
Targeted Degradation of Androgen Receptor by VNPP433-3β in Castration-Resistant Prostate Cancer Cells Implicates Interaction with E3 Ligase MDM2 Resulting in Ubiquitin-Proteasomal Degradation

 and
and 
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Immunoblotting
2.3. Preparation of Nuclear and Cytoplasmic Fractions
2.4. Molecular Docking
2.5. Co-Immunoprecipitation (Co-IP) Assay
2.6. Real Time Quantitative Studies of AR Degradation in CRISPR Knock-In HiBiT Reporter Cell Line
2.7. RNA-Sequencing and Transcriptome Analyses
2.8. Statistical Analysis
3. Results
3.1. VNPP433-3β Induces Targeted Degradation of AR/AR-V7
3.2. Degradation of f-AR by VNPP433-3β Leads to Its Decreased Nuclear Translocation
3.3. Decrease in f-AR/AR-V7 by VNPP433-3β Is Due to Enhanced Proteasomal Degradation
3.4. Real Time Quantitative Assay of VNPP433-3β-Induced AR Degradation in Live CWR22Rv1 Cells
3.5. VNPP433-3β Acts as Molecular Glue between AR and MDM2
3.6. RNA Sequencing Shows Transcriptional Downregulation of Several AR-Target Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mullard, A. Targeted protein degraders crowd into the clinic. Nat. Rev. Drug Discov. 2021, 20, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Stanton, B.Z.; Chory, E.J.; Crabtree, G.R. Chemically induced proximity in biology and medicine. Science 2018, 359, eaao5902. [Google Scholar] [CrossRef]
- Gerry, C.J.; Schreiber, S.L. Unifying principles of bifunctional, proximity-inducing small molecules. Nat. Chem. Biol. 2020, 16, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Ding, Y.; He, S.; Sheng, C. Molecular Glues for Targeted Protein Degradation: From Serendipity to Rational Discovery. J. Med. Chem. 2021, 64, 10606–10620. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Avgeris, I.; Pliatsika, D.; Nikolaropoulos, S.S.; Fousteris, M.A. Targeting androgen receptor for prostate cancer therapy: From small molecules to PROTACs. Bioorg. Chem. 2022, 128, 106089. [Google Scholar] [CrossRef]
- Beretta, G.L.; Zaffaroni, N. Androgen Receptor-Directed Molecular Conjugates for Targeting Prostate Cancer. Front. Chem. 2019, 7, 369. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Han, X. Targeting androgen receptor degradation with PROTACs from bench to bedside. Biomed. Pharmacother. 2023, 158, 114112. [Google Scholar] [CrossRef]
- Mohler, M.L.; Sikdar, A.; Ponnusamy, S.; Hwang, D.J.; He, Y.; Miller, D.D.; Narayanan, R. An Overview of Next-Generation Androgen Receptor-Targeted Therapeutics in Development for the Treatment of Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 2124. [Google Scholar] [CrossRef]
- Xiang, W.; Wang, S. Therapeutic Strategies to Target the Androgen Receptor. J. Med. Chem. 2022, 65, 8772–8797. [Google Scholar] [CrossRef]
- Kwegyir-Afful, A.K.; Ramalingam, S.; Ramamurthy, V.P.; Purushottamachar, P.; Murigi, F.N.; Vasaitis, T.S.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.; et al. Galeterone and The Next Generation Galeterone Analogs, VNPP414 and VNPP433-3beta Exert Potent Therapeutic Effects in Castration-/Drug-Resistant Prostate Cancer Preclinical Models In Vitro and In Vivo. Cancers 2019, 11, 1637. [Google Scholar] [CrossRef] [PubMed]
- Purushottamachar, P.; Thomas, E.; Thankan, R.S.; Rudchenko, V.; Huang, G.; Njar, V.C.O. Large-scale synthesis of galeterone and lead next generation galeterone analog VNPP433-3beta. Steroids 2022, 185, 109062. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.P.; Weber, D.J.; Njar, V.C.O. Novel AR/AR-V7 and Mnk1/2 Degrader, VNPP433-3beta: Molecular Mechanisms of Action and Efficacy in AR-Overexpressing Castration Resistant Prostate Cancer In Vitro and In Vivo Models. Cells 2022, 11, 2699. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.; Weber, D.J.; Njar, V.C.O. Transcriptome profiling reveals that VNPP433-3beta, the lead next-generation galeterone analog inhibits prostate cancer stem cells by downregulating epithelial-mesenchymal transition and stem cell markers. Mol. Carcinog. 2022, 61, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Sharp, A.; Coleman, I.; Yuan, W.; Sprenger, C.; Dolling, D.; Rodrigues, D.N.; Russo, J.W.; Figueiredo, I.; Bertan, C.; Seed, G.; et al. Androgen receptor splice variant-7 expression emerges with castration resistance in prostate cancer. J. Clin. Investig. 2019, 129, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Gopalakrishnan, V.; Somasagara, R.R.; Choudhary, B.; Raghavan, S.C. Extract of Vernonia condensata, Inhibits Tumor Progression and Improves Survival of Tumor-allograft Bearing Mouse. Sci. Rep. 2016, 6, 23255. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Cai, C.; Gao, S.; Simon, N.I.; Shen, H.C.; Balk, S.P. Galeterone prevents androgen receptor binding to chromatin and enhances degradation of mutant androgen receptor. Clin. Cancer Res. 2014, 20, 4075–4085. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Gopalakrishnan, V.; Hegde, M.; Kumar, S.; Karki, S.S.; Raghavan, S.C.; Choudhary, B. A Novel Resveratrol Based Tubulin Inhibitor Induces Mitotic Arrest and Activates Apoptosis in Cancer Cells. Sci. Rep. 2016, 6, 34653. [Google Scholar] [CrossRef]
- Riching, K.M.; Mahan, S.; Corona, C.R.; McDougall, M.; Vasta, J.D.; Robers, M.B.; Urh, M.; Daniels, D.L. Quantitative Live-Cell Kinetic Degradation and Mechanistic Profiling of PROTAC Mode of Action. ACS Chem. Biol. 2018, 13, 2758–2770. [Google Scholar] [CrossRef]
- D’Abronzo, L.S.; Bose, S.; Crapuchettes, M.E.; Beggs, R.E.; Vinall, R.L.; Tepper, C.G.; Siddiqui, S.; Mudryj, M.; Melgoza, F.U.; Durbin-Johnson, B.P.; et al. The androgen receptor is a negative regulator of eIF4E phosphorylation at S209: Implications for the use of mTOR inhibitors in advanced prostate cancer. Oncogene 2017, 36, 6359–6373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoodi, K.Z.; Xu, Y.; Dar, J.A.; Eisermann, K.; Pascal, L.E.; Parrinello, E.; Ai, J.; Johnston, P.A.; Nelson, J.B.; Wipf, P.; et al. Inhibition of Androgen Receptor Nuclear Localization and Castration-Resistant Prostate Tumor Growth by Pyrroloimidazole-based Small Molecules. Mol. Cancer Ther. 2017, 16, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Fan, L.; Hussain, A. Implications of ubiquitin ligases in castration-resistant prostate cancer. Curr. Opin. Oncol. 2015, 27, 172–176. [Google Scholar] [CrossRef]
- Schwinn, M.K.; Machleidt, T.; Zimmerman, K.; Eggers, C.T.; Dixon, A.S.; Hurst, R.; Hall, M.P.; Encell, L.P.; Binkowski, B.F.; Wood, K.V. CRISPR-Mediated Tagging of Endogenous Proteins with a Luminescent Peptide. ACS Chem. Biol. 2018, 13, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.K.; Wang, L.; Hu, Y.C.; Altuwaijri, S.; Chang, C. Phosphorylation-dependent ubiquitylation and degradation of androgen receptor by Akt require Mdm2 E3 ligase. EMBO J. 2002, 21, 4037–4048. [Google Scholar] [CrossRef]
- Gaughan, L.; Logan, I.R.; Neal, D.E.; Robson, C.N. Regulation of androgen receptor and histone deacetylase 1 by Mdm2-mediated ubiquitylation. Nucleic Acids Res. 2005, 33, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Vummidi Giridhar, P.; Williams, K.; VonHandorf, A.P.; Deford, P.L.; Kasper, S. Constant Degradation of the Androgen Receptor by MDM2 Conserves Prostate Cancer Stem Cell Integrity. Cancer Res. 2019, 79, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Fasan, R.; Dias, R.L.; Moehle, K.; Zerbe, O.; Obrecht, D.; Mittl, P.R.; Grutter, M.G.; Robinson, J.A. Structure-activity studies in a family of beta-hairpin protein epitope mimetic inhibitors of the p53-HDM2 protein-protein interaction. Chembiochem 2006, 7, 515–526. [Google Scholar] [CrossRef]
- Wu, D.; Terrian, D.M. Regulation of caveolin-1 expression and secretion by a protein kinase cepsilon signaling pathway in human prostate cancer cells. J. Biol. Chem. 2002, 277, 40449–40455. [Google Scholar] [CrossRef]
- Jin, H.J.; Kim, J.; Yu, J. Androgen receptor genomic regulation. Transl. Androl. Urol. 2013, 2, 157–177. [Google Scholar] [CrossRef]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Njar, V.C. Mechanistic insights on the effects of the lead next generation galeterone analog, VNPP433-3β in castration resistant prostate cancer. Molecular Cancer Therapeutics 20. Mechanistic insights on the effects of the lead next generation galeterone analog, VNPP433-3β in castration resistant prostate cancer. Mol. Cancer Ther. 2021, 20, LBA027. [Google Scholar]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Guo, J.; Parise, R.A.; Beumer, J.H.; Njar, V.C.O. Murine Toxicology and Pharmacokinetics of Lead Next Generation Galeterone Analog, VNPP433-3beta. Steroids 2023, 192, 109184. [Google Scholar] [CrossRef] [PubMed]
- Cato, L.; de Tribolet-Hardy, J.; Lee, I.; Rottenberg, J.T.; Coleman, I.; Melchers, D.; Houtman, R.; Xiao, T.; Li, W.; Uo, T.; et al. ARv7 Represses Tumor-Suppressor Genes in Castration-Resistant Prostate Cancer. Cancer Cell 2019, 35, 401–413.e6. [Google Scholar] [CrossRef] [PubMed]
- Lilja, H.; Ulmert, D.; Vickers, A.J. Prostate-specific antigen and prostate cancer: Prediction, detection and monitoring. Nat. Rev. Cancer 2008, 8, 268–278. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
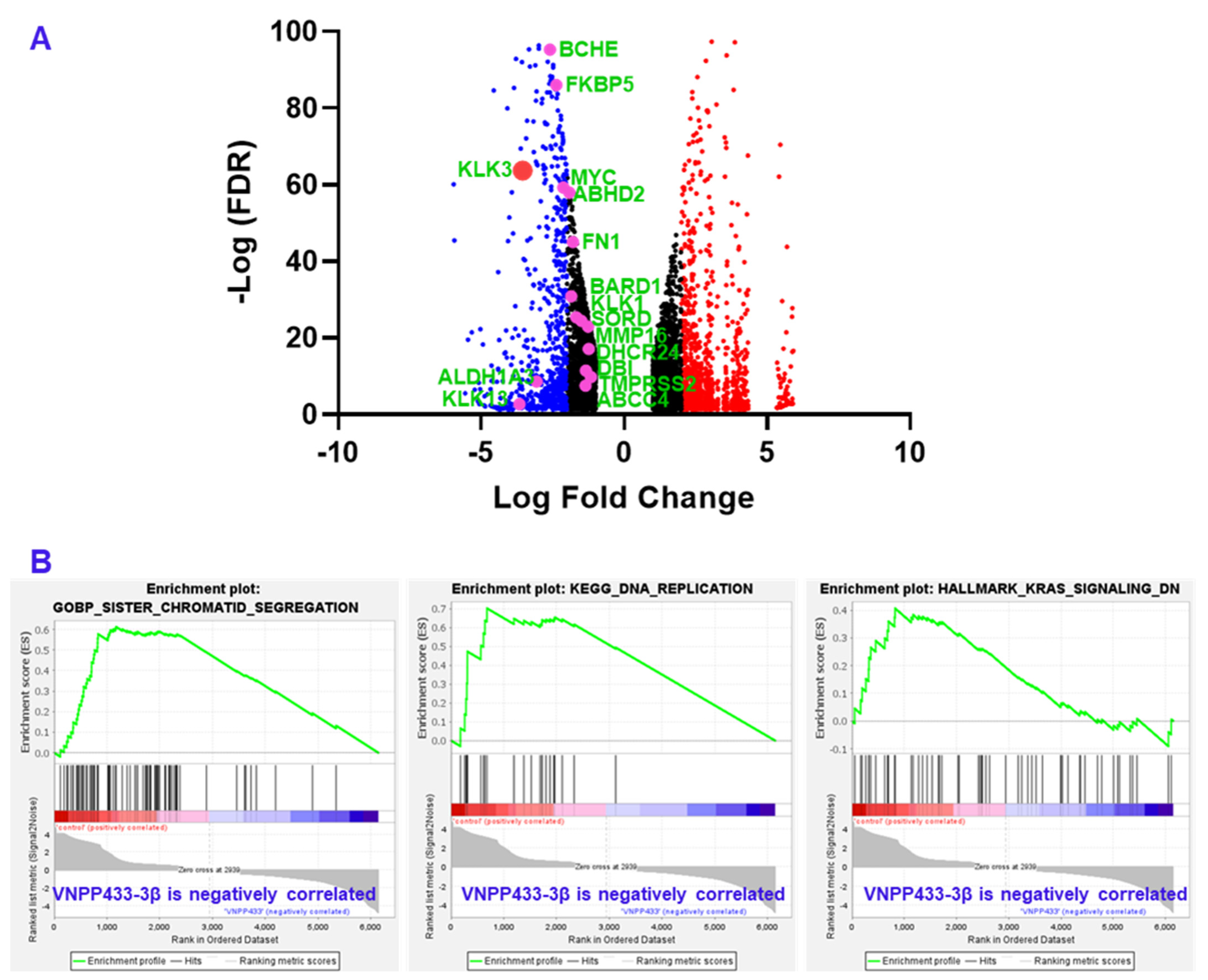
| Gene | LFC | −Log (FDR) | Remarks |
|---|---|---|---|
| KLK3 | −3.549878878 | 63.74105 | Prostate specific antigen (PSA) |
| KLK1 | −1.499349751 | 24.47005 | Serine protease; Oncoprotein |
| KLK13 | −3.661495531 | 2.685301 | Serine protease; Oncoprotein |
| NKX3-1 | −2.987220702 | 106.1704 | Transcription factor in prostate epithelium, tumor suppressor |
| DHCR24 | −1.242758268 | 17.15381 | Flavin adenine dinucleotide-dependent oxidoreductase; cholesterol biosynthesis |
| DBI | −1.340163994 | 11.44401 | acyl-CoA binding protein, cell signaling, steroid synthesis |
| MMP16 | −1.264127518 | 22.95686 | Cancer metastasis, matrix metalloproteinase |
| FKBP5 | −2.367045284 | 86.07973 | Co-chaperone that regulates steroid hormone receptors, immunoregulation |
| ALDH1A3 | −3.05609345 | 8.603198 | Tumorigenesis, detoxification of aldehydes and lipid peroxidation |
| BCHE | −2.590390622 | 95.29679 | Nonspecific cholinesterase |
| ABHD2 | −1.915809808 | 57.9697 | Enables nuclear steroid receptor activity, highly expressed in spermatozoa |
| SORD | −1.673855387 | 25.39808 | Carbohydrate metabolism |
| ABCC4 | −1.351567948 | 7.512045 | Multidrug resistance, regulates intracellular yclic nucleotide levels, cAMP-dependent signaling to the nucleus |
| BARD1 | −1.850384403 | 30.88807 | DNA repair, E3 ubiquitin ligase, transcriptional regulation to maintain genomic stability |
| TMPRSS2 | −1.160624783 | 9.802972 | Transmembrane Serine Protease 2, upregulated by androgenic hormones in prostate cancer, facilitates virus entry |
| FN1 | −1.795774135 | 45.01869 | Component of extracellular matrix, upregulated in multiple cancers |
| MYC | −2.129324177 | 59.28253 | Oncoprotein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Weber, D.J.; Njar, V.C.O. Targeted Degradation of Androgen Receptor by VNPP433-3β in Castration-Resistant Prostate Cancer Cells Implicates Interaction with E3 Ligase MDM2 Resulting in Ubiquitin-Proteasomal Degradation. Cancers 2023, 15, 1198. https://doi.org/10.3390/cancers15041198
Thomas E, Thankan RS, Purushottamachar P, Weber DJ, Njar VCO. Targeted Degradation of Androgen Receptor by VNPP433-3β in Castration-Resistant Prostate Cancer Cells Implicates Interaction with E3 Ligase MDM2 Resulting in Ubiquitin-Proteasomal Degradation. Cancers. 2023; 15(4):1198. https://doi.org/10.3390/cancers15041198
Chicago/Turabian StyleThomas, Elizabeth, Retheesh S. Thankan, Puranik Purushottamachar, David J. Weber, and Vincent C. O. Njar. 2023. "Targeted Degradation of Androgen Receptor by VNPP433-3β in Castration-Resistant Prostate Cancer Cells Implicates Interaction with E3 Ligase MDM2 Resulting in Ubiquitin-Proteasomal Degradation" Cancers 15, no. 4: 1198. https://doi.org/10.3390/cancers15041198
APA StyleThomas, E., Thankan, R. S., Purushottamachar, P., Weber, D. J., & Njar, V. C. O. (2023). Targeted Degradation of Androgen Receptor by VNPP433-3β in Castration-Resistant Prostate Cancer Cells Implicates Interaction with E3 Ligase MDM2 Resulting in Ubiquitin-Proteasomal Degradation. Cancers, 15(4), 1198. https://doi.org/10.3390/cancers15041198