
PKCθ Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-κB/IL-1β-Dependent Manner

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis of PA
2.2. Clinical Samples
2.3. Animal Model of Bone-Invasive PAs
2.4. Tissue Sections Stained
2.5. Microcomputed Tomography (µCT) Analysis
2.6. Cells Treatment and Transfection
2.7. Cell Viability Assays
2.8. Ki-67
2.9. Wound Healing Assay
2.10. Transwell Assay
2.11. Osteoclast Differentiation Induction
2.12. Quantitative PCR (qPCR)
2.13. Western Blotting
2.14. Coimmunoprecipitation (co-IP)
2.15. Enzyme-Linked Immunosorbent Assay (ELISA)
2.16. Statistical Analysis
3. Results
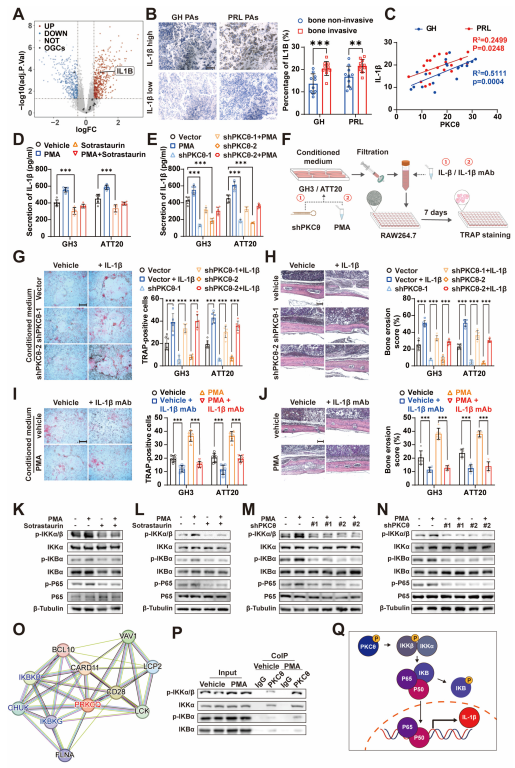
3.1. Bone Invasion of PA Is Associated with Proinflammatory Signals
3.2. PKCθ Is Potentially the Key Factor Related to Inflammation Status of PAs
3.3. PKCθ Is Involved in the Proliferation, Migration, and Bone Invasion of Pituitary Tumors
3.4. PKCθ of Pituitary Adenoma Affects Tumor Bone Invasion by Regulating Monocyte-osteoclast Differentiation
3.5. PKCθ of Pituitary Adenoma Promotes Monocyte–Osteoclast Differentiation by Regulating IL-1β Expression and Release
3.6. Celastrol Alleviates the Bone Invasion of PAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Melmed, S. Pathogenesis of pituitary tumors. Nat. Rev. Endocrinol. 2011, 7, 257–266. [Google Scholar] [CrossRef]
- Molitch, M.E. Diagnosis and Treatment of Pituitary Adenomas: A Review. JAMA 2017, 317, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Miermeister, C.P.; Petersenn, S.; Buchfelder, M.; Fahlbusch, R.; Ludecke, D.K.; Holsken, A.; Bergmann, M.; Knappe, H.U.; Hans, V.H.; Flitsch, J.; et al. Histological criteria for atypical pituitary adenomas-data from the German pituitary adenoma registry suggests modifications. Acta Neuropathol. Commun. 2015, 3, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Li, S.; Zhang, W.; Zang, Z.; Hu, J.; Yang, H. Current biomarkers of invasive sporadic pituitary adenomas. Ann. Endocrinol. 2016, 77, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Guo, J.; Shen, Y.; Dong, W.; Gao, H.; Miao, Y.; Li, C.; Zhang, Y. Functions and Mechanisms of Tumor Necrosis Factor-alpha and Noncoding RNAs in Bone-Invasive Pituitary Adenomas. Clin. Cancer Res. 2018, 24, 5757–5766. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Li, B.; Li, C.; Liu, C.; Wang, X.; Gui, S.; Zhao, P.; Bai, J.; Cao, L.; Zhang, Y. The clinical features, recurrence risks and surgical strategies of bone invasive pituitary adenomas. Clin. Neurol. Neurosurg. 2021, 201, 106455. [Google Scholar] [CrossRef]
- Nomikos, P.; Buchfelder, M.; Fahlbusch, R. The outcome of surgery in 668 patients with acromegaly using current criteria of biochemical 'cure'. Eur. J. Endocrinol. 2005, 152, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Cheng, J.; Wang, H.; Zhao, S.; Zhu, H.; Li, C.; Zhang, Y.; Zhao, P. CCNB1 affects cavernous sinus invasion in pituitary adenomas through the epithelial-mesenchymal transition. J. Transl. Med. 2019, 17, 336. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, H.; Ji, Z.; Cheng, Y.; Du, Y.; Liu, H.; Liao, J.; Peng, H. RASSF10 regulates bone invasion of growth hormone-secreting adenomas via exosomes. Biochem. Biophys. Res. Commun. 2020, 527, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Zhu, H.B.; Li, B.; Guo, J.; Miao, Y.Z.; Shen, Y.T.; Zhang, Y.Z.; Zhao, P.; Li, C.Z. LncRNA MEG8 promotes TNF-alpha expression by sponging miR-454-3p in bone-invasive pituitary adenomas. Aging 2021, 13, 14342–14354. [Google Scholar] [CrossRef] [PubMed]
- Isakov, N. Protein kinase C (PKC) isoforms in cancer, tumor promotion and tumor suppression. Semin. Cancer Biol. 2018, 48, 36–52. [Google Scholar] [CrossRef]
- Lei, T.; Adams, E.F.; Buchfelder, M.; Fahlbusch, R. Relationship between protein kinase C and adenylyl cyclase activity in the regulation of growth hormone secretion by human pituitary somatotrophinomas. Neurosurgery 1996, 39, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Ye, F.; Xu, T.; Wang, S.; Wang, X.; Wang, H.; Wan, F.; Lei, T. GHRP-6 induces CREB phosphorylation and growth hormone secretion via a protein kinase Csigma-dependent pathway in GH3 cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 30, 183–187. [Google Scholar] [CrossRef]
- Lei, Z.; Wang, J.; Sun, W.; Chen, X.; Jiao, W.; Zhang, H.; Lei, T.; Li, F. PKCdelta reveals a tumor promoter function by promoting cell proliferation and migration in somatotropinomas. Int. J. Clin. Exp. Pathol. 2018, 11, 208–215. [Google Scholar] [PubMed]
- Alvaro, V.; Levy, L.; Dubray, C.; Roche, A.; Peillon, F.; Querat, B.; Joubert, D. Invasive human pituitary tumors express a point-mutated alpha-protein kinase-C. J. Clin. Endocrinol. Metab. 1993, 77, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Xue, D.; Adams, E.F.; Buchfelder, M.; Fahlbusch, R. Relationship between invasiveness of pituitary somatotrophinomas and structural abnormalities of protein kinase C gene in human. J. Tongji Med. Univ. 1997, 17, 68–71. [Google Scholar] [CrossRef]
- Schiemann, U.; Assert, R.; Moskopp, D.; Gellner, R.; Hengst, K.; Gullotta, F.; Domschke, W.; Pfeiffer, A. Analysis of a protein kinase C-alpha mutation in human pituitary tumours. J. Endocrinol. 1997, 153, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Nicolle, A.; Zhang, Y.; Belguise, K. The Emerging Function of PKCtheta in Cancer. Biomolecules 2021, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Yun, H.; Kim, S.; Lee, N.; Kim, H.; Rho, J. Regulation of Osteoclast Differentiation by Cytokine Networks. Immune Netw. 2018, 18, e8. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Morrison, N.A.; Johnson, N.W.; Gao, J. MCP-1 as a potential target to inhibit the bone invasion by oral squamous cell carcinoma. J. Cell. Biochem. 2014, 115, 1787–1798. [Google Scholar] [CrossRef]
- Zanin-Zhorov, A.; Ding, Y.; Kumari, S.; Attur, M.; Hippen, K.L.; Brown, M.; Blazar, B.R.; Abramson, S.B.; Lafaille, J.J.; Dustin, M.L. Protein kinase C-theta mediates negative feedback on regulatory T cell function. Science 2010, 328, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Dienz, O.; Hehner, S.P.; Droge, W.; Schmitz, M.L. Synergistic activation of NF-kappa B by functional cooperation between vav and PKCtheta in T lymphocytes. J. Biol. Chem. 2000, 275, 24547–24551. [Google Scholar] [CrossRef] [Green Version]
- Coudronniere, N.; Villalba, M.; Englund, N.; Altman, A. NF-kappa B activation induced by T cell receptor/CD28 costimulation is mediated by protein kinase C-theta. Proc. Natl. Acad. Sci. USA 2000, 97, 3394–3399. [Google Scholar] [CrossRef] [PubMed]
- You, D.; Jeong, Y.; Yoon, S.Y.; Kim, S.A.; Kim, S.W.; Nam, S.J.; Lee, J.E.; Kim, S. Celastrol attenuates the inflammatory response by inhibiting IL-1beta expression in triple-negative breast cancer cells. Oncol. Rep. 2021, 45, 89. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Fu, R.J.; Zhang, S.; Yue, S.J.; Chen, Y.Y.; Xu, D.Q.; Tang, Y.P. Potential medicinal value of celastrol and its synthesized analogues for central nervous system diseases. Biomed. Pharmacother. 2021, 139, 111551. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, Y.; Luo, W.; Chen, S.; Lin, F.; Zhang, X.; Fan, S.; Shen, X.; Wang, Y.; Liang, G. Celastrol induces ROS-mediated apoptosis via directly targeting peroxiredoxin-2 in gastric cancer cells. Theranostics 2020, 10, 10290–10308. [Google Scholar] [CrossRef] [PubMed]
- Gan, K.; Xu, L.; Feng, X.; Zhang, Q.; Wang, F.; Zhang, M.; Tan, W. Celastrol attenuates bone erosion in collagen-Induced arthritis mice and inhibits osteoclast differentiation and function in RANKL-induced RAW264.7. Int. Immunopharmacol. 2015, 24, 239–246. [Google Scholar] [CrossRef]
- McHugh, K.P.; Shen, Z.; Crotti, T.N.; Flannery, M.R.; O'Sullivan, R.P.; Purdue, P.E.; Goldring, S.R. The role of cell-substrate interaction in regulating osteoclast activation: Potential implications in targeting bone loss in rheumatoid arthritis. Ann. Rheum. Dis. 2010, 69 (Suppl. 1), i83–i85. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, Q.; Chen, Q. The cytokine network involved in the host immune response to periodontitis. Int. J. Oral Sci. 2019, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, A.; Wu, F.; Hardy, K.; Li, J.; Tu, W.J.; McCuaig, R.; Harris, J.; Khanna, K.K.; Attema, J.; Gregory, P.A.; et al. Chromatinized protein kinase C-theta directly regulates inducible genes in epithelial to mesenchymal transition and breast cancer stem cells. Mol. Cell. Biol. 2014, 34, 2961–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaya, Z.E.; Grebennikova, T.A.; Melnichenko, G.A.; Nikitin, A.G.; Solodovnikov, A.G.; Brovkina, O.I.; Grigoriev, A.U.; Rozhinskaya, L.Y.; Dedov, I.I. Effects of endogenous hypercortisolism on bone mRNA and microRNA expression in humans. Osteoporos. Int. 2018, 29, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.H.; Dudics, S.; Astry, B.; Moudgil, K.D. Control of autoimmune inflammation by celastrol, a natural triterpenoid. Pathog. Dis. 2016, 74, ftw059. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | Overall (n = 40) | BIPAs (n = 20) | Non-BIPAs (n = 20) | p-Value | |
|---|---|---|---|---|---|
| Subtype (%) | GH | 20 (50.00) | 10 (50.00) | 10 (50.00) | 1 * |
| PRL | 20 (50.00) | 10 (50.00) | 10 (50.00) | ||
| Age (median [IQR]) | 46.000 [39.000, 53.250] | 48.500 [42.000, 55.000] | 45.000 [34.000, 48.500] | 0.0784 ** | |
| Gander (%) | Female | 21 (52.50) | 11 (55.00) | 10 (50.00) | 1 * |
| Male | 19 (47.50) | 9 (45.00) | 10 (50.00) | ||
| Knosp grade (%) | <Ⅲ | 18 (45.00) | 4 (20.00) | 14 (70.00) | 0.0036 *** |
| ≥Ⅲ | 22 (55.00) | 16 (80.00) | 6 (30.00) | ||
| Diameter (%) | <1 cm | 20 (50.00) | 5 (25.00) | 15 (75.00) | 0.0044 * |
| ≥1 cm | 20 (50.00) | 15 (75.00) | 5 (25.00) | ||
| Ki-67 (%) | <3% | 19 (47.50) | 5 (25.00) | 14 (70.00) | 0.0113 * |
| ≥3% | 21 (52.50) | 15 (75.00) | 6 (30.00) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Lei, Z.; Wang, Z.; Jiang, Q.; Zhang, Z.; Liu, X.; Xing, B.; Li, S.; Guo, X.; Liu, Y.; et al. PKCθ Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-κB/IL-1β-Dependent Manner. Cancers 2023, 15, 1624. https://doi.org/10.3390/cancers15051624
Wang Q, Lei Z, Wang Z, Jiang Q, Zhang Z, Liu X, Xing B, Li S, Guo X, Liu Y, et al. PKCθ Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-κB/IL-1β-Dependent Manner. Cancers. 2023; 15(5):1624. https://doi.org/10.3390/cancers15051624
Chicago/Turabian StyleWang, Quanji, Zhuowei Lei, Zihan Wang, Qian Jiang, Zhuo Zhang, Xiaojin Liu, Biao Xing, Sihan Li, Xiang Guo, Yanchao Liu, and et al. 2023. "PKCθ Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-κB/IL-1β-Dependent Manner" Cancers 15, no. 5: 1624. https://doi.org/10.3390/cancers15051624
APA StyleWang, Q., Lei, Z., Wang, Z., Jiang, Q., Zhang, Z., Liu, X., Xing, B., Li, S., Guo, X., Liu, Y., Li, X., Shu, K., Zhang, H., Huang, Y., & Lei, T. (2023). PKCθ Regulates Pituitary Adenoma Bone Invasion by Activating Osteoclast in NF-κB/IL-1β-Dependent Manner. Cancers, 15(5), 1624. https://doi.org/10.3390/cancers15051624