Induction of Immunological Antitumor Effects by the Combination of Adenovirus-Mediated Gene Transfer of B7-1 and Anti-Programmed Cell Death-1 Antibody in a Murine Squamous Cell Carcinoma Model

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Cell Line
2.2. Adenoviral Vectors
2.3. Detection of B7-1 Expression on KLN205 Cells Infected with AdB7
Determination of Optimal Dose for AdB7 Infection
2.4. In Vivo Tumor Studies
2.5. Immunohistochemical Analysis
2.6. Multiplex Immunohistochemical Analysis
3. Results
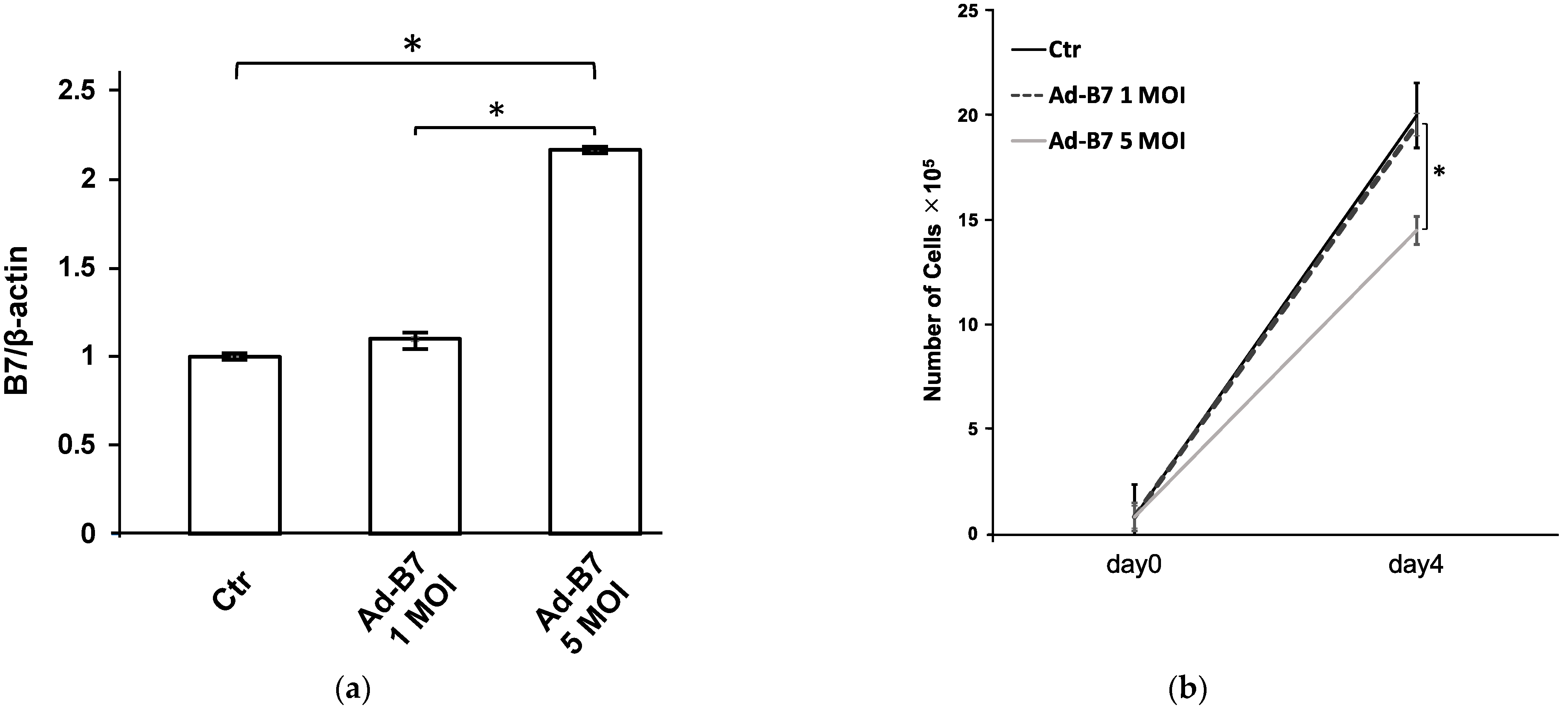
3.1. Detection of B7-1 Expression in KLN205 Cells Infected with AdB7
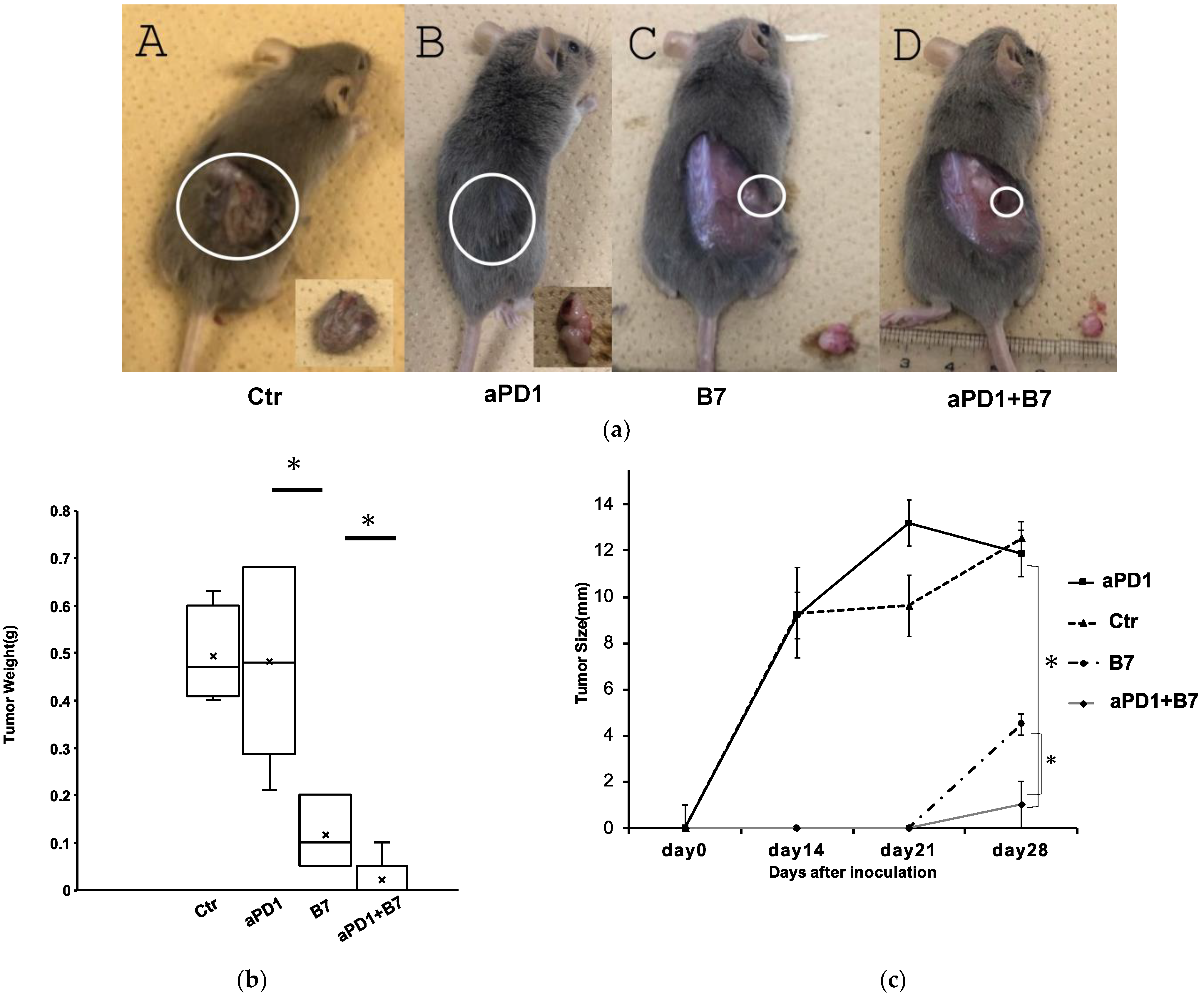
3.2. Antitumor Effects of B7-1 and Anti-PD-1 Antibody
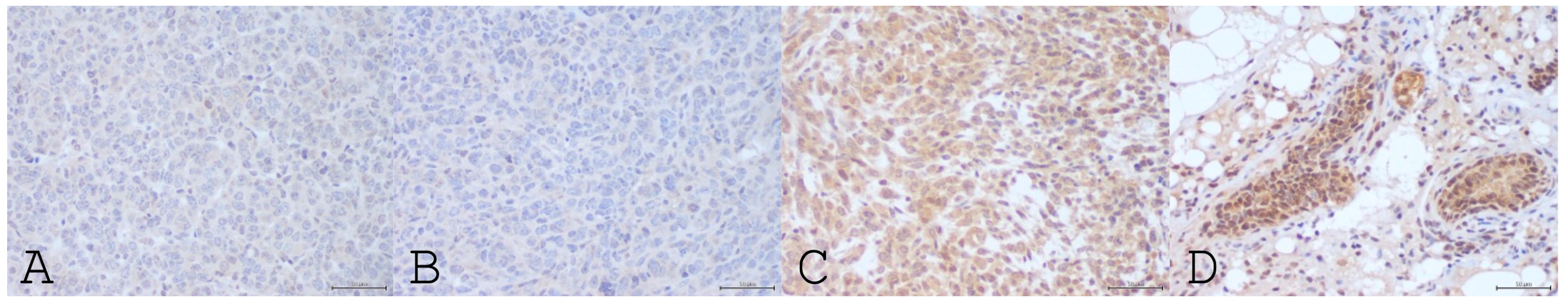
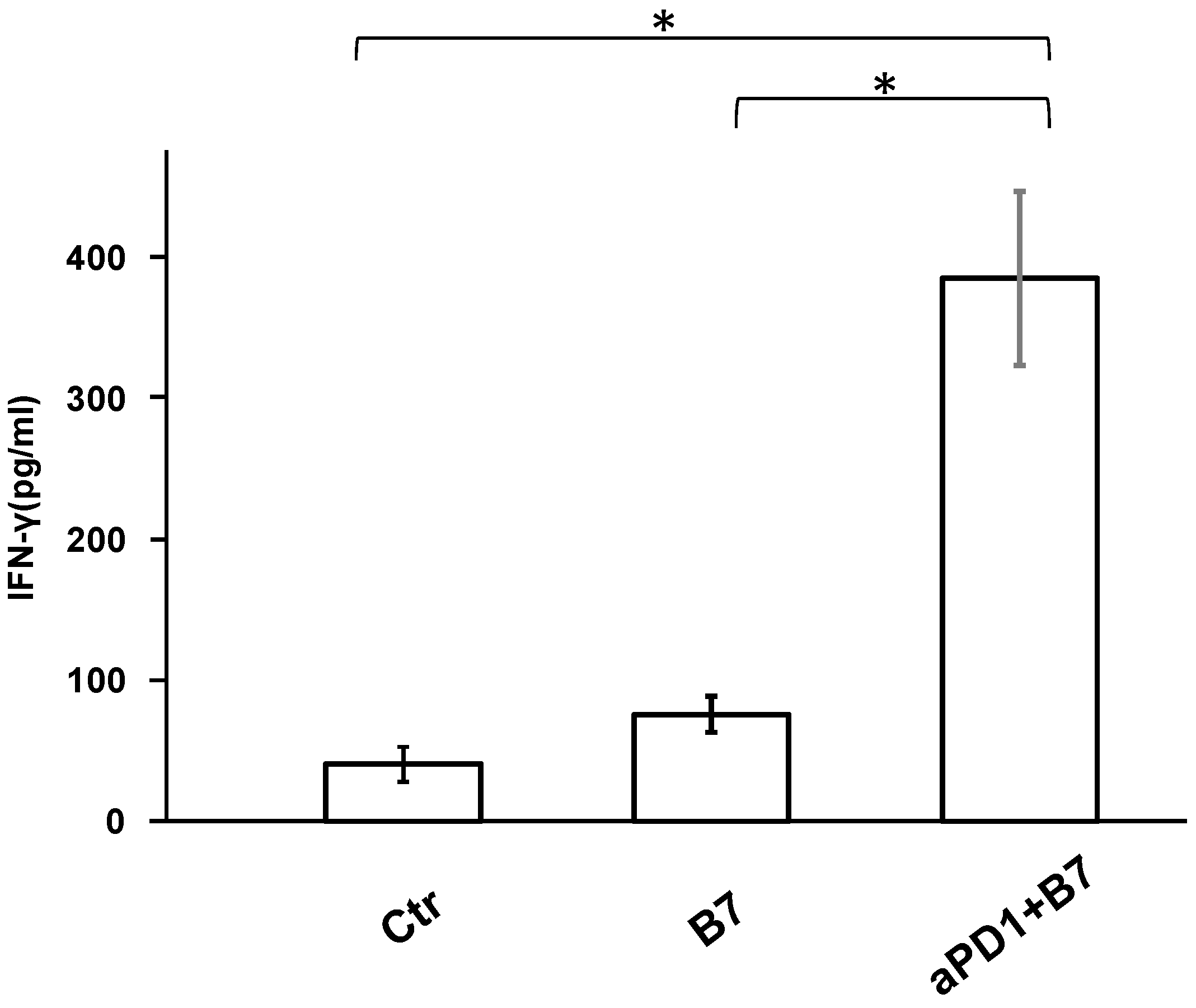
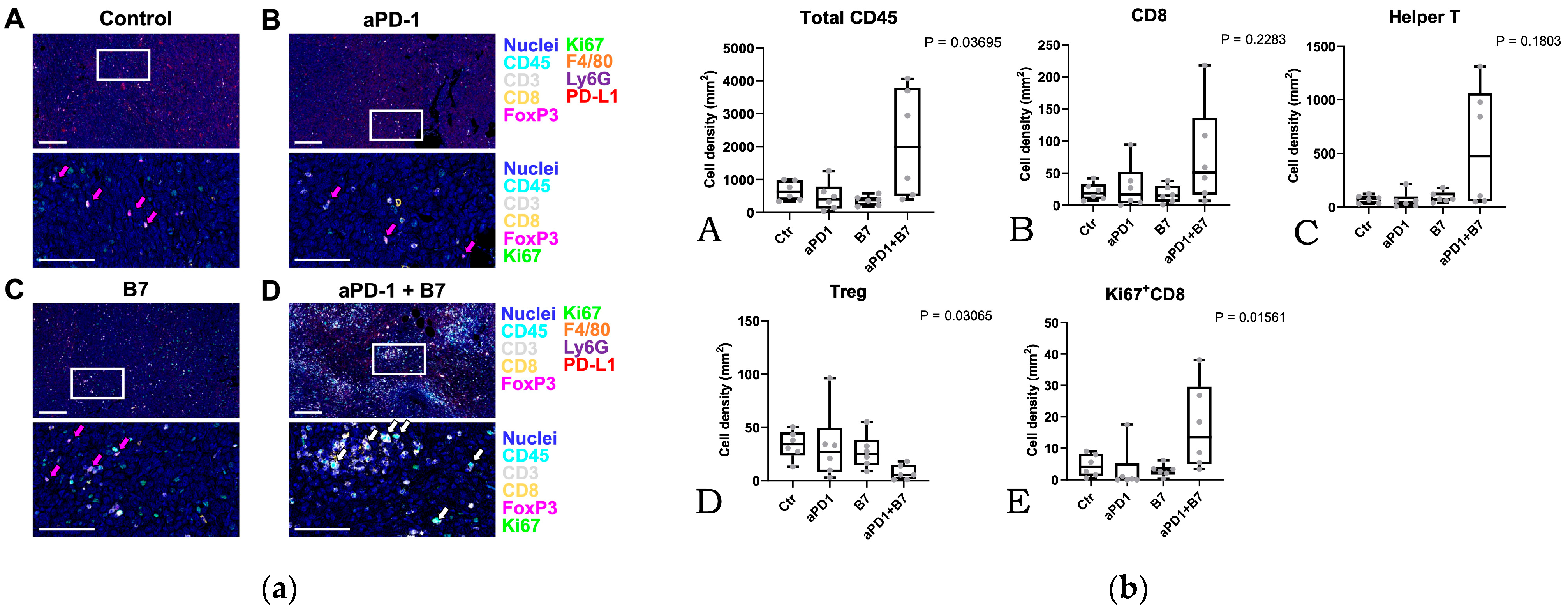
3.3. Tumor Microenvironment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- La, V. Head and neck cancers New etiological insights. In World Cancer Report 2020; Wild, C.P., Weiderpass, E., Stewart, B.W., Eds.; IARC Publication: Lyon, France, 2020; pp. 310–322. [Google Scholar]
- Vermorken, J.B.; Mesia, R.; Rivera, F.; Remenar, E.; Kawecki, A.; Rottey, S.; Erfan, J.; Zabolotnyy, D.; Kienzer, H.-R.; Cupissol, D.; et al. Platinum-Based Chemotherapy plus Cetuximab in Head and Neck Cancer. N. Engl. J. Med. 2008, 359, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarraf, M.; LeBlanc, M.; Giri, P.G.; Fu, K.K.; Cooper, J.; Vuong, T.; Forastiere, A.A.; Adams, G.; Sakr, W.A.; Schuller, D.E.; et al. Chemoradiotherapy versus radiotherapy in patients with advanced nasopharyngeal cancer: Phase III randomized Intergroup study 0099. J. Clin. Oncol. 1998, 16, 1310–1317. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, H.; Kawakita, D.; Taniyama, Y.; Oze, I.; Koyanagi, Y.N.; Hori, M.; Nakata, K.; Sugiyama, H.; Miyashiro, I.; Oki, I.; et al. Subsite-specific trends in mid- and long-term survival for head and neck cancer patients in Japan: A population-based study. Cancer Sci. 2023, 115, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Blumenschein, G., Jr.; Fayette, J.; Guigay, J.; Colevas, A.D.; Licitra, L.; Harrington, K.; Kasper, S.; Vokes, E.E.; Even, C.; et al. Nivolumab for Recurrent Squamous-Cell Carcinoma of the Head and Neck. N. Engl. J. Med. 2016, 375, 1856–1867. [Google Scholar] [CrossRef] [PubMed]
- Burtness, B.; Harrington, K.J.; Greil, R.; Soulières, D.; Tahara, M.; de Castro, G., Jr.; Psyrri, A.; Basté, N.; Neupane, P.; Bratland, A.; et al. Pembrolizumab alone or with chemotherapy versus cetuximab with chemotherapy for recurrent or metastatic squamous cell carcinoma of the head and neck (KEYNOTE-048): A randomised, open-label, phase 3 study. Lancet 2019, 394, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lee, N.Y. JAVELIN Head and Neck 100: A Phase III trial of avelumab and chemoradiation for locally advanced head and neck cancer. Futur. Oncol. 2019, 15, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Haddad, R.I.; Harrington, K.; Tahara, M.; Ferris, R.L.; Gillison, M.; Fayette, J.; Daste, A.; Koralewski, P.; Zurawski, B.; Taberna, M.; et al. Nivolumab Plus Ipilimumab Versus EXTREME Regimen as First-Line Treatment for Recurrent/Metastatic Squamous Cell Carcinoma of the Head and Neck: The Final Results of CheckMate 651. J. Clin. Oncol. 2022, 41, 2166–2180. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. MHC-dependent antigen processing and peptide presentation: Providing ligands for T lymphocyte activation. Cell 1994, 76, 287–299. [Google Scholar] [CrossRef]
- Schwartz, R.H. Costimulation of T lymphocytes: The role of CD28, CTLA-4, and B7/BB1 in interleukin-2 production and immunotherapy. Cell 1992, 71, 1065–1068. [Google Scholar] [CrossRef]
- Bolandi, N.; Derakhshani, A.; Hemmat, N.; Baghbanzadeh, A.; Asadzadeh, Z.; Nour, M.A.; Brunetti, O.; Bernardini, R.; Silvestris, N.; Baradaran, B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 10719. [Google Scholar] [CrossRef]
- Tirapu, I.; Huarte, E.; Guiducci, C.; Arina, A.; Zaratiegui, M.; Murillo, O.; Gonzalez, A.; Berasain, C.; Berraondo, P.; Fortes, P.; et al. Low surface expression of B7-1 (CD80) is an immunoescape mechanism of colon carcinoma. Cancer Res. 2006, 66, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-Y.; Yu, Z.-L.; Fu, Q.-Y.; Zhang, L.-Z.; Li, J.-B.; Wu, M.; Liu, B.; Chen, G. Immunosuppressive effect of small extracellular vesicle PD-L1 is restricted by co-expression of CD80. Br. J. Cancer 2023, 129, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Hoshitani, Y.; Ishida, H.; Otsuki, N.; Shirakawa, T.; Gotoh, A.; Nibu, K. Induction of immunological antitumor effects by adenovirus-mediated gene transfer of B7-1 in a murine squamous cell carcinoma cell line. Arch. Otolaryngol. Head Neck Surg. 2007, 133, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Sakate, Y.; Yashiro, M.; Tanaka, H.; Kosaka, K.; Ohira, M.; Hirakawa, K. Suppression of peritoneal metastasis in human gastric carcinoma by enhanced immunogenicity of B7-1 transfection. Oncol. Rep. 2004, 12, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Sakate, Y.; Yashiro, M.; Tanaka, H.; Sunami, T.; Kosaka, K.; Hirakawa, K. Usefulness of inhibiting the lymph node metastasis in human gastric carcinoma by B7–1 gene transfection1. J. Surg. Res. 2004, 122, 89–95. [Google Scholar] [CrossRef]
- Morioka, J.; Kajiwara, K.; Yoshikawa, K.; Ideguchi, M.; Uchida, T.; Ohmoto, Y.; Suzuki, M. Adenovirus-mediated Gene Transfer of B7.1 Induces Immunological Anti-tumor Effects in a Murine Brain Tumor. J. Neuro-Oncol. 2002, 60, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, T.; Shimada, M.; Gabriela, G.; Kosugi, T.; Sato, W.; Lee, P.Y.; Lanaspa, M.A.; Rivard, C.; Maruyama, S.; Garin, E.H.; et al. Toll-like receptor 3 ligand, polyIC, induces proteinuria and glomerular CD80, and increases urinary CD80 in mice. Nephrol. Dial. Transplant. 2013, 28, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wu, Y.; Zhang, G.; Wang, M.; Yang, H.; Li, Q. γδT Cells Exacerbate Podocyte Injury via the CD28/B7-1-Phosphor-SRC Kinase Pathway. BioMed Res. Int. 2018, 2018, 5647120. [Google Scholar] [CrossRef] [PubMed]
- Tsujikawa, T.; Kumar, S.; Borkar, R.N.; Azimi, V.; Thibault, G.; Chang, Y.H.; Balter, A.; Kawashima, R.; Choe, G.; Sauer, D.; et al. Quantitative Multiplex Immunohistochemistry Reveals Myeloid-Inflamed Tumor-Immune Complexity Associated with Poor Prognosis. Cell Rep. 2017, 19, 203–217. [Google Scholar] [CrossRef]
- Netzer, C.; von Arps-Aubert, V.; Mačinković, I.; Von Der Grün, J.; Küffer, S.; Ströbel, P.; von Knethen, A.; Weigert, A.; Beutner, D. Association between spatial distribution of leukocyte subsets and clinical presentation of head and neck squamous cell carcinoma. Front. Immunol. 2024, 14, 1240394. [Google Scholar] [CrossRef]
- Chaves, P.; Garrido, M.; Oliver, J.; Pérez-Ruiz, E.; Barragan, I.; Rueda-Domínguez, A. Preclinical models in head and neck squamous cell carcinoma. Br. J. Cancer 2023, 128, 1819–1827. [Google Scholar] [CrossRef]
- Miserocchi, G.; Spadazzi, C.; Calpona, S.; De Rosa, F.; Usai, A.; De Vita, A.; Liverani, C.; Cocchi, C.; Vanni, S.; Calabrese, C.; et al. Precision Medicine in Head and Neck Cancers: Genomic and Preclinical Approaches. J. Pers. Med. 2022, 12, 854. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; McGowan, P.; Hellström, I.; Hellström, K.E.; Chen, L. Costimulation of tumor-reactive CD4+ and CD8+ T lymphocytes by B7, a natural ligand for CD28, can be used to treat established mouse melanoma. J. Immunol. 1994, 153, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Boxhorn, H.K.E.; Smith, J.G.; Chang, Y.J.; Guerry, D.; Lee, W.M.F.; Rodeck, U.; Turka, L.A.; Eck, S.L. Adenoviral transduction of melanoma cells with B7-1: Antitumor immunity and immunosuppressive factors. Cancer Immunol. Immunother. 1998, 46, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Takenoyama, M.; Yoshino, I.; Eifuku, R.; So, T.; Imahayashi, S.; Sugaya, M.; Yasuda, M.; Inoue, M.; Ichiyoshi, Y.; Osaki, T.; et al. Successful Induction of Tumor-specific Cytotoxic T Lymphocytes from Patients with Non-small Cell Lung Cancer Using CD80-transfected Autologous Tumor Cells. Jpn. J. Cancer Res. 2001, 92, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Joki, T.; Kikuchi, T.; Akasaki, Y.; Saitoh, S.; Abe, T.; Ohno, T. Induction of effective antitumor immunity in a mouse brain tumor model using B7-1 (CD80) and intercellular adhesive molecule 1 (ICAM-1; CD54) transfection and recombinant interleukin 12. Int. J. Cancer 1999, 82, 714–720. [Google Scholar] [CrossRef]
- Morioka, J.; Kajiwara, K.; Yoshikawa, K.; Ideguchi, M.; Uchida, T.; Suzuki, M. Vaccine Therapy for Murine Glioma Using Tumor Cells Genetically Modified to Express B7.1. Neurosurgery 2004, 54, 182–189, Discussion 189–190. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, J.J.; Sullivan, T.J.; Allison, J.P. CTLA-4 Overexpression Inhibits T Cell Responses through a CD28-B7-Dependent Mechanism. J. Immunol. 2006, 177, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Krummel, M.F.; Allison, J.P. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J. Exp. Med. 1995, 182, 459–465. [Google Scholar] [CrossRef]
- Chang, S.R.; Chou, C.H.; Liu, C.J.; Lin, Y.C.; Tu, H.F.; Chang, K.W.; Lin, S.C. The Concordant Disruption of B7/CD28 Immune Regulators Predicts the Prognosis of Oral Carcinomas. Int. J. Mol. Sci. 2023, 24, 5931. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycle 1 | Cycle 2 | Cycle 3 | Cycle 4 | Cycle 5 | Cycle 6 | Cycle 7 | Cycle 8 | Cycle 9 | |
|---|---|---|---|---|---|---|---|---|---|
| Primary Ab | Hematoxylin | PDL1 | CD8 | CD3 | F4/80 | CD45 | Foxp3 | Ki67 | Ly6G |
| Supplier | Dako | Proteintech | eBioscience | Gene Tex | Abcam | BD Pharmingen | Invitrogen | Cell Signaling | ABA |
| Clone/Product #number | S330130-2 | 17952-1-AP | 14-0808-82 | SP7 | ab6640 | 30-F11 | FJK16s | D3B5 | 1A8 |
| Conc | Original | 1/200 | 1/50 | 1/400 | 1/400 | 1/200 | 1/100 | 1/10,000 | 1/2000 |
| Reaction time | 2 min | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min |
| Secondary Ab | Anti-rabbit | Anti-rat | Anti-rabbit | Anti-rat | Anti-rat | Anti-rat | Anti-rabbit | Anti-rat | |
| Reaction | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min | 30 min | |
| AEC | 20 min | 20 min | 20 min | 20 min | 20 min | 20 min | 20 min | ||
| AMEC | 5 min | ||||||||
| Heat treatment | Citrate | Citrate | Citrate | Citrate | Citrate | Citrate | Citrate | Citrate | Citrate |
| Cell Type | Identification Biomarkers |
|---|---|
| CD8+ T cells | CD45+ CD3+ CD8+ |
| Proliferating CD8+ T cells | CD45+ CD3− CD8− Ki-67+ |
| Hepler T cells # | CD45+ CD3+ CD8− FoxP3− |
| Regulatory T cells (Treg) | CD45+ CD3+ CD8− FoxP3+ |
| Tumor-Associated Macrophages | CD45+ CD3− CD20− F4/80+ |
| Granulocytes | CD45+ CD3− CD8− Ly6G+ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hara, M.; Saburi, S.; Uehara, N.; Tsujikawa, T.; Kubo, M.; Furukawa, T.; Teshima, M.; Shinomiya, H.; Hirano, S.; Nibu, K.-i. Induction of Immunological Antitumor Effects by the Combination of Adenovirus-Mediated Gene Transfer of B7-1 and Anti-Programmed Cell Death-1 Antibody in a Murine Squamous Cell Carcinoma Model. Cancers 2024, 16, 1359. https://doi.org/10.3390/cancers16071359
Hara M, Saburi S, Uehara N, Tsujikawa T, Kubo M, Furukawa T, Teshima M, Shinomiya H, Hirano S, Nibu K-i. Induction of Immunological Antitumor Effects by the Combination of Adenovirus-Mediated Gene Transfer of B7-1 and Anti-Programmed Cell Death-1 Antibody in a Murine Squamous Cell Carcinoma Model. Cancers. 2024; 16(7):1359. https://doi.org/10.3390/cancers16071359
Chicago/Turabian StyleHara, Makiko, Sumiyo Saburi, Natsumi Uehara, Takahiro Tsujikawa, Mie Kubo, Tatsuya Furukawa, Masanori Teshima, Hirotaka Shinomiya, Shigeru Hirano, and Ken-ichi Nibu. 2024. "Induction of Immunological Antitumor Effects by the Combination of Adenovirus-Mediated Gene Transfer of B7-1 and Anti-Programmed Cell Death-1 Antibody in a Murine Squamous Cell Carcinoma Model" Cancers 16, no. 7: 1359. https://doi.org/10.3390/cancers16071359
APA StyleHara, M., Saburi, S., Uehara, N., Tsujikawa, T., Kubo, M., Furukawa, T., Teshima, M., Shinomiya, H., Hirano, S., & Nibu, K. -i. (2024). Induction of Immunological Antitumor Effects by the Combination of Adenovirus-Mediated Gene Transfer of B7-1 and Anti-Programmed Cell Death-1 Antibody in a Murine Squamous Cell Carcinoma Model. Cancers, 16(7), 1359. https://doi.org/10.3390/cancers16071359