

Cell-Free RNA from Plasma in Patients with Neuroblastoma: Exploring the Technical and Clinical Potential

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Patients and Samples
2.2. Preparation of Platelets from Peripheral Blood
2.3. Isolation of Cell-Free RNA and cDNA Synthesis
2.4. Design and Optimization of the Multiplex ddPCR Assays
2.5. Isolation of Cell-Free DNA and ddPCR Assays
2.6. Isolation of EVs from Plasma and Electron Microscopy on EVs
2.7. Western Blot
2.8. Statistical Analysis
3. Results
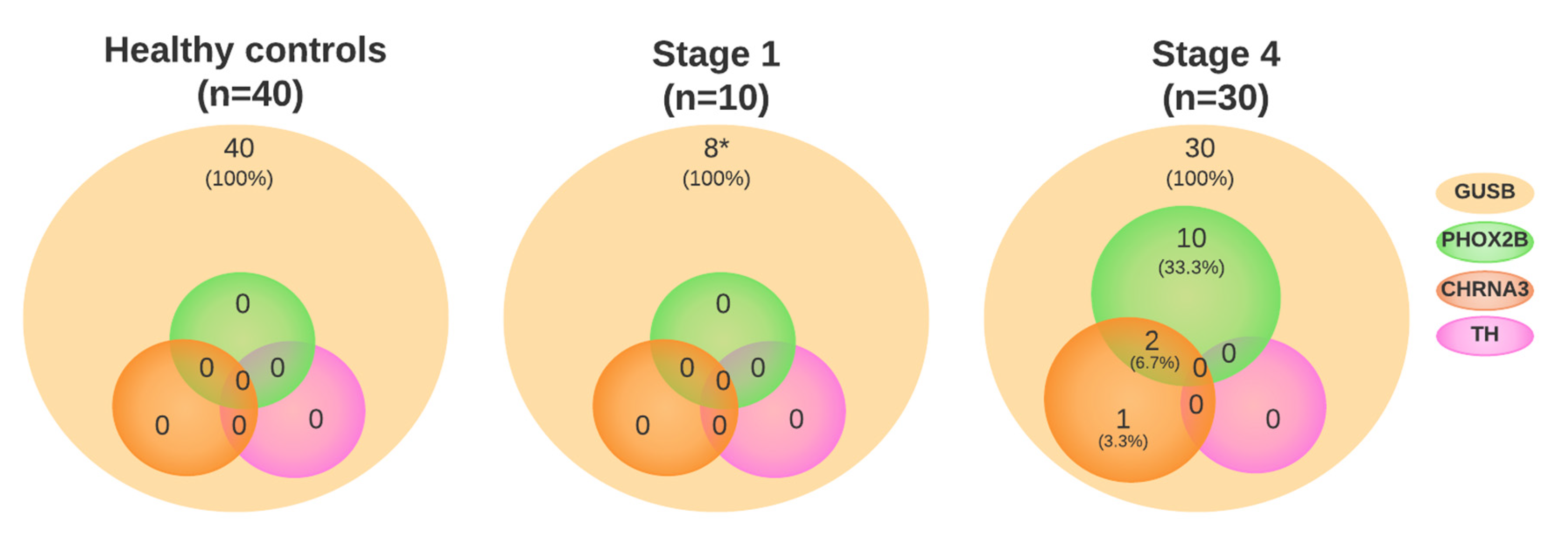
3.1. Neuroblastoma-Specific mRNA Is Present in Plasma
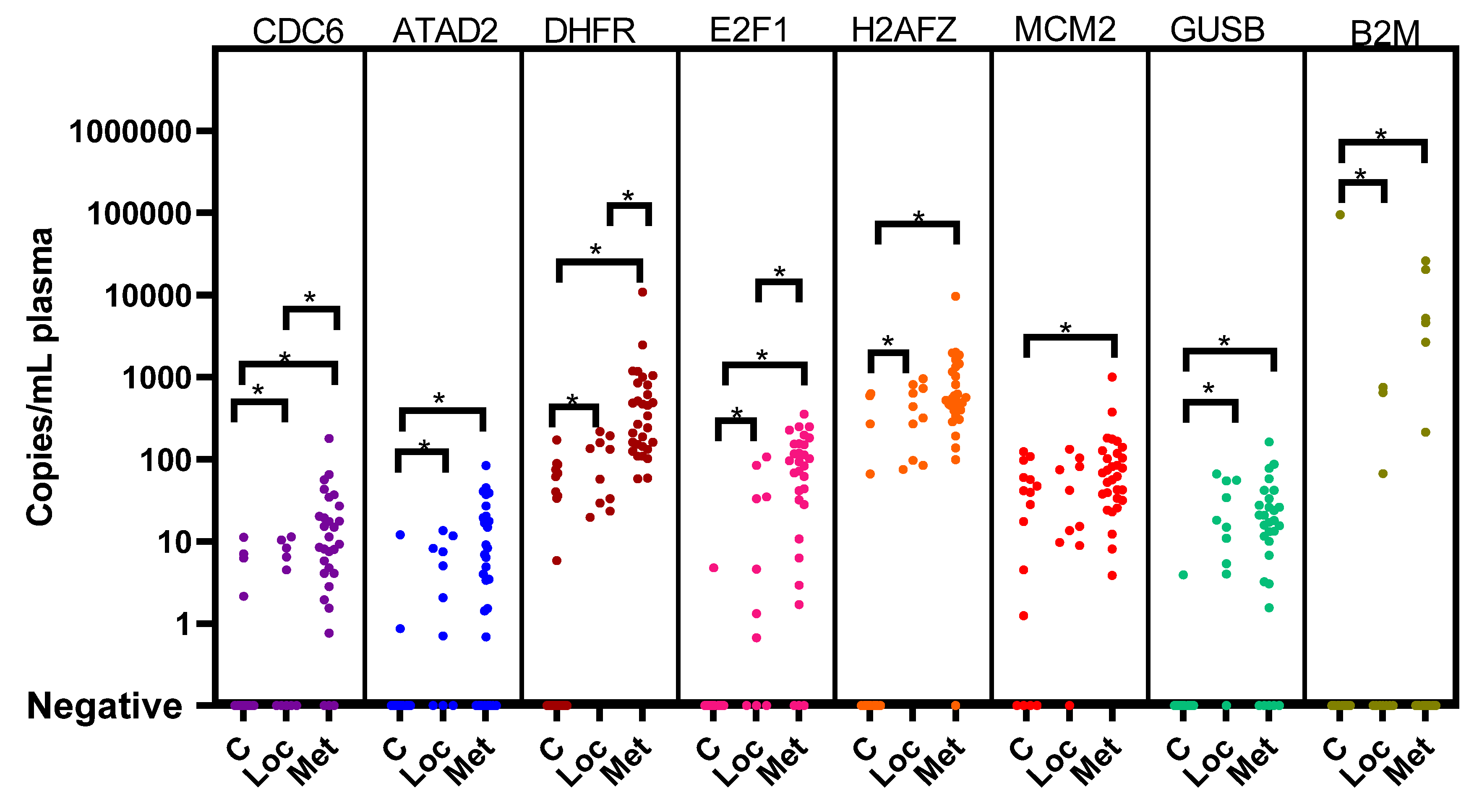
3.2. Cell Cycle Genes in Plasma and Correction for the Presence of Platelets
3.3. Cell Cycle Genes in Plasma from Patients at Diagnosis
3.4. cfRNA during Treatment
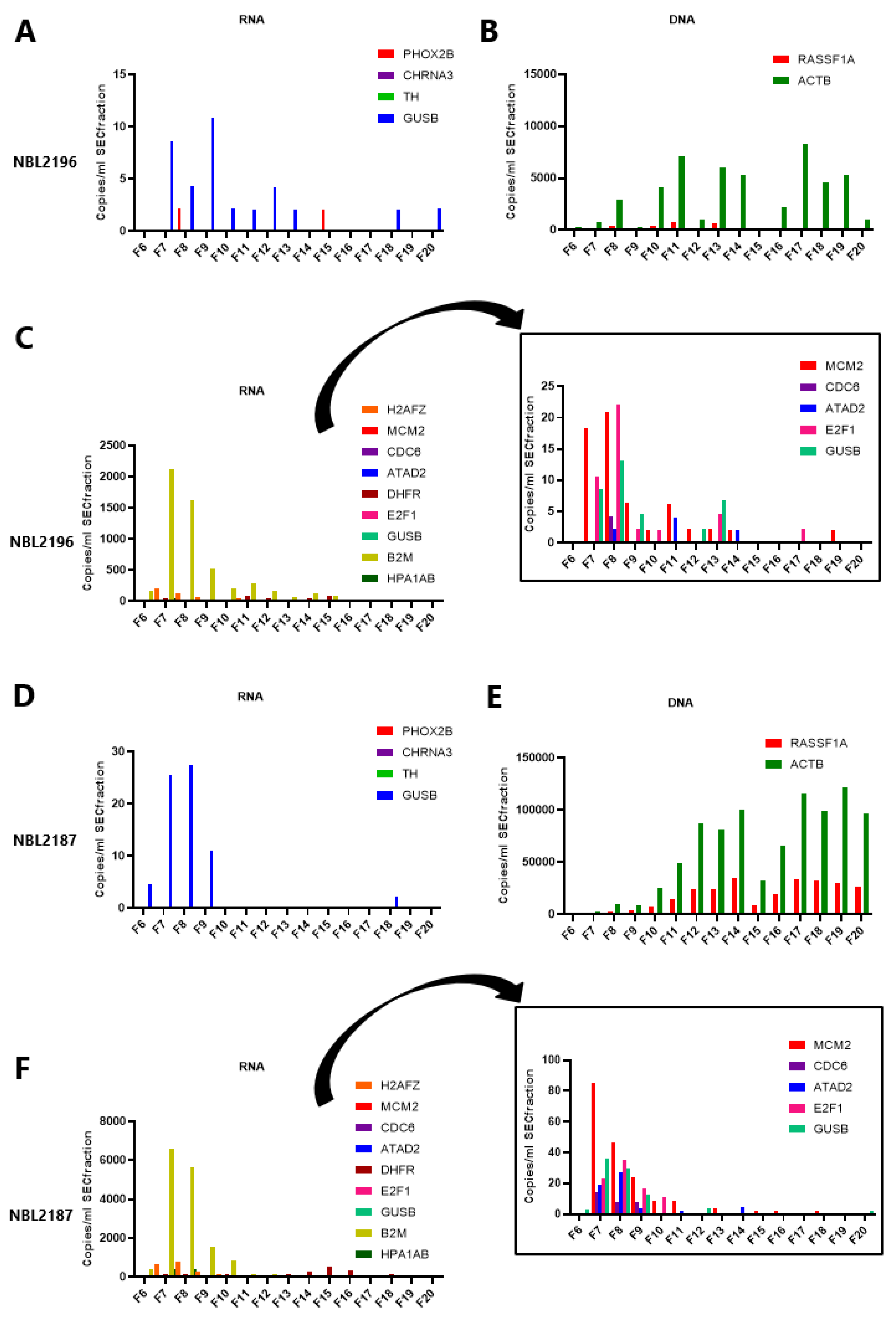
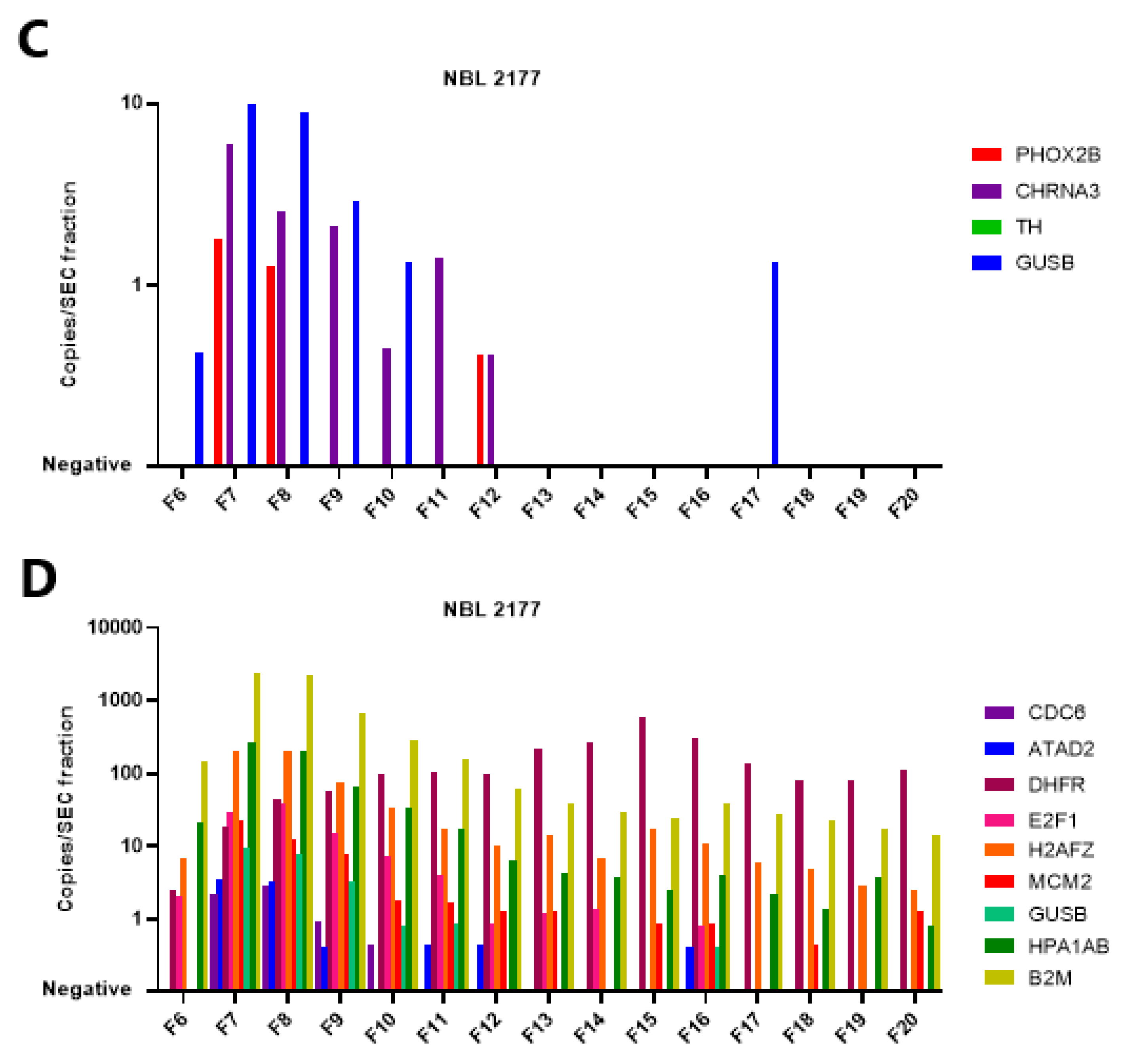
3.5. The mRNA in Plasma Is Concentrated in EV-Enriched SEC Fractions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Pearson, A.D.; Pinkerton, C.R.; Lewis, I.J.; Imeson, J.; Ellershaw, C.; Machin, D. High-dose rapid and standard induction chemotherapy for patients aged over 1 year with stage 4 neuroblastoma: A randomised trial. Lancet Oncol. 2008, 9, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Kreissman, S.G.; Seeger, R.C.; Matthay, K.K.; London, W.B.; Sposto, R.; A Grupp, S.; A Haas-Kogan, D.; LaQuaglia, M.P.; Yu, A.L.; Diller, L.; et al. Purged versus non-purged peripheral blood stem-cell transplantation for high-risk neuroblastoma (COG A3973): A randomised phase 3 trial. Lancet Oncol. 2013, 14, 999–1008. [Google Scholar] [CrossRef] [Green Version]
- Van Paemel, R.; Vlug, R.; De Preter, K.; Van Roy, N.; Speleman, F.; Willems, L.; Lammens, T.; Laureys, G.; Schleiermacher, G.; Tytgat, G.A.M.; et al. The pitfalls and promise of liquid biopsies for diagnosing and treating solid tumors in children: A review. Eur. J. Pediatr. 2020, 179, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Alix-Panabières, C.; Pantel, K. Clinical Applications of Circulating Tumor Cells and Circulating Tumor DNA as Liquid Biopsy. Cancer Discov. 2016, 6, 479–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stutterheim, J.; Gerritsen, A.; Zappeij-Kannegieter, L.; Kleijn, I.; Dee, R.; Hooft, L.; van Noesel, M.M.; Bierings, M.; Berthold, F.; Versteeg, R.; et al. PHOX2B Is a Novel and Specific Marker for Minimal Residual Disease Testing in Neuroblastoma. J. Clin. Oncol. 2008, 26, 5443–5449. [Google Scholar] [CrossRef]
- Yáñez, Y.; Hervás, D.; Grau, E.; Oltra, S.; Pérez, G.; Palanca, S.; Bermúdez, M.; Márquez, C.; Cañete, A.; Castel, V. TH and DCX mRNAs in peripheral blood and bone marrow predict outcome in metastatic neuroblastoma patients. J. Cancer Res. Clin. Oncol. 2016, 142, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Van Wezel, E.M.; van Zogchel, L.M.J.; van Wijk, J.; Timmerman, I.; Vo, N.K.; Zappeij-Kannegieter, L.; deCarolis, B.; Simon, T.; van Noesel, M.M.; Molenaar, J.J.; et al. Mesenchymal Neuroblastoma Cells Are Undetected by Current mRNA Marker Panels: The Development of a Specific Neuroblastoma Mesenchymal Minimal Residual Disease Panel. JCO Precis. Oncol. 2019, 3, 1–11. [Google Scholar] [CrossRef]
- Stutterheim, J.; Gerritsen, A.; Zappeij-Kannegieter, L.; Yalcin, B.; Dee, R.; van Noesel, M.M.; Berthold, F.; Versteeg, R.; Caron, H.N.; van der Schoot, C.E.; et al. Detecting Minimal Residual Disease in Neuroblastoma: The Superiority of a Panel of Real-Time Quantitative PCR Markers. Clin. Chem. 2009, 55, 1316–1326. [Google Scholar] [CrossRef] [Green Version]
- Van Zogchel, L.M.J.; Lak, N.S.M.; Verhagen, O.J.H.M.; Tissoudali, A.; Nuru, M.G.; Gelineau, N.U.; Zappeij-Kannengieter, L.; Javadi, A.; Zijtregtop, E.A.M.; Merks, J.H.M.; et al. Novel Circulating Hypermethylated RASSF1A ddPCR for Liquid Biopsies in Patients With Pediatric Solid Tumors. JCO Precis. Oncol. 2021, 5, 1738–1748. [Google Scholar] [CrossRef]
- Lodrini, M.; Sprüssel, A.; Astrahantseff, K.; Tiburtius, D.; Konschak, R.; Lode, H.N.; Fischer, M.; Keilholz, U.; Eggert, A.; Deubzer, H.E. Using droplet digital PCR to analyze MYCN and ALK copy number in plasma from patients with neuroblastoma. Oncotarget 2017, 8, 85234–85251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodrini, M.; Graef, J.; Thole-Kliesch, T.M.; Astrahantseff, K.; Sprüssel, A.; Grimaldi, M.; Peitz, C.; Linke, R.B.; Hollander, J.F.; Lankes, E.; et al. Targeted Analysis of Cell-free Circulating Tumor DNA is Suitable for Early Relapse and Actionable Target Detection in Patients with Neuroblastoma. Clin. Cancer Res. 2022, 28, 1809–1820. [Google Scholar] [CrossRef] [PubMed]
- Van Zogchel, L.M.J.; van Wezel, E.M.; van Wijk, J.; Stutterheim, J.; Bruins, W.S.C.; Zappeij-Kannegieter, L.; Tirza, J.E.; Iedan, R.N.; Verly, C.; Godelieve, A.M.; et al. Hypermethylated RASSF1A as Circulating Tumor DNA Marker for Disease Monitoring in Neuroblastoma. JCO Precis. Oncol. 2020, 4, 291–306. [Google Scholar] [CrossRef]
- Wan, J.C.M.; Massie, C.; Garcia-Corbacho, J.; Mouliere, F.; Brenton, J.D.; Caldas, C.; Pacey, S.; Baird, R.; Rosenfeld, N. Liquid biopsies come of age: Towards implementation of circulating tumour DNA. Nat. Rev. Cancer 2017, 17, 223–238. [Google Scholar] [CrossRef]
- Skog, J.; Wurdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T., Jr.; Sena-Esteves, M.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Hulstaert, E.; Morlion, A.; Avila Cobos, F.; Verniers, K.; Nuytens, J.; Vanden Eynde, E.; Yigit, N.; Anckaert, J.; Geerts, A.; Hindryckx, P.; et al. Charting Extracellular Transcriptomes in The Human Biofluid RNA Atlas. Cell Rep. 2020, 33, 108552. [Google Scholar] [CrossRef] [PubMed]
- Herriott, R.M.; Connolly, J.H.; Gupta, S. Blood Nucleases and Infectious Viral Nucleic Acids. Nature 1961, 189, 817–820. [Google Scholar] [CrossRef]
- Bebelman, M.P.; Smit, M.J.; Pegtel, D.M.; Baglio, S.R. Biogenesis and function of extracellular vesicles in cancer. Pharmacol. Ther. 2018, 188, 1–11. [Google Scholar] [CrossRef]
- Xavier, C.P.R.; Caires, H.R.; Barbosa, M.A.G.; Bergantim, R.; Guimarães, J.E.; Vasconcelos, M.H. The Role of Extracellular Vesicles in the Hallmarks of Cancer and Drug Resistance. Cells 2020, 9, 1141. [Google Scholar] [CrossRef]
- Grölz, D.; Hauch, S.; Schlumpberger, M.; Guenther, K.; Voss, T.; Sprenger-Haussels, M.; Oelmüller, U. Liquid Biopsy Preservation Solutions for Standardized Pre-Analytical Workflows—Venous Whole Blood and Plasma. Curr. Pathobiol. Rep. 2018, 6, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurdinger, T.; In ‘t Veld, S.G.J.G.; Best, M.G. Platelet RNA as Pan-Tumor Biomarker for Cancer Detection. Cancer Res. 2020, 80, 1371–1373. [Google Scholar] [CrossRef] [Green Version]
- Rowley, J.W.; Oler, A.J.; Tolley, N.D.; Hunter, B.N.; Low, E.N.; Nix, D.A.; Yost, C.C.; Zimmerman, G.A.; Weyrich, A. Genome-wide RNA-seq analysis of human and mouse platelet transcriptomes. Blood 2011, 118, e101–e111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowley, J.W.; Schwertz, H.; Weyrich, A.S. Platelet mRNA. Curr. Opin. Hematol. 2012, 19, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Morini, M.; Cangelosi, D.; Segalerba, D.; Marimpietri, D.; Raggi, F.; Castellano, A.; Fruci, D.; de Mora, J.F.; Cañete, A.; Yáñez, Y.; et al. Exosomal microRNAs from Longitudinal Liquid Biopsies for the Prediction of Response to Induction Chemotherapy in High-Risk Neuroblastoma Patients: A Proof of Concept SIOPEN Study II. Cancers 2019, 11, 1476. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Xu, M.; Yin, M.; Hong, J.; Chen, H.; Gao, Y.; Xie, C.; Shen, N.; Gu, S.; Mo, X. Exosomal hsa-miR199a-3p Promotes Proliferation and Migration in Neuroblastoma. Front. Oncol. 2019, 9, 459. [Google Scholar] [CrossRef]
- Larson, M.H.; Pan, W.; Kim, H.J.; Mauntz, R.E.; Stuart, S.M.; Pimentel, M.; Zhou, Y.; Knudsgaard, P.; Demas, V.; Aravanis, A.M.; et al. A comprehensive characterization of the cell-free transcriptome reveals tissue- and subtype-specific biomarkers for cancer detection. Nat. Commun. 2021, 12, 2357. [Google Scholar] [CrossRef]
- Harashima, H.; Dissmeyer, N.; Schnittger, A. Cell cycle control across the eukaryotic kingdom. Trends Cell Biol. 2013, 23, 345–356. [Google Scholar] [CrossRef]
- Matthews, H.K.; Bertoli, C.; de Bruin, R.A.M. Cell cycle control in cancer. Nat. Rev. Mol. Cell Biol. 2021, 23, 74–88. [Google Scholar] [CrossRef]
- Thurlings, I.; de Bruin, A. E2F Transcription Factors Control the Roller Coaster Ride of Cell Cycle Gene Expression. Methods Mol. Biol. 2016, 1342, 71–88. [Google Scholar]
- Bongiovanni, L.; Andriessen, A.; Silvestri, S.; Porcellato, I.; Brachelente, C.; de Bruin, A. H2AFZ: A Novel Prognostic Marker in Canine Melanoma and a Predictive Marker for Resistance to CDK4/6 Inhibitor Treatment. Front. Vet. Sci. 2021, 8, 705359. [Google Scholar] [CrossRef]
- Andriessen, A.; Bongiovanni, L.; Driedonks, T.A.P.; van Liere, E.; Seijger, A.; Hegeman, C.V.; van Nimwegen, S.A.; Galac, S.; Westendorp, B.; Hoen, E.N.M.N.; et al. CDC6: A novel canine tumour biomarker detected in circulating extracellular vesicles. Veter Comp. Oncol. 2021, 20, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Nakagawara, A. Acceleration or Brakes: Which Is Rational for Cell Cycle-Targeting Neuroblastoma Therapy? Biomolecules 2021, 11, 750. [Google Scholar] [CrossRef] [PubMed]
- Van Zogchel, L.; de Carolis, B.; van Wezel, E.; Stutterheim, J.; Zappeij-Kannegieter, L.; van Doornum, M.; Schumacher-Kuckelkorn, R.; Gecht, J.; Simon, T.; Caron, H.; et al. Detection of Minimal Residual Disease (MRD) in High Risk Neuroblastoma Correlates with Outcome: Final Results of International GPOH-DCOG Prospective Validation Study. In Pediatric Blood & Cancer; Wiley: Hoboken, NJ, USA, 2018; p. S38. [Google Scholar]
- Beillard, E.; Pallisgaard, N.; van der Velden, V.H.J.; Bi, W.; Dee, R.; van der Schoot, E.; Delabesse, E.; Macintyre, E.; Gottardi, E.; Saglio, G.; et al. Evaluation of candidate control genes for diagnosis and residual disease detection in leukemic patients using ‘real-time’ quantitative reverse-transcriptase polymerase chain reaction (RQ-PCR)—A Europe against cancer program. Leukemia 2003, 17, 2474–2486. [Google Scholar] [CrossRef] [Green Version]
- Corrias, M.V.; Occhino, M.; Croce, M.; De Ambrosis, A.; Pistillo, M.P.; Bocca, P.; Pistoia, V.; Ferrini, S. Lack of HLA-class I antigens in human neuroblastoma cells: Analysis of its relationship to TAP and tapasin expression. Tissue Antigens 2001, 57, 110–117. [Google Scholar] [CrossRef]
- Spel, L.; Schiepers, A.; Boes, M. NFκB and MHC-1 Interplay in Neuroblastoma and Immunotherapy. Trends Cancer 2018, 4, 715–717. [Google Scholar] [CrossRef]
- Corrias, M.V.; Pistorio, A.; Cangemi, G.; Tripodi, G.; Bs, B.C.; Scaruffi, P.; Fardin, P.; Garaventa, A.; Pistoia, V.; Haupt, R. Detection of cell-free RNA in children with neuroblastoma and comparison with that of whole blood cell RNA. Pediatr. Blood Cancer 2010, 54, 897–903. [Google Scholar] [CrossRef]
- Koppen, A.; Ait-Aissa, R.; Koster, J.; Van Sluis, P.G.; Øra, I.; Caron, H.N.; Volckmann, R.; Versteeg, R.; Valentijn, L.J. Direct regulation of the minichromosome maintenance complex by MYCN in neuroblastoma. Eur. J. Cancer 2007, 43, 2413–2422. [Google Scholar] [CrossRef]
- Liu, X.; Cai, Y.; Cheng, C.; Gu, Y.; Hu, X.; Chen, K.; Wu, Y.; Wu, Z. PCLAF promotes neuroblastoma G1/S cell cycle progression via the E2F1/PTTG1 axis. Cell Death Dis. 2022, 13, 178. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Xu, L.; Zhang, J.; Cao, H. Prognostic significance of MYCN related genes in pediatric neuroblastoma: A study based on TARGET and GEO datasets. BMC Pediatr. 2020, 20, 314. [Google Scholar] [CrossRef]
- Feng, L.; Barnhart, J.R.; Seeger, R.C.; Wu, L.; Keshelava, N.; Huang, S.-H.; Jong, A. Cdc6 knockdown inhibits human neuroblastoma cell proliferation. Mol. Cell. Biochem. 2008, 311, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, K.; Kumar, N.; Gulati, H.K.; Sharma, A.; Kaur, A.; Singh, J.V.; Singh, H.; Bedi, P.M.S. Dihydrofolate reductase inhibitors: Patent landscape and phases of clinical development (2001–2021). Expert Opin. Ther. Patents 2022, 32, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, M.V.; Randazzo, O.; La Franca, M.; Barone, G.; Vignoni, E.; Rossi, D.; Collina, S. DHFR Inhibitors: Reading the Past for Discovering Novel Anticancer Agents. Molecules 2019, 24, 1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.; Boilard, E.; Buzas, E.I.; Cheng, L.; Falcón-Perez, J.M.; Gardiner, C.; Gustafson, D.; Gualerzi, A.; Hendrix, A.; Hoffman, A.; et al. Considerations towards a roadmap for collection, handling and storage of blood extracellular vesicles. J. Extracell. Vesicles 2019, 8, 1647027. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Coumans, F.A.W.; Brisson, A.R.; Buzas, E.I.; Dignat-George, F.; Drees, E.E.E.; El-Andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F.; et al. Methodological Guidelines to Study Extracellular Vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef]
- Liu, H.; Tian, Y.; Xue, C.; Niu, Q.; Chen, C.; Yan, X. Analysis of extracellular vesicle DNA at the single-vesicle level by nano-flow cytometry. J. Extracell. Vesicles 2022, 11, 12206. [Google Scholar] [CrossRef]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [Green Version]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 3, 24858. [Google Scholar] [CrossRef] [Green Version]
- Vergauwen, G.; Tulkens, J.; Pinheiro, C.; Cobos, F.A.; Dedeyne, S.; De Scheerder, M.; Vandekerckhove, L.; Impens, F.; Miinalainen, I.; Braems, G.; et al. Robust sequential biophysical fractionation of blood plasma to study variations in the biomolecular landscape of systemically circulating extracellular vesicles across clinical conditions. J. Extracell. Vesicles 2021, 10, e12122. [Google Scholar] [CrossRef] [PubMed]
- Stegmaier, S.; Sparber-Sauer, M.; Aakcha-Rudel, E.; Münch, P.; Reeh, T.; Feuchtgruber, S.; Hallmen, E.; Blattmann, C.; Bielack, S.; Klingebiel, T.; et al. Fusion transcripts as liquid biopsy markers in alveolar rhabdomyosarcoma and synovial sarcoma: A report of the Cooperative Weichteilsarkom Studiengruppe (CWS). Pediatr. Blood Cancer 2022, 69, 29652. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lak, N.S.M.; Seijger, A.; van Zogchel, L.M.J.; Gelineau, N.U.; Javadi, A.; Zappeij-Kannegieter, L.; Bongiovanni, L.; Andriessen, A.; Stutterheim, J.; van der Schoot, C.E.; et al. Cell-Free RNA from Plasma in Patients with Neuroblastoma: Exploring the Technical and Clinical Potential. Cancers 2023, 15, 2108. https://doi.org/10.3390/cancers15072108
Lak NSM, Seijger A, van Zogchel LMJ, Gelineau NU, Javadi A, Zappeij-Kannegieter L, Bongiovanni L, Andriessen A, Stutterheim J, van der Schoot CE, et al. Cell-Free RNA from Plasma in Patients with Neuroblastoma: Exploring the Technical and Clinical Potential. Cancers. 2023; 15(7):2108. https://doi.org/10.3390/cancers15072108
Chicago/Turabian StyleLak, Nathalie S. M., Anne Seijger, Lieke M. J. van Zogchel, Nina U. Gelineau, Ahmad Javadi, Lily Zappeij-Kannegieter, Laura Bongiovanni, Anneloes Andriessen, Janine Stutterheim, C. Ellen van der Schoot, and et al. 2023. "Cell-Free RNA from Plasma in Patients with Neuroblastoma: Exploring the Technical and Clinical Potential" Cancers 15, no. 7: 2108. https://doi.org/10.3390/cancers15072108
APA StyleLak, N. S. M., Seijger, A., van Zogchel, L. M. J., Gelineau, N. U., Javadi, A., Zappeij-Kannegieter, L., Bongiovanni, L., Andriessen, A., Stutterheim, J., van der Schoot, C. E., de Bruin, A., & Tytgat, G. A. M. (2023). Cell-Free RNA from Plasma in Patients with Neuroblastoma: Exploring the Technical and Clinical Potential. Cancers, 15(7), 2108. https://doi.org/10.3390/cancers15072108