tRNA-Derived Fragments as Biomarkers in Bladder Cancer

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Samples
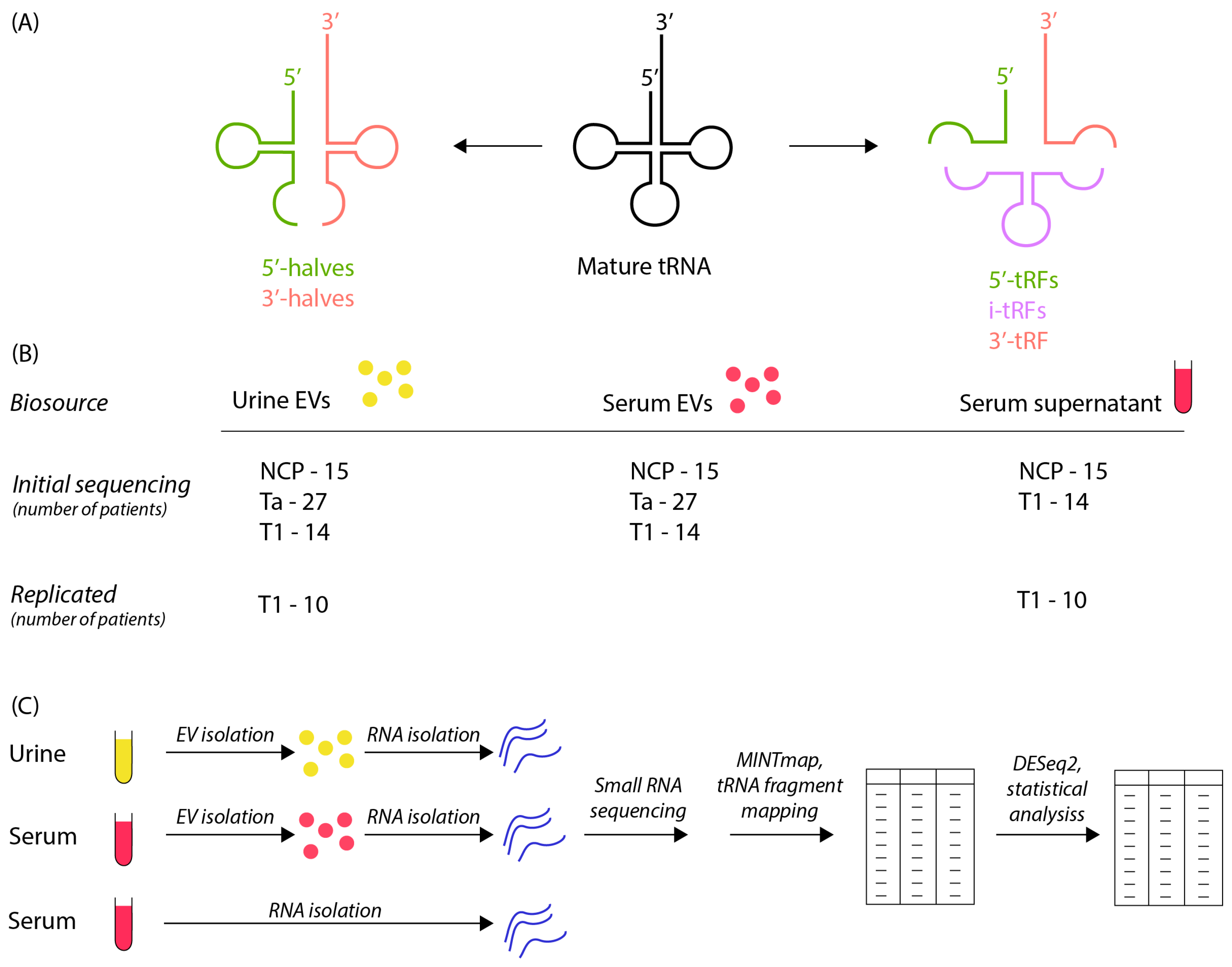
2.2. Workflow and Study Design
2.3. RNA Isolation
2.4. Small RNA Sequencing
2.5. Statistical Analysis
2.6. BLAST Search
3. Results
3.1. Sample Characterization
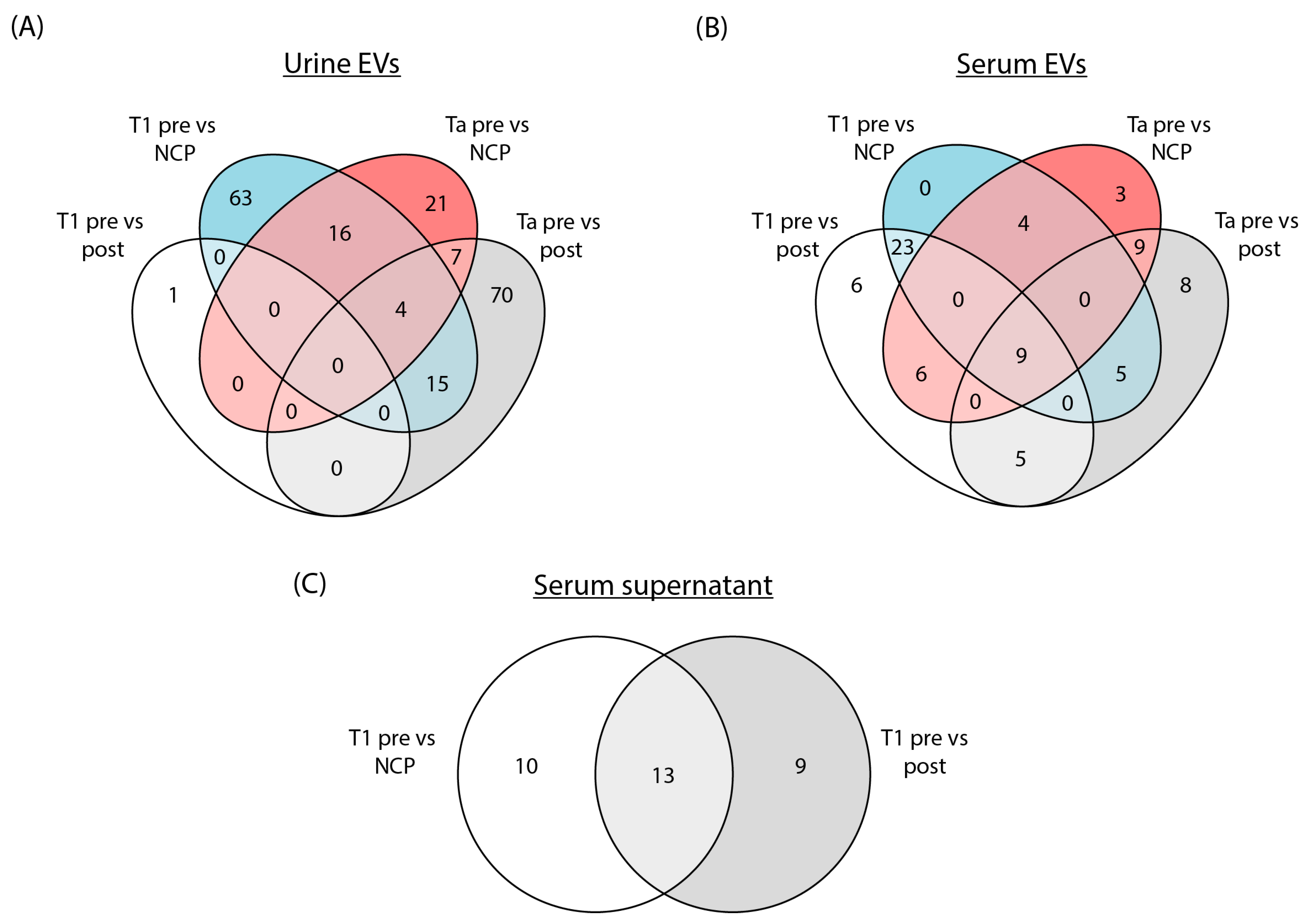
3.2. Differential Expression of tRFs in Urine EVs, Serum EVs and Serum Supernatant in the Initial Sequencing Run
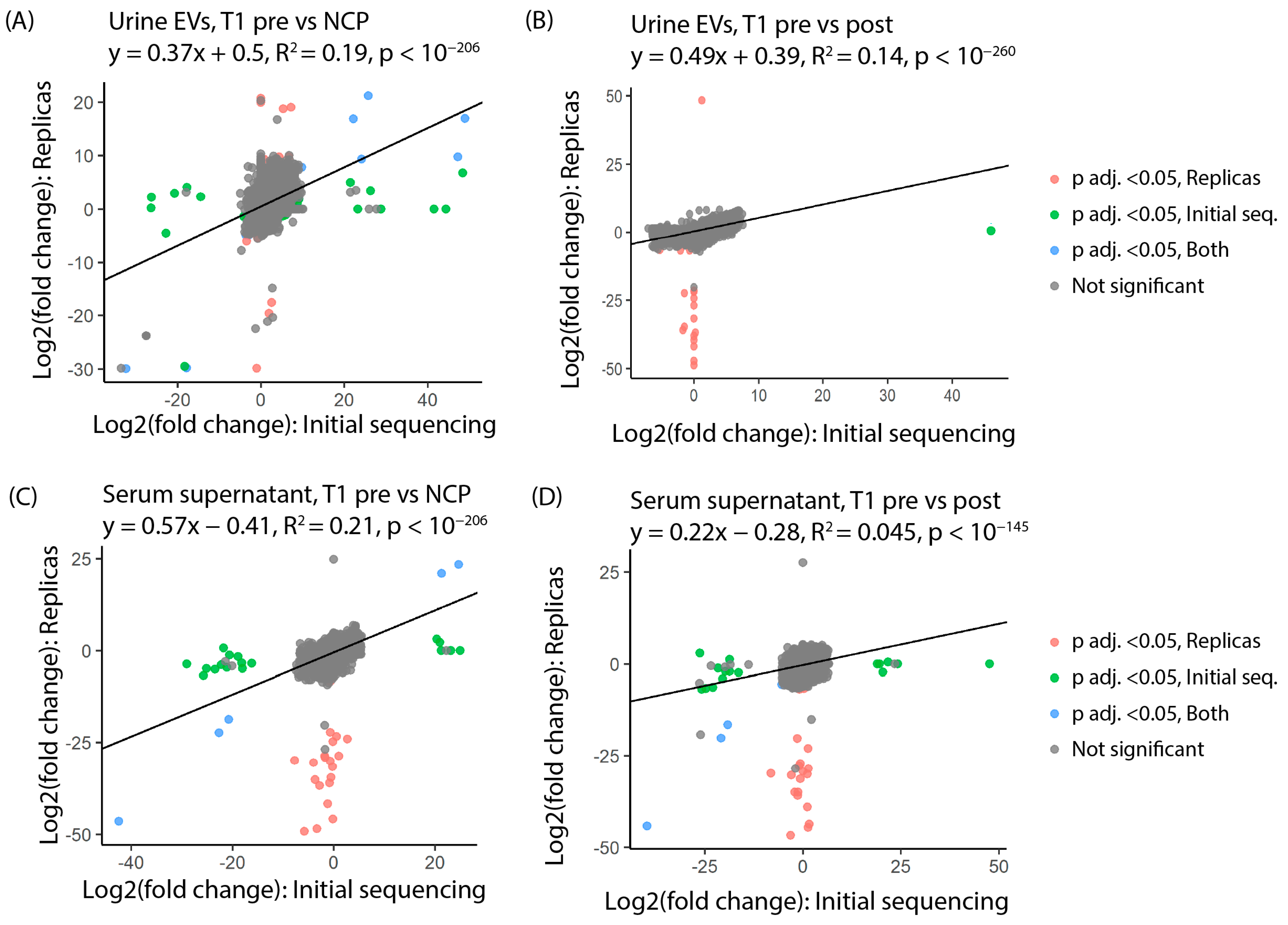
3.3. tRFs Are Confirmed as Differentially Expressed among Patients with Stage T1 Disease by Replica Sequencing
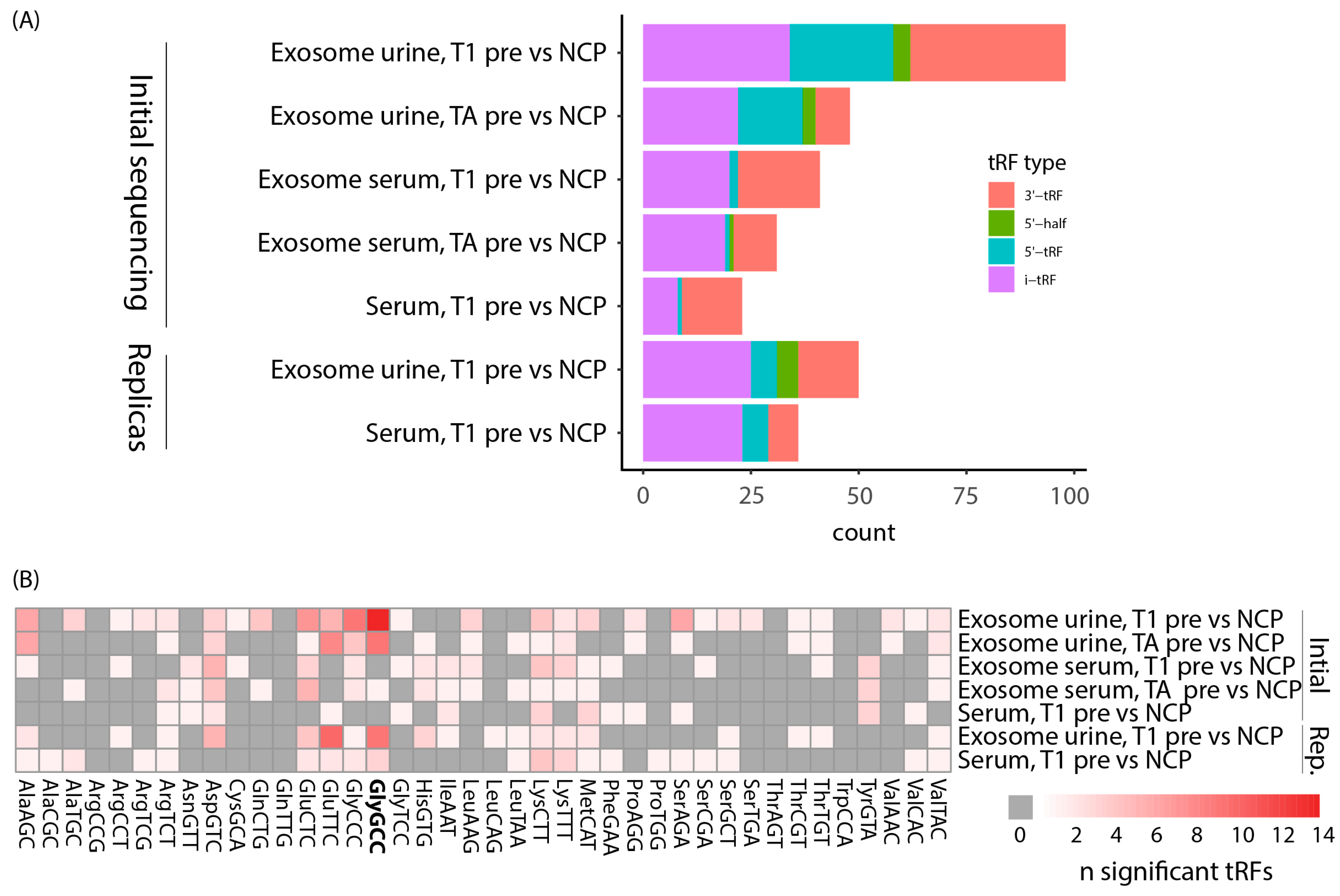
3.4. tRFs Differentially Expressed between Bladder Cancer and Noncancer Patients Are of Diverse Types and Originate from a Multitude of tRNAs
3.5. Assessment of Potential Role for tRFs in Biological Processes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trail, M.; Cullen, J.; Fulton, E.; Clayton, F.; McGregor, E.; McWilliam, F.; Dick, L.; Kalima, P.; Donat, R.; Mariappan, P. Evaluating the Safety of Performing Flexible Cystoscopy When Urinalysis Suggests Presence of “Infection”: Results of a Prospective Clinical Study in 2350 patients. Eur. Urol. Open Sci. 2021, 31, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Toden, S.; Goel, A. Non-coding RNAs as liquid biopsy biomarkers in cancer. Br. J. Cancer 2022, 126, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, D.; Jiang, L.; Wu, Y.; Yuan, H.; Shi, G.; Liu, F.; Wu, P.; Jiang, K. Biological functions and clinical significance of tRNA-derived small fragment (tsRNA) in tumors: Current state and future perspectives. Cancer Lett. 2024, 587, 216701. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, M.B.; Stephenson, M.L.; Scott, J.F.; Hecht, L.I.; Zamecnik, P.C. A soluble ribonucleic acid intermediate in protein synthesis. J. Biol. Chem. 1958, 231, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Speer, J.; Gehrke, C.W.; Kuo, K.C.; Waalkes, T.P.; Borek, E. tRNA breakdown products as markers for cancer. Cancer 1979, 44, 2120–2123. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, T.P.; Gehrke, C.W.; Zumwalt, R.W.; Chang, S.Y.; Lakings, D.B.; Tormey, D.C.; Ahmann, D.L.; Moertel, C.G. The urinary excretion of nucleosides of ribonucleic acid by patients with advanced cancer. Cancer 1975, 36, 390–398. [Google Scholar] [CrossRef]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes. Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef]
- Weng, Q.; Wang, Y.; Xie, Y.; Yu, X.; Zhang, S.; Ge, J.; Li, Z.; Ye, G.; Guo, J. Extracellular vesicles-associated tRNA-derived fragments (tRFs): Biogenesis, biological functions, and their role as potential biomarkers in human diseases. J. Mol. Med. 2022, 100, 679–695. [Google Scholar] [CrossRef]
- Wilson, B.; Dutta, A. Function and Therapeutic Implications of tRNA Derived Small RNAs. Front. Mol. Biosci. 2022, 9, 888424. [Google Scholar] [CrossRef]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA-Related Fragments (tRFs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef]
- Zhang, M.; Li, F.; Wang, J.; He, W.; Li, Y.; Li, H.; Wei, Z.; Cao, Y. tRNA-derived fragment tRF-03357 promotes cell proliferation, migration and invasion in high-grade serous ovarian cancer. Onco. Targets Ther. 2019, 12, 6371–6383. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Chen, W.; Huang, X.; Ye, D. Role of tRNA-derived small RNAs(tsRNAs) in the diagnosis and treatment of malignant tumours. Cell Commun. Signal 2023, 21, 178. [Google Scholar] [CrossRef]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Gu, J.; Wang, M.; Zhang, J.; Chen, Y.; Jiang, P.; Zhu, T.; Zhang, X. Emerging roles of tRNA-derived fragments in cancer. Mol. Cancer 2023, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Endres, L. tRNA modification and cancer: Potential for therapeutic prevention and intervention. Futire Med. Cemistry 2019, 8, 885–900. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.P.; Hutvagner, G. tRNA-Derived Fragments (tRFs): Emerging New Roles for an Ancient RNA in the Regulation of Gene Expression. Life 2015, 5, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yao, L.; Yu, X.; Ruan, Y.; Li, Z.; Guo, J. Action mechanisms and research methods of tRNA-derived small RNAs. Signal Transduct. Target. Ther. 2020, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Monshaugen, I.; Klungland, A.; Ougland, R.; Dutta, A. Characterization of novel small non-coding RNAs and their modifications in bladder cancer using an updated small RNA-seq workflow. Front. Mol. Biosci. 2022, 9, 887686. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D‘Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M.; Thery, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Kroemer, G.; Zitvogel, L. Extracellular vesicles: Masters of intercellular communication and potential clinical interventions. J. Clin. Invest. 2016, 126, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Bebelman, M.P.; Janssen, E.; Pegtel, D.M.; Crudden, C. The forces driving cancer extracellular vesicle secretion. Neoplasia 2021, 23, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.E.; Korbie, D.; Hill, M.M.; Trau, M. Extracellular vesicles as circulating cancer biomarkers: Opportunities and challenges. Clin. Transl. Med. 2018, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, J.; Gong, Y.; Wu, Q.; Tan, S.; Sun, D.; Xu, X.; Zuo, Y.; Zhao, Y.; Wei, Y.Q.; et al. Exosomal tRNA-derived small RNA as a promising biomarker for cancer diagnosis. Mol. Cancer 2019, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Lin, C. tRFs as Potential Exosome tRNA-Derived Fragment Biomarkers for Gastric Carcinoma. Clin. Lab. 2020, 66. [Google Scholar] [CrossRef] [PubMed]
- Guzman, N.; Agarwal, K.; Asthagiri, D.; Yu, L.; Saji, M.; Ringel, M.D.; Paulaitis, M.E. Breast Cancer-Specific miR Signature Unique to Extracellular Vesicles Includes “microRNA-like” tRNA Fragments. Mol. Cancer Res. 2015, 13, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Koi, Y.; Tsutani, Y.; Nishiyama, Y.; Ueda, D.; Ibuki, Y.; Sasada, S.; Akita, T.; Masumoto, N.; Kadoya, T.; Yamamoto, Y.; et al. Predicting the presence of breast cancer using circulating small RNAs, including those in the extracellular vesicles. Cancer Sci. 2020, 111, 2104–2115. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Green, B.B.; Seigne, J.D.; Schned, A.R.; Marsit, C.J. MicroRNA molecular profiling from matched tumor and bio-fluids in bladder cancer. Mol. Cancer 2015, 14, 194. [Google Scholar] [CrossRef]
- Stromme, O.; Heck, K.A.; Brede, G.; Lindholm, H.T.; Otterlei, M.; Arum, C.J. Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors. Curr. Issues Mol. Biol. 2021, 43, 286–300. [Google Scholar] [CrossRef]
- Loher, P.; Telonis, A.G.; Rigoutsos, I. MINTmap: Fast and exhaustive profiling of nuclear and mitochondrial tRNA fragments from short RNA-seq data. Sci. Rep. 2017, 7, 41184. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Srinivasan, S.; Yeri, A.; Cheah, P.S.; Chung, A.; Danielson, K.; De Hoff, P.; Filant, J.; Laurent, C.D.; Laurent, L.D.; Magee, R.; et al. Small RNA Sequencing across Diverse Biofluids Identifies Optimal Methods for exRNA Isolation. Cell 2019, 177, 446–462 e416. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.; Pham, P.; Dedon, P.C.; Begley, T.J. Lifestyle modifications: Coordinating the tRNA epitranscriptome with codon bias to adapt translation during stress responses. Genome Biol. 2018, 19, 228. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, X.; Li, J.; Fu, Z.; Hua, Y.; Zeng, T.; He, Y.; Duan, N.; Yang, F.; Liang, Y.; et al. Exosome-Transmitted tRF-16-K8J7K1B Promotes Tamoxifen Resistance by Reducing Drug-Induced Cell Apoptosis in Breast Cancer. Cancers 2023, 15, 899. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; Tan, X.; Mao, X.; Wei, D.; Yao, Y.; Jiang, P.; Mo, D.; Wang, T.; Yan, F. Identification of tRNA-Derived Fragments Expression Profile in Breast Cancer Tissues. Curr. Genomics 2019, 20, 199–213. [Google Scholar] [CrossRef]
- Su, Z.; Monshaugen, I.; Wilson, B.; Wang, F.; Klungland, A.; Ougland, R.; Dutta, A. TRMT6/61A-dependent base methylation of tRNA-derived fragments regulates gene-silencing activity and the unfolded protein response in bladder cancer. Nat. Commun. 2022, 13, 2165. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Djingo, D. Mammalian-wide interspersed repeat (MIR)-derived enhancers and the regulation of human gene expression. Mobile DNA 2014, 5, 1–12. [Google Scholar]
- Maraver, A.; Fernandez-Marcos, P.J.; Cash, T.P.; Mendez-Pertuz, M.; Duenas, M.; Maietta, P.; Martinelli, P.; Munoz-Martin, M.; Martinez-Fernandez, M.; Canamero, M.; et al. NOTCH pathway inactivation promotes bladder cancer progression. J. Clin. Invest. 2015, 125, 824–830. [Google Scholar] [CrossRef]
- Yu, M.; Yi, J.; Qiu, Q.; Yao, D.; Li, J.; Yang, J.; Mi, C.; Zhou, L.; Lu, B.; Lu, W.; et al. Pan-cancer tRNA-derived fragment CAT1 coordinates RBPMS to stabilize NOTCH2 mRNA to promote tumorigenesis. Cell Rep. 2023, 42, 113408. [Google Scholar] [CrossRef] [PubMed]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O‘Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Chen, Z.H.; Cao, R.; Shi, M.J.; Tian, Y. Differential Expression Profiles and Bioinformatics Analysis of tRNA-Derived Small RNAs in Muscle-Invasive Bladder Cancer in a Chinese Population. Genes 2022, 13, 601. [Google Scholar] [CrossRef]
- Papadimitriou, M.A.; Avgeris, M.; Levis, P.; Papasotiriou, E.C.; Kotronopoulos, G.; Stravodimos, K.; Scorilas, A. tRNA-Derived Fragments (tRFs) in Bladder Cancer: Increased 5‘-tRF-LysCTT Results in Disease Early Progression and Patients‘ Poor Treatment Outcome. Cancers 2020, 12, 3661. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phrase | n | tRF Names |
|---|---|---|
| FCGR2A | 6 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-17-D9W1X6K |
| GPD2 | 6 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-17-D9W1X6K |
| H3K4me1 hESC enhancer | 11 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-25-7P596VW631, tRF-20-40KK5Y93, tRF-17-I7XUK8N, tRF-17-D9W1X6K, tRF-17-8R6546J, tRF-16-F1R3WEE |
| HES7 | 6 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-17-D9W1X6K |
| HSPA6 | 6 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-17-D9W1X6K |
| lncRNA | 7 | tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-25-7P596VW631, tRF-21-86J8WPMNB, tRF-18-MBQ4NKDJ, tRF-17-D9W1X6K, tRF-16-F1R3WEE |
| LTA4 | 1 | tRF-16-F1R3WEE |
| piRNA | 3 | tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-18-MBQ4NKDJ |
| regulatory element | 7 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-18-MBQ4NKDJ, tRF-17-D9W1X6K |
| tRNA | 12 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-25-7P596VW631, tRF-21-86J8WPMNB, tRF-20-40KK5Y93, tRF-18-MBQ4NKDJ, tRF-17-I7XUK8N, tRF-17-D9W1X6K, tRF-16-F1R3WEE |
| VAC14 | 6 | tRF-38-PNR8YP9LON4VN18, tRF-32-PER8YP9LON4V3, tRF-31-PER8YP9LON4VD, tRF-27-J87383RPD95, tRF-26-IK9NJ4S2I7D, tRF-17-D9W1X6K |
| Phrase | n | tRF Names |
|---|---|---|
| H3K4me1 hESC enhancer | 2 | tRF-29-W4R951KHZK1V, tRF-16-F1R3WEE |
| lncRNA | 3 | tRF-29-W4R951KHZK1V, tRF-18-F8DHXYD9, tRF-16-F1R3WEE |
| LTA4 | 1 | tRF-16-F1R3WEE |
| piRNA | 3 | tRF-30-87R95RM3Y826, tRF-29-KY7SHRRNWJE2, tRF-18-F8DHXYD9 |
| regulatory element | 1 | tRF-18-F8DHXYD9 |
| tRNA | 5 | tRF-30-87R95RM3Y826, tRF-29-KY7SHRRNWJE2, tRF-19-DRXSE5I2, tRF-18-F8DHXYD9, tRF-16-F1R3WEE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strømme, O.; Heck, K.A.; Brede, G.; Lindholm, H.T.; Otterlei, M.; Arum, C.-J. tRNA-Derived Fragments as Biomarkers in Bladder Cancer. Cancers 2024, 16, 1588. https://doi.org/10.3390/cancers16081588
Strømme O, Heck KA, Brede G, Lindholm HT, Otterlei M, Arum C-J. tRNA-Derived Fragments as Biomarkers in Bladder Cancer. Cancers. 2024; 16(8):1588. https://doi.org/10.3390/cancers16081588
Chicago/Turabian StyleStrømme, Olaf, Kathleen A. Heck, Gaute Brede, Håvard T. Lindholm, Marit Otterlei, and Carl-Jørgen Arum. 2024. "tRNA-Derived Fragments as Biomarkers in Bladder Cancer" Cancers 16, no. 8: 1588. https://doi.org/10.3390/cancers16081588
APA StyleStrømme, O., Heck, K. A., Brede, G., Lindholm, H. T., Otterlei, M., & Arum, C. -J. (2024). tRNA-Derived Fragments as Biomarkers in Bladder Cancer. Cancers, 16(8), 1588. https://doi.org/10.3390/cancers16081588