
SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer

 ,
,  , ,
, ,  ,
,  and
and 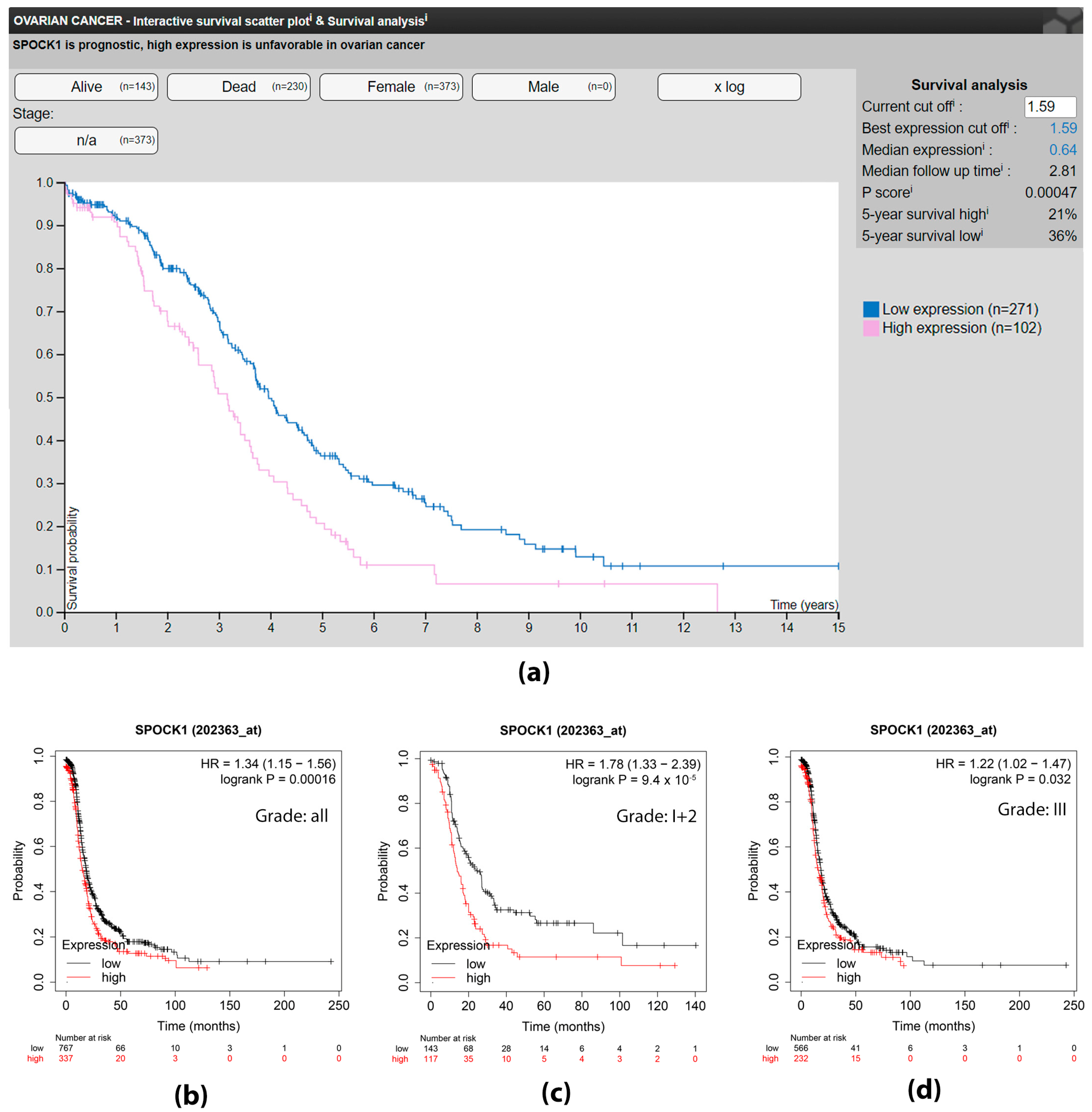{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Cultures
2.3. Transfection
2.4. Immunocytochemistry
2.5. Automated Western Blotting (WES)
2.6. BrdU Incorporation Assay
2.7. Migration Assay
2.8. Human Materials
2.9. Immunhistochemistry
2.10. SPOCK1 ELISA
2.11. In Silico Analysis
2.12. Statistical Analysis
3. Results
3.1. The Effect of SPOCK1 Level on Survival—In Silico Approach
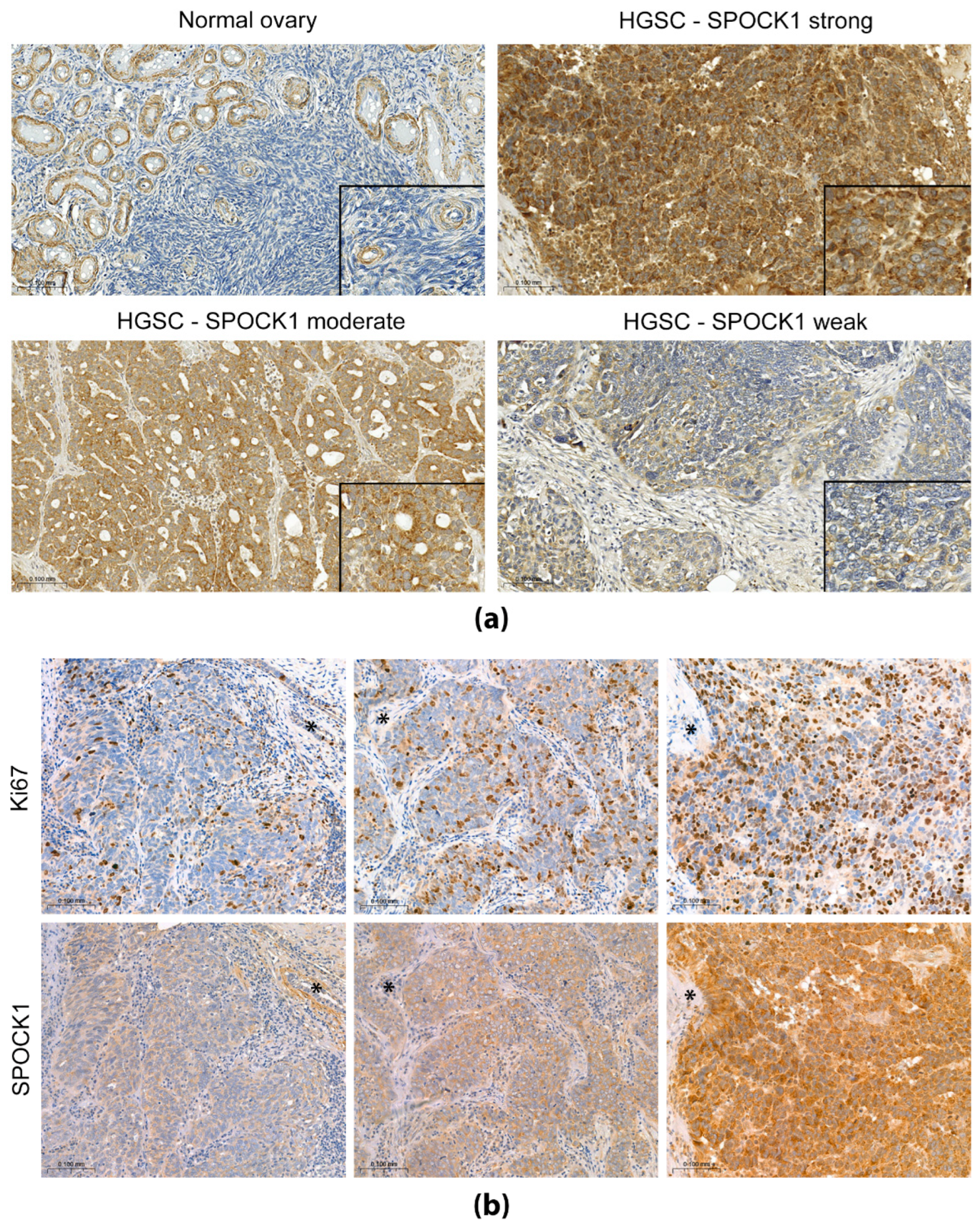
3.2. SPOCK1 Localization and Expression in Human Ovary Samples
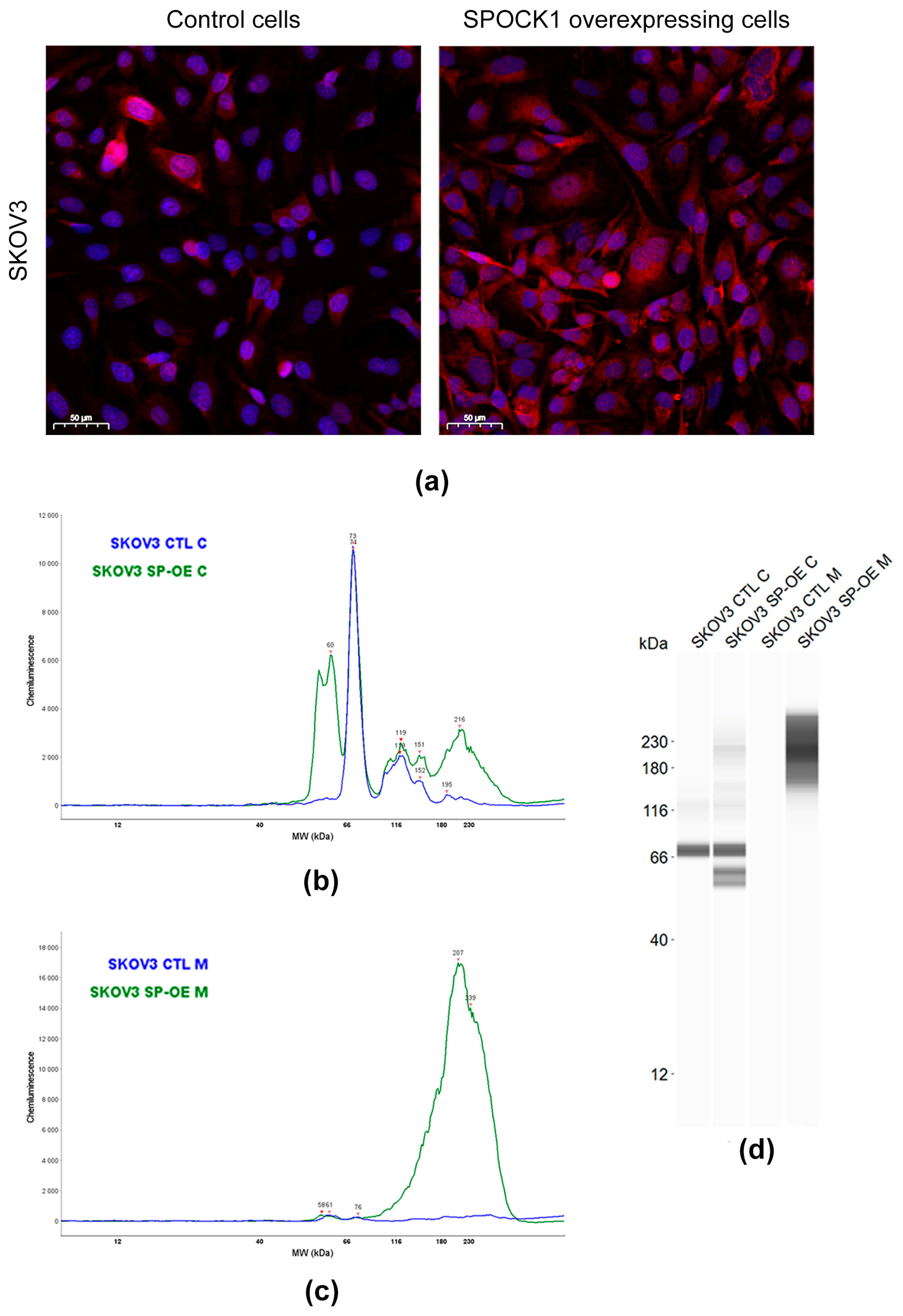
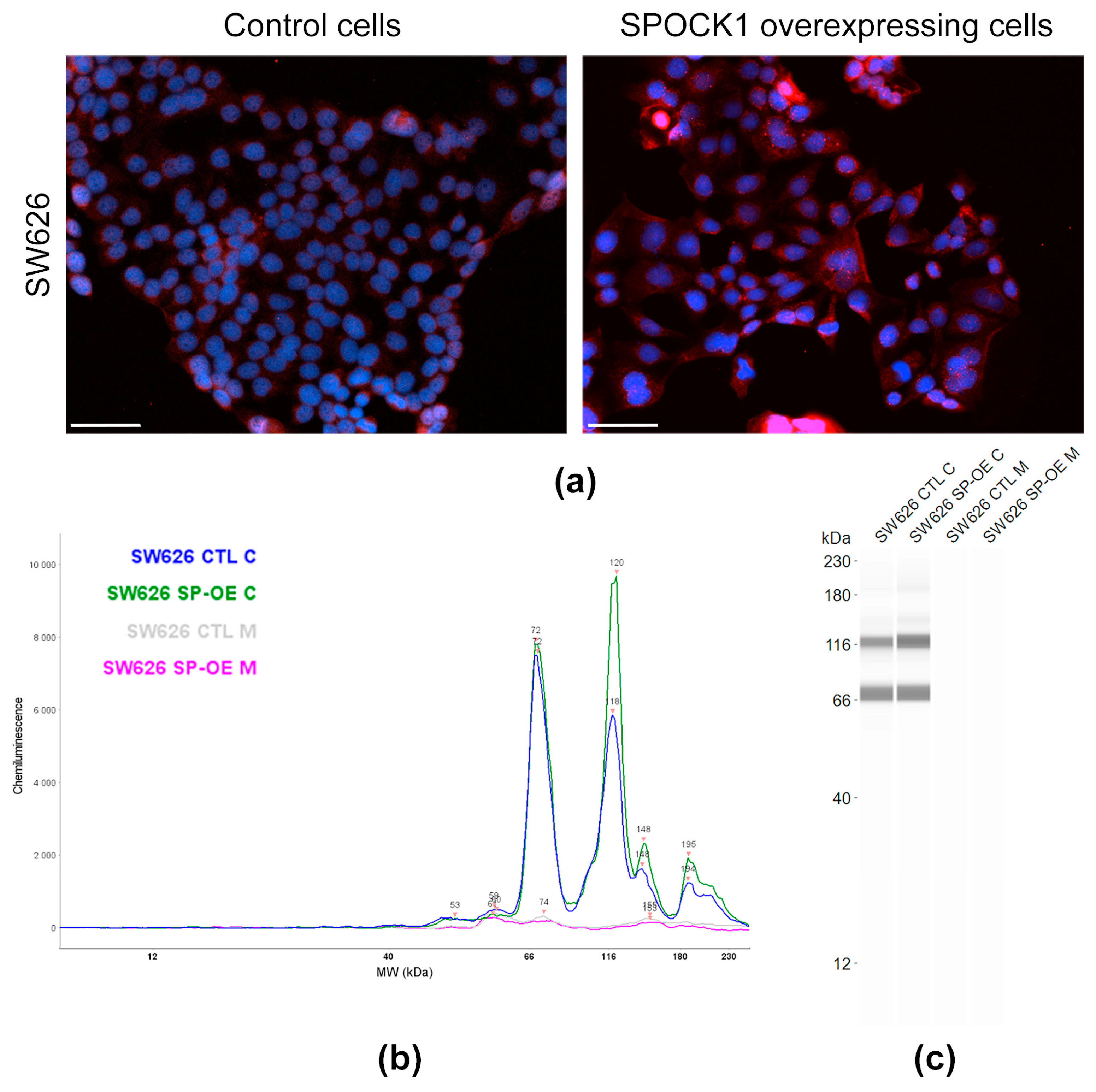
3.3. SPOCK1 Expression in Control and Transfected Tumor Cell Lines
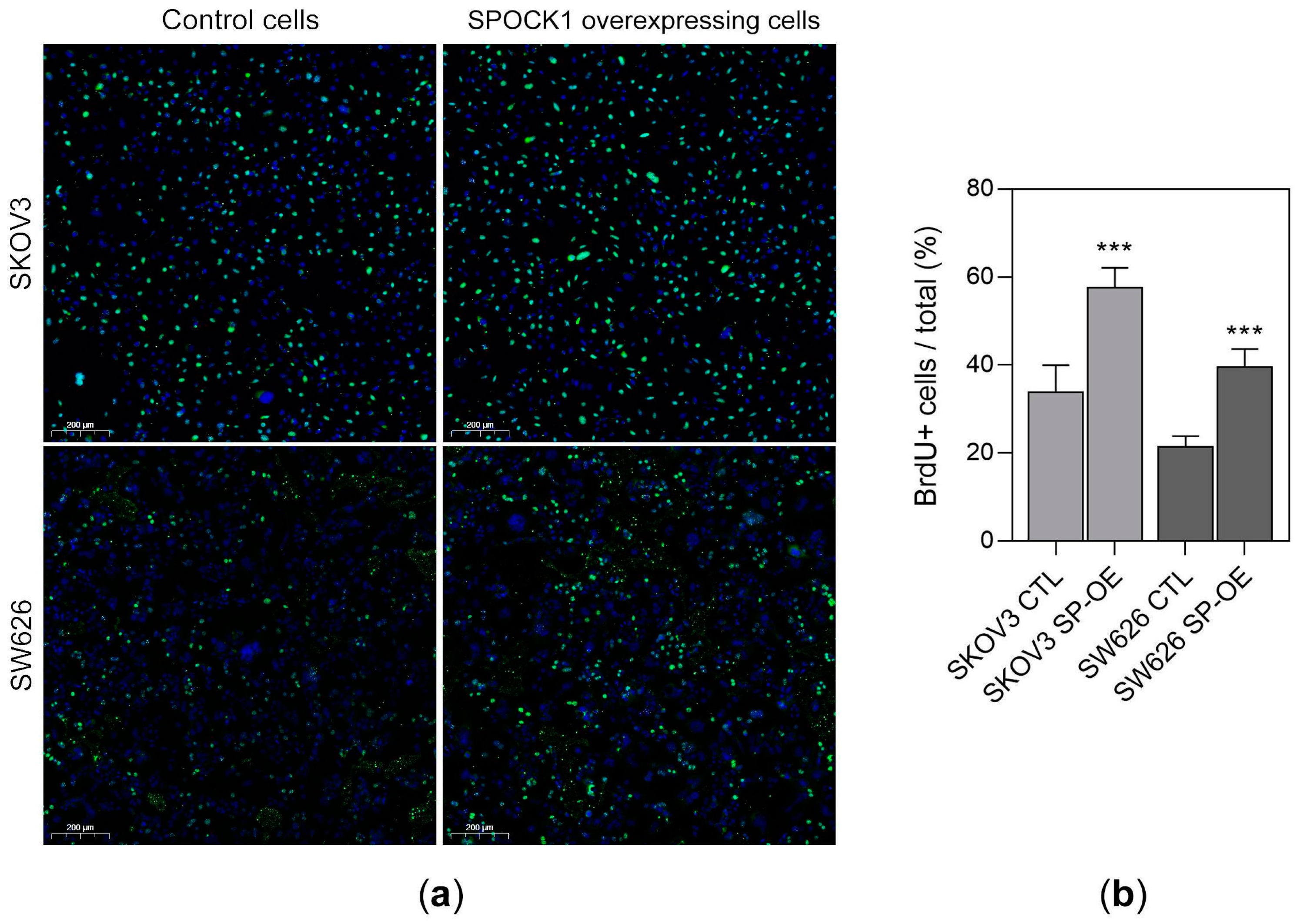
3.4. SPOCK1 Overexpression Promotes DNA Synthesis
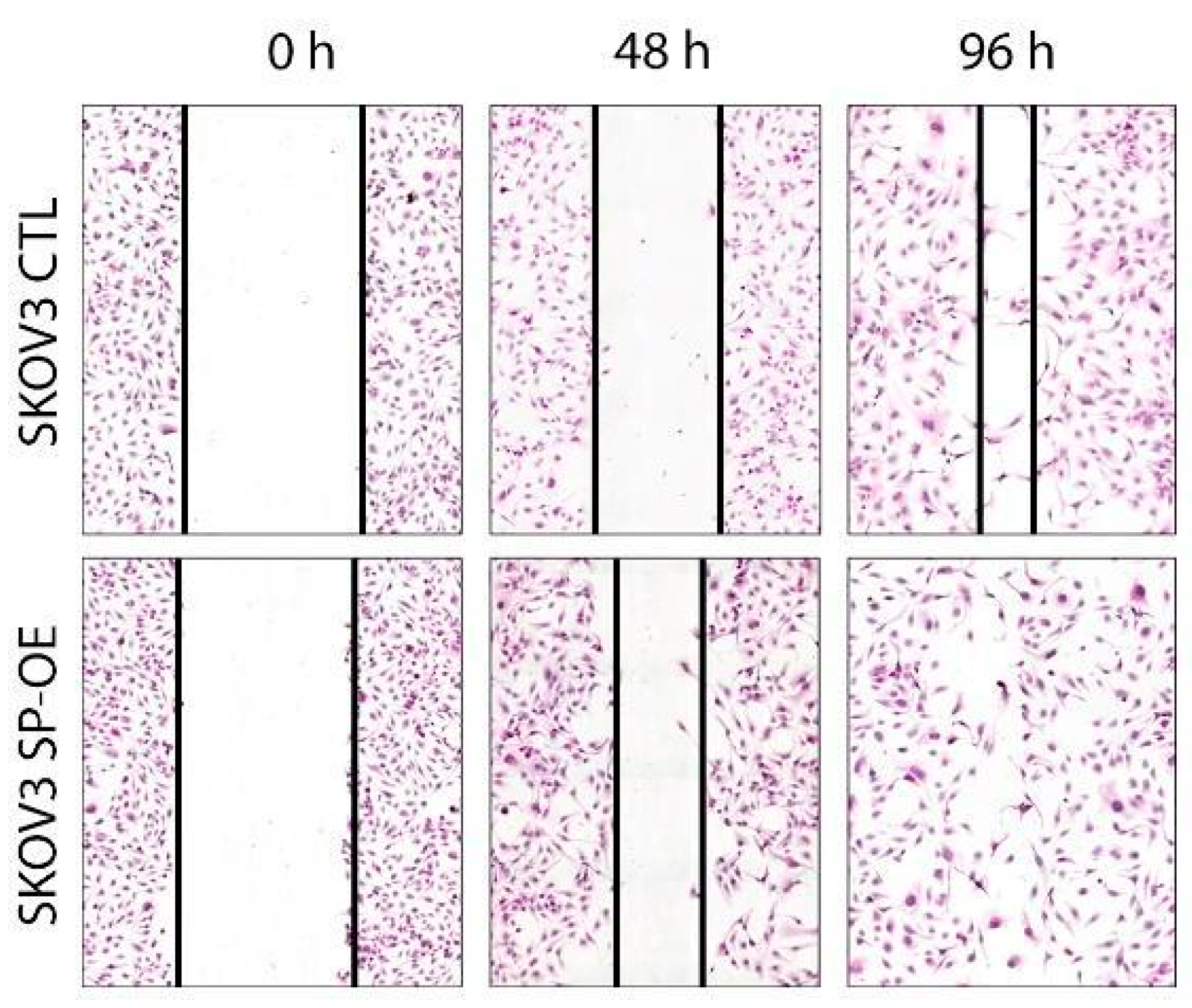
3.5. The Impact of SPOCK1 on Tumor Cell Migration
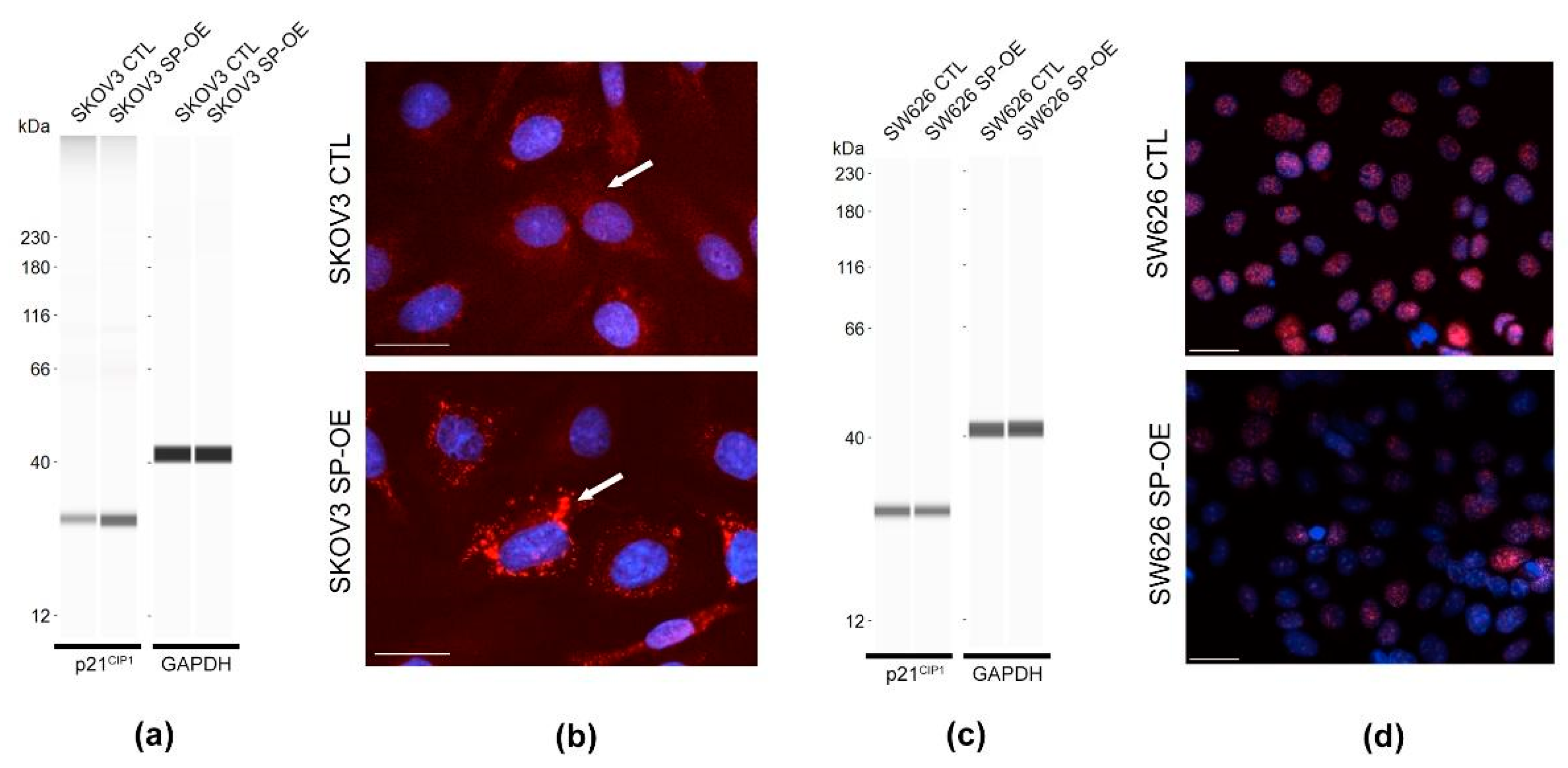
3.6. Tumor Promoting Effect of SPOCK1 via p21CIP1
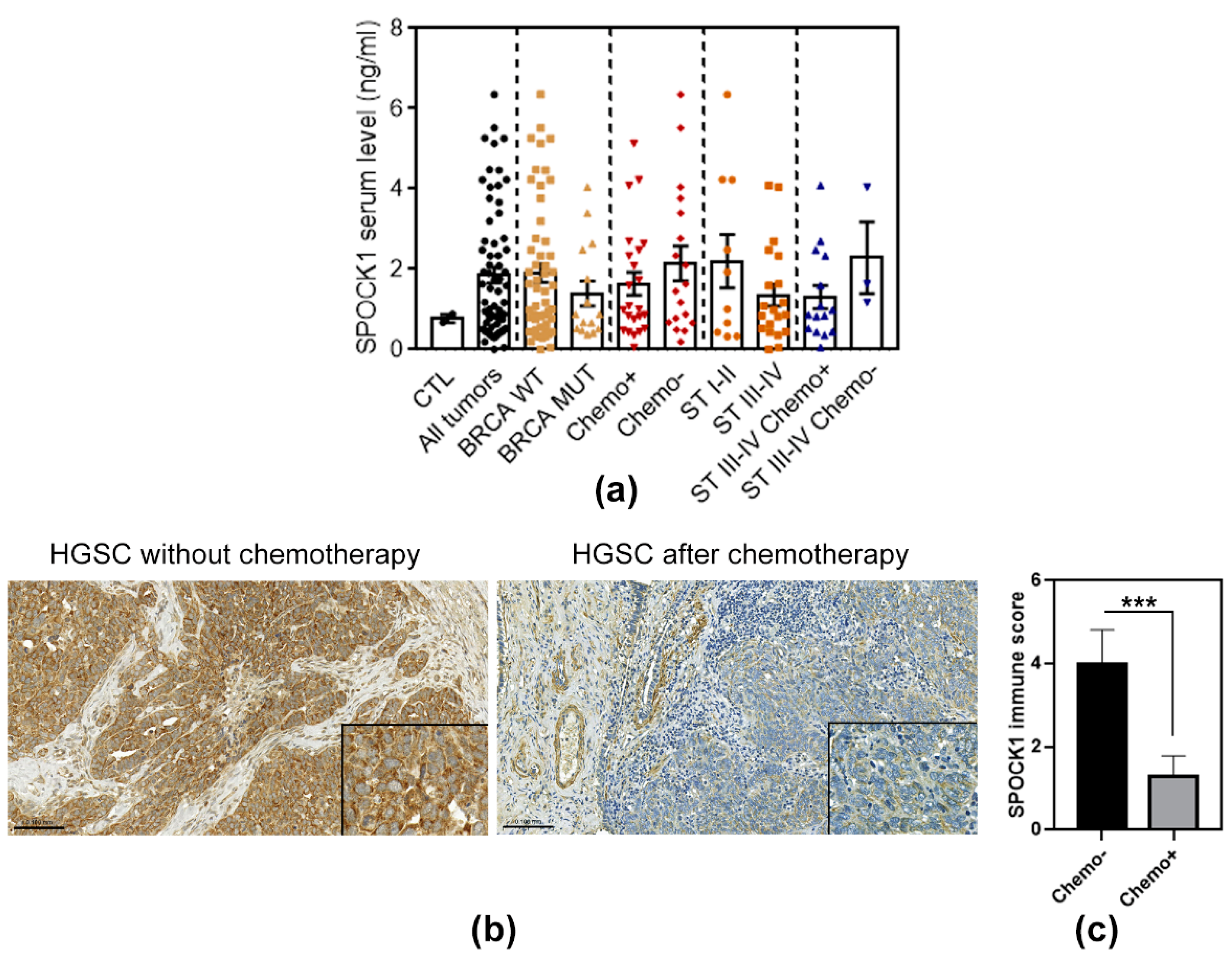
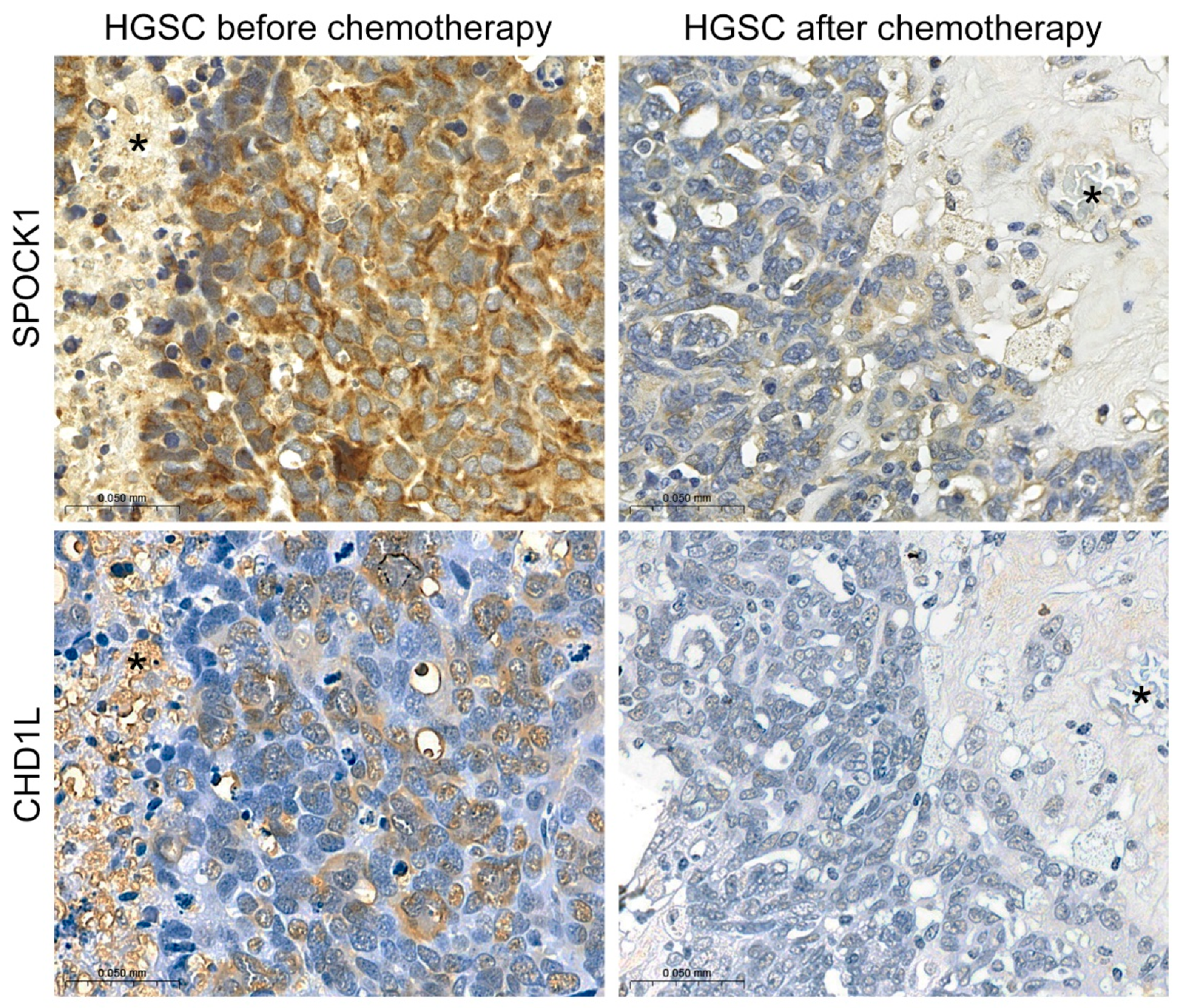
3.7. SPOCK1 Serum and Tissue Levels in Patients with Ovarian Cancer
3.8. SPOCK1 Expression Is Related to the CHD1L Transcription Factor
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Girolimetti, G.; Perrone, A.M.; Santini, D.; Barbieri, E.; Guerra, F.; Ferrari, S.; Zamagni, C.; De Iaco, P.; Gasparre, G.; Turchetti, D. BRCA-associated ovarian cancer: From molecular genetics to risk management. BioMed Res. Int. 2014, 2014, 787143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, X.; Jing, H.; Li, Z. The clinical features and management of Lynch syndrome-associated ovarian cancer. J. Obstet. Gynaecol. Res. 2022, 48, 1538–1545. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Men, X.; Zhang, W.; Lei, P. Advances in tumor markers of ovarian cancer for early diagnosis. Indian J. Cancer 2014, 51 (Suppl. S3), e72–e76. [Google Scholar] [CrossRef]
- Seelenmeyer, C.; Wegehingel, S.; Lechner, J.; Nickel, W. The cancer antigen CA125 represents a novel counter receptor for galectin-1. J. Cell Sci. 2003, 116, 1305–1318. [Google Scholar] [CrossRef] [Green Version]
- Nebgen, D.R.; Lu, K.H.; Bast, R.C. Novel Approaches to Ovarian Cancer Screening. Curr. Oncol. Rep. 2019, 21, 75. [Google Scholar] [CrossRef]
- Orr, B.; Edwards, R.P. Diagnosis and Treatment of Ovarian Cancer. Hematol./Oncol. Clin. North Am. 2018, 32, 943–964. [Google Scholar] [CrossRef] [PubMed]
- Annaval, T.; Wild, R.; Crétinon, Y.; Sadir, R.; Vivès, R.R.; Lortat-Jacob, H. Heparan Sulfate Proteoglycans Biosynthesis and Post Synthesis Mechanisms Combine Few Enzymes and Few Core Proteins to Generate Extensive Structural and Functional Diversity. Molecules 2020, 25, 4215. [Google Scholar] [CrossRef]
- Charbonnier, F.; Périn, J.P.; Roussel, G.; Nussbaum, J.L.; Alliel, P.M. Cloning of testican/SPOCK in man and mouse. Neuromuscular expression perspectives in pathology. Comptes Rendus Seances Soc. Biol. Ses Fil. 1997, 191, 127–133. [Google Scholar]
- Edgell, C.J.; BaSalamah, M.A.; Marr, H.S. Testican-1: A differentially expressed proteoglycan with protease inhibiting activities. Int. Rev. Cytol. 2004, 236, 101–122. [Google Scholar] [CrossRef]
- Chen, D.; Zhou, H.; Liu, G.; Zhao, Y.; Cao, G.; Liu, Q. SPOCK1 promotes the invasion and metastasis of gastric cancer through Slug-induced epithelial-mesenchymal transition. J. Cell. Mol. Med. 2018, 22, 797–807. [Google Scholar] [CrossRef]
- Shu, Y.-J.; Weng, H.; Ye, Y.-Y.; Hu, Y.-P.; Bao, R.-F.; Cao, Y.; Wang, X.-A.; Zhang, F.; Xiang, S.-S.; Li, H.-F.; et al. SPOCK1 as a potential cancer prognostic marker promotes the proliferation and metastasis of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol. Cancer 2015, 14, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Rong, Y.; Gao, X. Multiomic analysis of the function of SPOCK1 across cancers: An integrated bioinformatics approach. J. Int. Med. Res. 2021, 49, 300060520962659. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, V.L.; Damhofer, H.; Waasdorp, C.; Steins, A.; Kocher, H.M.; Medema, J.P.; van Laarhoven, H.W.; Bijlsma, M.F. Stromal SPOCK1 supports invasive pancreatic cancer growth. Mol. Oncol. 2017, 11, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.C.; Jeng, Y.M.; Lu, Y.T.; Lien, H.C. SPOCK1 Is a Novel Transforming Growth Factor-β-Induced Myoepithelial Marker That Enhances Invasion and Correlates with Poor Prognosis in Breast Cancer. PLoS ONE 2016, 11, e0162933. [Google Scholar] [CrossRef] [Green Version]
- Váncza, L.; Karászi, K.; Péterfia, B.; Turiák, L.; Dezső, K.; Sebestyén, A.; Reszegi, A.; Petővári, G.; Kiss, A.; Schaff, Z.; et al. SPOCK1 Promotes the Development of Hepatocellular Carcinoma. Front. Oncol. 2022, 12, 819883. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, W.; Chang, H.H.; Kim, S.W.; Kim, J.; Bae, J.S. Testican-1, as a novel diagnosis of sepsis. J. Cell. Biochem. 2018, 119, 4216–4223. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Wang, Y.; Zhang, L. Effects of shRNA-mediated knockdown of SPOCK1 on ovarian cancer growth and metastasis. Cell. Mol. Biol. 2015, 61, 102–109. [Google Scholar]
- Lanczky, A.; Gyorffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Bradshaw, A.D. Diverse biological functions of the SPARC family of proteins. Int. J. Biochem. Cell Biol. 2012, 44, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Zhang, Z.; Tang, J.; Tao, H.; Zhang, Z. Correlation between SPARC, TGFβ1, Endoglin and angiogenesis mechanism in lung cancer. J. Biol. Regul. Homeost. Agents 2018, 32, 1525–1531. [Google Scholar]
- Rivera, L.B.; Brekken, R.A. SPARC promotes pericyte recruitment via inhibition of endoglin-dependent TGF-β1 activity. J. Cell Biol. 2011, 193, 1305–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Váncza, L.; Tátrai, P.; Reszegi, A.; Baghy, K.; Kovalszky, I. SPOCK1 with unexpected function. The start of a new career. Am. J. Physiol. Cell Physiol. 2022, 322, C688–C693. [Google Scholar] [CrossRef] [PubMed]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. J. Int. Soc. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yao, Y.T.; Xu, H.; Chen, Y.B.; Gu, M.; Cai, Z.K.; Wang, Z. SPOCK1 promotes tumor growth and metastasis in human prostate cancer. Drug Des. Dev. Ther. 2016, 10, 2311–2321. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ke, J.; Fang, J.; Chen, J.P. A potential prognostic marker and therapeutic target: SPOCK1 promotes the proliferation, metastasis, and apoptosis of pancreatic ductal adenocarcinoma cells. J. Cell. Biochem. 2020, 121, 743–754. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Chan, T.H.; Liu, M.; Kong, K.L.; Qiu, J.L.; Li, Y.; Yuan, Y.F.; Guan, X.Y. SPOCK1 is regulated by CHD1L and blocks apoptosis and promotes HCC cell invasiveness and metastasis in mice. Gastroenterology 2013, 144, 179–191.e174. [Google Scholar] [CrossRef]
- Liu, H.X.; Cao, Y.Y.; Qu, J.Y. SPOCK1 promotes the proliferation and migration of colon cancer cells by regulating the NF-κB pathway and inducing EMT. Neoplasma 2021, 68, 702–710. [Google Scholar] [CrossRef]
- Wang, T.; Liu, X.; Tian, Q.; Liang, T.; Chang, P. Reduced SPOCK1 expression inhibits non-small cell lung cancer cell proliferation and migration through Wnt/β-catenin signaling. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 637–644. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, X.; Zhang, S.; Piao, J.; Yang, Y.; Wang, X.; Lin, Z. SPOCK1/SIX1axis promotes breast cancer progression by activating AKT/mTOR signaling. Aging 2020, 13, 1032–1050. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Q.; Yu, J.; Li, X.; Yu, S.; Zhang, X. SPOCK1 promotes the proliferation, migration and invasion of glioma cells through PI3K/AKT and Wnt/β-catenin signaling pathways. Oncol. Rep. 2016, 35, 3566–3576. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhi, X.; Shi, S.; Tao, R.; Chen, P.; Sun, S.; Bian, L.; Xu, Z.; Ma, L. SPOCK1 is up-regulated and promotes tumor growth via the PI3K/AKT signaling pathway in colorectal cancer. Biochem. Biophys. Res. Commun. 2017, 482, 870–876. [Google Scholar] [CrossRef]
- Zhao, P.; Guan, H.T.; Dai, Z.J.; Ma, Y.G.; Liu, X.X.; Wang, X.J. Knockdown of SPOCK1 Inhibits the Proliferation and Invasion in Colorectal Cancer Cells by Suppressing the PI3K/Akt Pathway. Oncol. Res. 2016, 24, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Qian, Y.; Chen, X. Examination of the expanding pathways for the regulation of p21 expression and activity. Cell. Signal. 2010, 22, 1003–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.P.; Liao, Y.; Xia, W.; Spohn, B.; Lee, M.H.; Hung, M.C. Cytoplasmic localization of p21Cip1/WAF1 by Akt-induced phosphorylation in HER-2/neu-overexpressing cells. Nat. Cell Biol. 2001, 3, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.R.; Li, S.Y.; Guo, Q.S.; Zhou, W.; Zhang, H.M. SPOCK1 Involvement in Epithelial-to-Mesenchymal Transition: A New Target in Cancer Therapy? Cancer Manag. Res. 2020, 12, 3561–3569. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Lin, Z.; Wang, Z.; Liu, D.; Tian, D.; Xia, L. SPOCK1 overexpression induced by platelet-derived growth factor-BB promotes hepatic stellate cell activation and liver fibrosis through the integrin α5β1/PI3K/Akt signaling pathway. Lab. Investig. J. Tech. Methods Pathol. 2020, 100, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Koster, R.; di Pietro, A.; Timmer-Bosscha, H.; Gibcus, J.H.; van den Berg, A.; Suurmeijer, A.J.; Bischoff, R.; Gietema, J.A.; de Jong, S. Cytoplasmic p21 expression levels determine cisplatin resistance in human testicular cancer. J. Clin. Investig. 2010, 120, 3594–3605. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Ma, Q.; Li, X.; Ji, T.; Chen, P.; Xu, H.; Li, K.; Fang, Y.; Weng, D.; Weng, Y.; et al. Cytoplasmic p21 is a potential predictor for cisplatin sensitivity in ovarian cancer. BMC Cancer 2011, 11, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgakilas, A.G.; Martin, O.A.; Bonner, W.M. p21: A Two-Faced Genome Guardian. Trends Mol. Med. 2017, 23, 310–319. [Google Scholar] [CrossRef]
- Qu, Y.L.; Liu, X.L.; Zhao, S.Y.; Zhai, X.F. SPOCK1 silencing decreases 5-FU resistance through PRRX1 in colorectal cancer. Pathol. Res. Pract. 2022, 234, 153895. [Google Scholar] [CrossRef]
- Babaei-Jadidi, R.; Kashfi, H.; Alelwani, W.; Karimi Bakhtiari, A.; Kattan, S.W.; Mansouri, O.A.; Mukherjee, A.; Lobo, D.N.; Nateri, A.S. Anti-miR-135/SPOCK1 axis antagonizes the influence of metabolism on drug response in intestinal/colon tumour organoids. Oncogenesis 2022, 11, 4. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, M.; Ma, M.; Zhuang, Y.; Qiu, X.; Zhao, Q.; Dai, J.; Cai, H.; Yan, X. SPOCK1 contributes to the third-generation EGFR tyrosine kinase inhibitors resistance in lung cancer. J. Cell. Biochem. 2019, 120, 12566–12573. [Google Scholar] [CrossRef]
- Yu, F.; Li, G.; Gao, J.; Sun, Y.; Liu, P.; Gao, H.; Li, P.; Lei, T.; Chen, Y.; Cheng, Y.; et al. SPOCK1 is upregulated in recurrent glioblastoma and contributes to metastasis and Temozolomide resistance. Cell Prolif. 2016, 49, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.P.; Han, S.W.; Song, S.H.; Jeong, E.G.; Lee, M.Y.; Hwang, D.; Im, S.A.; Bang, Y.J.; Kim, T.Y. Testican-1-mediated epithelial-mesenchymal transition signaling confers acquired resistance to lapatinib in HER2-positive gastric cancer. Oncogene 2014, 33, 3334–3341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Su, Y.; Xu, F. CHD1L: A novel oncogene. Mol. Cancer 2013, 12, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.P.; Zhou, J.; Cai, M.Y.; Xiao, X.S.; Liao, Y.J.; Kung, H.F.; Guan, X.Y.; Xie, D.; Yang, G.F. CHD1L protein is overexpressed in human ovarian carcinomas and is a novel predictive biomarker for patients survival. BMC Cancer 2012, 12, 437. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Ma, Z.; Zhou, Q.; Wang, S.; Yan, Q.; Zhuang, H.; Zhou, Z.; Liu, C.; Wu, Z.; Zhao, J.; et al. Identification by genetic algorithm optimized back propagation artificial neural network and validation of a four-gene signature for diagnosis and prognosis of pancreatic cancer. Heliyon 2022, 8, e11321. [Google Scholar] [CrossRef]
- Li, F.; Song, Q.Z.; Zhang, Y.F.; Wang, X.R.; Cao, L.M.; Li, N.; Zhao, L.X.; Zhang, S.X.; Zhuang, X.F. Identifying the EMT-related signature to stratify prognosis and evaluate the tumor microenvironment in lung adenocarcinoma. Front. Genet. 2022, 13, 1008416. [Google Scholar] [CrossRef]
- Chen, J.; Ye, Z.; Liu, L.; Xuan, B. Assessment of the prognostic value of SPOCK1 in clear cell renal cell carcinoma: A bioinformatics analysis. Transl. Androl. Urol. 2022, 11, 509–518. [Google Scholar] [CrossRef]
- Chen, M.L.; Ho, C.J.; Yeh, C.M.; Chen, S.L.; Sung, W.W.; Wang, S.C.; Chen, C.J. High SPOCK1 Expression is Associated with Advanced Stage, T Value, and Gleason Grade in Prostate Cancer. Medicina 2019, 55, 343. [Google Scholar] [CrossRef] [Green Version]
- Chien, M.H.; Lin, Y.W.; Wen, Y.C.; Yang, Y.C.; Hsiao, M.; Chang, J.L.; Huang, H.C.; Lee, W.J. Targeting the SPOCK1-snail/slug axis-mediated epithelial-to-mesenchymal transition by apigenin contributes to repression of prostate cancer metastasis. J. Exp. Clin. Cancer Res. 2019, 38, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Váncza, L.; Horváth, A.; Seungyeon, L.; Rókusz, A.; Dezső, K.; Reszegi, A.; Petővári, G.; Götte, M.; Kovalszky, I.; Baghy, K. SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer. Cancers 2023, 15, 2037. https://doi.org/10.3390/cancers15072037
Váncza L, Horváth A, Seungyeon L, Rókusz A, Dezső K, Reszegi A, Petővári G, Götte M, Kovalszky I, Baghy K. SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer. Cancers. 2023; 15(7):2037. https://doi.org/10.3390/cancers15072037
Chicago/Turabian StyleVáncza, Lóránd, Anna Horváth, Lee Seungyeon, András Rókusz, Katalin Dezső, Andrea Reszegi, Gábor Petővári, Martin Götte, Ilona Kovalszky, and Kornélia Baghy. 2023. "SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer" Cancers 15, no. 7: 2037. https://doi.org/10.3390/cancers15072037
APA StyleVáncza, L., Horváth, A., Seungyeon, L., Rókusz, A., Dezső, K., Reszegi, A., Petővári, G., Götte, M., Kovalszky, I., & Baghy, K. (2023). SPOCK1 Overexpression Suggests Poor Prognosis of Ovarian Cancer. Cancers, 15(7), 2037. https://doi.org/10.3390/cancers15072037