
Prospects of POLD1 in Human Cancers: A Review

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
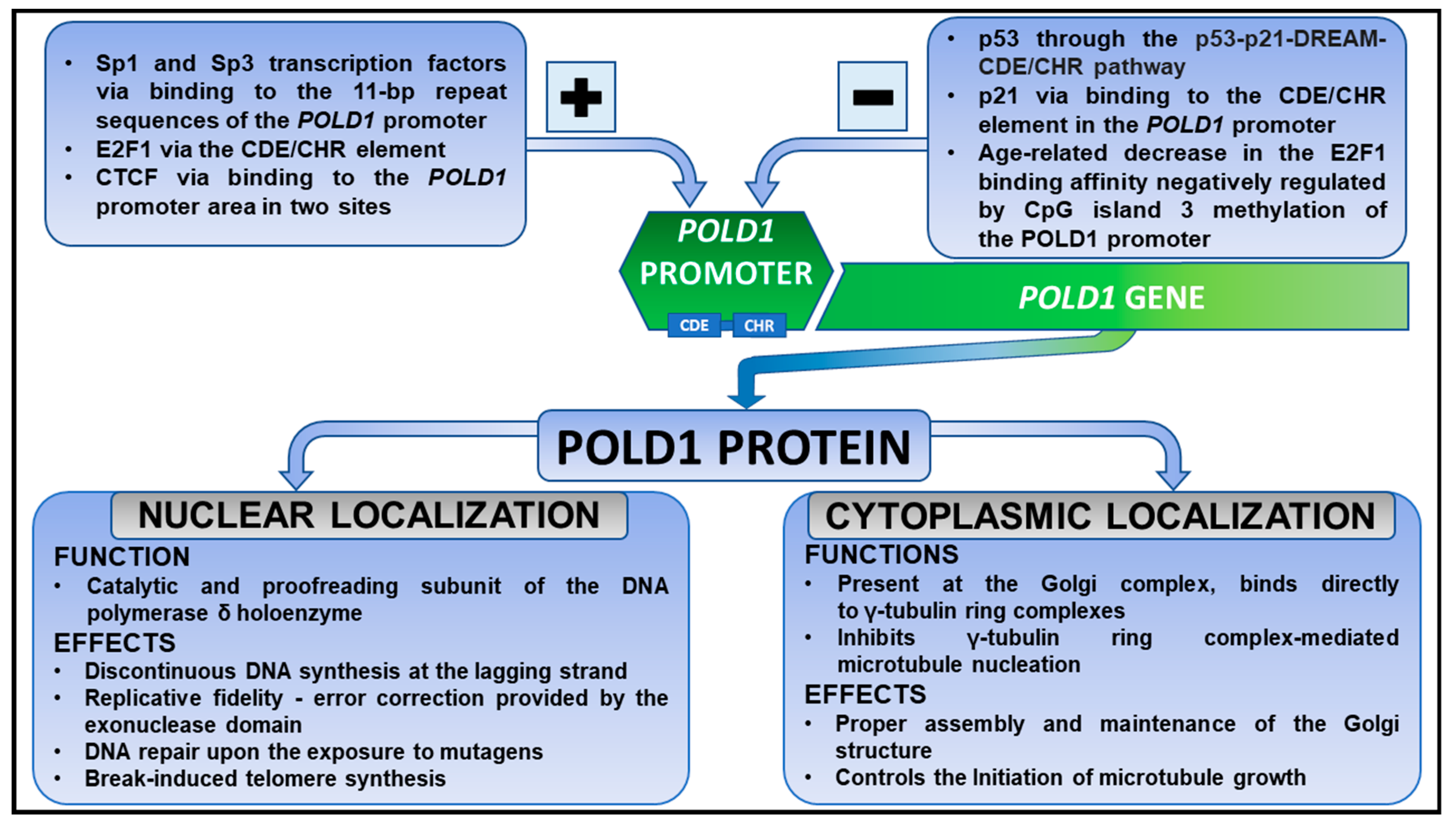
2. POLD1 Gene and a Polymerase Delta Holoenzyme in Health
3. POLD1 Gene and a Polymerase Delta Holoenzyme in Non-Oncogenic Processes
4. POLD1 in Cancers
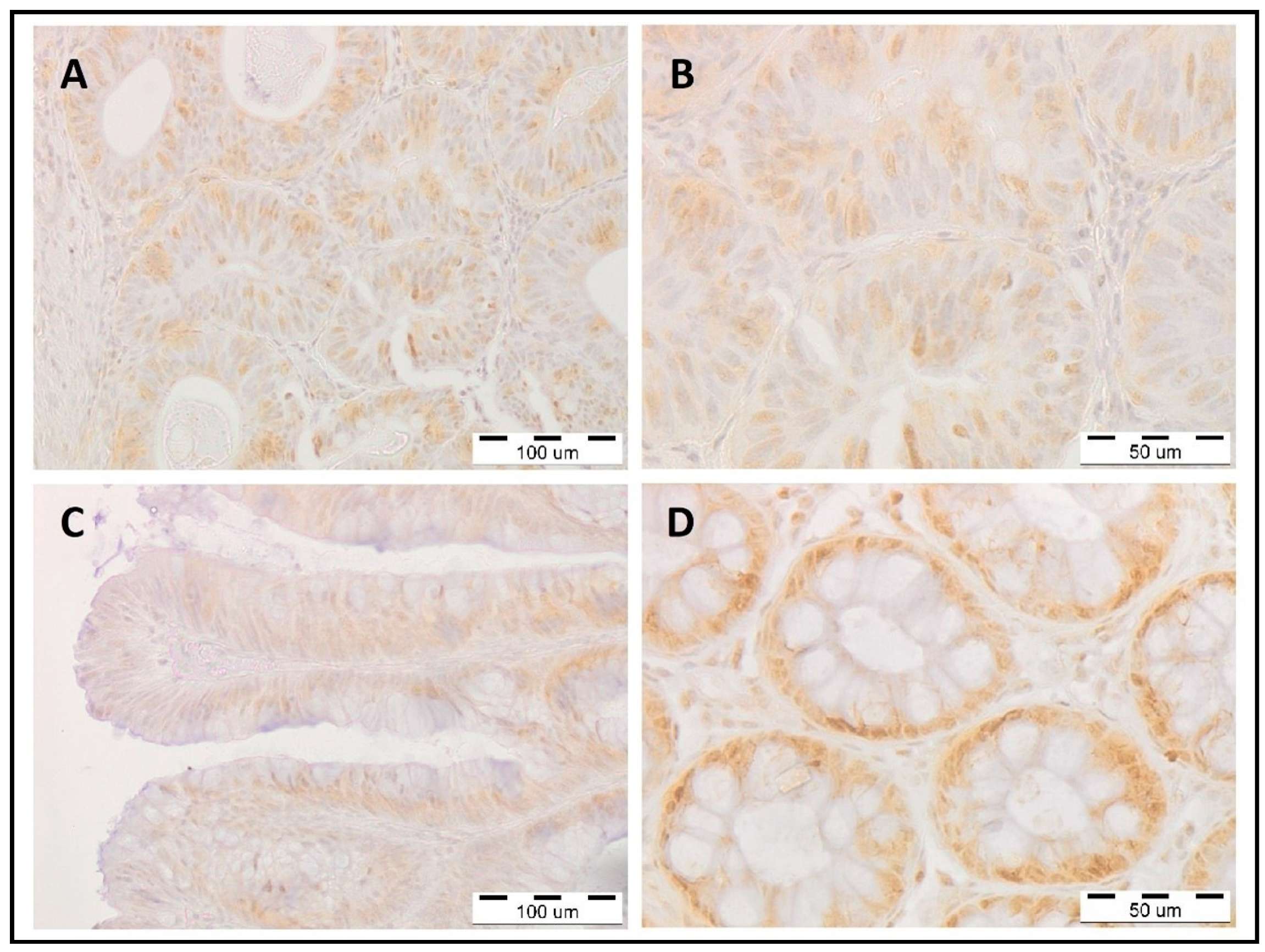
4.1. Colorectal Cancer
4.2. Endometrial Cancer
4.3. Renal Cancer
4.4. Liver Cancer
4.5. Breast Cancer
4.6. Other Cancers
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ganesh, K.; Massagué, J. Targeting metastatic cancer. Nat. Med. 2021, 27, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.; Cheblal, A.; Gasser, S.M. Underappreciated Roles of DNA Polymerase δ in Replication Stress Survival. Trends Genet. 2021, 37, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.R.; Hao, H.; Jiang, Y.; Lee, M.Y. Regulation of human DNA polymerase delta during the cell cycle. J. Biol. Chem. 1994, 269, 24027–24033. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Rice, W.J.; Malik, R.; Johnson, R.E.; Prakash, L.; Prakash, S.; Ubarretxena-Belandia, I.; Aggarwal, A.K. Cryo-EM structure and dynamics of eukaryotic DNA polymerase δ holoenzyme. Nat. Struct. Mol. Biol. 2019, 26, 955–962. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, Y.; Zhang, S.; Lee, E.Y.; Frick, D.N.; Lee, M.Y. DNA damage alters DNA polymerase delta to a form that exhibits increased discrimination against modified template bases and mismatched primers. Nucleic Acids Res. 2009, 37, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, E.; Golemis, E.A.; Arora, S. POLD1: Central mediator of DNA replication and repair, and implication in cancer and other pathologies. Gene 2016, 590, 128–141. [Google Scholar] [CrossRef]
- Lemoine, F.J.; Degtyareva, N.P.; Kokoska, R.J.; Petes, T.D. Reduced levels of DNA polymerase delta induce chromosome fragile site instability in yeast. Mol. Cell. Biol. 2008, 28, 5359–5368. [Google Scholar] [CrossRef] [Green Version]
- Rayner, E.; van Gool, I.C.; Palles, C.; Kearsey, S.E.; Bosse, T.; Tomlinson, I.; Church, D.N. A panoply of errors: Polymerase proofreading domain mutations in cancer. Nat. Rev. Cancer. 2016, 16, 71–81. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, Q.; Wang, Y.N.; Jin, Y.; He, M.M.; Liu, Z.X.; Xu, R.H. Evaluation of POLE and POLD1 Mutations as Biomarkers for Immunotherapy Outcomes Across Multiple Cancer Types. JAMA Oncol. 2019, 5, 1504–1506. [Google Scholar] [CrossRef] [Green Version]
- Briggs, S.; Tomlinson, I. Germline and somatic polymerase ε and δ mutations define a new class of hypermutated colorectal and endometrial cancers. J. Pathol. 2013, 230, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, S.A.; Morreau, H.; de Miranda, N.F.C.C.; van Wezel, T. The missing heritability of familial colorectal cancer. Mutagenesis 2020, 35, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siraj, A.K.; Bu, R.; Iqbal, K.; Parvathareddy, S.K.; Masoodi, T.; Siraj, N.; Al-Rasheed, M.; Kong, Y.; Ahmed, S.O.; Al-Obaisi, K.A.S.; et al. POLE and POLD1 germline exonuclease domain pathogenic variants, a rare event in colorectal cancer from the Middle East. Mol. Genet. Genom. Med. 2020, 8, e1368. [Google Scholar] [CrossRef]
- Siraj, A.K.; Parvathareddy, S.K.; Bu, R.; Iqbal, K.; Siraj, S.; Masoodi, T.; Concepcion, R.M.; Ghazwani, L.O.; AlBadawi, I.; Al-Dayel, F.; et al. Germline POLE and POLD1 proofreading domain mutations in endometrial carcinoma from Middle Eastern region. Cancer Cell Int. 2019, 19, 334. [Google Scholar] [CrossRef] [PubMed]
- Godlewski, J.; Stefaniak, P.; Kiezun, J.; Krazinski, B.E. DNA Polymerase Delta 1 Catalytic Subunit (POLD1) as a Prognostic Factor in Clear Cell Renal Cell Carcinoma Patients. In Vivo 2022, 36, 1188–1194. [Google Scholar] [CrossRef]
- Sanefuji, K.; Taketomi, A.; Iguchi, T.; Sugimachi, K.; Ikegami, T.; Yamashita, Y.; Gion, T.; Soejima, Y.; Shirabe, K.; Maehara, Y. Significance of DNA polymerase delta catalytic subunit p125 induced by mutant p53 in the invasive potential of human hepatocellular carcinoma. Oncology 2010, 79, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; You, T.; Sun, Z.; Bai, C. A Comprehensive Prognostic Analysis of POLD1 in Hepatocellular Carcinoma. BMC Cancer 2022, 22, 197. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, W.; Zhu, X.; Wei, C. p53 inhibits the expression of p125 and the methylation of POLD1 gene promoter by downregulating the Sp1-induced DNMT1 activities in breast cancer. Onco Targets Ther. 2016, 9, 1351–1360. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.; Tan, Q.; Li, J.; Yang, W.; Lian, B.; Mo, Q.; Wei, C. Elevated expression of POLD1 is associated with poor prognosis in breast cancer. Oncol. Lett. 2018, 16, 5591–5598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, J.; Yang, H.; Pan, C.; Li, H.; Luo, Y.; Cheng, T. Multiple microarray analyses identify key genes associated with the development of Non-Small Cell Lung Cancer from Chronic Obstructive Pulmonary Disease. J. Cancer 2021, 12, 996–1010. [Google Scholar] [CrossRef]
- Siraj, A.K.; Bu, R.; Arshad, M.; Iqbal, K.; Parvathareddy, S.K.; Masoodi, T.; Ghazwani, L.O.; Al-Sobhi, S.S.; Al-Dayel, F.; Al-Kuraya, K.S. POLE and POLD1 pathogenic variants in the proofreading domain in papillary thyroid cancer. Endocr. Connect. 2020, 9, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Kucherlapati, M. Examining transcriptional changes to DNA replication and repair factors over uveal melanoma subtypes. BMC Cancer 2018, 18, 818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Liu, D.; Wang, L.; Liu, M.; Duan, W.; Yi, J.; Yi, Y. DNA Repair Genes Are Associated with Subtype Classification, Prognosis, and Immune Infiltration in Uveal Melanoma. J. Oncol. 2022, 2022, 1965451. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Deng, L.; Bai, H.X.; Sun, J.; Neale, N.; Wu, J.; Wang, Y.; Chang, K.; Huang, R.Y.; Zhang, P.J.; et al. Reduced expression of DNA repair genes and chemosensitivity in 1p19q codeleted lower-grade gliomas. J. Neurooncol. 2018, 139, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, C.; Wang, W.; Liu, W.; Zhang, G. Abnormally high expression of POLD1, MCM2, and PLK4 promotes relapse of acute lymphoblastic leukemia. Medicine 2018, 97, e10734. [Google Scholar] [CrossRef]
- Gao, S.; Song, Q.; Liu, J.; Zhang, X.; Ji, X.; Wang, P. E2F1 mediates the downregulation of POLD1 in replicative senescence. Cell. Mol. Life Sci. 2019, 76, 2833–2850. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.W.; Zhang, J.A.; Tan, C.K.; Davie, E.W.; So, A.G.; Downey, K.M. Primary structure of the catalytic subunit of human DNA polymerase delta and chromosomal location of the gene. Proc. Natl. Acad. Sci. USA 1991, 88, 11197–11201. [Google Scholar] [CrossRef] [Green Version]
- Kemper, R.R.; Ahn, E.R.; Zhang, P.; Lee, M.Y.; Rabin, M. Human DNA polymerase delta gene maps to region 19q13.3-q13.4 by in situ hybridization. Genomics 1992, 14, 205–206. [Google Scholar] [CrossRef]
- Zhao, L.; Chang, L.S. The human POLD1 gene. Identification of an upstream activator sequence, activation by Sp1 and Sp3, and cell cycle regulation. J. Biol. Chem. 1997, 272, 4869–4882. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Song, Q.; Gao, S.; Zhang, X.; Wang, Y.; Liu, J.; Fu, J.; Cao, M.; Wang, P. CTCF Mediates Replicative Senescence Through POLD1. Front. Cell Dev. Biol. 2021, 9, 618586. [Google Scholar] [CrossRef]
- Li, B.; Lee, M.Y. Transcriptional regulation of the human DNA polymerase delta catalytic subunit gene POLD1 by p53 tumor suppressor and Sp1. J. Biol. Chem. 2001, 276, 29729–29739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, N.; Zhu, X.; Shi, L.; An, J.; Wu, Y.; Sang, J. Identification and functional analysis of a CDE/CHR element in the POLD1 promoter. Sci. China C Life Sci. 2009, 52, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Quaas, M.; Steiner, L.; Engeland, K. The p53-p21-DREAM-CDE/CHR pathway regulates G2/M cell cycle genes. Nucleic Acids Res. 2016, 44, 164–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czochor, J.R.; Sulkowski, P.; Glazer, P.M. miR-155 Overexpression Promotes Genomic Instability by Reducing High-fidelity Polymerase Delta Expression and Activating Error-Prone DSB Repair. Mol. Cancer Res. 2016, 14, 363–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.L.; Guo, H.L.; Wang, P.C.; Liu, C.G. Age-dependent down-regulation of DNA polymerase δ1 in human lymphocytes. Mol. Cell. Biochem. 2012, 371, 157–163. [Google Scholar] [CrossRef]
- Lee, M.Y.W.T.; Wang, X.; Zhang, S.; Zhang, Z.; Lee, E.Y.C. Regulation and Modulation of Human DNA Polymerase δ Activity and Function. Genes 2017, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Khandagale, P.; Peroumal, D.; Manohar, K.; Acharya, N. Human DNA polymerase delta is a pentameric holoenzyme with a dimeric p12 subunit. Life Sci. Alliance 2019, 2, e201900323. [Google Scholar] [CrossRef] [Green Version]
- Morrison, A.; Bell, J.B.; Kunkel, T.A.; Sugino, A. Eukaryotic DNA polymerase amino acid sequence required for 3′----5′ exonuclease activity. Proc. Natl. Acad. Sci. USA 1991, 88, 9473–9477. [Google Scholar] [CrossRef] [Green Version]
- Dilley, R.L.; Verma, P.; Cho, N.W.; Winters, H.D.; Wondisford, A.R.; Greenberg, R.A. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 2016, 539, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Drosopoulos, W.C.; Vierra, D.A.; Kenworthy, C.A.; Coleman, R.A.; Schildkraut, C.L. Dynamic Assembly and Disassembly of the Human DNA Polymerase δ Holoenzyme on the Genome In Vivo. Cell Rep. 2020, 30, 1329–1341.e5. [Google Scholar] [CrossRef]
- Murga, M.; Lecona, E.; Kamileri, I.; Díaz, M.; Lugli, N.; Sotiriou, S.K.; Anton, M.E.; Méndez, J.; Halazonetis, T.D.; Fernandez-Capetillo, O. POLD3 Is Haploinsufficient for DNA Replication in Mice. Mol. Cell. 2016, 63, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Liu, P.; Jiang, T.; Hu, Y.; Au, F.K.C.; Qi, R.Z. The catalytic subunit of DNA polymerase δ inhibits γTuRC activity and regulates Golgi-derived microtubules. Nat. Commun. 2017, 8, 554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Wang, K.; Qi, R.Z. The catalytic subunit of DNA polymerase δ is a nucleocytoplasmic shuttling protein. Exp. Cell Res. 2019, 375, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Zhang, S.; Lin, S.H.; Wang, X.; Darzynkiewicz, Z.; Zhang, Z.; Lee, E.Y. The tail that wags the dog: p12, the smallest subunit of DNA polymerase δ, is degraded by ubiquitin ligases in response to DNA damage and during cell cycle progression. Cell Cycle 2014, 13, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhao, H.; Darzynkiewicz, Z.; Zhou, P.; Zhang, Z.; Lee, E.Y.; Lee, M.Y. A novel function of CRL4(Cdt2): Regulation of the subunit structure of DNA polymerase δ in response to DNA damage and during the S phase. J. Biol. Chem. 2013, 288, 29550–29561. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhou, Y.; Trusa, S.; Meng, X.; Lee, E.Y.; Lee, M.Y. A novel DNA damage response: Rapid degradation of the p12 subunit of dna polymerase delta. J. Biol. Chem. 2007, 282, 15330–15340. [Google Scholar] [CrossRef] [Green Version]
- Weedon, M.N.; Ellard, S.; Prindle, M.J.; Caswell, R.; Lango Allen, H.; Oram, R.; Godbole, K.; Yajnik, C.S.; Sbraccia, P.; Novelli, G.; et al. An in-frame deletion at the polymerase active site of POLD1 causes a multisystem disorder with lipodystrophy. Nat. Genet. 2013, 45, 947–950. [Google Scholar] [CrossRef] [Green Version]
- Elouej, S.; Beleza-Meireles, A.; Caswell, R.; Colclough, K.; Ellard, S.; Desvignes, J.P.; Béroud, C.; Lévy, N.; Mohammed, S.; De Sandre-Giovannoli, A. Exome sequencing reveals a de novo POLD1 mutation causing phenotypic variability in mandibular hypoplasia, deafness, progeroid features, and lipodystrophy syndrome (MDPL). Metabolism 2017, 71, 213–225. [Google Scholar] [CrossRef]
- Oh, D.Y.; Matsumoto, Y.; Kitajiri, S.I.; Kim, N.K.D.; Kim, M.Y.; Kim, A.R.; Lee, M.; Lee, C.; Tomkinson, A.E.; Katsuno, T.; et al. POLD1 variants leading to reduced polymerase activity can cause hearing loss without syndromic features. Hum. Mutat. 2020, 41, 913–920. [Google Scholar] [CrossRef]
- Conde, C.D.; Petronczki, Ö.Y.; Baris, S.; Willmann, K.L.; Girardi, E.; Salzer, E.; Weitzer, S.; Ardy, R.C.; Krolo, A.; Ijspeert, H.; et al. Polymerase δ deficiency causes syndromic immunodeficiency with replicative stress. J. Clin. Investig. 2019, 129, 4194–4206. [Google Scholar] [CrossRef]
- Cui, Y.; Keles, S.; Charbonnier, L.M.; Julé, A.M.; Henderson, L.; Celik, S.C.; Reisli, I.; Shen, C.; Xie, W.J.; Schmitz-Abe, K.; et al. Combined immunodeficiency caused by a loss-of-function mutation in DNA polymerase delta 1. J. Allergy Clin. Immunol. 2020, 145, 391–401.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols-Vinueza, D.X.; Delmonte, O.M.; Bundy, V.; Bosticardo, M.; Zimmermann, M.T.; Dsouza, N.R.; Pala, F.; Dobbs, K.; Stoddard, J.; Niemela, J.E.; et al. POLD1 Deficiency Reveals a Role for POLD1 in DNA Repair and T and B Cell Development. J. Clin. Immunol. 2021, 41, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Krautscheid, P.; Graham, R.P.; Ganguly, A.; Shankar, S.; Ferber, M.; Hegde, M.; ACMG Laboratory Quality Assurance Committee. Genetic testing for inherited colorectal cancer and polyposis, 2021 revision: A technical standard of the American College of Medical Genetics and Genomics (ACMG). Genet. Med. 2021, 23, 1807–1817. [Google Scholar] [CrossRef]
- Jasperson, K.W.; Tuohy, T.M.; Neklason, D.W.; Burt, R.W. Hereditary and familial colon cancer. Gastroenterology 2010, 138, 2044–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palles, C.; Cazier, J.B.; Howarth, K.M.; Domingo, E.; Jones, A.M.; Broderick, P.; Kemp, Z.; Spain, S.L.; Guarino, E.; Salguero, I.; et al. Germline mutations affecting the proofreading domains of POLE and POLD1 predispose to colorectal adenomas and carcinomas. Nat. Genet. 2013, 45, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayed, F.A.; Tops, C.M.J.; Nielsen, M.; Ruano, D.; Vasen, H.F.A.; Morreau, H.J.; Hes, F.; van Wezel, T. Low frequency of POLD1 and POLE exonuclease domain variants in patients with multiple colorectal polyps. Mol. Genet. Genom. Med. 2019, 7, e00603. [Google Scholar] [CrossRef] [Green Version]
- Mur, P.; García-Mulero, S.; Del Valle, J.; Magraner-Pardo, L.; Vidal, A.; Pineda, M.; Cinnirella, G.; Martín-Ramos, E.; Pons, T.; López-Doriga, A.; et al. Role of POLE and POLD1 in familial cancer. Genet. Med. 2020, 22, 2089–2100. [Google Scholar] [CrossRef]
- Keshinro, A.; Vanderbilt, C.; Kim, J.K.; First, C.; Chen, C.T.; Yaeger, R.; Ganesh, K.; Segal, N.H.; Gonen, M.; Shia, J.; et al. Tumor-Infiltrating Lymphocytes, Tumor Mutational Burden, and Genetic Alterations in Microsatellite Unstable, Microsatellite Stable, or Mutant POLE/POLD1 Colon Cancer. JCO Precis. Oncol. 2021, 5, PO.20.00456. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Ward, E.M.; Johnson, C.J.; Cronin, K.A.; Ma, J.; Ryerson, B.; Mariotto, A.; Lake, A.J.; Wilson, R.; Sherman, R.L.; et al. Annual Report to the Nation on the Status of Cancer, 1975-2014, Featuring Survival. J. Natl. Cancer Inst. 2017, 109, djx030. [Google Scholar] [CrossRef] [Green Version]
- Church, D.N.; Briggs, S.E.; Palles, C.; Domingo, E.; Kearsey, S.J.; Grimes, J.M.; Gorman, M.; Martin, L.; Howarth, K.M.; Hodgson, S.V.; et al. DNA polymerase ε and δ exonuclease domain mutations in endometrial cancer. Hum. Mol. Genet. 2013, 22, 2820–2828. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Kuick, C.H.; Wong, W.L.; Tham, J.M.; Mansor, S.; Loh, E.; Jain, S.; Vikas, N.N.; Tan, S.H.; Chan, S.H.; et al. Mutation spectrum of POLE and POLD1 mutations in South East Asian women presenting with grade 3 endometrioid endometrial carcinomas. Gynecol. Oncol. 2016, 141, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Palles, C.; Martin, L.; Domingo, E.; Chegwidden, L.; McGuire, J.; Cuthill, V.; Heitzer, E.; CORGI Consortium; Kerr, R.; Kerr, D.; et al. The clinical features of polymerase proof-reading associated polyposis (PPAP) and recommendations for patient management. Fam. Cancer 2022, 21, 197–209. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ouyang, W.; Zhao, W.; Shao, L.; Li, B.; Liu, B.; Wang, D.; Han-Zhang, H.; Zhang, Z.; Shao, L.; et al. Distinctive genomic characteristics in POLE/POLD1-mutant cancers can potentially predict beneficial clinical outcomes in patients who receive immune checkpoint inhibitor. Ann. Transl. Med. 2021, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Haraldsdottir, S.; Hampel, H.; Tomsic, J.; Frankel, W.L.; Pearlman, R.; de la Chapelle, A.; Pritchard, C.C. Colon and endometrial cancers with mismatch repair deficiency can arise from somatic, rather than germline, mutations. Gastroenterology 2014, 147, 1308–1316.e1. [Google Scholar] [CrossRef] [Green Version]
- Haradhvala, N.J.; Kim, J.; Maruvka, Y.E.; Polak, P.; Rosebrock, D.; Livitz, D.; Hess, J.M.; Leshchiner, I.; Kamburov, A.; Mouw, K.W.; et al. Distinct mutational signatures characterize concurrent loss of polymerase proofreading and mismatch repair. Nat. Commun. 2018, 9, 1746. [Google Scholar] [CrossRef] [Green Version]
- Schamschula, E.; Kinzel, M.; Wernstedt, A.; Oberhuber, K.; Gottschling, H.; Schnaiter, S.; Friedrichs, N.; Merkelbach-Bruse, S.; Zschocke, J.; Gallon, R.; et al. Teenage-Onset Colorectal Cancers in a Digenic Cancer Predisposition Syndrome Provide Clues for the Interaction between Mismatch Repair and Polymerase δ Proofreading Deficiency in Tumorigenesis. Biomolecules 2022, 12, 1350. [Google Scholar] [CrossRef]
- Jansen, A.M.; van Wezel, T.; van den Akker, B.E.; Ventayol Garcia, M.; Ruano, D.; Tops, C.M.; Wagner, A.; Letteboer, T.G.; Gómez-García, E.B.; Devilee, P.; et al. Combined mismatch repair and POLE/POLD1 defects explain unresolved suspected Lynch syndrome cancers. Eur. J. Hum. Genet. 2016, 24, 1089–1092. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kitazono, I.; Akahane, T.; Yanazume, S.; Kamio, M.; Togami, S.; Nohara, S.; Sakamoto, I.; Yokoyama, S.; Tabata, K.; et al. Molecular Evaluation of Endometrial Dedifferentiated Carcinoma, Endometrioid Carcinoma, Carcinosarcoma, and Serous Carcinoma Using a Custom-Made Small Cancer Panel. Pathol. Oncol. Res. 2021, 27, 1610013. [Google Scholar] [CrossRef]
- Li, K.; Luo, H.; Huang, L.; Luo, H.; Zhu, X. Microsatellite instability: A review of what the oncologist should know. Cancer Cell Int. 2020, 20, 16. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Dong, L.; Liu, X.; Ou, K.; Yang, L. POLE/POLD1 mutation and tumor immunotherapy. J. Exp. Clin. Cancer Res. 2022, 41, 216. [Google Scholar] [CrossRef]
- Cohen, H.T.; McGovern, F.J. Renal-cell carcinoma. N. Engl. J. Med. 2005, 353, 2477–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Tang, H.; Xu, W.H.; Tang, H.; Wei, S.; Anwaier, A.; Huang, H.; Qu, Y.Y.; Zhang, H.; Zhao, S.; et al. Protumorigenic Role of Elevated Levels of DNA Polymerase Epsilon Predicts an Immune-Suppressive Microenvironment in Clear Cell Renal Cell Carcinoma. Front. Genet. 2021, 12, 751977. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Zhang, Z.; Zhang, Y.; Li, J.; Liang, G.; Ling, J. Effect of Smilax china L.-containing serum on the expression of POLD1 mRNA in human hepatocarcinoma SMMC-7721 cells. Exp. Ther. Med. 2013, 6, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.J.; Wan, Y.; Zhu, D.D.; Wang, M.X.; Jiang, H.M.; Huang, D.L.; Luo, L.F.; Chen, M.J.; Yang, W.P.; Li, H.M.; et al. Resveratrol Mediates the Apoptosis of Triple Negative Breast Cancer Cells by Reducing POLD1 Expression. Front. Oncol. 2021, 11, 569295. [Google Scholar] [CrossRef]
- Ho, D.W.; Lam, W.M.; Chan, L.K.; Ng, I.O. Investigation of Functional Synergism of CENPF and FOXM1 Identifies POLD1 as Downstream Target in Hepatocellular Carcinoma. Front. Med. 2022, 9, 860395. [Google Scholar] [CrossRef] [PubMed]
- Sigurdson, A.J.; Hauptmann, M.; Chatterjee, N.; Alexander, B.H.; Doody, M.M.; Rutter, J.L.; Struewing, J.P. Kin-cohort estimates for familial breast cancer risk in relation to variants in DNA base excision repair, BRCA1 interacting and growth factor genes. BMC Cancer 2004, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Bellido, F.; Pineda, M.; Aiza, G.; Valdés-Mas, R.; Navarro, M.; Puente, D.A.; Pons, T.; González, S.; Iglesias, S.; Darder, E.; et al. POLE and POLD1 mutations in 529 kindred with familial colorectal cancer and/or polyposis: Review of reported cases and recommendations for genetic testing and surveillance. Genet. Med. 2016, 18, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Nik-Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of somatic mutations in 560 breast cancer whole-genome sequences. Nature 2016, 534, 47–54, Erratum in Nature 2019, 566, E1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Encinas, G.; Sabelnykova, V.Y.; de Lyra, E.C.; Hirata Katayama, M.L.; Maistro, S.; de Vasconcellos Valle, P.W.M.; de Lima Pereira, G.F.; Rodrigues, L.M.; de Menezes Pacheco Serio, P.A.; de Gouvêa, A.C.R.C.; et al. Somatic mutations in early onset luminal breast cancer. Oncotarget 2018, 9, 22460–22479. [Google Scholar] [CrossRef] [Green Version]
- Anbazhagan, R.; Fujii, H.; Gabrielson, E. Microsatellite instability is uncommon in breast cancer. Clin. Cancer Res. 1999, 5, 839–844. [Google Scholar] [PubMed]
- da Silva, C.M.; Shimba, D.; Oishi, D.; Cagnacci, A.; de Gouvea, A.C.R.C.; Ades, F. VUS-type alteration in POLD1 and microsatellite instability in a metastatic luminal B breast cancer patient. Ecancermedicalscience 2020, 14, 1002. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qin, Q.; Chen, R.; Wei, C.; Mo, Q. SIRT1 promotes proliferation, migration, and invasion of breast cancer cell line MCF-7 by upregulating DNA polymerase delta1 (POLD1). Biochem. Biophys. Res. Commun. 2018, 502, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Anurag, M.; Jaehnig, E.J.; Krug, K.; Lei, J.T.; Bergstrom, E.J.; Kim, B.J.; Vashist, T.D.; Huynh, A.M.T.; Dou, Y.; Gou, X.; et al. Proteogenomic Markers of Chemotherapy Resistance and Response in Triple-Negative Breast Cancer. Cancer Discov. 2022, 12, 2586–2605. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Gong, Y.; Zhao, W.; Han, Z.; Guo, S.; Liu, H.; Peng, X.; Xiao, W.; Li, Y.; Dang, S.; et al. Comprehensive analysis of POLE and POLD1 Gene Variations identifies cancer patients potentially benefit from immunotherapy in Chinese population. Sci. Rep. 2019, 9, 15767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Røe, O.D.; Szulkin, A.; Anderssen, E.; Flatberg, A.; Sandeck, H.; Amundsen, T.; Erlandsen, S.E.; Dobra, K.; Sundstrøm, S.H. Molecular resistance fingerprint of pemetrexed and platinum in a long-term survivor of mesothelioma. PLoS ONE 2012, 7, e40521. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhai, L.; Ma, S.; Zhang, C.; Zhao, L.; Li, N.; Xu, Y.; Zhang, T.; Guo, Z.; Zhang, H.; et al. Down-regulation of RIP3 potentiates cisplatin chemoresistance by triggering HSP90-ERK pathway mediated DNA repair in esophageal squamous cell carcinoma. Cancer Lett. 2018, 418, 97–108. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Liu, G.; Zha, W.; Liu, Y. Cadherin-16 inhibits thyroid carcinoma cell proliferation and invasion. Oncol Lett. 2022, 23, 145. [Google Scholar] [CrossRef]
- Zhu, M.; Cui, H.; Zhang, L.; Zhao, K.; Jia, X.; Jin, H. Assessment of POLE and POLD1 mutations as prognosis and immunotherapy biomarkers for stomach adenocarcinoma. Transl. Cancer Res. 2022, 11, 193–205. [Google Scholar] [CrossRef]
- Johnson, A.; Severson, E.; Gay, L.; Vergilio, J.A.; Elvin, J.; Suh, J.; Daniel, S.; Covert, M.; Frampton, G.M.; Hsu, S.; et al. Comprehensive Genomic Profiling of 282 Pediatric Low- and High-Grade Gliomas Reveals Genomic Drivers, Tumor Mutational Burden, and Hypermutation Signatures. Oncologist 2017, 22, 1478–1490. [Google Scholar] [CrossRef] [Green Version]
- Mackay, A.; Burford, A.; Molinari, V.; Jones, D.T.W.; Izquierdo, E.; Brouwer-Visser, J.; Giangaspero, F.; Haberler, C.; Pietsch, T.; Jacques, T.S.; et al. Molecular, Pathological, Radiological, and Immune Profiling of Non-brainstem Pediatric High-Grade Glioma from the HERBY Phase II Randomized Trial. Cancer Cell 2018, 33, 829–842.e5. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.A.; Kim, D.; Lee, B.; Shim, J.H.; Suh, Y.L. Incidence, clinicopathologic, and genetic characteristics of mismatch repair gene-mutated glioblastomas. J. Neurooncol. 2021, 153, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wei, C.; Chen, L.; Liu, N.; Ou, Q.; Yin, J.C.; Pang, J.; Fang, Z.; Wu, X.; Wang, X.; et al. Genomic Correlates of Unfavorable Outcome in Locally Advanced Cervical Cancer Treated with Neoadjuvant Chemoradiation. Cancer Res. Treat. 2022, 54, 1209–1218. [Google Scholar] [CrossRef]
- Xing, L.; Mi, W.; Zhang, Y.; Tian, S.; Zhang, Y.; Qi, R.; Lou, G.; Zhang, C. The identification of six risk genes for ovarian cancer platinum response based on global network algorithm and verification analysis. J. Cell. Mol. Med. 2020, 24, 9839–9852. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.X.; Lujan, S.A.; Burkholder, A.B.; St Charles, J.; Dahl, J.; Farrell, C.E.; Williams, J.S.; Kunkel, T.A. How asymmetric DNA replication achieves symmetrical fidelity. Nat. Struct. Mol. Biol. 2021, 28, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Andrianova, M.A.; Seplyarskiy, V.B.; Terradas, M.; Sánchez-Heras, A.B.; Mur, P.; Soto, J.L.; Aiza, G.; Kondrashov, F.A.; Kondrashov, A.S.; Bazykin, G.A.; et al. Extended family with germline pathogenic variant in polymerase delta provides strong evidence for recessive effect of proofreading inactivation. bioRxiv 2022, 20, 500591. [Google Scholar] [CrossRef]
- Bonjoch, L.; Soares de Lima, Y.; Díaz-Gay, M.; Dotti, I.; Muñoz, J.; Moreira, L.; Carballal, S.; Ocaña, T.; Cuatrecasas, M.; Ortiz, O.; et al. Unraveling the impact of a germline heterozygous POLD1 frameshift variant in serrated polyposis syndrome. Front. Mol. Biosci. 2023, 10, 1119900. [Google Scholar] [CrossRef]
- Twomey, J.D.; Zhang, B. Cancer Immunotherapy Update: FDA-Approved Checkpoint Inhibitors and Companion Diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef]
- Esfahani, K.; Roudaia, L.; Buhlaiga, N.; Del Rincon, S.V.; Papneja, N.; Miller, W.H., Jr. A review of cancer immunotherapy: From the past, to the present, to the future. Curr. Oncol. 2020, 27, S87–S97. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Cai, X.; Xie, Z.; Zhou, L.; Cheng, B.; Zhong, R.; Xiong, S.; Li, J.; Chen, Z.; et al. The association between CD8+ tumor-infiltrating lymphocytes and the clinical outcome of cancer immunotherapy: A systematic review and meta-analysis. EClinicalMedicine 2021, 41, 101134. [Google Scholar] [CrossRef]
- Brummel, K.; Eerkens, A.L.; de Bruyn, M.; Nijman, H.W. Tumour-infiltrating lymphocytes: From prognosis to treatment selection. Br. J. Cancer. 2023, 128, 451–458. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, Q.; Xu, R.H. Evaluation of POLE/POLD1 Variants as Potential Biomarkers for Immune Checkpoint Inhibitor Treatment Outcomes-Reply. JAMA Oncol. 2020, 6, 590. [Google Scholar] [CrossRef] [PubMed]
- Marron, T.U.; Ryan, A.E.; Reddy, S.M.; Kaczanowska, S.; Younis, R.H.; Thakkar, D.; Zhang, J.; Bartkowiak, T.; Howard, R.; Anderson, K.G.; et al. Considerations for treatment duration in responders to immune checkpoint inhibitors. J. Immunother. Cancer 2021, 9, e001901. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, L.B.; Kim, J.; Haradhvala, N.J.; Huang, M.N.; Tian Ng, A.W.; Wu, Y.; Boot, A.; Covington, K.R.; Gordenin, D.A.; Bergstrom, E.N.; et al. The repertoire of mutational signatures in human cancer. Nature 2020, 578, 94–101, Erratum in Nature 2023, 614, E412023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, P.S.; Coorens, T.H.H.; Palles, C.; Mitchell, E.; Abascal, F.; Olafsson, S.; Lee, B.C.H.; Lawson, A.R.J.; Lee-Six, H.; Moore, L.; et al. Increased somatic mutation burdens in normal human cells due to defective DNA polymerases. Nat. Genet. 2021, 53, 1434–1442. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.W.; Gout, A.M.; Zhang, J. Therapeutic and prognostic insights from the analysis of cancer mutational signatures. Trends Genet. 2022, 38, 194–208. [Google Scholar] [CrossRef]
- Ma, X.; Riaz, N.; Samstein, R.M.; Lee, M.; Makarov, V.; Valero, C.; Chowell, D.; Kuo, F.; Hoen, D.; Fitzgerald, C.W.R.; et al. Functional landscapes of POLE and POLD1 mutations in checkpoint blockade-dependent antitumor immunity. Nat. Genet. 2022, 54, 996–1012. [Google Scholar] [CrossRef]
- Liu, C.; Yang, M.; Zhang, D.; Chen, M.; Zhu, D. Clinical cancer immunotherapy: Current progress and prospects. Front. Immunol. 2022, 13, 961805. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Cancer | Number of Patients | POLD1 Status | Stage/ Grade | Detection Method | Clinicopathological Relevance | Ref. |
|---|---|---|---|---|---|---|
| Colorectal cancer | 1069 | Low | I–IV | IHC |
| [13] |
| Endometrial cancer | 419 | Low | I–IV; mainly type I EC (88.2%) | IHC |
| [14] |
| Clear cell renal cell carcinoma | 56 | High | I–III; mainly lower grades (G1–G2 in 73.2%) | IHC |
| [15] |
| Hepatocellular carcinoma | 339 | High | I–IV; mainly stage I and II tumors (50.1% and 24.8%, respectively) |
mRNA data retrieved from the GEPIA2 database IHC |
| [17] |
|
Breast cancer | 84 | High | G1–G3; invasive breast cancer treated with radical mastectomy | RT-qPCR Western blot |
| [19] |
|
Lung cancer | – | High | I–IV | RT-qPCR |
| [20] |
| Papillary thyroid cancer |
286 (females in 77.7%) | Low | I–IV; mainly stage I tumors (68.3%) | IHC |
| [21] |
| Lower-grade gliomas with 1p19q codeletion | 280 | Low | WHO grade II–III |
RT-qPCR Western blot |
| [24] |
| Childhood Acute lymphoblastic leukemia | 90 | High | – | – |
| [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gola, M.; Stefaniak, P.; Godlewski, J.; Jereczek-Fossa, B.A.; Starzyńska, A. Prospects of POLD1 in Human Cancers: A Review. Cancers 2023, 15, 1905. https://doi.org/10.3390/cancers15061905
Gola M, Stefaniak P, Godlewski J, Jereczek-Fossa BA, Starzyńska A. Prospects of POLD1 in Human Cancers: A Review. Cancers. 2023; 15(6):1905. https://doi.org/10.3390/cancers15061905
Chicago/Turabian StyleGola, Michał, Przemysław Stefaniak, Janusz Godlewski, Barbara Alicja Jereczek-Fossa, and Anna Starzyńska. 2023. "Prospects of POLD1 in Human Cancers: A Review" Cancers 15, no. 6: 1905. https://doi.org/10.3390/cancers15061905
APA StyleGola, M., Stefaniak, P., Godlewski, J., Jereczek-Fossa, B. A., & Starzyńska, A. (2023). Prospects of POLD1 in Human Cancers: A Review. Cancers, 15(6), 1905. https://doi.org/10.3390/cancers15061905