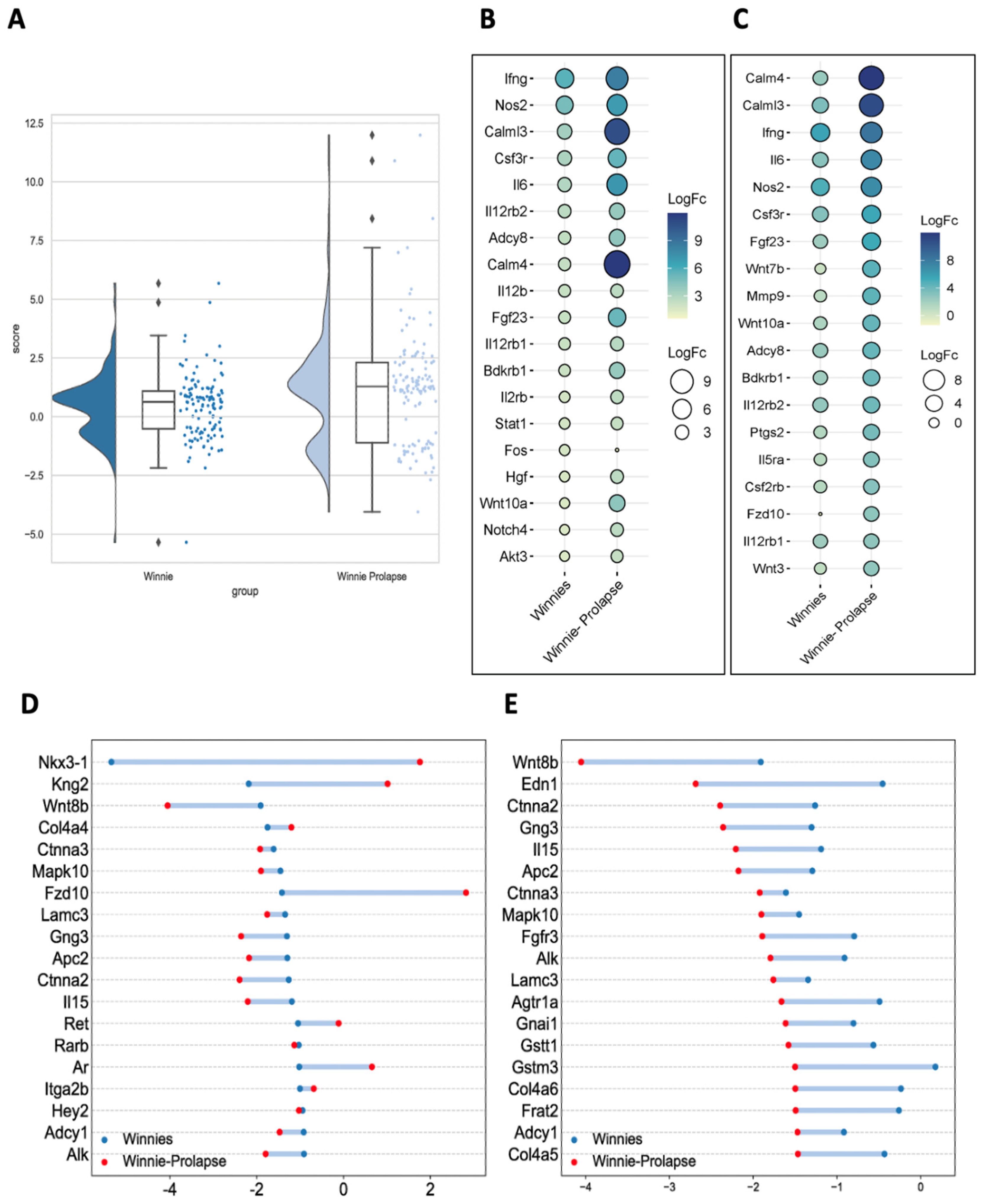
3.2. Gene Analysis
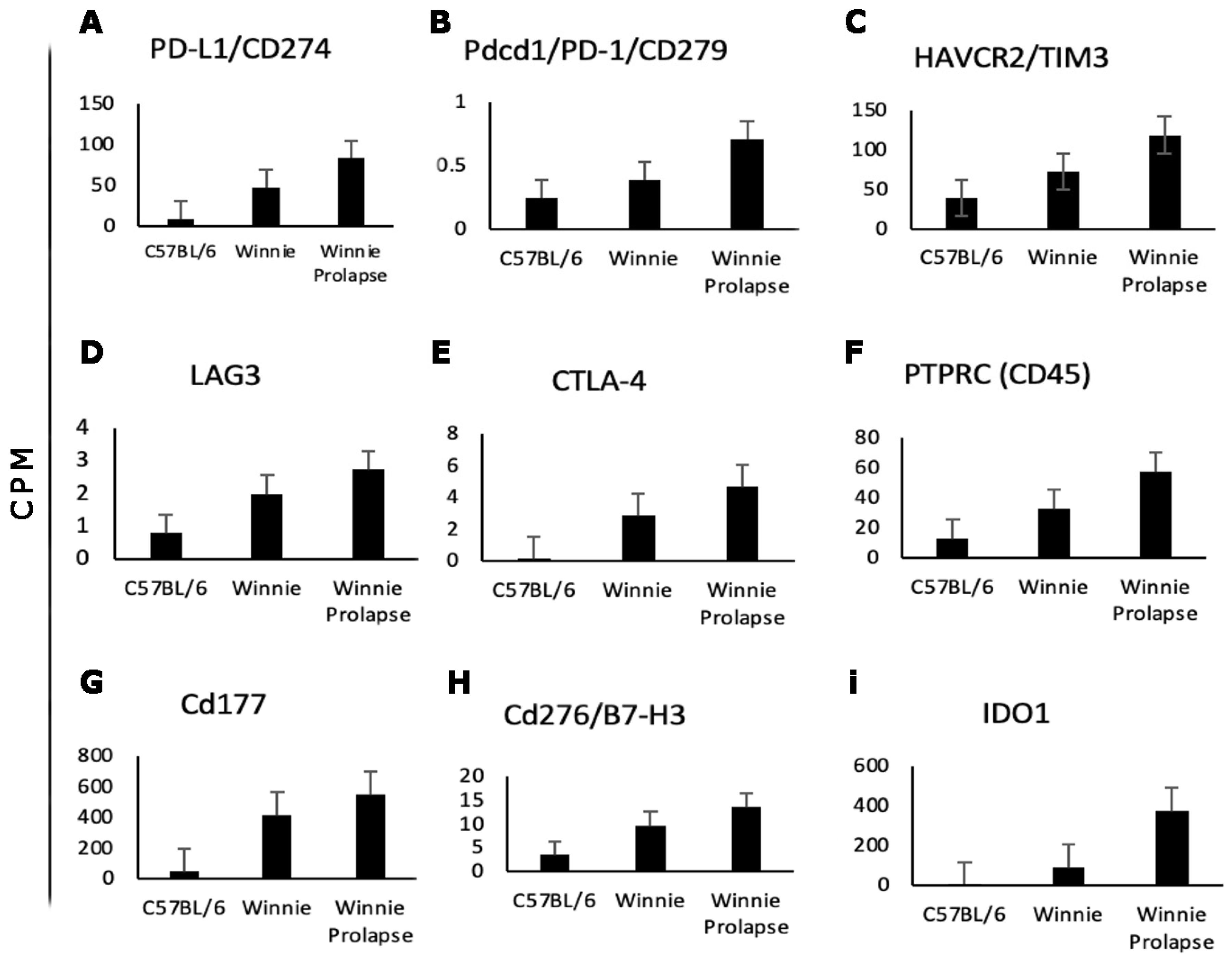
Transcriptomes of Winnie and Winnie-Prolapse Mice Are Highly Expressed in Cancer Genes (Oncogenes)
3.3. Other Genes Linked to CAC and CRC
4. Future Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cebon, J. Perspective: Cancer vaccines in the era of immune checkpoint blockade. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 2018, 29, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Korman, A.J.; Peggs, K.S.; Allison, J.P. Checkpoint blockade in cancer immunotherapy. Adv. Immunol. 2006, 90, 297–339. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; McKenzie, I.F.C. Cellular Mucins: Targets for Immunotherapy. Crit. Rev. Immunol. 2017, 37, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Pietersz, G.A.; McKenzie, I.F. Cell-mediated immune responses to MUC1 fusion protein coupled to mannan. Vaccine 1996, 14, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Pietersz, G.A.; Tsibanis, A.; Tsikkinis, A.; Stojanovska, L.; McKenzie, I.F.; Vassilaros, S. Dendritic cell immunotherapy: Clinical outcomes. Clin. Transl. Immunol. 2014, 3, e21. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef]
- Rahman, T.; Das, A.; Abir, M.H.; Nafiz, I.H.; Mahmud, A.R.; Sarker, M.R.; Emran, T.B.; Hassan, M.M. Cytokines and their role as immunotherapeutics and vaccine Adjuvants: The emerging concepts. Cytokine 2023, 169, 156268. [Google Scholar] [CrossRef]
- Barriga, V.; Kuol, N.; Nurgali, K.; Apostolopoulos, V. The Complex Interaction between the Tumor Micro-Environment and Immune Checkpoints in Breast Cancer. Cancers 2019, 11, 1205. [Google Scholar] [CrossRef]
- Ephraim, R.; Feehan, J.; Fraser, S.; Nurgali, K.; Apostolopoulos, V. Cancer Immunotherapy: The Checkpoint between Chronic Colitis and Colorectal Cancer. Cancers 2022, 14, 6131. [Google Scholar] [CrossRef]
- Ephraim, R.; Fraser, S.; Nurgali, K.; Apostolopoulos, V. Checkpoint Markers and Tumor Microenvironment: What Do We Know? Cancers 2022, 14, 3788. [Google Scholar] [CrossRef]
- Iso, H.; Hisakane, K.; Terashi, N.; Mikami, E.; Matsuki, S.; Sonokawa, T.; Atsumi, K.; Yoshino, N.; Nagata, K.; Seike, M.; et al. A remarkable response to combination chemotherapy with nivolumab and ipilimumab in a patient with primary pulmonary choriocarcinoma: A case report. Transl. Cancer Res. 2023, 12, 2212–2218. [Google Scholar] [CrossRef] [PubMed]
- Khadela, A.; Chavda, V.P.; Postwala, H.; Ephraim, R.; Apostolopoulos, V.; Shah, Y. Configuring Therapeutic Aspects of Immune Checkpoints in Lung Cancer. Cancers 2023, 15, 543. [Google Scholar] [CrossRef]
- Kuol, N.; Stojanovska, L.; Nurgali, K.; Apostolopoulos, V. The mechanisms tumor cells utilize to evade the host’s immune system. Maturitas 2017, 105, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kuol, N.; Stojanovska, L.; Nurgali, K.; Apostolopoulos, V. PD-1/PD-L1 in disease. Immunotherapy 2018, 10, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Mabbitt, J.; Holyer, I.D.; Roper, J.A.; Nilsson, U.J.; Zetterberg, F.R.; Vuong, L.; Mackinnon, A.C.; Pedersen, A.; Slack, R.J. Resistance to anti-PD-1/anti-PD-L1: Galectin-3 inhibition with GB1211 reverses galectin-3-induced blockade of pembrolizumab and atezolizumab binding to PD-1/PD-L1. Front. Immunol. 2023, 14, 1250559. [Google Scholar] [CrossRef] [PubMed]
- Suga, T.; Kimura, Y.; Furuya, K.; Sato, H. Hepatocellular carcinoma with gastric adenocarcinoma treated with atezolizumab and bevacizumab. Clin. Case Rep. 2023, 11, e7875. [Google Scholar] [CrossRef]
- Xie, M.; Wang, X.; Wang, P.; Liu, A.; Wen, Y.; Xiao, B. Efficacy of immune checkpoint inhibitors along with chemotherapy in non-small cell lung cancer and the impact on adverse reactions and serum tumor markers. Am. J. Transl. Res. 2023, 15, 5276–5283. [Google Scholar]
- Gun, S.Y.; Lee, S.W.L.; Sieow, J.L.; Wong, S.C. Targeting immune cells for cancer therapy. Redox Biol. 2019, 25, 101174. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J. Current status and future directions of cancer immunotherapy. J. Cancer 2018, 9, 1773–1781. [Google Scholar] [CrossRef]
- Koury, J.; Lucero, M.; Cato, C.; Chang, L.; Geiger, J.; Henry, D.; Hernandez, J.; Hung, F.; Kaur, P.; Teskey, G.; et al. Immunotherapies: Exploiting the Immune System for Cancer Treatment. J. Immunol. Res. 2018, 2018, 9585614. [Google Scholar] [CrossRef]
- Johnson, D.B.; Nebhan, C.A.; Moslehi, J.J.; Balko, J.M. Immune-checkpoint inhibitors: Long-term implications of toxicity. Nat. Rev. Clin. Oncol. 2022, 19, 254–267. [Google Scholar] [CrossRef]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef]
- Kreamer, K.M. Immune Checkpoint Blockade: A New Paradigm in Treating Advanced Cancer. J. Adv. Pract. Oncol. 2014, 5, 418–431. [Google Scholar] [CrossRef]
- Kaser, A.; Zeissig, S.; Blumberg, R.S. Inflammatory bowel disease. Annu. Rev. Immunol. 2010, 28, 573–621. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Götz, M.; Herrlinger, K.; Steurer, W.; Stange, E.F. Inflammatory Bowel Disease. Dtsch. Arztebl. Int. 2016, 113, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, T.L.; Friel, C.M. Colonic crohn disease. Clin. Colon Rectal. Surg. 2013, 26, 84–89. [Google Scholar] [CrossRef]
- Dave, M.; Papadakis, K.A.; Faubion, W.A., Jr. Immunology of inflammatory bowel disease and molecular targets for biologics. Gastroenterol. Clin. N. Am. 2014, 43, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Kim, S.Y. The Era of Janus Kinase Inhibitors for Inflammatory Bowel Disease Treatment. Int. J. Mol. Sci. 2021, 22, 11322. [Google Scholar] [CrossRef]
- Yun, S.M.; Kim, S.H.; Kim, E.H. The Molecular Mechanism of Transforming Growth Factor-β Signaling for Intestinal Fibrosis: A Mini-Review. Front. Pharmacol. 2019, 10, 162. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Dotan, I.; Allez, M.; Danese, S.; Keir, M.; Tole, S.; McBride, J. The role of integrins in the pathogenesis of inflammatory bowel disease: Approved and investigational anti-integrin therapies. Med. Res. Rev. 2020, 40, 245–262. [Google Scholar] [CrossRef]
- Lee, T.W.; Fedorak, R.N. Tumor necrosis factor-α monoclonal antibodies in the treatment of inflammatory bowel disease: Clinical practice pharmacology. Gastroenterol. Clin. N. Am. 2010, 39, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, G.R. Comprehensive review: Antitumor necrosis factor agents in inflammatory bowel disease and factors implicated in treatment response. Ther. Adv. Gastroenterol. 2013, 6, 269–293. [Google Scholar] [CrossRef]
- Phipps, A.I.; Lindor, N.M.; Jenkins, M.A.; Baron, J.A.; Win, A.K.; Gallinger, S.; Gryfe, R.; Newcomb, P.A. Colon and rectal cancer survival by tumor location and microsatellite instability: The Colon Cancer Family Registry. Dis. Colon Rectum 2013, 56, 937–944. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef]
- Hossain, M.S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.K.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef] [PubMed]
- Hnatyszyn, A.; Hryhorowicz, S.; Kaczmarek-Ryś, M.; Lis, E.; Słomski, R.; Scott, R.J.; Pławski, A. Colorectal carcinoma in the course of inflammatory bowel diseases. Hered. Cancer Clin. Pract. 2019, 17, 18. [Google Scholar] [CrossRef]
- Low, D.; Mino-Kenudson, M.; Mizoguchi, E. Recent advancement in understanding colitis-associated tumorigenesis. Inflamm. Bowel Dis. 2014, 20, 2115–2123. [Google Scholar] [CrossRef]
- Sebastian, S.; Hernández, H.V.; Myrelid, P.; Kariv, R.; Tsianos, E.; Toruner, M.; Marti-Gallostra, M.; Spinelli, A.; van der Meulen-de Jong, A.E.; Yuksel, E.S. Colorectal cancer in inflammatory bowel disease: Results of the 3rd ECCO pathogenesis scientific workshop (I). J. Crohn’s Colitis 2014, 8, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yao, J.; Xie, J.; Liu, Z.; Zhou, Y.; Pan, H.; Han, W. The role of autophagy in colitis-associated colorectal cancer. Signal Transduct. Target. Ther. 2018, 3, 31. [Google Scholar] [CrossRef]
- Kanneganti, M.; Mino-Kenudson, M.; Mizoguchi, E. Animal Models of Colitis-Associated Carcinogenesis. J. Biomed. Biotechnol. 2011, 2011, 342637. [Google Scholar] [CrossRef]
- Potack, J.; Itzkowitz, S.H. Colorectal cancer in inflammatory bowel disease. Gut Liver 2008, 2, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Barkin, J.A.; Sussman, D.A.; Abreu, M.T. Chromoendoscopy and advanced imaging technologies for surveillance of patients with IBD. Gastroenterol. Hepatol. 2012, 8, 796. [Google Scholar]
- Grivennikov, S.I. Inflammation and colorectal cancer: Colitis-associated neoplasia. Semin. Immunopathol. 2013, 35, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Long, A.G.; Lundsmith, E.T.; Hamilton, K.E. Inflammation and Colorectal Cancer. Curr. Color. Cancer Rep. 2017, 13, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Itzkowitz, S.H.; Yio, X. Inflammation and Cancer IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2004, 287, G7–G17. [Google Scholar] [CrossRef] [PubMed]
- Randall-Demllo, S.; Fernando, R.; Brain, T.; Sohal, S.S.; Cook, A.L.; Guven, N.; Kunde, D.; Spring, K.; Eri, R. Characterisation of colonic dysplasia-like epithelial atypia in murine colitis. World J. Gastroenterol. 2016, 22, 8334–8348. [Google Scholar] [CrossRef]
- Murphy, K.T.; Lynch, G.S. Update on emerging drugs for cancer cachexia. Expert Opin. Emerg. Drugs 2009, 14, 619–632. [Google Scholar] [CrossRef]
- Shibata, M.; Fukahori, M.; Kasamatsu, E.; Machii, K.; Hamauchi, S. A Retrospective Cohort Study to Investigate the Incidence of Cachexia During Chemotherapy in Patients with Colorectal Cancer. Adv. Ther. 2020, 37, 5010–5022. [Google Scholar] [CrossRef]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef]
- Kooshkaki, O.; Derakhshani, A.; Hosseinkhani, N.; Torabi, M.; Safaei, S.; Brunetti, O.; Racanelli, V.; Silvestris, N.; Baradaran, B. Combination of Ipilimumab and Nivolumab in Cancers: From Clinical Practice to Ongoing Clinical Trials. Int. J. Mol. Sci. 2020, 21, 4427. [Google Scholar] [CrossRef]
- Zhen, Y.; Luo, C.; Zhang, H. Early detection of ulcerative colitis-associated colorectal cancer. Gastroenterol. Rep. 2018, 6, 83–92. [Google Scholar] [CrossRef]
- Axelrad, J.E.; Lichtiger, S.; Yajnik, V. Inflammatory bowel disease and cancer: The role of inflammation, immunosuppression, and cancer treatment. World J. Gastroenterol. 2016, 22, 4794–4801. [Google Scholar] [CrossRef] [PubMed]
- Currey, N.; Jahan, Z.; Caldon, C.E.; Tran, P.N.; Benthani, F.; De Lacavalerie, P.; Roden, D.L.; Gloss, B.S.; Campos, C.; Bean, E.G.; et al. Mouse Model of Mutated in Colorectal Cancer Gene Deletion Reveals Novel Pathways in Inflammation and Cancer. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 819–839. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.C.; Itzkowitz, S.H. Colorectal Cancer in Inflammatory Bowel Disease: Mechanisms and Management. Gastroenterology 2022, 162, 715–730.e713. [Google Scholar] [CrossRef]
- Heazlewood, C.K.; Cook, M.C.; Eri, R.; Price, G.R.; Tauro, S.B.; Taupin, D.; Thornton, D.J.; Png, C.W.; Crockford, T.L.; Cornall, R.J.; et al. Aberrant mucin assembly in mice causes endoplasmic reticulum stress and spontaneous inflammation resembling ulcerative colitis. PLoS Med. 2008, 5, e54. [Google Scholar] [CrossRef]
- Kang, Y.; Park, H.; Choe, B.H.; Kang, B. The Role and Function of Mucins and Its Relationship to Inflammatory Bowel Disease. Front. Med. 2022, 9, 848344. [Google Scholar] [CrossRef]
- Dalal, S.R.; Chang, E.B. The microbial basis of inflammatory bowel diseases. J. Clin. Investig. 2014, 124, 4190–4196. [Google Scholar] [CrossRef]
- Stavely, R.; Fraser, S.; Sharma, S.; Rahman, A.A.; Stojanovska, V.; Sakkal, S.; Apostolopoulos, V.; Bertrand, P.; Nurgali, K. The Onset and Progression of Chronic Colitis Parallels Increased Mucosal Serotonin Release via Enterochromaffin Cell Hyperplasia and Downregulation of the Serotonin Reuptake Transporter. Inflamm. Bowel Dis. 2018, 24, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Stavely, R.; Rahman, A.A.; Sahakian, L.; Prakash, M.D.; Robinson, A.M.; Hassanzadeganroudsari, M.; Filippone, R.T.; Fraser, S.; Eri, R.; Bornstein, J.C.; et al. Divergent Adaptations in Autonomic Nerve Activity and Neuroimmune Signaling Associated With the Severity of Inflammation in Chronic Colitis. Inflamm. Bowel Dis. 2022, 28, 1229–1243. [Google Scholar] [CrossRef]
- Tsyganov, K.; James Perry, A.; Kenneth Archer, S.; Powell, D. RNAsik: A Pipeline for complete and reproducible RNA-seq analysis that runs anywhere with speed and ease. J. Open Source Softw. 2018, 3, 583. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Law, C.; Alhamdoosh, M.; Su, S.; Dong, X.; Tian, L.; Smyth, G.; Ritchie, M. RNA-seq analysis is easy as 1-2-3 with limma, Glimma and edgeR [version 3; peer review: 3 approved]. F1000Research 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Moniruzzaman, M.; Wong, K.Y.; Wiid, P.; Harding, A.; Giri, R.; Tong, W.H.; Creagh, J.; Begun, J.; McGuckin, M.A.; et al. Gut microbiota shape the inflammatory response in mice with an epithelial defect. Gut Microbes 2021, 13, 1887720. [Google Scholar] [CrossRef] [PubMed]
- Perše, M. Oxidative stress in the pathogenesis of colorectal cancer: Cause or consequence? Biomed. Res. Int. 2013, 2013, 725710. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.M.; Gondalia, S.V.; Karpe, A.V.; Eri, R.; Beale, D.J.; Morrison, P.D.; Palombo, E.A.; Nurgali, K. Fecal Microbiota and Metabolome in a Mouse Model of Spontaneous Chronic Colitis: Relevance to Human Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2016, 22, 2767–2787. [Google Scholar] [CrossRef]
- Gimple, R.C.; Wang, X. RAS: Striking at the Core of the Oncogenic Circuitry. Front. Oncol. 2019, 9, 965. [Google Scholar] [CrossRef]
- Gutierrez-Hoya, A.; Soto-Cruz, I. Role of the JAK/STAT Pathway in Cervical Cancer: Its Relationship with HPV E6/E7 Oncoproteins. Cells 2020, 9, 2297. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Wang, R.C.; Cheng, K.; Ring, B.Z.; Su, L. Roles of Rap1 signaling in tumor cell migration and invasion. Cancer Biol. Med. 2017, 14, 90–99. [Google Scholar] [CrossRef]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef]
- Han, Z.; Meng, L.; Huang, X.; Tan, J.; Liu, W.; Chen, W.; Zou, Y.; Cai, Y.; Huang, S.; Chen, A.; et al. Inhibition of p38 MAPK increases the sensitivity of 5-fluorouracil-resistant SW480 human colon cancer cells to noscapine. Oncol. Lett. 2022, 23, 52. [Google Scholar] [CrossRef]
- Koveitypour, Z.; Panahi, F.; Vakilian, M.; Peymani, M.; Seyed Forootan, F.; Nasr Esfahani, M.H.; Ghaedi, K. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Hansmannel, F.; Arnone, D.; Choukour, M.; Ndiaye, N.C.; Kokten, T.; Houlgatte, R.; Peyrin-Biroulet, L. Genomic and molecular alterations in human inflammatory bowel disease-associated colorectal cancer. United Eur. Gastroenterol. J. 2020, 8, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Seidel, J.A.; Otsuka, A.; Kabashima, K. Anti-PD-1 and Anti-CTLA-4 Therapies in Cancer: Mechanisms of Action, Efficacy, and Limitations. Front. Oncol. 2018, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Villalobo, A.; Berchtold, M.W. The Role of Calmodulin in Tumor Cell Migration, Invasiveness, and Metastasis. Int. J. Mol. Sci. 2020, 21, 765. [Google Scholar] [CrossRef] [PubMed]
- Gurel, B.; Ali, T.Z.; Montgomery, E.A.; Begum, S.; Hicks, J.; Goggins, M.; Eberhart, C.G.; Clark, D.P.; Bieberich, C.J.; Epstein, J.I.; et al. NKX3.1 as a marker of prostatic origin in metastatic tumors. Am. J. Surg. Pathol. 2010, 34, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ming, T.; Tang, S.; Ren, S.; Yang, H.; Liu, M.; Tao, Q.; Xu, H. Wnt signaling in colorectal cancer: Pathogenic role and therapeutic target. Mol. Cancer 2022, 21, 144. [Google Scholar] [CrossRef]
- Nie, X.; Liu, H.; Liu, L.; Wang, Y.D.; Chen, W.D. Emerging Roles of Wnt Ligands in Human Colorectal Cancer. Front. Oncol. 2020, 10, 1341. [Google Scholar] [CrossRef]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef]
- Fu, X.; Luo, H.; Zheng, Y.; Wang, S.; Zhong, Z.; Wang, Y.; Yang, Y. CTLA-4 immunotherapy exposes differences in immune response along with different tumor progression in colorectal cancer. Aging 2020, 12, 15656–15669. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, B.R.; Zafar, S.Y. Capecitabine in the management of colorectal cancer. Cancer Manag. Res. 2011, 3, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef]
- Derakhshani, A.; Hashemzadeh, S.; Asadzadeh, Z.; Shadbad, M.A.; Rasibonab, F.; Safarpour, H.; Jafarlou, V.; Solimando, A.G.; Racanelli, V.; Singh, P.K.; et al. Cytotoxic T-Lymphocyte Antigen-4 in Colorectal Cancer: Another Therapeutic Side of Capecitabine. Cancers 2021, 13, 2414. [Google Scholar] [CrossRef]
- Zhou, X.; Ding, X.; Li, H.; Yang, C.; Ma, Z.; Xu, G.; Yang, S.; Zhang, D.; Xie, X.; Xin, L.; et al. Upregulation of TIGIT and PD-1 in Colorectal Cancer with Mismatch-repair Deficiency. Immunol. Invest. 2021, 50, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, Y.; Qin, S.L.; Mu, Y.F.; Qi, Y.; Yu, M.H.; Zhong, M. The clinical impact of ICOS signal in colorectal cancer patients. Oncoimmunology 2016, 5, e1141857. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Dang, Q.; Liu, Z.; Shao, B.; Chen, C.; Guo, Y.; Chen, Z.; Zhou, Q.; Hu, S.; Liu, J.; et al. LINC01272/miR-876/ITGB2 axis facilitates the metastasis of colorectal cancer via epithelial-mesenchymal transition. J. Cancer 2021, 12, 3909–3919. [Google Scholar] [CrossRef]
- Korniluk, A.; Kamińska, J.; Kiszło, P.; Kemona, H.; Dymicka-Piekarska, V. Lectin adhesion proteins (P-, L- and E-selectins) as biomarkers in colorectal cancer. Biomarkers 2017, 22, 629–634. [Google Scholar] [CrossRef]
- Wang, B.; Sun, Y. SELPLG Expression Was Potentially Correlated With Metastasis and Prognosis of Osteosarcoma. Pathol. Oncol. Res. 2022, 28, 1610047. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Z.; Skrzypczynska, K.M.; Fang, Q.; Zhang, W.; O’Brien, S.A.; He, Y.; Wang, L.; Zhang, Q.; Kim, A.; et al. Single-Cell Analyses Inform Mechanisms of Myeloid-Targeted Therapies in Colon Cancer. Cell 2020, 181, 442–459.e429. [Google Scholar] [CrossRef]
- Ma, C.; Wu, W.; Lin, R.; Ge, Y.; Zhang, C.; Sun, S.; Cong, Y.; Li, X.; Liu, Z. Critical Role of CD6highCD4+ T Cells in Driving Th1/Th17 Cell Immune Responses and Mucosal Inflammation in IBD. J. Crohn’s Colitis 2019, 13, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Ruth, J.H.; Gurrea-Rubio, M.; Athukorala, K.S.; Rasmussen, S.M.; Weber, D.P.; Randon, P.M.; Gedert, R.J.; Lind, M.E.; Amin, M.A.; Campbell, P.L.; et al. CD6 is a target for cancer immunotherapy. JCI Insight 2021, 6, e145662. [Google Scholar] [CrossRef]
- Kumara, H.; Bellini, G.A.; Caballero, O.L.; Herath, S.A.C.; Su, T.; Ahmed, A.; Njoh, L.; Cekic, V.; Whelan, R.L. P-Cadherin (CDH3) is overexpressed in colorectal tumors and has potential as a serum marker for colorectal cancer monitoring. Oncoscience 2017, 4, 139–147. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.Y.; Jang, G.B.; Choi, J.H.; Kim, J.H.; Lee, C.J.; Lee, S.; Baek, J.H.; Park, K.K.; Kim, J.M.; et al. Aberrant activation of the CD45-Wnt signaling axis promotes stemness and therapy resistance in colorectal cancer cells. Theranostics 2021, 11, 8755–8770. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liang, J.; Liu, Z.; Zhang, C.; Wang, Y.; Watson, A.H.; Zhou, C.; Zhang, F.; Wu, K.; Zhang, F.; et al. The Role of CD276 in Cancers. Front. Oncol. 2021, 11, 654684. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Peng, K.; Song, Y.; Yang, W.; Shu, W.; Yu, T.; Yu, L.; Lin, M.; Wei, Q.; Chen, C.; et al. CD177+ neutrophils suppress epithelial cell tumourigenesis in colitis-associated cancer and predict good prognosis in colorectal cancer. Carcinogenesis 2017, 39, 272–282. [Google Scholar] [CrossRef]
- Zenonos, K.; Kyprianou, K. RAS signaling pathways, mutations and their role in colorectal cancer. World J. Gastrointest. Oncol. 2013, 5, 97–101. [Google Scholar] [CrossRef]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Dillon, M.; Lopez, A.; Lin, E.; Sales, D.; Perets, R.; Jain, P. Progress on Ras/MAPK Signaling Research and Targeting in Blood and Solid Cancers. Cancers 2021, 13, 5059. [Google Scholar] [CrossRef]
- Wei, J.; Hu, M.; Huang, K.; Lin, S.; Du, H. Roles of Proteoglycans and Glycosaminoglycans in Cancer Development and Progression. Int. J. Mol. Sci. 2020, 21, 5983. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Horgan, P.G.; McMillan, D.C.; Edwards, J. NF-κB pathways in the development and progression of colorectal cancer. Transl. Res. 2018, 197, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Browning, D.D.; Kwon, I.K.; Wang, R. cGMP-dependent protein kinases as potential targets for colon cancer prevention and treatment. Future Med. Chem. 2010, 2, 65–80. [Google Scholar] [CrossRef]
- Looi, C.K.; Hii, L.W.; Ngai, S.C.; Leong, C.O.; Mai, C.W. The Role of Ras-Associated Protein 1 (Rap1) in Cancer: Bad Actor or Good Player? Biomedicines 2020, 8, 334. [Google Scholar] [CrossRef]
- Li, H.; Han, G.; Li, X.; Li, B.; Wu, B.; Jin, H.; Wu, L.; Wang, W. MAPK-RAP1A Signaling Enriched in Hepatocellular Carcinoma Is Associated with Favorable Tumor-Infiltrating Immune Cells and Clinical Prognosis. Front. Oncol. 2021, 11, 649980. [Google Scholar] [CrossRef]
- Saunders, A.S.; Bender, D.E.; Ray, A.L.; Wu, X.; Morris, K.T. Colony-stimulating factor 3 signaling in colon and rectal cancers: Immune response and CMS classification in TCGA data. PLoS ONE 2021, 16, e0247233. [Google Scholar] [CrossRef]
- Pan, Z.; He, Y.; Zhu, W.; Xu, T.; Hu, X.; Huang, P. A Dynamic Transcription Factor Signature Along the Colorectal Adenoma-Carcinoma Sequence in Patients With Co-Occurrent Adenoma and Carcinoma. Front. Oncol. 2021, 11, 597447. [Google Scholar] [CrossRef]
- Cui, J.; Tian, J.; Wang, W.; He, T.; Li, X.; Gu, C.; Wang, L.; Wu, J.; Shang, A. IGF2BP2 promotes the progression of colorectal cancer through a YAP-dependent mechanism. Cancer Sci. 2021, 112, 4087–4099. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Qiang, P. The role of IGF2BP2, an m6A reader gene, in human metabolic diseases and cancers. Cancer Cell Int. 2021, 21, 99. [Google Scholar] [CrossRef]
- Chen, H.; Liu, Y.; Jiang, C.J.; Chen, Y.M.; Li, H.; Liu, Q.A. Calcium-Activated Chloride Channel A4 (CLCA4) Plays Inhibitory Roles in Invasion and Migration through Suppressing Epithelial-Mesenchymal Transition via PI3K/AKT Signaling in Colorectal Cancer. Med. Sci. Monit. 2019, 25, 4176–4185. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Chen, W.; Zhao, J.; Fang, Y.; Lin, J. Downregulation of CLCA4 expression is associated with the development and progression of colorectal cancer. Oncol. Lett. 2020, 20, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhang, S.; Li, S.; Wang, Y.; Zhang, X.; Xue, F. Expression of the CLCA4 Gene in Esophageal Carcinoma and Its Impact on the Biologic Function of Esophageal Carcinoma Cells. J. Oncol. 2021, 2021, 1649344. [Google Scholar] [CrossRef] [PubMed]
- Ling, A.; Löfgren-Burström, A.; Larsson, P.; Li, X.; Wikberg, M.L.; Öberg, Å.; Stenling, R.; Edin, S.; Palmqvist, R. TAP1 down-regulation elicits immune escape and poor prognosis in colorectal cancer. Oncoimmunology 2017, 6, e1356143. [Google Scholar] [CrossRef]
- Li, X.; Zeng, S.; Ding, Y.; Nie, Y.; Yang, M. Comprehensive Analysis of the Potential Immune-Related Biomarker Transporter Associated with Antigen Processing 1 That Inhibits Metastasis and Invasion of Ovarian Cancer Cells. Front. Mol. Biosci. 2021, 8, 763958. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ephraim, R.; Fraser, S.; Devereaux, J.; Stavely, R.; Feehan, J.; Eri, R.; Nurgali, K.; Apostolopoulos, V. Differential Gene Expression of Checkpoint Markers and Cancer Markers in Mouse Models of Spontaneous Chronic Colitis. Cancers 2023, 15, 4793. https://doi.org/10.3390/cancers15194793
Ephraim R, Fraser S, Devereaux J, Stavely R, Feehan J, Eri R, Nurgali K, Apostolopoulos V. Differential Gene Expression of Checkpoint Markers and Cancer Markers in Mouse Models of Spontaneous Chronic Colitis. Cancers. 2023; 15(19):4793. https://doi.org/10.3390/cancers15194793
Chicago/Turabian StyleEphraim, Ramya, Sarah Fraser, Jeannie Devereaux, Rhian Stavely, Jack Feehan, Rajaraman Eri, Kulmira Nurgali, and Vasso Apostolopoulos. 2023. "Differential Gene Expression of Checkpoint Markers and Cancer Markers in Mouse Models of Spontaneous Chronic Colitis" Cancers 15, no. 19: 4793. https://doi.org/10.3390/cancers15194793
APA StyleEphraim, R., Fraser, S., Devereaux, J., Stavely, R., Feehan, J., Eri, R., Nurgali, K., & Apostolopoulos, V. (2023). Differential Gene Expression of Checkpoint Markers and Cancer Markers in Mouse Models of Spontaneous Chronic Colitis. Cancers, 15(19), 4793. https://doi.org/10.3390/cancers15194793