

EPHA3 Could Be a Novel Prognosis Biomarker and Correlates with Immune Infiltrates in Bladder Cancer

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Data Source
2.2. Survival Curves, ROC Curves, and Clinicopathologic Features Analysis
2.3. Co-Expressed Gene Exploration and Enrichment
2.4. Cell Lines and Cell Culture
2.5. Cell Transfection
2.6. Cell Proliferation Assay
2.7. Colony Formation Assay
2.8. Cell Migration Assay
2.9. Cell Invasion Assay
2.10. Cell Apoptosis Assay
2.11. Animal Experiments
2.12. RNA Isolation and Real-Time Quantitative PCR (RT-qPCR)
2.13. Western Blotting
2.14. Data Source and Immune Infiltration Expression Analysis
2.15. Statistical Analysis
3. Results
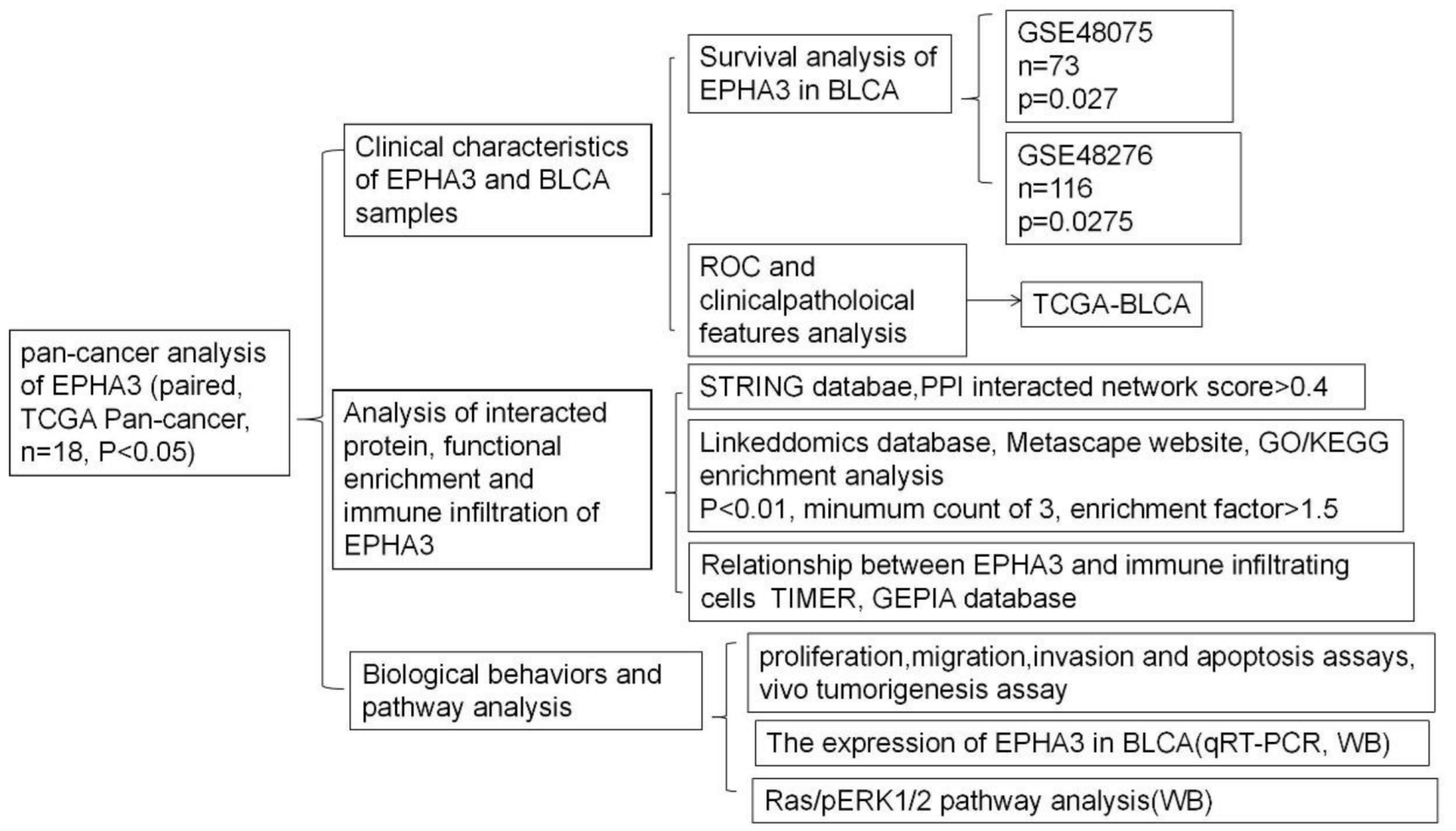
3.1. Flow Charts and Patient Features
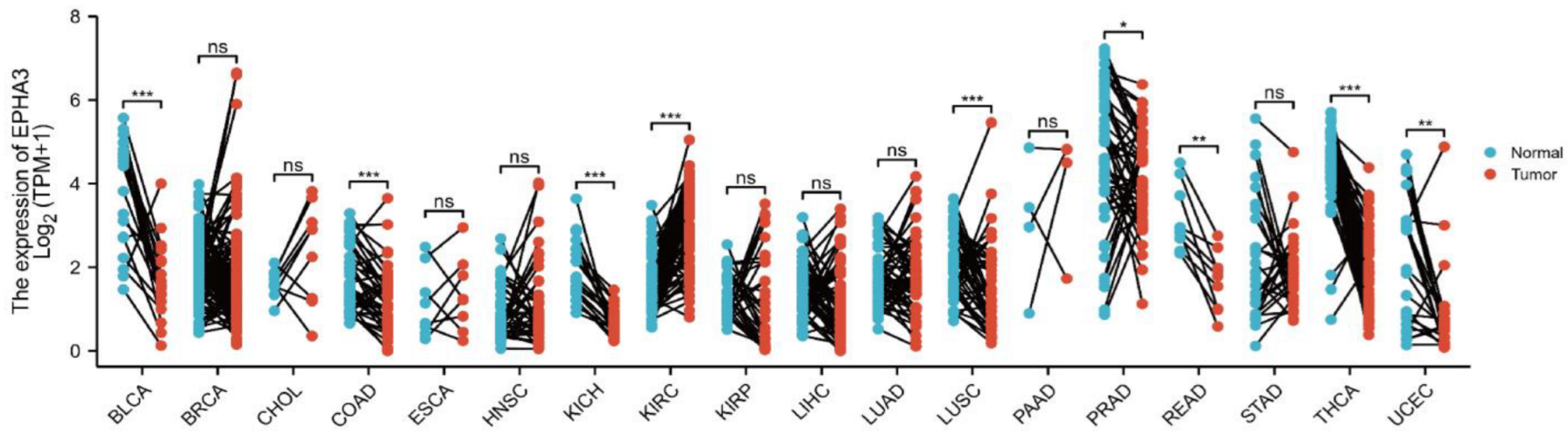
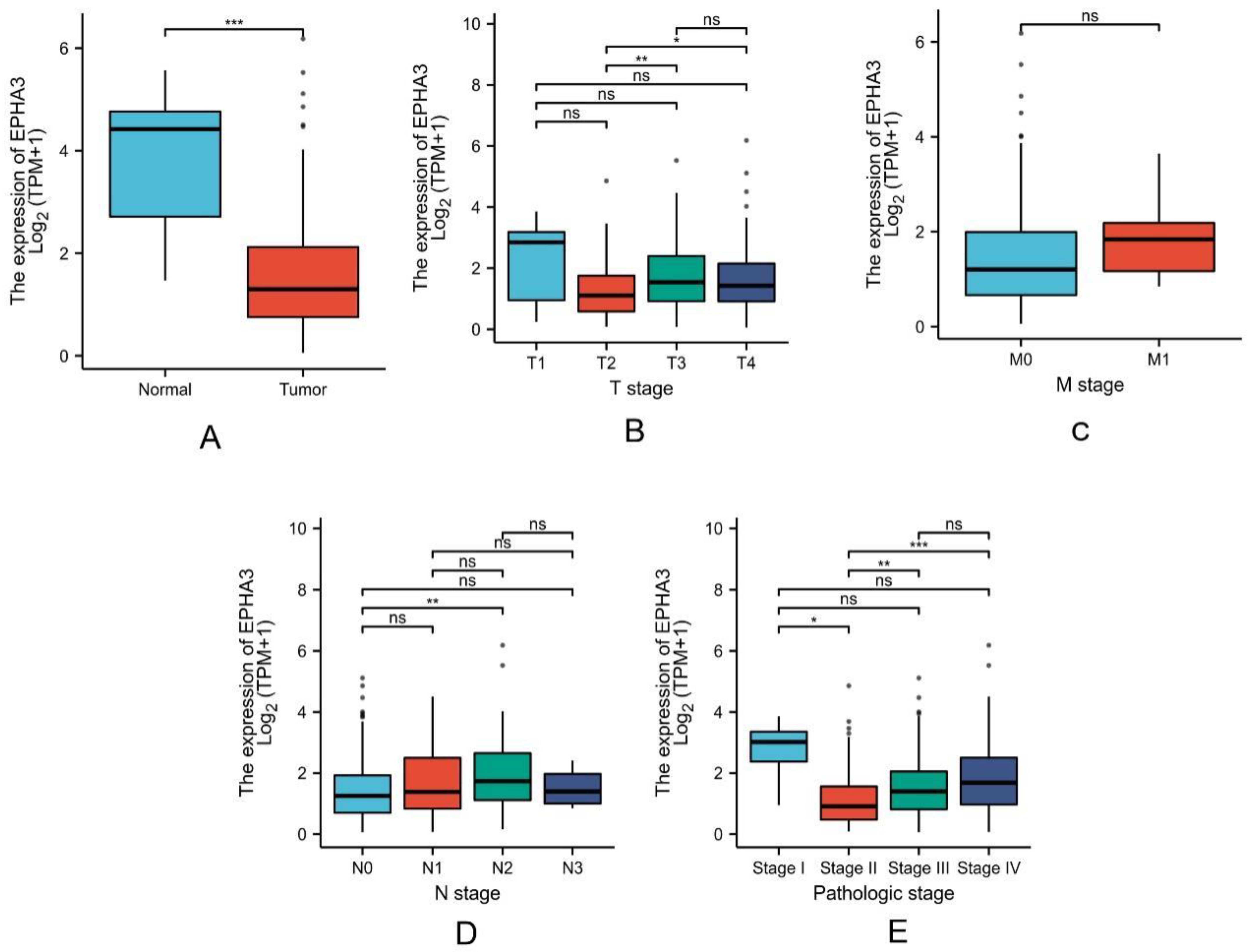
3.2. Expression Level of EPHA3 in Normal Samples Was Higher Than in BLCA
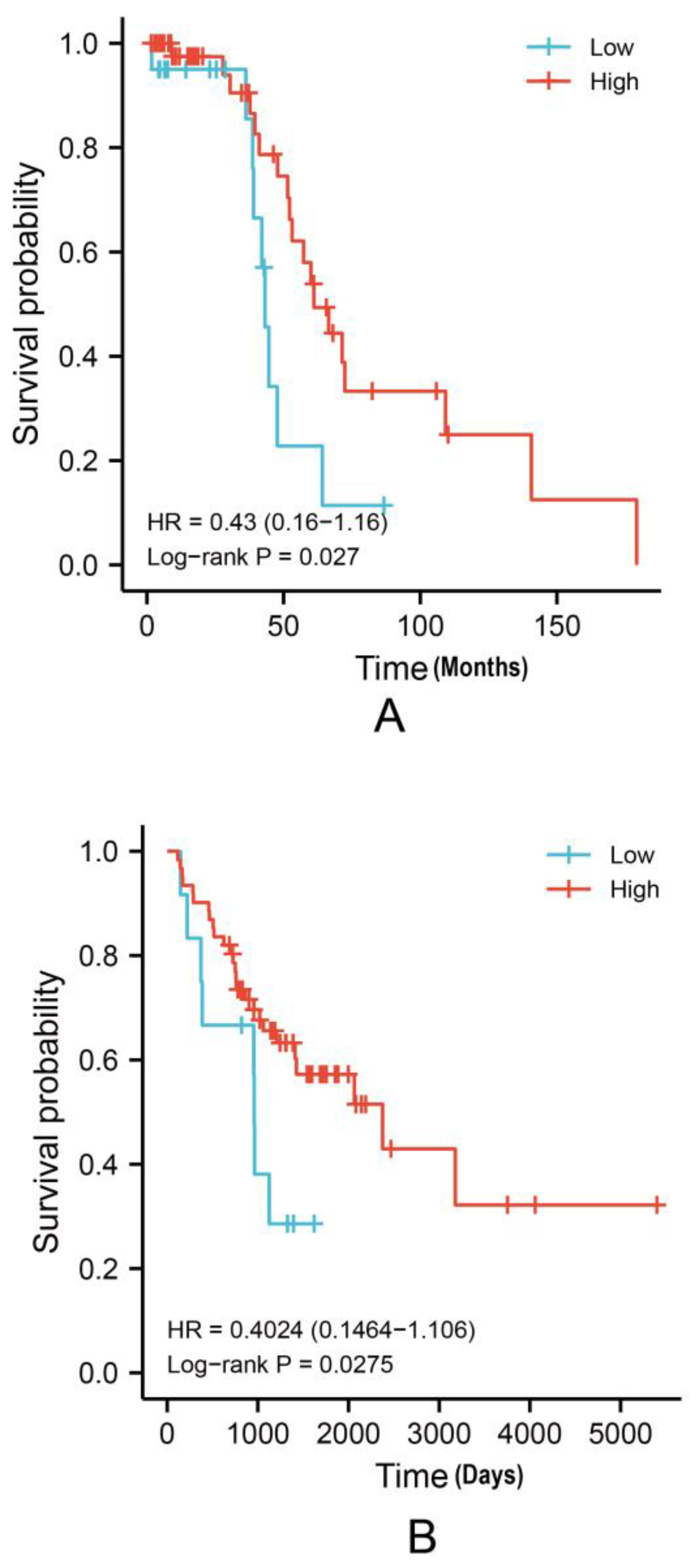
3.3. Higher EPHA3 mRNA Expression Levels May Predict a Better OS Rate in BLCA Patients
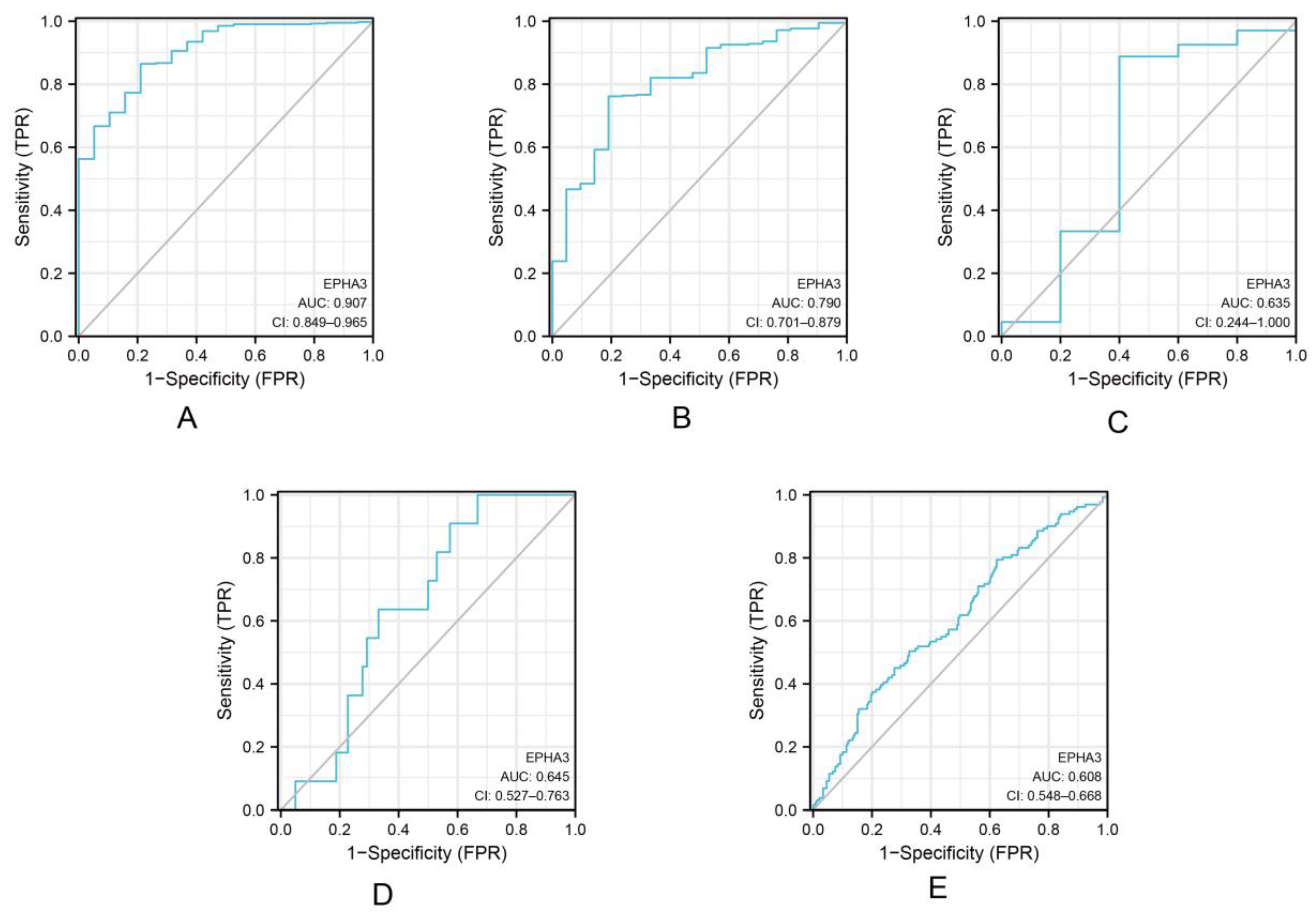
3.4. EPHA3 Was a Good Predictor of Histologic Grade and Status in BLCA
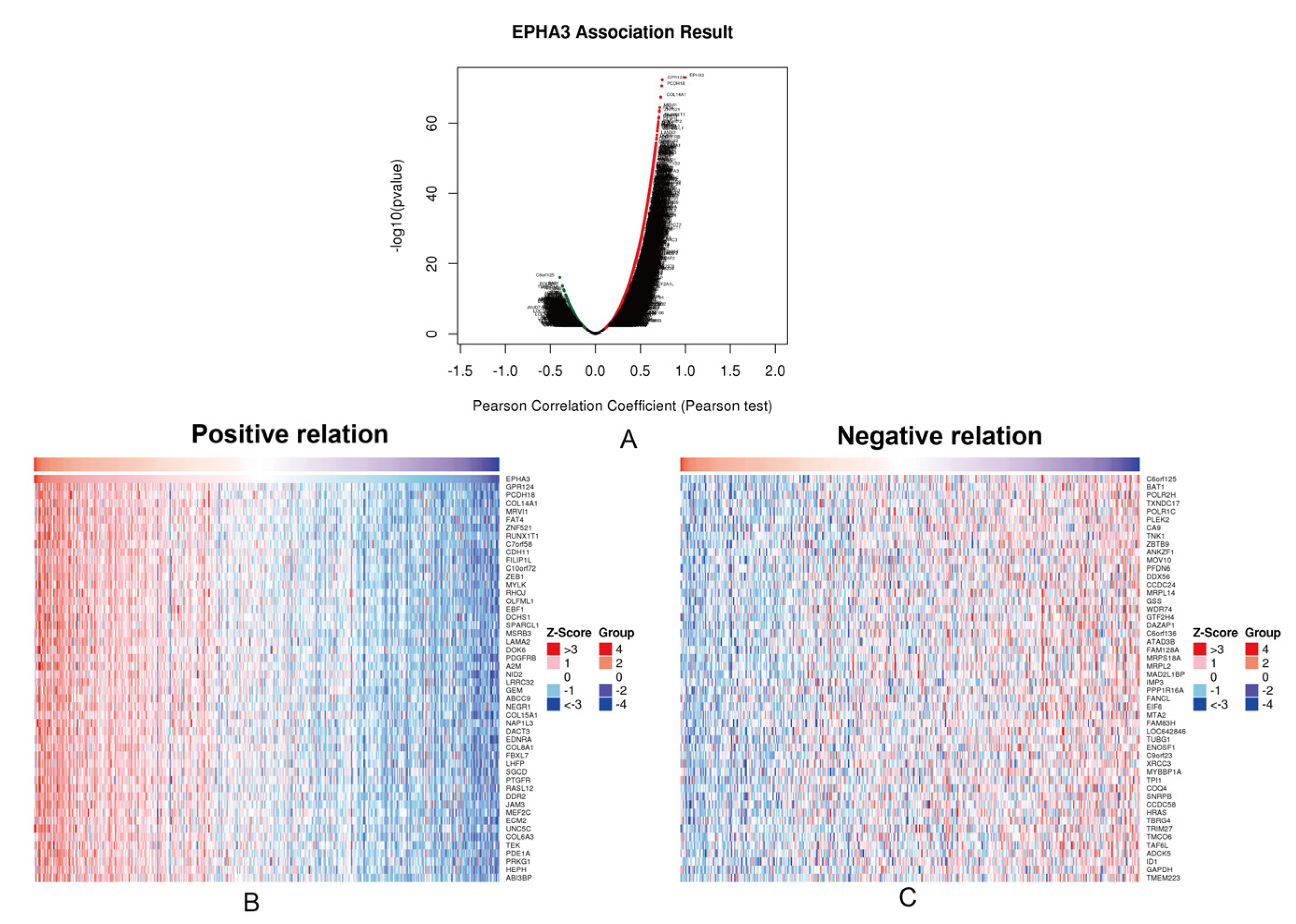
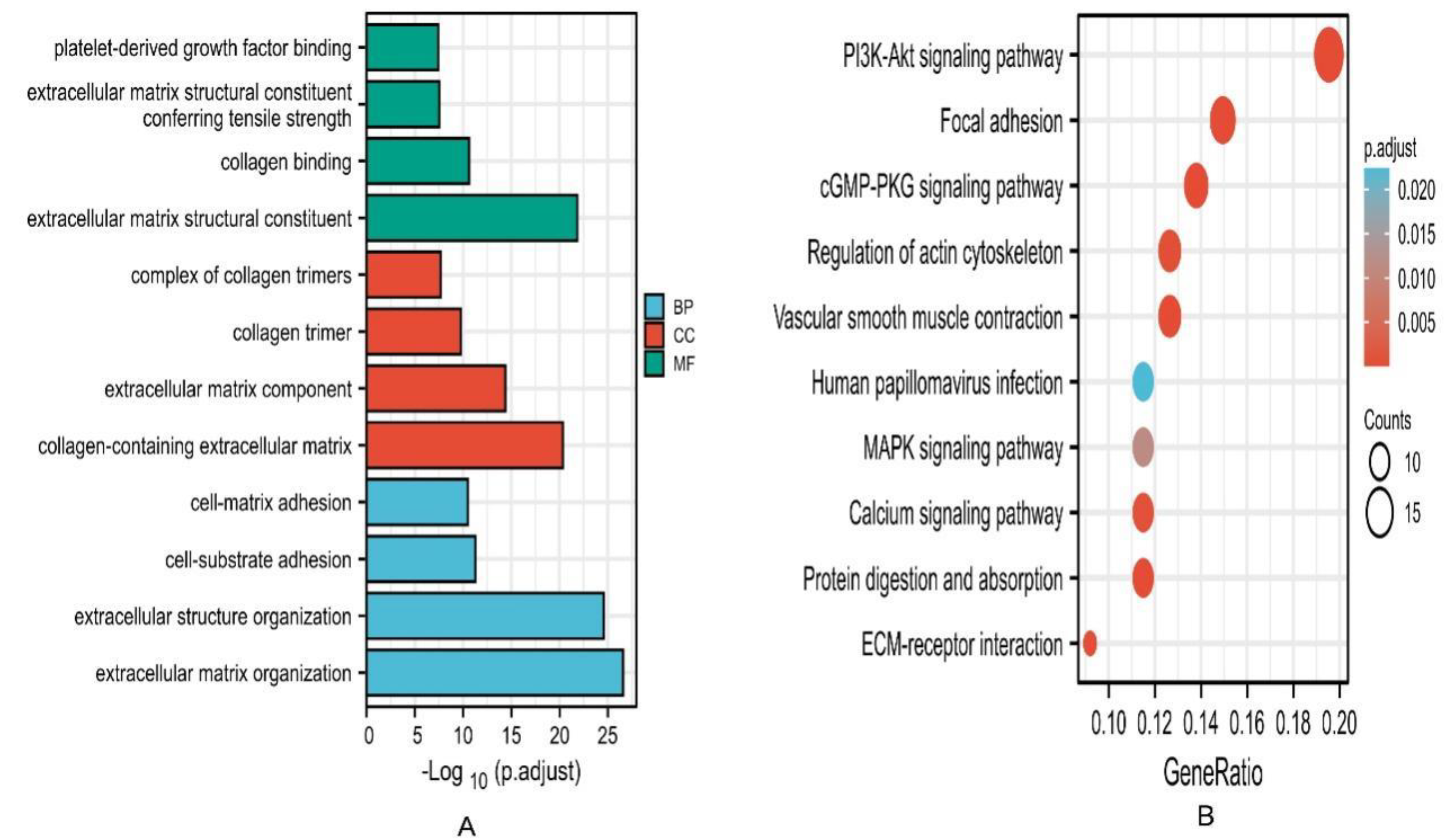
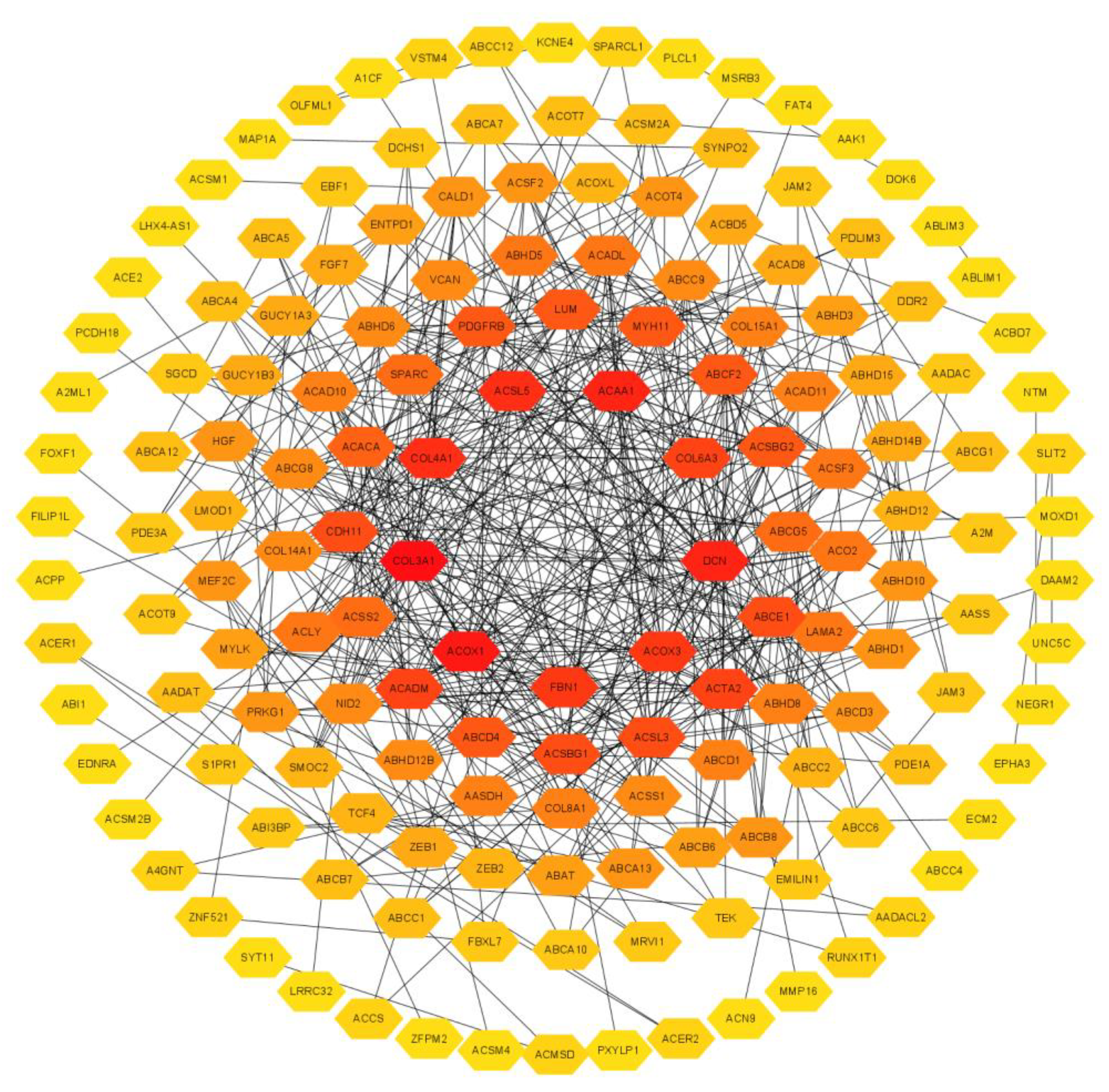
3.5. Enrichment Analysis and Interacted Genes Network Construction for EPHA3 in BLCA
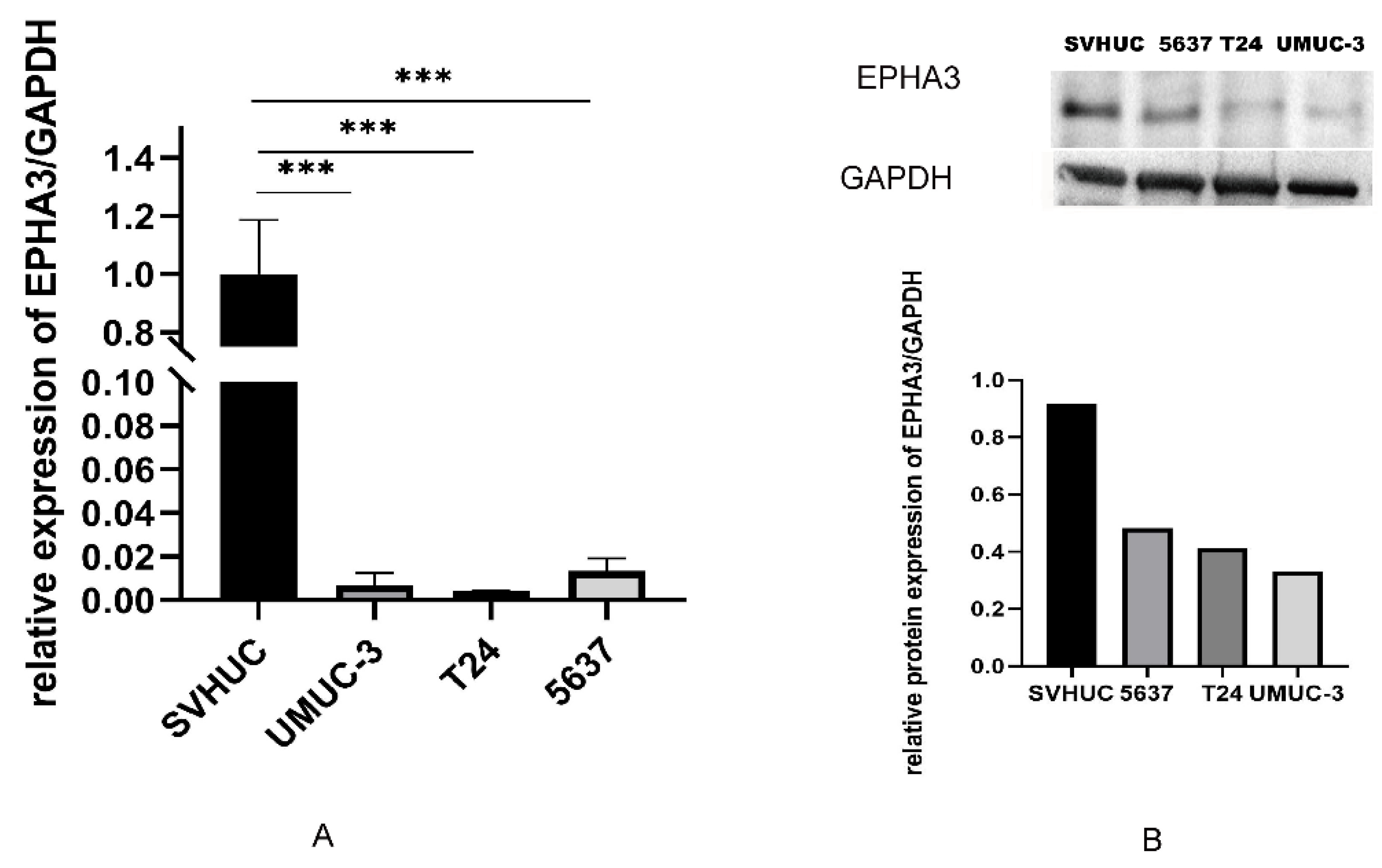
3.6. EPHA3 Expression within BLCA Cell Lines
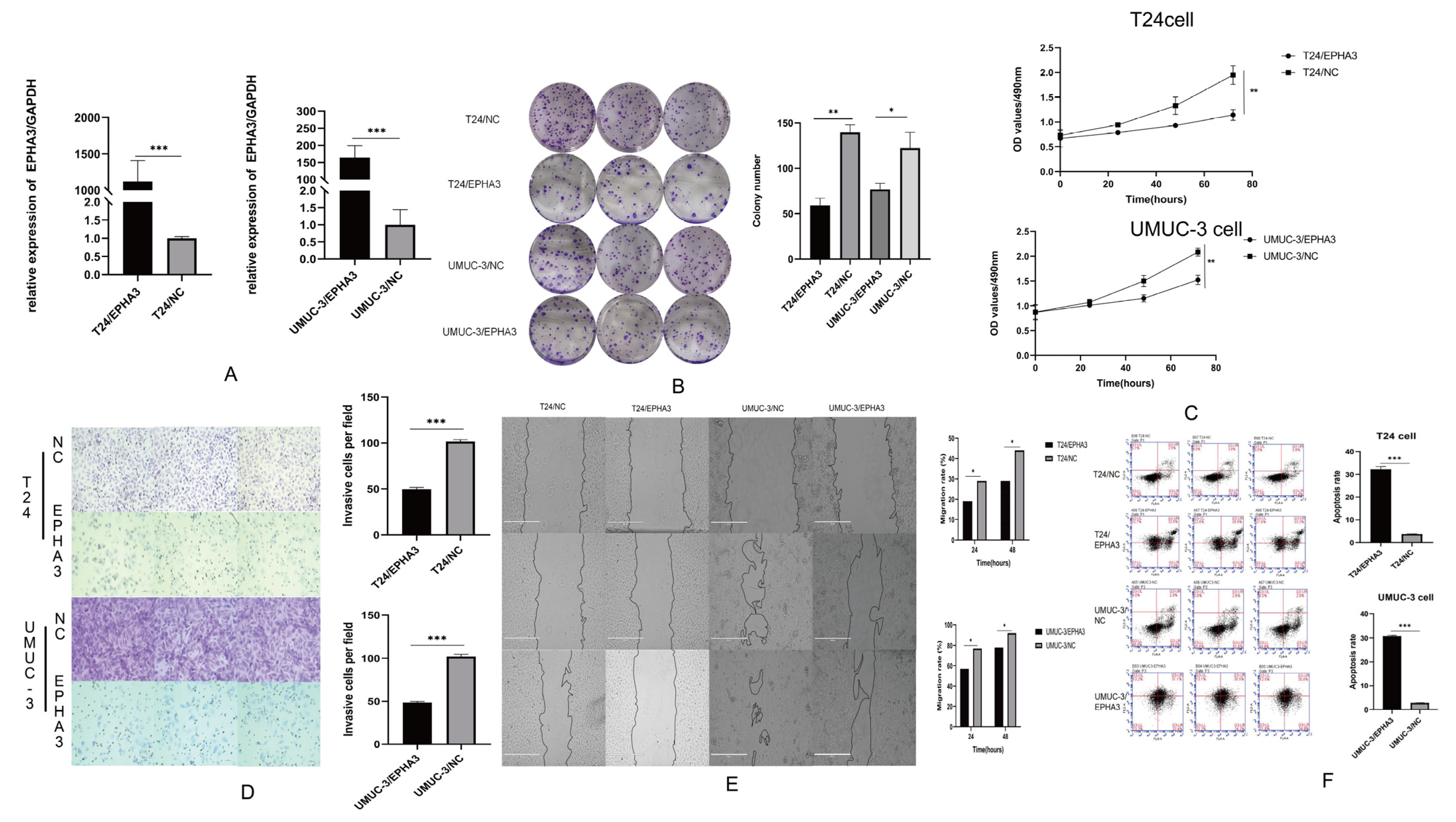
3.7. Upregulating EPHA3 Hindered BLCA Cell Growth, Invasion, Migration, and It Enhanced Their Apoptosis
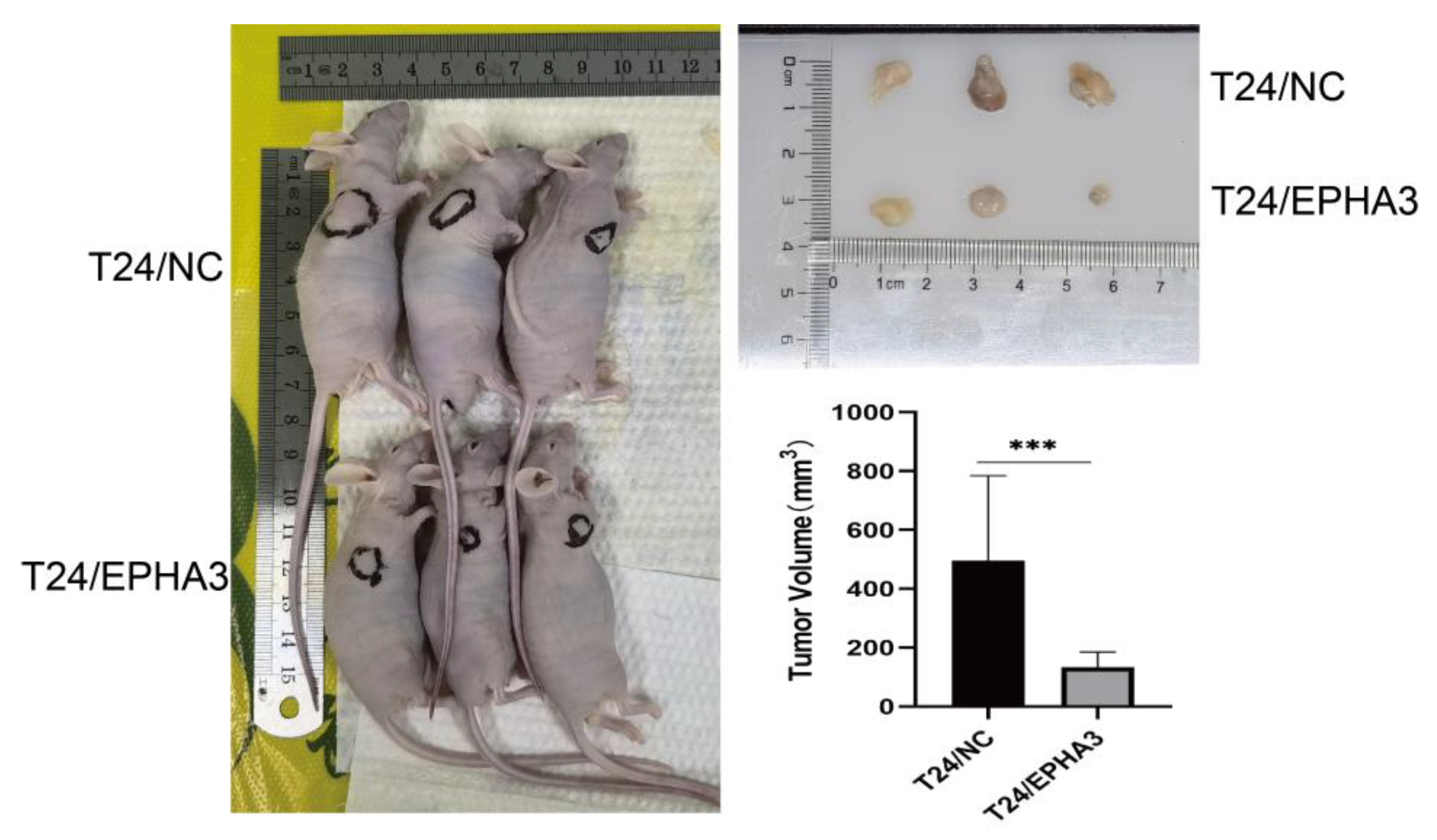
3.8. EPHA3 Inhibited BLCA Growth In Vivo
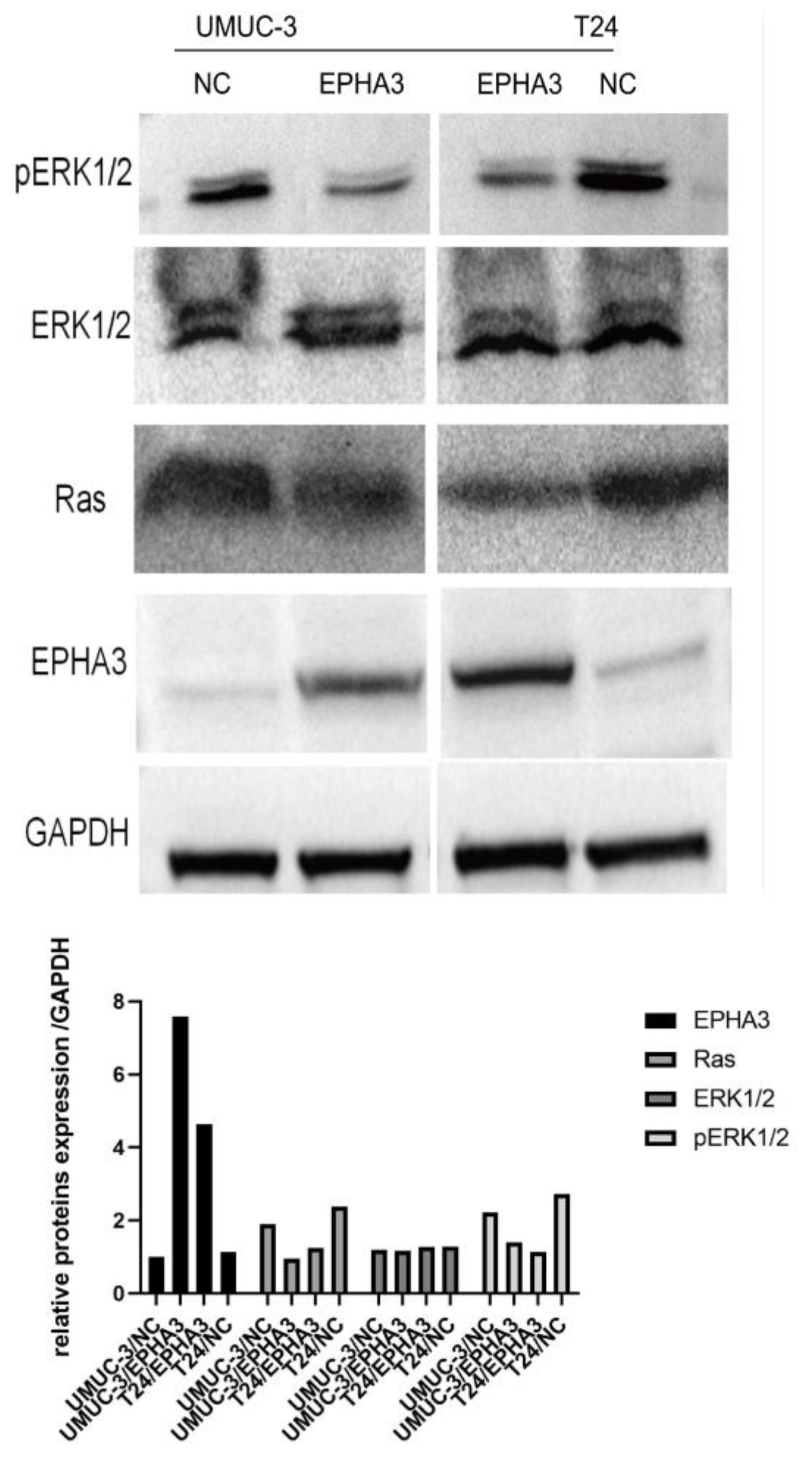
3.9. EPHA3 Regulated BLCA via the Ras/pERK1/2 Pathway
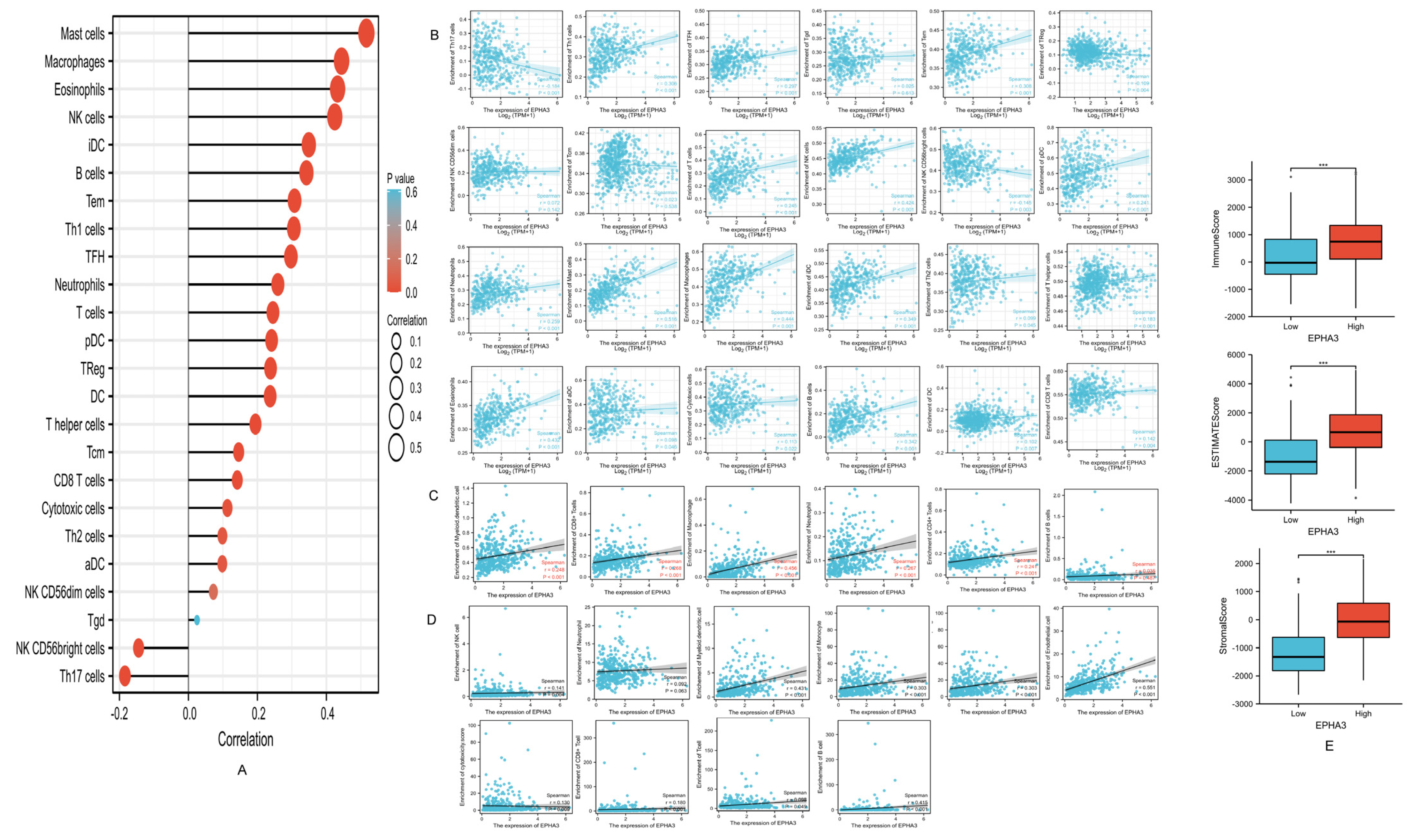
3.10. EPHA3 Was Correlated with Immune-Infiltrating Cells in BLCA
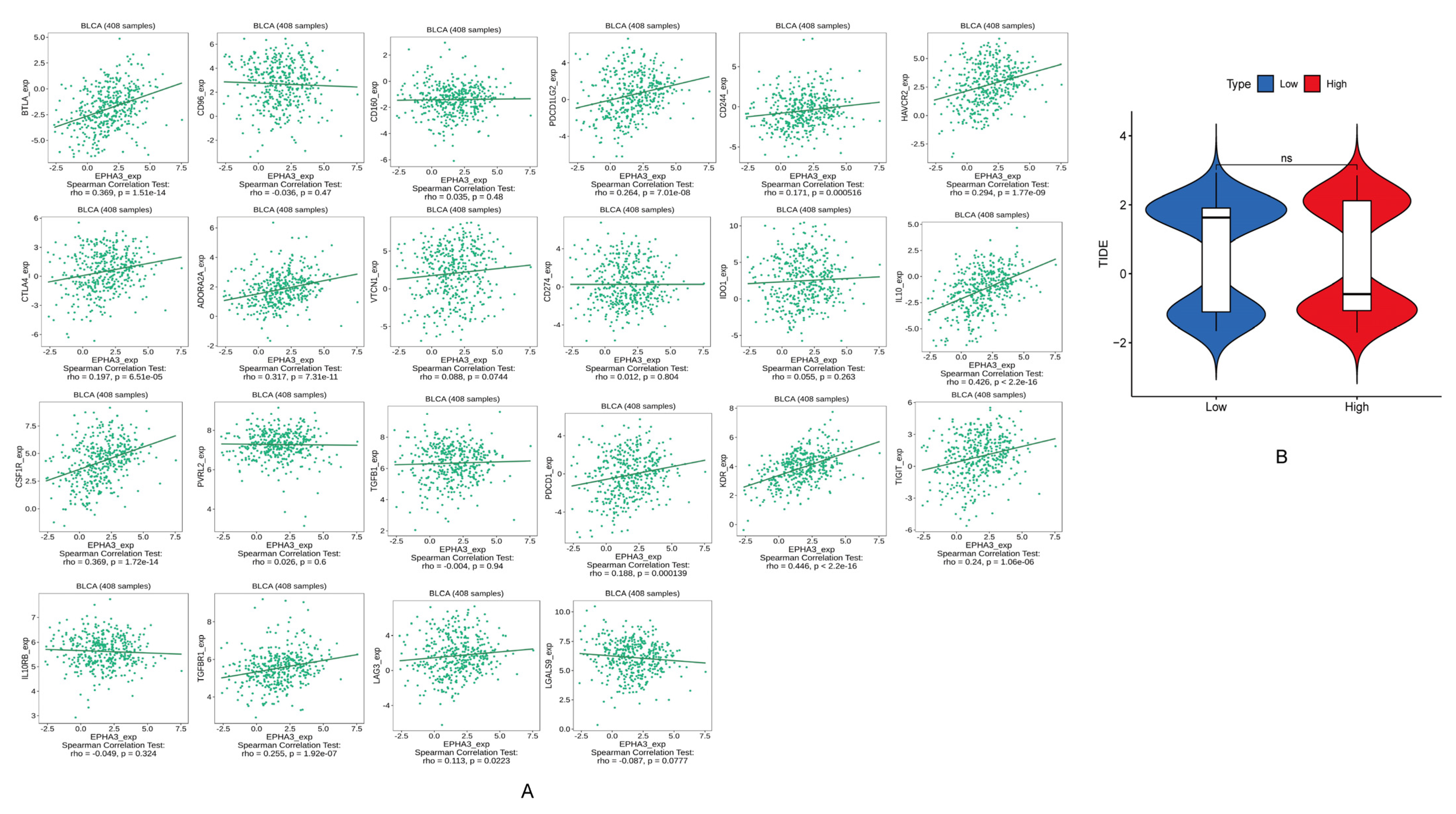
3.11. EPHA3 Was Associated with ICGs in BLCA
3.12. Correlation between EPHA3 and Immune Cells Marker Genes in BLCA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| EPHA3 | EPH receptor A3 |
| BLCA | bladder cancer |
| NMIBC | non-muscle-invasive bladder cancer |
| MIBC | muscle-invasive bladder cancer |
| TCGA | The Cancer Genome Atlas |
| ROC | receiver operating characteristic |
| GEO | Gene Expression Omnibus |
| ICG | immune checkpoint gene |
| ICI | immune checkpoint inhibitor |
| TIDE | tumor immune dysfunction and exclusion |
| OS | overall survival |
| AUC | area under ROC |
| pERK1/2 | phosphorylation-ERK 1/2 |
| ESTIMATE | Estimation of STromal and Immune cells in Malignant Tumor tissues |
| TIMER | Tumor Immune Estimation Resource |
| RTKs | Receptor tyrosine-kinases |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PPI | protein–protein interaction |
| PI | propidium iodide |
| CCK8 | counting kit-8 |
| RT-qPCR | real-time quantitative PCR |
| MCP-counter | microenvironment cell populations-counter |
References
- Babjuk, M.; Burger, M.; Compérat, E.M.; Gontero, P.; Mostafid, A.H.; Palou, J.; van Rhijn, B.W.G.; Rouprêt, M.; Shariat, S.F.; Sylvester, R.; et al. European Association of Urology Guidelines on Non-muscle-invasive Bladder Cancer (TaT1 and Carcinoma In Situ)—2019 Update. Eur. Urol. 2019, 76, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Oswald, D.; Pallauf, M.; Herrmann, T.R.W.; Netsch, C.; Becker, B.; Lehrich, K.; Miernik, A.; Schöb, D.S.; Sievert, K.D.; Gross, A.J.; et al. Transurethral resection of bladder tumors (TURBT). Urol. A 2022, 61, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Vuky, J. Combination Therapy With Immune Checkpoint Inhibitors in Urothelial Carcinoma: Current Data and Future Outlook. Oncology 2021, 35, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.S.; Boorjian, S.A.; Chou, R.; Clark, P.E.; Daneshmand, S.; Konety, B.R.; Pruthi, R.; Quale, D.Z.; Ritch, C.R.; Seigne, J.D.; et al. Diagnosis and Treatment of Non-Muscle Invasive Bladder Cancer: AUA/SUO Guideline. J. Urol. 2016, 196, 1021–1029. [Google Scholar] [CrossRef]
- Murthy, V.; Bakshi, G.; Manjali, J.J.; Prakash, G.; Pal, M.; Joshi, A.; Dholakia, K.; Bhattacharjee, A.; Talole, S.; Puppalwar, A.; et al. Locoregional recurrence after cystectomy in muscle invasive bladder cancer: Implications for adjuvant radiotherapy. Urol. Oncol. 2021, 39, 496.e9–496.e15. [Google Scholar] [CrossRef]
- Mueller, J.; Schrader, A.J.; Schrader, M.; Schnoeller, T.; Jentzmik, F. Management of muscle-invasive bladder cancer. Minerva Urol. Nefrol. 2013, 65, 235–248. [Google Scholar]
- Martini, A.; Sfakianos, J.P.; Renström-Koskela, L.; Mortezavi, A.; Falagario, U.G.; Egevad, L.; Hosseini, A.; Mehrazin, R.; Galsky, M.D.; Steineck, G.; et al. The natural history of untreated muscle-invasive bladder cancer. BJU Int. 2020, 125, 270–275. [Google Scholar] [CrossRef]
- Moghbeli, M.; Abbaszadegan, M.R.; Farshchian, M.; Montazer, M.; Raeisossadati, R.; Abdollahi, A.; Forghanifard, M.M. Association of PYGO2 and EGFR in esophageal squamous cell carcinoma. Med. Oncol. 2013, 30, 516. [Google Scholar] [CrossRef]
- Moghbeli, M.; Makhdoumi, Y.; Delgosha, M.S.; Aarabi, A.; Dadkhah, E.; Memar, B.; Abdollahi, A.; Abbaszadegan, M.R. ErbB1 and ErbB3 co-over expression as a prognostic factor in gastric cancer. Biol. Res. 2019, 52, 2. [Google Scholar] [CrossRef]
- Janes, P.W.; Slape, C.I.; Farnsworth, R.H.; Atapattu, L.; Scott, A.M.; Vail, M.E. EphA3 biology and cancer. Growth Factors 2014, 32, 176–189. [Google Scholar] [CrossRef]
- Boyd, A.W.; Ward, L.D.; Wicks, I.P.; Simpson, R.J.; Salvaris, E.; Wilks, A.; Welch, K.; Loudovaris, M.; Rockman, S.; Busmanis, I. Isolation and characterization of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line. J. Biol. Chem. 1992, 267, 3262–3267. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, Q.; Liu, H.; Ye, M.; Wu, X.; Guo, L. EPHA3 regulates the multidrug resistance of small cell lung cancer via the PI3K/BMX/STAT3 signaling pathway. Tumour Biol. 2016, 37, 11959–11971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, M.; Gallo, E. Critical role of EphA3 in cancer and current state of EphA3 drug therapeutics. Mol. Biol. Rep. 2020, 47, 5523–5533. [Google Scholar] [CrossRef] [PubMed]
- Verschueren, E.; Husain, B.; Yuen, K.; Sun, Y.; Paduchuri, S.; Senbabaoglu, Y.; Lehoux, I.; Arena, T.A.; Wilson, B.; Lianoglou, S.; et al. The Immunoglobulin Superfamily Receptome Defines Cancer-Relevant Networks Associated with Clinical Outcome. Cell 2020, 182, 329–344.e19. [Google Scholar] [CrossRef] [PubMed]
- Samatar, A.A.; Poulikakos, P.I. Targeting RAS-ERK signalling in cancer: Promises and challenges. Nat. Rev. Drug Discov. 2014, 13, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, Y.; Guo, X.B.; Zhao, B. CDC42 Regulates Cell Proliferation and Apoptosis in Bladder Cancer via the IQGAP3-Mediated Ras/ERK Pathway. Biochem. Genet. 2022, 60, 2383–2398. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; Meylan, M.; de Reyniès, A.; Sautès-Fridman, C.; Fridman, W.H. The Tumor Microenvironment in the Response to Immune Checkpoint Blockade Therapies. Front. Immunol. 2020, 11, 784. [Google Scholar] [CrossRef]
- Schneider, A.K.; Chevalier, M.F.; Derré, L. The multifaceted immune regulation of bladder cancer. Nat. Rev. Urol. 2019, 16, 613–630. [Google Scholar] [CrossRef]
- Crispen, P.L.; Kusmartsev, S. Mechanisms of immune evasion in bladder cancer. Cancer Immunol. Immunother. 2020, 69, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.C.; Bondaruk, J.; Yao, H.; Wang, Z.; Zhang, L.; Lee, S.; Lee, J.G.; Cogdell, D.; Zhang, M.; Yang, G.; et al. Assessment of Luminal and Basal Phenotypes in Bladder Cancer. Sci. Rep. 2020, 10, 9743. [Google Scholar] [CrossRef]
- Choi, W.; Porten, S.; Kim, S.; Willis, D.; Plimack, E.R.; Hoffman-Censits, J.; Roth, B.; Cheng, T.; Tran, M.; Lee, I.L.; et al. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell 2014, 25, 152–165. [Google Scholar] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautès-Fridman, C.; Fridman, W.H.; et al. Estimating the population abundance of tissue-infiltrating immune and stromal cell populations using gene expression. Genome Biol. 2016, 17, 218. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, S.; Yuan, R.; Engleman, E.G. Immune Checkpoint Inhibitors for the Treatment of Cancer: Clinical Impact and Mechanisms of Response and Resistance. Annu. Rev. Pathol. 2021, 16, 223–249. [Google Scholar] [CrossRef]
- Degirmenci, U.; Wang, M.; Hu, J. Targeting Aberrant RAS/RAF/MEK/ERK Signaling for Cancer Therapy. Cells 2020, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Lenis, A.T.; Lec, P.M.; Chamie, K.; Mshs, M.D. Bladder Cancer: A Review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef]
- Teoh, J.Y.; Kamat, A.M.; Black, P.; Grivas, P.; Shariat, S.F.; Babjuk, M. Recurrence mechanisms of non-muscle-invasive bladder cancer—A clinical perspective. Nat. Rev. Urol. 2022, 19, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Vlaming, M.; Kiemeney, L.; van der Heijden, A.G. Survival after radical cystectomy: Progressive versus De novo muscle invasive bladder cancer. Cancer Treat. Res. Commun. 2020, 25, 100264. [Google Scholar] [CrossRef] [PubMed]
- Grayson, M. Bladder cancer. Nature 2017, 551, S33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Trinh, Q.D. Diagnosis and staging of bladder cancer. Hematol. Oncol. Clin. N. Am. 2015, 29, 205–218, vii. [Google Scholar] [CrossRef]
- Guidance, N.I. Bladder cancer: Diagnosis and management of bladder cancer: © NICE (2015) Bladder cancer: Diagnosis and management of bladder cancer. BJU Int. 2017, 120, 755–765. [Google Scholar]
- Tan, W.S.; Tan, W.P.; Tan, M.Y.; Khetrapal, P.; Dong, L.; deWinter, P.; Feber, A.; Kelly, J.D. Novel urinary biomarkers for the detection of bladder cancer: A systematic review. Cancer Treat. Rev. 2018, 69, 39–52. [Google Scholar] [CrossRef]
- Anderton, M.; van der Meulen, E.; Blumenthal, M.J.; Schäfer, G. The Role of the Eph Receptor Family in Tumorigenesis. Cancers 2021, 13, 206. [Google Scholar] [CrossRef]
- Pliego Zamora, A.C.; Ranasinghe, H.; Lisle, J.E.; Ng, C.K.; Huang, S.; Wadlow, R.; Scott, A.M.; Boyd, A.W.; Slape, C.I. Cell-Extrinsic Differentiation Block Mediated by EphA3 in Pre-Leukaemic Thymus Contributes to Disease Progression. Cancers 2021, 13, 3858. [Google Scholar] [CrossRef]
- Lisabeth, E.M.; Fernandez, C.; Pasquale, E.B. Cancer somatic mutations disrupt functions of the EphA3 receptor tyrosine kinase through multiple mechanisms. Biochemistry 2012, 51, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Li, F.; Xu, D.; Hou, K.; Fang, W.; Li, Y. The Function of RAS Mutation in Cancer and Advances in its Drug Research. Curr. Pharm. Des. 2019, 25, 1105–1114. [Google Scholar] [CrossRef]
- Bos, J.L. ras oncogenes in human cancer: A review. Cancer Res. 1989, 49, 4682–4689. [Google Scholar] [PubMed]
- Tripathi, K.; Goel, A.; Singhai, A.; Garg, M. Promoter hypomethylation as potential confounder of Ras gene overexpression and their clinical significance in subsets of urothelial carcinoma of bladder. Mol. Biol. Rep. 2021, 48, 2183–2199. [Google Scholar] [CrossRef] [PubMed]
- Reissig, T.M.; Sara, L.; Ting, S.; Reis, H.; Metzenmacher, M.; Eberhardt, W.E.E.; Zaun, G.; Herold, T.; Aigner, C.; Darwiche, K.; et al. ERK phosphorylation as a marker of RAS activity and its prognostic value in non-small cell lung cancer. Lung Cancer 2020, 149, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.K.; Kwak, Y.; Kim, A.; Seok, J.; Kwak, H.J.; Lee, M.; Dayem, A.A.; Song, K.; Park, J.Y.; Park, K.S.; et al. TTYH3 Modulates Bladder Cancer Proliferation and Metastasis via FGFR1/H-Ras/A-Raf/MEK/ERK Pathway. Int. J. Mol. Sci. 2022, 23, 10496. [Google Scholar] [CrossRef]
- Michael, J.V.; Wurtzel, J.G.; Goldfinger, L.E. Regulation of H-Ras-driven MAPK signaling, transformation and tumorigenesis, but not PI3K signaling and tumor progression, by plasma membrane microdomains. Oncogenesis 2016, 5, e228. [Google Scholar] [CrossRef] [Green Version]
- Miyake, M.; Tatsumi, Y.; Gotoh, D.; Ohnishi, S.; Owari, T.; Iida, K.; Ohnishi, K.; Hori, S.; Morizawa, Y.; Itami, Y.; et al. Regulatory T Cells and Tumor-Associated Macrophages in the Tumor Microenvironment in Non-Muscle Invasive Bladder Cancer Treated with Intravesical Bacille Calmette-Guérin: A Long-Term Follow-Up Study of a Japanese Cohort. Int. J. Mol. Sci. 2017, 18, 2186. [Google Scholar] [CrossRef] [Green Version]
- Lang, P.A.; Crome, S.Q.; Xu, H.C.; Lang, K.S.; Chapatte, L.; Deenick, E.K.; Grusdat, M.; Pandyra, A.A.; Pozdeev, V.I.; Wang, R.; et al. NK Cells Regulate CD8(+) T Cell Mediated Autoimmunity. Front. Cell. Infect. Microbiol. 2020, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Yrigoyen, M.; Cassetta, L.; Pollard, J.W. Macrophage targeting in cancer. Ann. N. Y. Acad. Sci. 2021, 1499, 18–41. [Google Scholar] [CrossRef]
- Martínez-Lostao, L.; Anel, A.; Pardo, J. How Do Cytotoxic Lymphocytes Kill Cancer Cells? Clin. Cancer Res. 2015, 21, 5047–5056. [Google Scholar] [CrossRef] [Green Version]
- Mollinedo, F. Neutrophil Degranulation, Plasticity, and Cancer Metastasis. Trends Immunol. 2019, 40, 228–242. [Google Scholar] [CrossRef]
- Liu, S.T.; Hui, G.; Mathis, C.; Chamie, K.; Pantuck, A.J.; Drakaki, A. The Current Status and Future Role of the Phosphoinositide 3 Kinase/AKT Signaling Pathway in Urothelial Cancer: An Old Pathway in the New Immunotherapy Era. Clin. Genitourin. Cancer 2018, 16, e269–e276. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Levels | Low EPHA3 Expression | High EPHA3 Expression | p |
|---|---|---|---|---|
| n | 207 | 207 | ||
| Gender, n (%) | Female | 47 (11.4%) | 62 (15%) | 0.118 |
| Male | 160 (38.6%) | 145 (35%) | ||
| Age, n (%) | ≤70 | 120 (29%) | 114 (27.5%) | 0.620 |
| >70 | 87 (21%) | 93 (22.5%) | ||
| T stage, n (%) | T1 | 2 (0.5%) | 3 (0.8%) | 0.017 |
| T2 | 71 (18.7%) | 48 (12.6%) | ||
| T3 | 84 (22.1%) | 112 (29.5%) | ||
| T4 | 25 (6.6%) | 35 (9.2%) | ||
| N stage, n (%) | N0 | 122 (33%) | 117 (31.6%) | 0.225 |
| N1 | 21 (5.7%) | 25 (6.8%) | ||
| N2 | 29 (7.8%) | 48 (13%) | ||
| N3 | 4 (1.1%) | 4 (1.1%) | ||
| M stage, n (%) | M0 | 111 (52.1%) | 91 (42.7%) | 0.371 |
| M1 | 4 (1.9%) | 7 (3.3%) | ||
| Pathologic stage, n (%) | Stage I | 1 (0.2%) | 3 (0.7%) | <0.001 |
| Stage II | 84 (20.4%) | 46 (11.2%) | ||
| Stage III | having | 77 (18.7%) | ||
| Stage IV | 55 (13.3%) | 81 (19.7%) | ||
| Histologic grade, n (%) | High Grade | 187 (45.5%) | 203 (49.4%) | 0.002 |
| Low Grade | 18 (4.4%) | 3 (0.7%) | ||
| Subtype, n (%) | Non-Papillary | 122 (29.8%) | 153 (37.4%) | 0.001 |
| Papillary | 83 (20.3%) | 51 (12.5%) | ||
| OS event, n (%) | Alive | 132 (31.9%) | 99 (23.9%) | 0.002 |
| Dead | 75 (18.1%) | 108 (26.1%) | ||
| DFS event, n (%) | Alive | 155 (38.8%) | 119 (29.8%) | <0.001 |
| Dead | 48 (12%) | 78 (19.5%) | ||
| Lymphovascular invasion, n (%) | No | 75 (26.5%) | 55 (19.4%) | 0.120 |
| Yes | 73 (25.8%) | 80 (28.3%) | ||
| Age, median (IQR) | 68 (60, 75) | 69 (60.5, 76) | 0.379 |
| Genes | Sequence (5′–3′) |
|---|---|
| EPHA3 forward | ATTTTGGCAATGGGCATTTA |
| EPHA3 reverse | ATGTATGTGGGTCAACATAAGTCC |
| GAPDH forward | CAGGAGGCATTGCTGATGAT |
| GAPDH reverse | GAAGGCTGGGGCTCATTT |
| Primary Antibody | Source Species | Company | Product No. | Predicted Band Size | Dilution |
|---|---|---|---|---|---|
| EPHA3 | Rabbit | ABCAM | ab126261 | 110 kDa | 1:1000 |
| ERK1/2 | Rabbit | CST | #9102 | 4244 kDa | 1:1000 |
| pERK1/2 | Rabbit | CST | #4370 | 4244 kDa | 1:2000 |
| Ras | Rabbit | CST | #67648 | 21 kDa | 1:1000 |
| GAPDH | Rabbit | Beyotime | aF1186 | 36 kDa | 1:2000 |
| Cell Type | Gene Marker | None | Purity | ||
|---|---|---|---|---|---|
| Cor | p | Cor | p | ||
| B cells | CD19 | 0.361 | *** | 0.253 | *** |
| CD20 (KRT20) | 0.032 | 0.524 | 0.094 | 0.072 | |
| CD38 | 0.307 | *** | 0.16 | *** | |
| CD8 + cells | CD8A | 0.213 | *** | 0.063 | 0.231 |
| CD8B | 0.175 | *** | 0.066 | 0.209 | |
| Tfh | BCL6 | 0.074 | 0.134 | 0.093 | 0.0749 |
| ICOS | 0.254 | *** | 0.095 | 0.0686 | |
| CXCR5 | 0.371 | *** | 0.253 | *** | |
| Th1 | T-bet (TBX21) | 0.248 | *** | 0.084 | 0.106 |
| STAT4 | 0.241 | *** | 0.081 | 0.119 | |
| IL12RB2 | 0.107 | * | 0.008 | 0.879 | |
| WSX1 (IL27RA) | 0.164 | *** | 0.045 | 0.394 | |
| STAT1 | 0.133 | ** | 0.014 | 0.793 | |
| IFN-γ (IFNG) | 0.054 | 0.277 | −0.089 | 0.087 | |
| TNF-α (TNF) | 0.084 | 0.0913 | −0.028 | 0.599 | |
| Th2 | GATA3 | −0.058 | 0.24 | 0.029 | 0.585 |
| CCR3 | 0.158 | *** | 0.122 | * | |
| STAT6 | −0.023 | 0.637 | 0.031 | 0.557 | |
| STAT5A | 0.49 | *** | 0.42 | *** | |
| Th9 | TGFBR2 | −0.014 | 0.902 | −0.022 | 0.855 |
| IRF4 | 0.358 | *** | 0.215 | *** | |
| PU.1 (SPI1) | 0.384 | *** | 0.26 | *** | |
| Th17 | STAT3 | 0.244 | *** | 0.164 | ** |
| IL21R | 0.346 | *** | 0.215 | *** | |
| IL23R | 0.168 | *** | 0.127 | * | |
| IL17A | −0.084 | 0.094 | −0.136 | ** | |
| Th22 | CCR10 | 0.226 | *** | 0.211 | *** |
| AHR | 0.067 | 0.177 | 0.139 | ** | |
| Tregs | FOXP3 | 0.409 | *** | 0.319 | *** |
| CD25 (IL2RA) | 0.364 | *** | 0.227 | *** | |
| CCR8 | 0.441 | *** | 0.363 | *** | |
| T cells exhaustion | PD-1 (PDCD1) | 0.234 | *** | 0.071 | 0.176 |
| CTLA4 | 0.237 | *** | 0.079 | 0.129 | |
| LAG3 | 0.171 | *** | 0.014 | 0.718 | |
| TIM-3 (HAVCR2) | 0.345 | *** | 0.206 | *** | |
| Macrophages | CD68 | 0.177 | *** | 0.06 | 0.25 |
| CD11b (ITGAM) | 0.376 | *** | 0.255 | *** | |
| M1 | INOS (NOS2) | 0.225 | *** | 0.182 | *** |
| IRF5 | 0.072 | 0.141 | 0.074 | 0.155 | |
| COX2 (PTGS2) | 0.172 | *** | 0.122 | * | |
| M2 | CD163 | 0.42 | *** | 0.314 | *** |
| ARG1 | 0.136 | ** | 0.164 | ** | |
| MRC1 | 0.404 | *** | 0.304 | *** | |
| MS4A4A | 0.453 | *** | 0.356 | *** | |
| TAMs | CCL2 | 0.445 | *** | 0.329 | *** |
| CD80 | 0.257 | *** | 0.121 | * | |
| CD86 | 0.355 | *** | 0.229 | *** | |
| CCR5 | 0.323 | *** | 0.15 | ** | |
| Monocytes | CD14 | 0.333 | *** | 0.192 | *** |
| CD16(FCGR3B) | 0.193 | *** | 0.109 | ** | |
| CD115 (CSF1R) | 0.415 | *** | 0.312 | *** | |
| Neutrophils | CD66b (CEACAM8) | 0.047 | 0.341 | 0.032 | 0.539 |
| CD15(FUT4) | 0.284 | *** | 0.184 | *** | |
| CD11b (ITGAM) | 0.376 | *** | 0.255 | *** | |
| NK cells | XCL1 | 0.055 | 0.264 | 0.075 | 0.15 |
| CD7 | 0.208 | *** | 0.034 | 0.501 | |
| KIR3DL1 | 0.128 | ** | 0.044 | 0.397 | |
| DCs | CD1C(BDCA-1) | 0.324 | *** | 0.242 | *** |
| CD141(THBD) | 0.093 | 0.0601 | 0.037 | 0.479 | |
| CD11c (ITGAX) | 0.406 | *** | 0.285 | *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhou, Z.; Jiang, Y.; Lin, Y.; Yang, Y.; Tian, C.; Liu, J.; Lin, H.; Huang, B. EPHA3 Could Be a Novel Prognosis Biomarker and Correlates with Immune Infiltrates in Bladder Cancer. Cancers 2023, 15, 621. https://doi.org/10.3390/cancers15030621
Liu J, Zhou Z, Jiang Y, Lin Y, Yang Y, Tian C, Liu J, Lin H, Huang B. EPHA3 Could Be a Novel Prognosis Biomarker and Correlates with Immune Infiltrates in Bladder Cancer. Cancers. 2023; 15(3):621. https://doi.org/10.3390/cancers15030621
Chicago/Turabian StyleLiu, Junpeng, Zewen Zhou, Yifan Jiang, Yuzhao Lin, Yunzhi Yang, Chongjiang Tian, Jinwen Liu, Hao Lin, and Bin Huang. 2023. "EPHA3 Could Be a Novel Prognosis Biomarker and Correlates with Immune Infiltrates in Bladder Cancer" Cancers 15, no. 3: 621. https://doi.org/10.3390/cancers15030621
APA StyleLiu, J., Zhou, Z., Jiang, Y., Lin, Y., Yang, Y., Tian, C., Liu, J., Lin, H., & Huang, B. (2023). EPHA3 Could Be a Novel Prognosis Biomarker and Correlates with Immune Infiltrates in Bladder Cancer. Cancers, 15(3), 621. https://doi.org/10.3390/cancers15030621