

PATZ1 in Non-Small Cell Lung Cancer: A New Biomarker That Negatively Correlates with PD-L1 Expression and Suppresses the Malignant Phenotype




 , , ,
, , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohorts and Public Datasets
2.2. Tissue-Microarray and Immunohistochemical Study
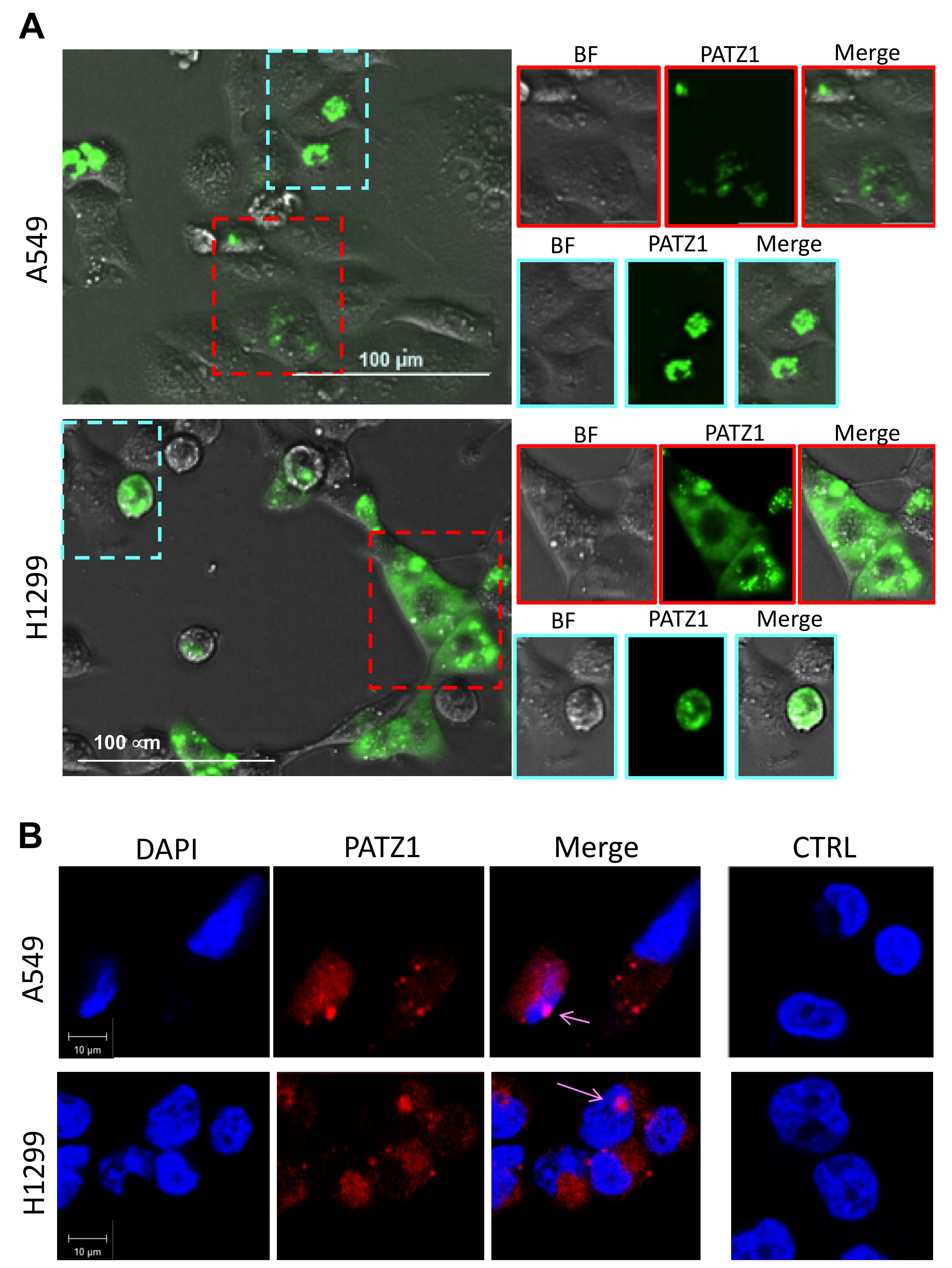
2.3. Cell Culture, Transfections, and PATZ1 Subcellular Localization
2.4. Cell Viability, Tripan Blue Exclusion, and Colony Formation Assays
2.5. Cell Migration and Invasion Assays
2.6. RNA Extraction, Reverse Transcription, and RTqPCR
2.7. Protein Extraction, Western Blotting, and Antibodies
2.8. Statistical Analysis and Kaplan–Meier Survival Curves
2.9. Animals
3. Results
3.1. Tissue Micro-Array Design and Clinicopathological Data
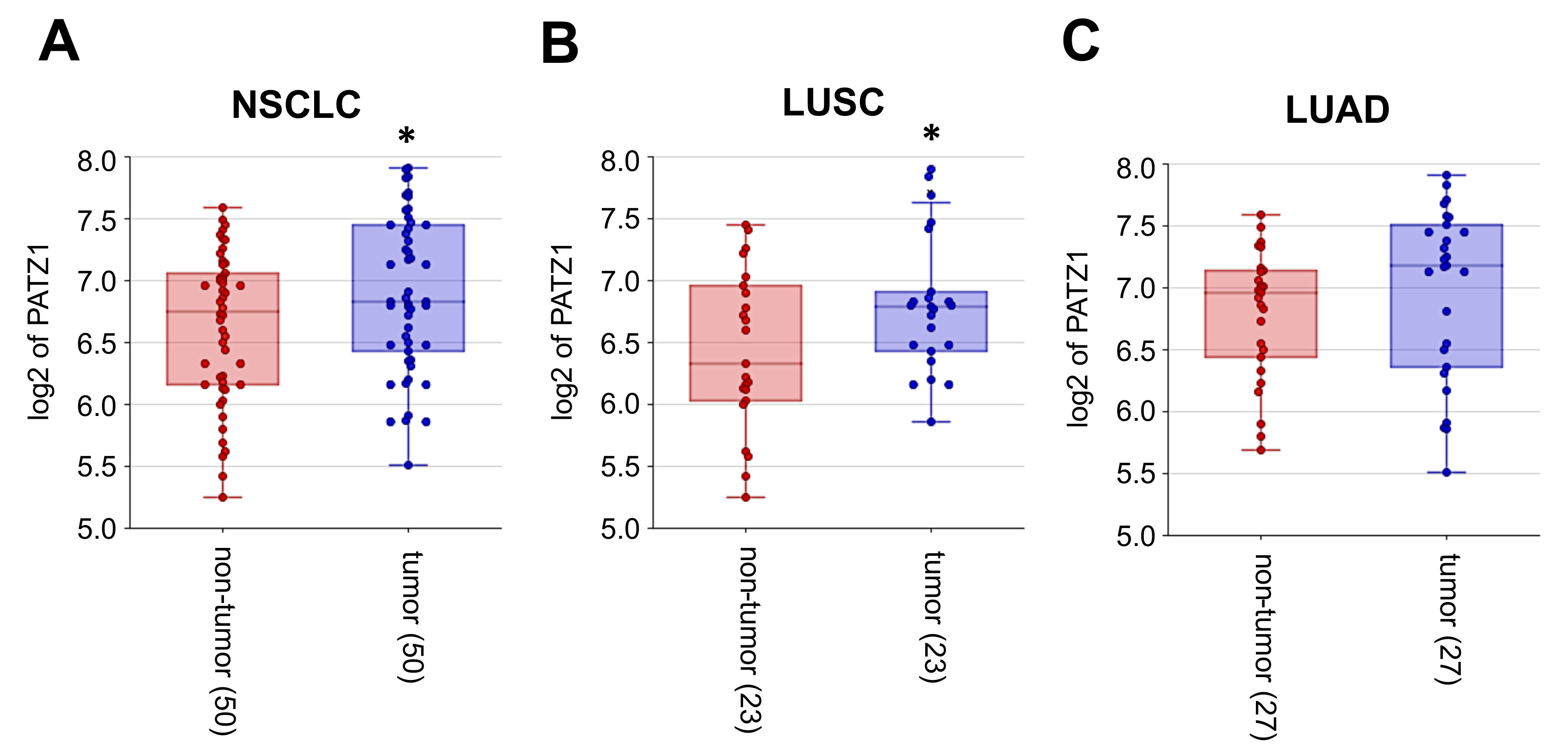
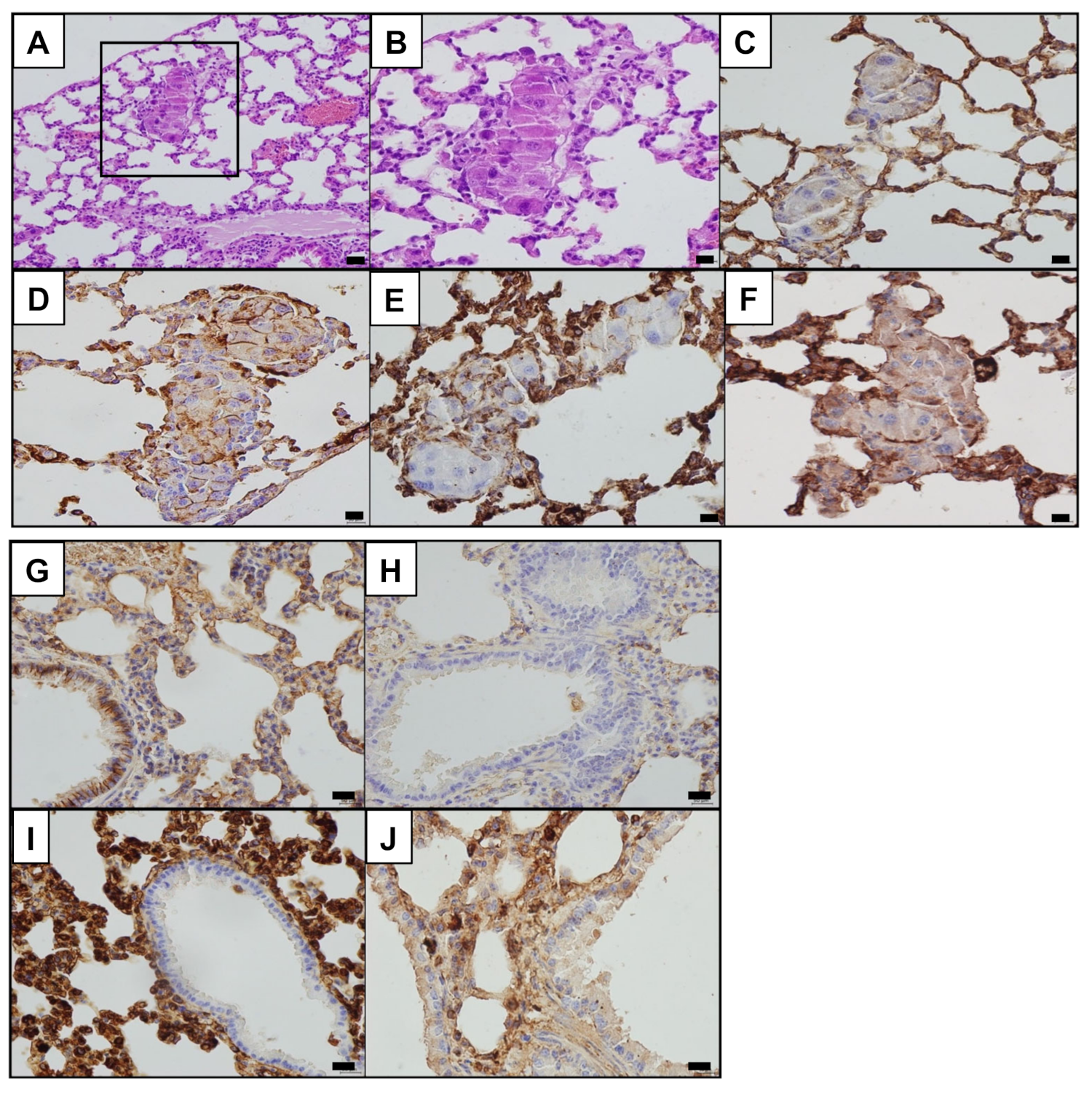
3.2. PATZ1 Is Differently Expressed and Mislocalized in NSCLC
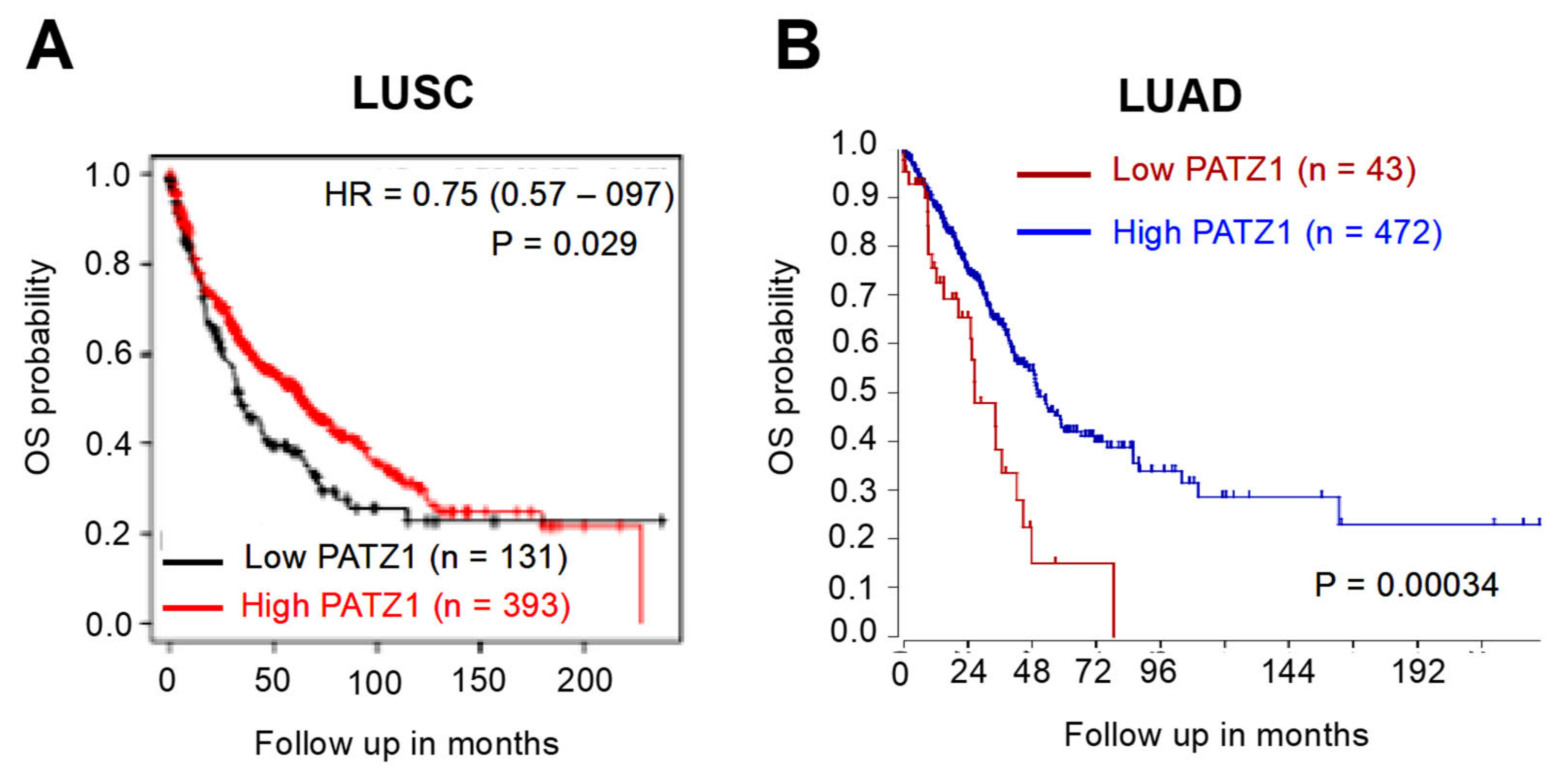
3.3. PATZ1 High Expression Correlates with the LUSC Subtype and Is a Favorable Prognostic Marker
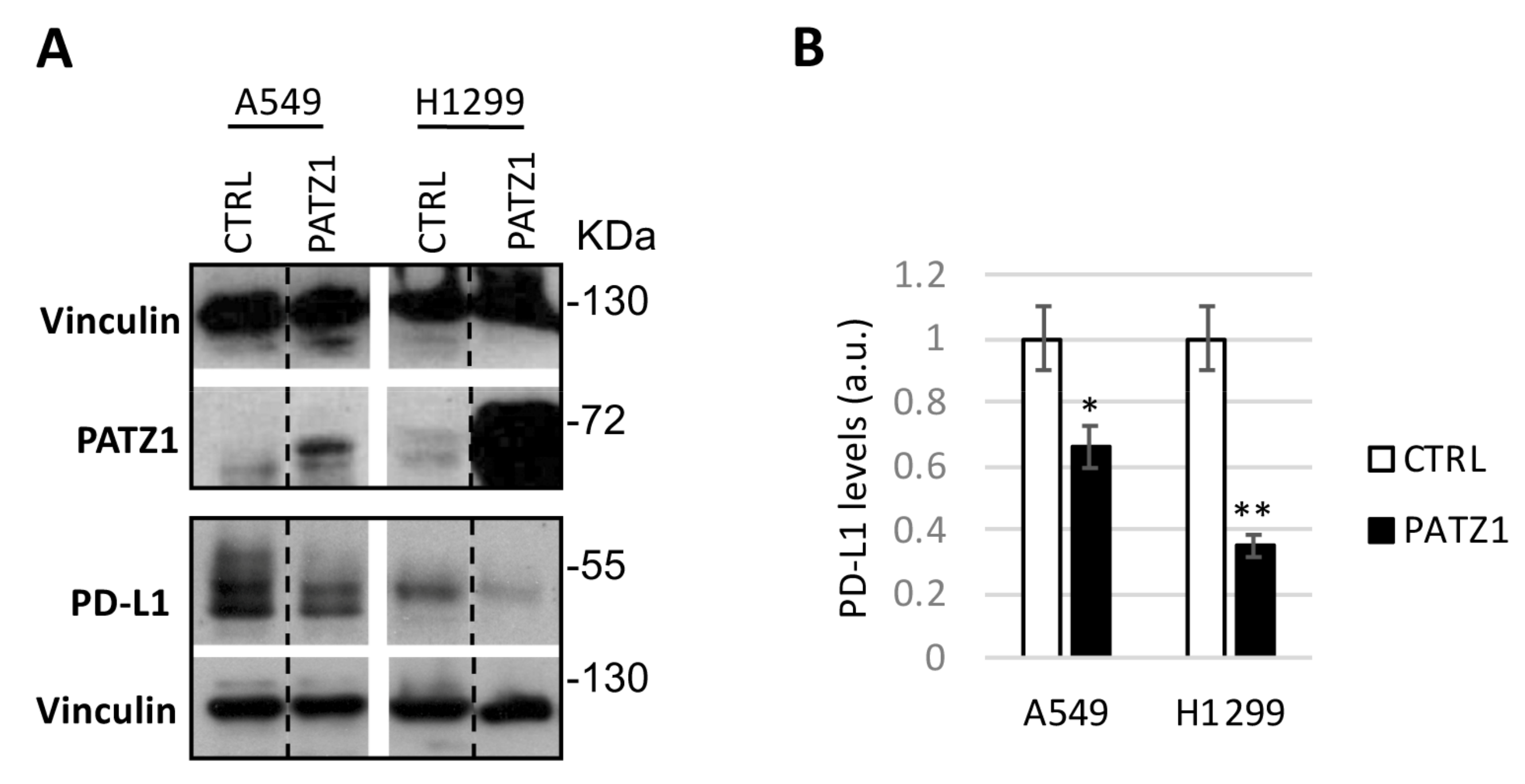
3.4. PATZ1 Expression Negatively Correlates with PD-L1 Expression
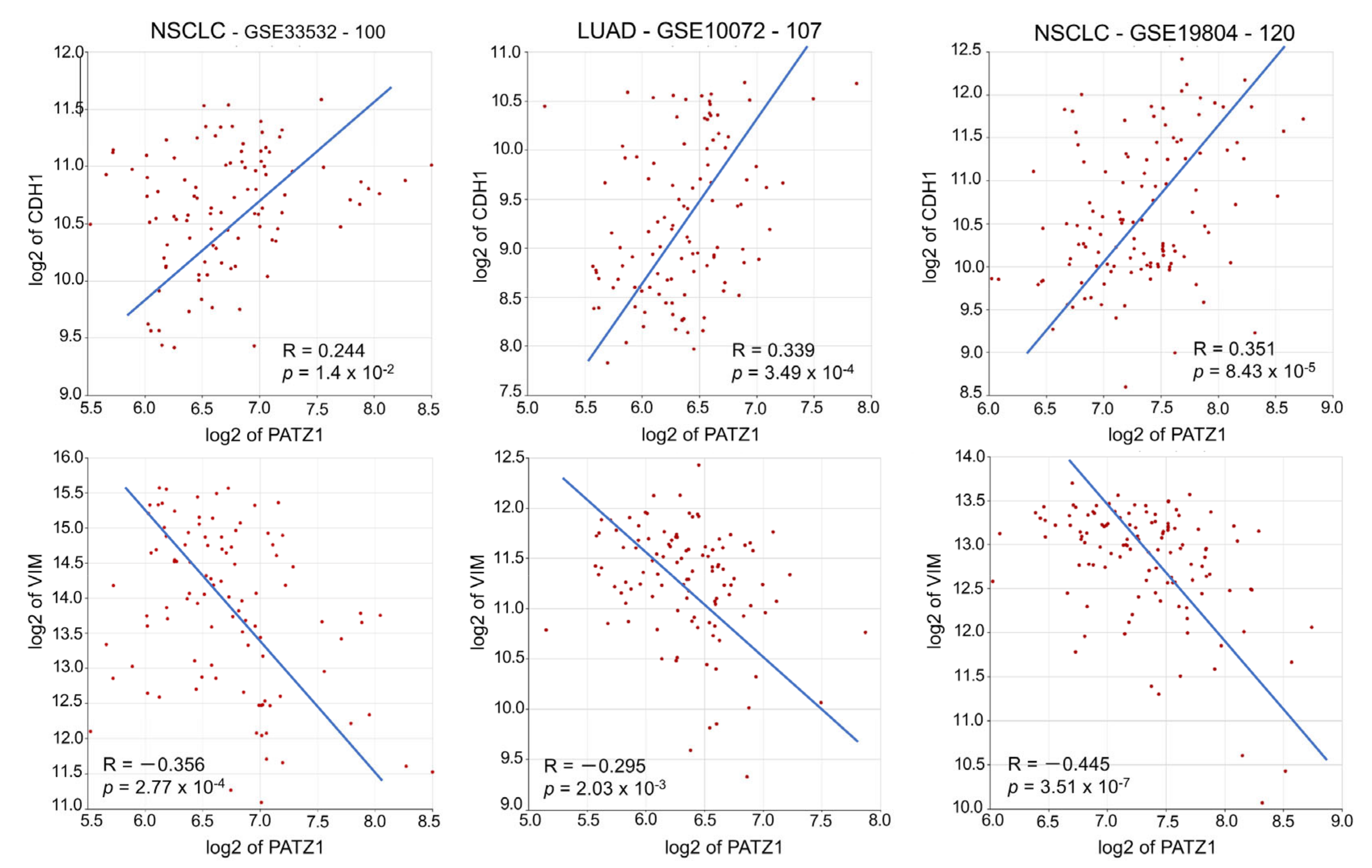
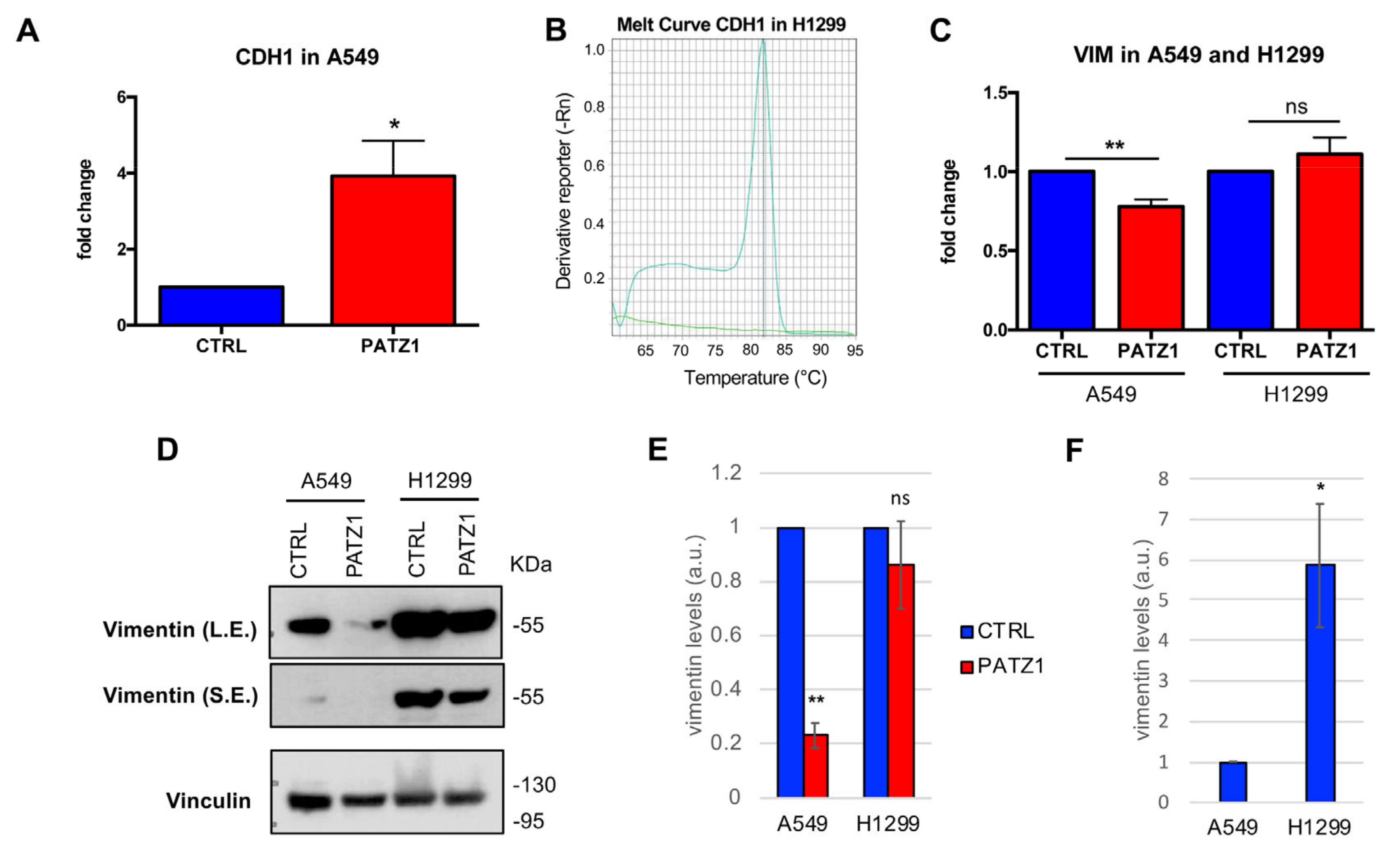
3.5. PATZ1 Expression Attenuated EMT in NSCLC Cells
3.6. PATZ1 Overexpression in A549 and H1299 NSCLC Cell Lines Inhibits Cell Proliferation, Migration, and Invasion
3.7. PATZ1 Knockout Can Induce LUAD in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartorius, B.; Sartorius, K. How much incident lung cancer was missed globally in 2012? An ecological country-level study. Geospat. Health 2016, 11, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, F.R.; Scagliotti, G.V.; Mulshine, J.L.; Kwon, R.; Curran, W.J., Jr.; Wu, Y.L.; Paz-Ares, L. Lung cancer: Current therapies and new targeted treatments. Lancet 2017, 389, 299–311. [Google Scholar] [CrossRef]
- Ullah, A.; Pulliam, S.; Karki, N.R.; Khan, J.; Jogezai, S.; Sultan, S.; Muhammad, L.; Khan, M.; Jamil, N.; Waheed, A.; et al. PD-L1 Over-Expression Varies in Different Subtypes of Lung Cancer: Will This Affect Future Therapies? Clin. Pract. 2022, 12, 653–671. [Google Scholar] [CrossRef]
- Pardoll, D.M. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Han, X. Anti-PD-1/PD-L1 therapy of human cancer: Past, present, and future. J. Clin. Investig. 2015, 125, 3384–3391. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Hua, Y.; Qiu, H.; Hao, J.; Zou, K.; Li, Z.; Hu, S.; Guo, P.; Chen, M.; Sui, S.; et al. PD-L1 promotes tumor growth and progression by activating WIP and β-catenin signaling pathways and predicts poor prognosis in lung cancer. Cell Death Dis. 2020, 11, 506. [Google Scholar] [CrossRef]
- Fedele, M.; Crescenzi, E.; Cerchia, L. The POZ/BTB and AT-Hook Containing Zinc Finger 1 (PATZ1) Transcription Regulator: Physiological Functions and Disease Involvement. Int. J. Mol. Sci. 2017, 18, 2524. [Google Scholar] [CrossRef] [Green Version]
- Valentino, T.; Palmieri, D.; Vitiello, M.; Pierantoni, G.M.; Fusco, A.; Fedele, M. PATZ1 interacts with p53 and regulates expression of p53-target genes enhancing apoptosis or cell survival based on the cellular context. Cell Death Dis. 2013, 4, e963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilic, I.; Koesters, C.; Unger, B.; Sekimata, M.; Hertweck, A.; Maschek, R.; Wilson, C.B.; Ellmeier, W. Negative regulation of CD8 expression via Cd8 enhancer-mediated recruitment of the zinc finger protein MAZR. Nat. Immunol. 2006, 7, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.H.; Kim, M.J.; Kim, K.J.; Kim, J.R. POZ/BTB and AT-hook-containing zinc finger protein 1 (PATZ1) inhibits endothelial cell senescence through a p53 dependent pathway. Cell Death Differ. 2012, 19, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Ow, J.R.; Tan, B.C.; Goh, Z.; Feng, B.; Loh, Y.H.; Fedele, M.; Li, H.; Wu, Q. The dosage of Patz1 modulates reprogramming process. Sci. Rep. 2014, 4, 7519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Z.L.; Siew, J.; Li, J.; Ji, G.; Huang, M.; Liao, X.; Yu, S.; Chew, Y.; Png, C.W.; Zhang, Y.; et al. PATZ1 (MAZR) Co-occupies Genomic Sites With p53 and Inhibits Liver Cancer Cell Proliferation via Regulating p27. Front. Cell Dev. Biol. 2021, 9, 586150. [Google Scholar] [CrossRef]
- Tian, X.; Sun, D.; Zhang, Y.; Zhao, S.; Xiong, H.; Fang, J. Zinc finger protein 278, a potential oncogene in human colorectal cancer. Acta Biochim. Biophys. Sin. 2008, 40, 289–296. [Google Scholar] [CrossRef]
- Tian, X.Q.; Guo, F.F.; Sun, D.F.; Wang, Y.C.; Yang, L.; Chen, S.L.; Hong, J.; Fang, J.Y. Downregulation of ZNF278 arrests the cell cycle and decreases the proliferation of colorectal cancer cells via inhibition of the ERK/MAPK pathway. Oncol. Rep. 2017, 38, 3685–3692. [Google Scholar] [CrossRef] [Green Version]
- Keskin, N.; Deniz, E.; Eryilmaz, J.; Un, M.; Batur, T.; Ersahin, T.; Cetin Atalay, R.; Sakaguchi, S.; Ellmeier, W.; Erman, B. PATZ1 Is a DNA Damage-Responsive Transcription Factor That Inhibits p53 Function. Mol. Cell Biol. 2015, 35, 1741–1753. [Google Scholar] [CrossRef] [Green Version]
- Tritz, R.; Mueller, B.M.; Hickey, M.J.; Lin, A.H.; Gomez, G.G.; Hadwiger, P.; Sah, D.W.; Muldoon, L.; Neuwelt, E.A.; Kruse, C.A. siRNA Down-regulation of the PATZ1 Gene in Human Glioma Cells Increases Their Sensitivity to Apoptotic Stimuli. Cancer Ther. 2008, 6, 865–876. [Google Scholar]
- Tao, X.; Zhang, G.; Liu, J.; Ji, B.; Xu, H.; Chen, Z. PATZ1 Induces Apoptosis through PUMA in Glioblastoma. J. Oncol. 2022, 2022, 4953107. [Google Scholar] [CrossRef] [PubMed]
- Chiappetta, G.; Valentino, T.; Vitiello, M.; Pasquinelli, R.; Monaco, M.; Palma, G.; Sepe, R.; Luciano, A.; Pallante, P.; Palmieri, D.; et al. PATZ1 acts as a tumor suppressor in thyroid cancer via targeting p53-dependent genes involved in EMT and cell migration. Oncotarget 2015, 6, 5310–5323. [Google Scholar] [CrossRef] [Green Version]
- Iesato, A.; Nakamura, T.; Izumi, H.; Uehara, T.; Ito, K.I. PATZ1 knockdown enhances malignant phenotype in thyroid epithelial follicular cells and thyroid cancer cells. Oncotarget 2017, 8, 82754–82772. [Google Scholar] [CrossRef] [Green Version]
- Monaco, M.; Palma, G.; Vitiello, M.; Capiluongo, A.; D’Andrea, B.; Vuttariello, E.; Luciano, A.; Cerchia, L.; Chiappetta, G.; Arra, C.; et al. Loss of One or Two PATZ1 Alleles Has a Critical Role in the Progression of Thyroid Carcinomas Induced by the RET/PTC1 Oncogene. Cancers 2018, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Yan, M.; Li, C.; Feng, Z. POZ/BTB and AT-Hook-Containing Zinc Finger Protein 1 (PATZ1) Suppresses Progression of Ovarian Cancer and Serves as an Independent Prognosis Factor. Med. Sci. Monit. 2018, 24, 4262–4270. [Google Scholar] [CrossRef]
- Fedele, M.; Franco, R.; Salvatore, G.; Paronetto, M.P.; Barbagallo, F.; Pero, R.; Chiariotti, L.; Sette, C.; Tramontano, D.; Chieffi, G.; et al. PATZ1 gene has a critical role in the spermatogenesis and testicular tumours. J. Pathol. 2008, 215, 39–47. [Google Scholar] [CrossRef]
- Yao, T.; Wang, Q.; Zhang, W.; Bian, A.; Zhang, J. Identification of genes associated with renal cell carcinoma using gene expression profiling analysis. Oncol. Lett. 2016, 12, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.W.; Park, H.; Seo, S.P.; Byun, Y.J.; Piao, X.M.; Kim, S.M.; Kim, W.T.; Yun, S.J.; Jang, W.; Shon, H.S.; et al. Methylation Signature for Prediction of Progression Free Survival in Surgically Treated Clear Cell Renal Cell Carcinoma. Korean Med. Sci. 2019, 34, e144. [Google Scholar] [CrossRef]
- Al-Obaidy, K.I.; Bridge, J.A.; Cheng, L.; Sumegi, J.; Reuter, V.E.; Benayed, R.; Hameed, M.; Williamson, S.R.; Hes, O.; Alruwaii, F.I.; et al. EWSR1-PATZ1 fusion renal cell carcinoma: A recurrent gene fusion characterizing thyroid-like follicular renal cell carcinoma. Mod. Pathol. 2021, 34, 1921–1934. [Google Scholar] [CrossRef] [PubMed]
- Guadagno, E.; Vitiello, M.; Francesca, P.; Calì, G.; Caponnetto, F.; Cesselli, D.; Camorani, S.; Borrelli, G.; Califano, M.; Cappabianca, P.; et al. PATZ1 is a new prognostic marker of glioblastoma associated with the stem-like phenotype and enriched in the proneural subtype. Oncotarget 2017, 8, 59282–59300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passariello, A.; Errico, M.E.; Donofrio, V.; Maestrini, M.; Zerbato, A.; Cerchia, L.; Capasso, M.; Capasso, M.; Fedele, M. PATZ1 Is Overexpressed in Pediatric Glial Tumors and Correlates with Worse Event-Free Survival in High-grade Gliomas. Cancers 2019, 11, 1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhalabi, K.T.; Stichel, D.; Sievers, P.; Peterziel, H.; Sommerkamp, A.C.; Sturm, D.; Wittmann, A.; Sill, M.; Jäger, N.; Beck, P.; et al. PATZ1 fusions define a novel molecularly distinct neuroepithelial tumor entity with a broad histological spectrum. Acta Neuropathol. 2021, 142, 841–857. [Google Scholar] [CrossRef]
- Park, K.W.; Cai, Y.; Benjamin, T.; Qorbani, A.; George, J. Round Cell Sarcoma with EWSR1-PATZ1 Gene Fusion in the Neck: Case Report and Review of the Literature. Laryngoscope 2020, 130, E833–E836. [Google Scholar] [CrossRef]
- Gyorffy, B.; Surowiak, P.; Budczies, J.; Lanczky, A. Online survival analysis software to assess the prognostic value of biomarkers using transcriptomic data in non-small-cell lung cancer. PLoS ONE 2013, 8, e82241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan Meyer Plotter. Available online: www.kmplot.com (accessed on 15 November 2022).
- Franco, R.; Scognamiglio, G.; Valentino, E.; Vitiello, M.; Luciano, A.; Palma, G.; Arra, C.; La Mantia, E.; Panico, L.; Tenneriello, V.; et al. PATZ1 expression correlates positively with BAX and negatively with BCL6 and survival in human diffuse large B cell lymphomas. Oncotarget 2016, 7, 59158–59172. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- R2: Genomic Analysis and Visualization Platform. Available online: http://r2.amc.nl (accessed on 16 November 2022).
- Valentino, T.; Palmieri, D.; Vitiello, M.; Simeone, A.; Palma, G.; Arra, C.; Chieffi, P.; Chiariotti, L.; Fusco, A.; Fedele, M. Embryonic defects and growth alteration in mice with homozygous disruption of the Patz1 gene. J. Cell Physiol. 2013, 228, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Hellyer, J.A.; Patel, M.I. Sex disparities in lung cancer incidence: Validation of a long-observed trend. Transl. Lung Cancer Res. 2019, 8, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Marquardt, G.; Mullapudi, N.; Wang, T.; Han, W.; Shi, M.; Keller, S.; Zhu, C.; Locker, J.; Spivack, S.D. Lung cancer transcriptomes refined with laser capture microdissection. Am. J. Pathol. 2014, 184, 2868–2884. [Google Scholar] [CrossRef] [Green Version]
- Lanczky, A.; Gyorffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Human Protein Atlas. Available online: www.proteinatlas.org (accessed on 15 November 2022).
- Pan, L.N.; Ma, Y.F.; Li, Z.; Hu, J.A.; Xu, Z.H. KRAS G12V mutation upregulates PD-L1 expression via TGF-β/EMT signaling pathway in human non-small-cell lung cancer. Cell Biol. Int. 2021, 45, 795–803. [Google Scholar] [CrossRef]
- Liu, G.; Guo, Z.; Zhang, Q.; Liu, Z.; Zhu, D. AHNAK2 Promotes Migration, Invasion, and Epithelial-Mesenchymal Transition in Lung Adenocarcinoma Cells via the TGF-β/Smad3 Pathway. Onco Targets Ther. 2020, 13, 12893–12903. [Google Scholar] [CrossRef]
- Albini, A. Tumor and endothelial cell invasion of basement membranes. The matrigel chemoinvasion assay as a tool for dissecting molecular mechanisms. Pathol. Oncol. Res. 1998, 4, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Osmani, L.; Askin, F.; Gabrielson, E.; Li, Q.K. Current WHO guidelines and the critical role of immunohistochemical markers in the subclassification of non-small cell lung carcinoma (NSCLC): Moving from targeted therapy to immunotherapy. Semin. Cancer Biol. 2018, 52, 103–109. [Google Scholar] [CrossRef]
- Relli, V.; Trerotola, M.; Guerra, E.; Alberti, S. Abandoning the Notion of Non-Small Cell Lung Cancer. Trends Mol. Med. 2019, 25, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yang, S.; Jiang, H.; Wang, Y.; Xiang, Z. Transcriptomic analysis of tumor tissues and organoids reveals the crucial genes regulating the proliferation of lung adenocarcinoma. J. Transl. Med. 2021, 19, 368. [Google Scholar] [CrossRef]
- Jamal-Hanjani, M.; Wilson, G.A.; McGranahan, N.; Birkbak, N.J.; Watkins, T.B.K.; Veeriah, S.; Shafi, S.; Johnson, D.H.; Mitter, R.; Rosenthal, R.; et al. Tracking the evolution of nonsmall-cell lung cancer. N. Engl. J. Med. 2017, 376, 2109–2121. [Google Scholar] [CrossRef] [Green Version]
- Ferone, G.; Song, J.Y.; Sutherland, K.D.; Bhaskaran, R.; Monkhorst, K.; Lambooij, J.P.; Proost, N.; Gargiulo, G.; Berns, A. SOX2 Is the Determining Oncogenic Switch in Promoting Lung Squamous Cell Carcinoma from Different Cells of Origin. Cancer Cell 2016, 30, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Danés, A.; Blanpain, C. Deciphering the cells of origin of squamous cell carcinomas. Nat. Rev. Cancer 2018, 18, 549–561. [Google Scholar] [CrossRef]
- Ow, J.R.; Ma, H.; Jean, A.; Goh, Z.; Lee, Y.H.; Chong, Y.M.; Soong, R.; Fu, X.Y.; Yang, H.; Wu, Q. Patz1 regulates embryonic stem cell identity. Stem Cells Dev. 2014, 23, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Mancinelli, S.; Vitiello, M.; Donnini, M.; Mantile, F.; Palma, G.; Luciano, A.; Arra, C.; Cerchia, L.; Liguori, G.L.; Fedele, M. The Transcription Regulator Patz1 Is Essential for Neural Stem Cell Maintenance and Proliferation. Front. Cell Dev. Biol. 2021, 9, 657149. [Google Scholar] [CrossRef]
- Vitiello, M.; Palma, G.; Monaco, M.; Bello, A.M.; Camorani, S.; Francesca, P.; Rea, D.; Barbieri, A.; Chiappetta, G.; De Vita, G.; et al. Dual Oncogenic/Anti-Oncogenic Role of PATZ1 in FRTL5 Rat Thyroid Cells Transformed by the Ha-RasV12 Oncogene. Genes 2019, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Segura, A.; de Jong, T.V.; Melov, S.; Guryev, V.; Campisi, J.; Demaria, M. Unmasking Transcriptional Heterogeneity in Senescent Cells. Curr. Biol. 2017, 27, 2652–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitiello, M.; Valentino, T.; De Menna, M.; Crescenzi, E.; Francesca, P.; Rea, D.; Arra, C.; Fusco, A.; De Vita, G.; Cerchia, L.; et al. PATZ1 is a target of miR-29b that is induced by Ha-Ras oncogene in rat thyroid cells. Sci. Rep. 2016, 6, 25268. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.X.; Hua, Y.Z.; Wang, N.; Chen, X.; Wang, F.; Liu, J.Y.; Wang, L.L.; Yan, C.Y.; Ma, Y.G.; Cao, Y.H.; et al. miR-24 suppression of POZ/BTB and AT-hook-containing zinc finger protein 1 (PATZ1) protects endothelial cell from diabetic damage. Biochem. Biophys. Res. Commun. 2016, 480, 682–689. [Google Scholar] [CrossRef]
- Kazempour Dizaji, M.; Farzanegan, B.; Bahrami, N.; Khoshnam, Z.; Fathi, M.; Dargahi, H.; Pejhan, S.; Khosravi, A.; Shirian, S.; Narimani, A.; et al. Expression of miRNA1, miRNA133, miRNA191, and miRNA24, as Good Biomarkers, in Non-Small Cell Lung Cancer Using Real-Time PCR Method. Asian Pac. J. Cancer Prev. 2022, 23, 1565–1570. [Google Scholar] [CrossRef]
- Wang, X.H.; Gan, C.Z.; Xie, J.Y. Inhibition of miR-24 suppresses malignancy of human non-small cell lung cancer cells by targeting WWOX in vitro and in vivo. Thorac. Cancer 2018, 9, 1583–1593. [Google Scholar] [CrossRef] [Green Version]
- Esposito, F.; Boscia, F.; Franco, R.; Tornincasa, M.; Fusco, A.; Kitazawa, S.; Looijenga, L.H.; Chieffi, P. Down-regulation of oestrogen receptor-β associates with transcriptional co-regulator PATZ1 delocalization in human testicular seminomas. J. Pathol. 2011, 224, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.L.; Ravatn, R.; Kudoh, K.; Alabanza, L.; Chin, K.V. Interaction of the regulatory subunit of the cAMP-dependent protein kinase with PATZ1 (ZNF278). Biochem. Biophys. Res. Commun. 2010, 391, 1318–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzillo, A.; Ragone, A.; Spina, A.; Naviglio, S.; Sapio, L. Forskolin affects proliferation, migration and Paclitaxel-mediated cytotoxicity in non-small-cell lung cancer cell lines via adenylyl cyclase/cAMP axis. Eur. J. Cell Biol. 2023, 102, 151292. [Google Scholar] [CrossRef]
- Pawelczyk, K.; Piotrowska, A.; Ciesielska, U.; Jablonska, K.; Glatzel-Plucinska, N.; Grzegrzolka, J.; Podhorska-Okolow, M.; Dziegiel, P.; Nowinska, K. Role of PD-L1 expression in non-small cell lung cancer and their prognostic significance according to clinicopathological factors and diagnostic markers. Int. J. Mol. Sci. 2019, 20, 824. [Google Scholar] [CrossRef] [Green Version]
- Passiglia, F.; Bronte, G.; Bazan, V.; Natoli, C.; Rizzo, S.; Galvano, A.; Listì, A.; Cicero, G.; Rolfo, C.; Santini, D.; et al. PD-L1 expression as predictive biomarker in patients with NSCLC: A pooled analysis. Oncotarget 2016, 7, 19738–19747. [Google Scholar] [CrossRef] [Green Version]
- Frafjord, A.; Buer, L.; Hammarström, C.; Aamodt, H.; Woldbæk, P.R.; Brustugun, O.T.; Helland, Å.; Øynebråten, I.; Corthay, A. The Immune Landscape of Human Primary Lung Tumors Is Th2 Skewed. Front. Immunol. 2021, 12, 764596. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.; Gülich, A.F.; Alteneder, M.; Preglej, T.; Orola, M.J.; Dhele, N.; Stolz, V.; Schebesta, A.; Hamminger, P.; Hladik, A.; et al. The Transcription Factor MAZR/PATZ1 Regulates the Development of FOXP3+ Regulatory T Cells. Cell Rep. 2019, 29, 4447–4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TraFaC (Transcription Factor binding site Comparison). Available online: http://trafac.chmcc.org (accessed on 17 June 2013).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarker | Tot (n = 104) | Subtype and Grade | Stadium | |||
|---|---|---|---|---|---|---|
| Gender | F | 29 (27.9%) | LUSC | 8 | ||
| LUAD | 21 | |||||
| M | 75 (72.1%) | LUSC | 35 | |||
| LUAD | 40 | |||||
| Median Age | 65.6 ± 2.77 | LUSC | 66.5 ± 3.20 | |||
| LUAD | 65.2 ± 2.64 | |||||
| Subtype, grade and stadium | LUSC | 43 (41.3%) | G1 | 2 | IA1 | 4 |
| G2 | 27 | IA2 | 11 | |||
| G3 | 14 | IB | 7 | |||
| IIA | 8 | |||||
| IIB | 8 | |||||
| IIIA | 4 | |||||
| IIIB | 1 | |||||
| LUAD | 61 (58.7%) | G1 | 19 | IA1 | 3 | |
| G2 | 29 | IA2 | 12 | |||
| G3 | 13 | IB | 23 | |||
| IIA | 10 | |||||
| IIB | 5 | |||||
| IIIA | 6 | |||||
| IIIB | 2 | |||||
| Subtype 1 | N. | PATZ1 Sub-Cellular Localization 2 | Negative | ||
|---|---|---|---|---|---|
| N | N/C | C | |||
| LUSC | 43 | 15 (35%) | 16 (37%) | 7 (16%) | 5 (12%) |
| LUAD | 61 | 7 (11%) | 29 (47%) | 18 (29%) | 7 (11%) |
| Bronchial epithelium | 4 | 1 (25%) | 3 (75%) | - | - |
| Alveolar epithelium | 30 | 16 (53%) | - | - | 14 (47%) |
| Nuclear PATZ1 | p | R | |||
|---|---|---|---|---|---|
| (−) | (+) | ||||
| LUSC | (−) | 47 (87%) | 7 (13%) | 0.003 | 0.306 |
| (+) | 23 (60.5%) | 15 (39.5%) | |||
| LUAD | (−) | 23 (60.5%) | 15 (39.5%) | 0.003 | −0.306 |
| (+) | 47 (87%) | 7 (13%) | |||
| Subtype 1 | Low PATZ1 (Absent or <20%) | Medium PATZ1 (≥20%–<70%) | High PATZ1 (≥70%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (−) | N | N/C | C | N | N/C | C | N | N/C | C | |
| LUSC | 5 (11%) | 2 (5%) | 2 (5%) | 2 (5%) | 11 (26%) | 9 (21%) | 5 (11%) | 2 (5%) | 5 (11%) | 0 |
| LUAD | 7 (11%) | 1 (2%) | 0 | 5 (8%) | 5 (8%) | 28 (46%) | 13 (21%) | 1 (2%) | 1 (2%) | 0 |
| PATZ1 ≥ 70% | p | R | |||
|---|---|---|---|---|---|
| (−) | (+) | ||||
| LUSC | (−) | 58 (97%) | 2 (3%) | 0.022 | 0.226 |
| (+) | 36 (84%) | 7 (16%) | |||
| LUAD | (−) | 36 (84%) | 7 (16%) | 0.022 | −0.226 |
| (+) | 58 (97%) | 2 (3%) | |||
| PD-L1 | p | R | |||
|---|---|---|---|---|---|
| (−) | (+) | ||||
| High PATZ1 | (−) | 40 (47%) | 45 (53%) | 0.029 | −0.227 |
| (+) | 7 (87.5%) | 1 (12.5%) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucà, S.; Franco, R.; Napolitano, A.; Soria, V.; Ronchi, A.; Zito Marino, F.; Della Corte, C.M.; Morgillo, F.; Fiorelli, A.; Luciano, A.; et al. PATZ1 in Non-Small Cell Lung Cancer: A New Biomarker That Negatively Correlates with PD-L1 Expression and Suppresses the Malignant Phenotype. Cancers 2023, 15, 2190. https://doi.org/10.3390/cancers15072190
Lucà S, Franco R, Napolitano A, Soria V, Ronchi A, Zito Marino F, Della Corte CM, Morgillo F, Fiorelli A, Luciano A, et al. PATZ1 in Non-Small Cell Lung Cancer: A New Biomarker That Negatively Correlates with PD-L1 Expression and Suppresses the Malignant Phenotype. Cancers. 2023; 15(7):2190. https://doi.org/10.3390/cancers15072190
Chicago/Turabian StyleLucà, Stefano, Renato Franco, Antonella Napolitano, Valeria Soria, Andrea Ronchi, Federica Zito Marino, Carminia Maria Della Corte, Floriana Morgillo, Alfonso Fiorelli, Antonio Luciano, and et al. 2023. "PATZ1 in Non-Small Cell Lung Cancer: A New Biomarker That Negatively Correlates with PD-L1 Expression and Suppresses the Malignant Phenotype" Cancers 15, no. 7: 2190. https://doi.org/10.3390/cancers15072190
APA StyleLucà, S., Franco, R., Napolitano, A., Soria, V., Ronchi, A., Zito Marino, F., Della Corte, C. M., Morgillo, F., Fiorelli, A., Luciano, A., Palma, G., Arra, C., Battista, S., Cerchia, L., & Fedele, M. (2023). PATZ1 in Non-Small Cell Lung Cancer: A New Biomarker That Negatively Correlates with PD-L1 Expression and Suppresses the Malignant Phenotype. Cancers, 15(7), 2190. https://doi.org/10.3390/cancers15072190