
Enzymatic Pretreatment Improved the In Vitro Ruminal Degradability of Oil Palm Fronds

 ,
,  , , , , ,
, , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
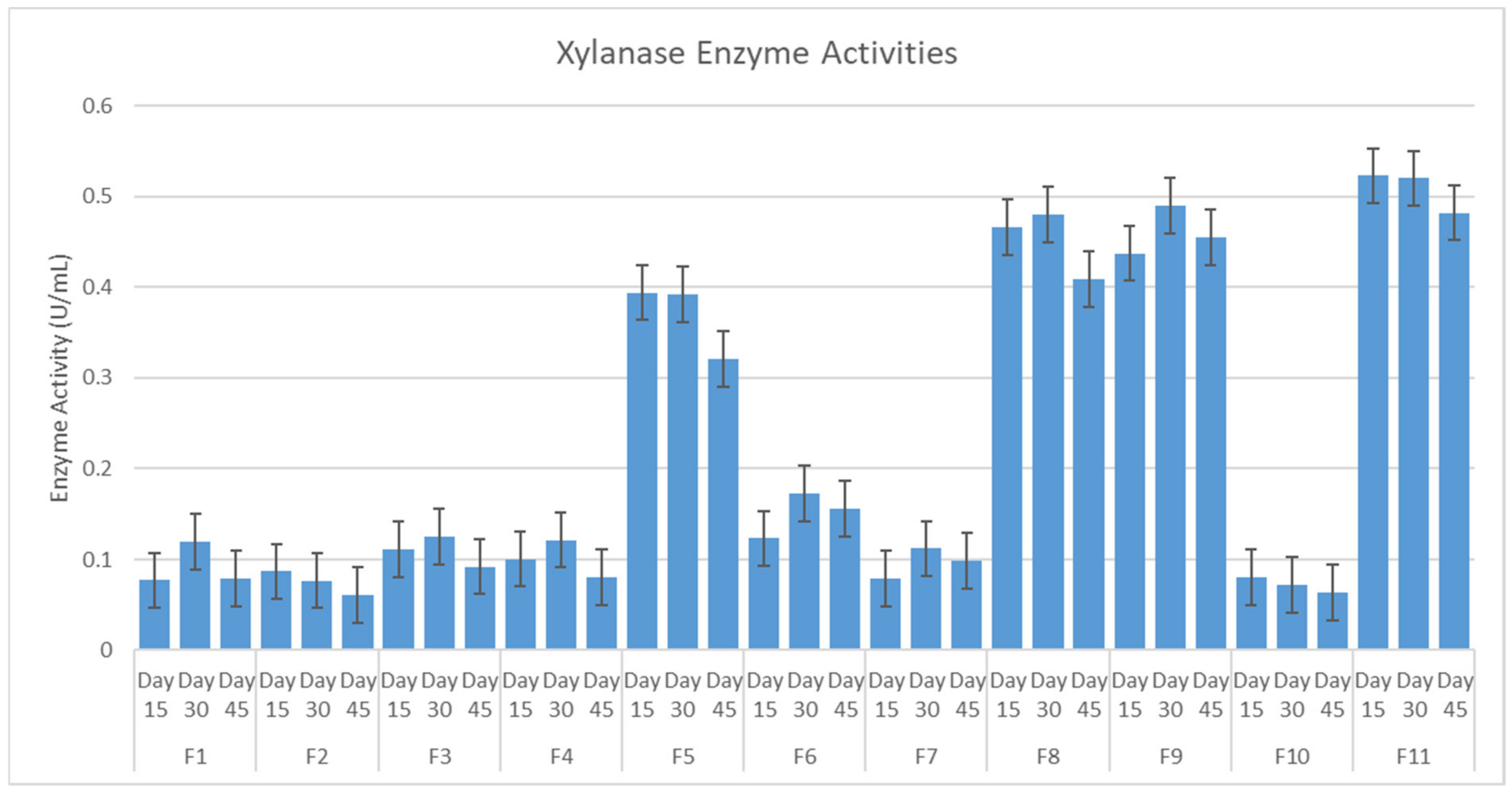
2.1. Isolation, Enzyme Activities and the Selection of Fungi
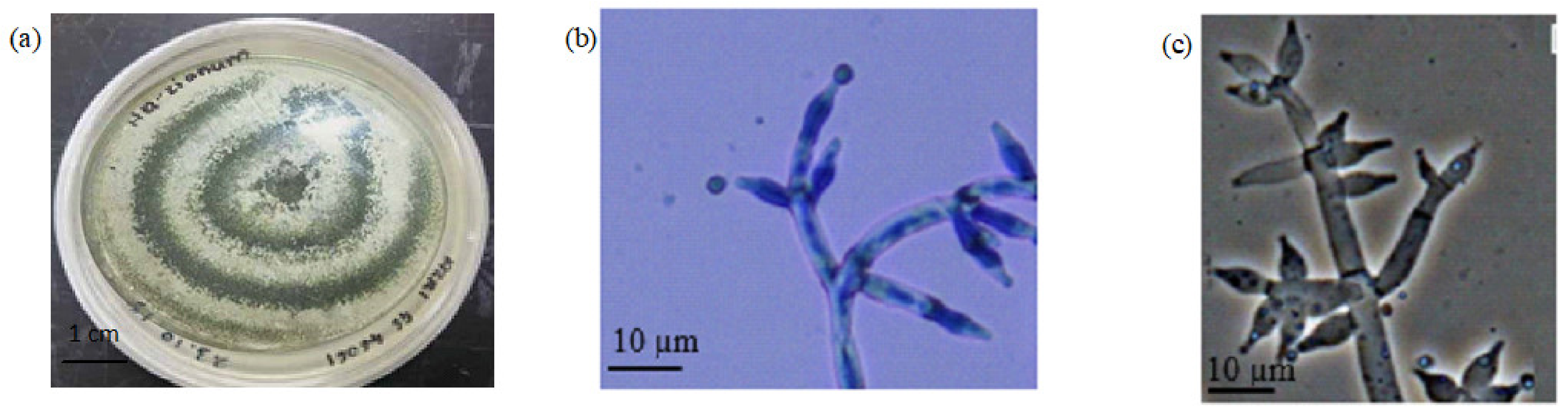
2.2. Morphological Observation and Molecular Identification of Selected Fungi
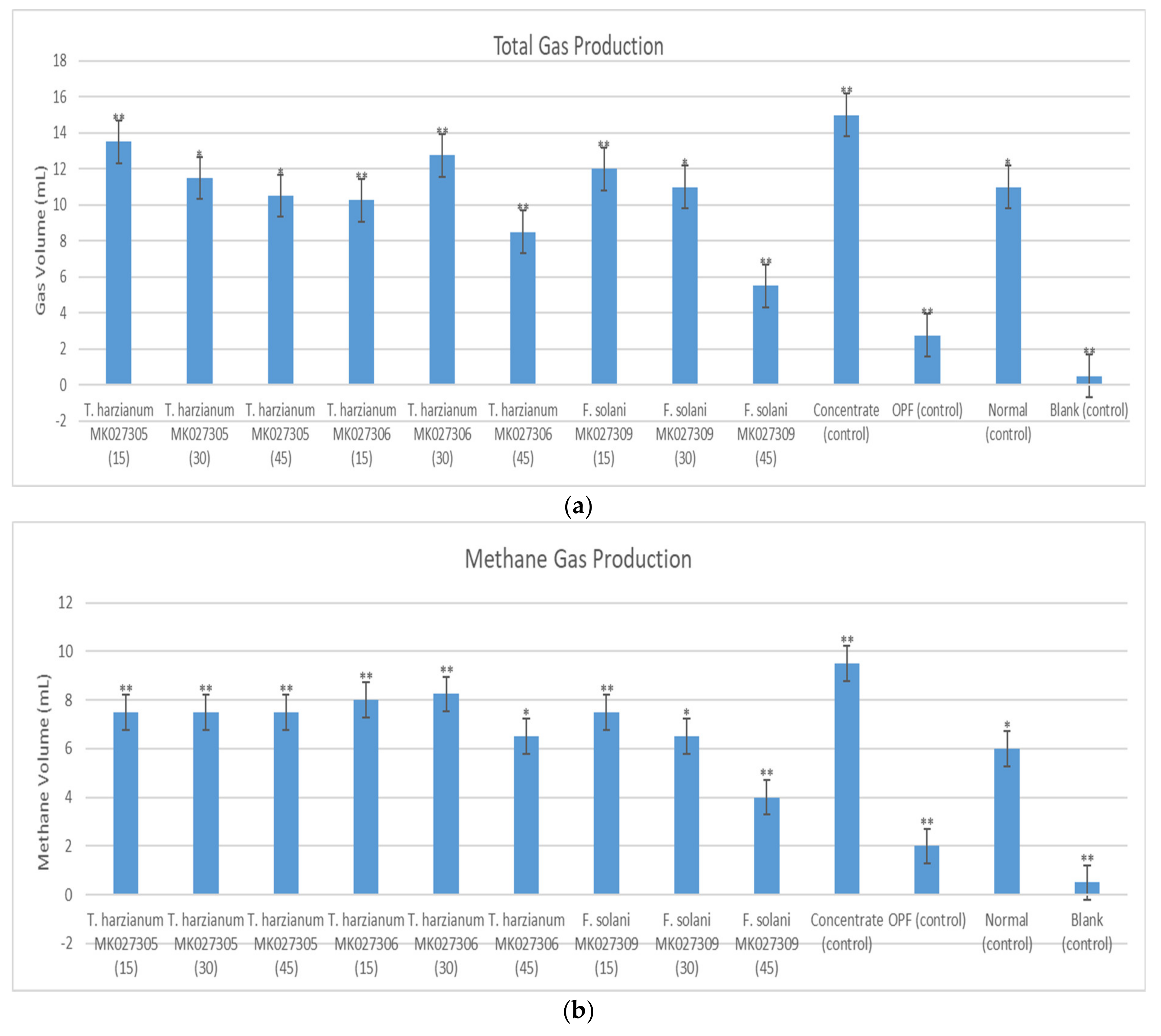
2.3. Total Gas, Methane, Volatile Fatty Acid Production and Apparent Rumen Degradable Carbohydrate
3. Discussion
4. Materials and Methods
4.1. Enzyme Extraction of Filamentous Fungi
4.2. Determination of Enzymes Activity
4.2.1. Lignolytic Enzymes
4.2.2. Cellulolytic Enzymes
4.2.3. Hemicellulolytic Enzyme
4.3. Selection of Fungi Isolate with Optimum Enzyme Activity
4.4. Identification of Selected Fungi Species
4.4.1. Morphological Identification
4.4.2. Molecular Identification
4.5. Pretreatment of OPFs with Enzyme Extracts
4.6. In Vitro Fermentation Trial
4.7. Volatile Fatty Acid Determination
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pakiam, R. Malaysia Keeps Palm Oil Export Tax Unchanged to Spur Shipments. Bloomberg. 2013. Available online: https://www.bloomberg.com/news/articles/2013-09-17/malaysia-keeps-palm-oil-export-tax-unchanged-to-spur-shipments (accessed on 31 August 2020).
- Mahlia, T.M.I.; Ismail, N.; Hossain, N.; Silitonga, A.S.; Shamsuddin, A.H. Palm oil and its wastes as bioenergy sources: A comprehensive review. Environ. Sci. Pollut. Res. Int. 2019, 26, 14849–14866. [Google Scholar] [CrossRef] [PubMed]
- Wan Zahari, M.; Farid, M.M. Oil palm by products as feeds for livestock in Malaysia. In Proceedings of the International Palm Oil Congress 2011 (PIPOC 2011), KLCC, Kuala Lumpur, Malaysia, 15–17 November 2011; pp. 1–21. [Google Scholar]
- Shanmuganvelu, S. Decision Support System in Livestock Production; Research Inaugural Lecture; Malaysian Agricultural Research and Development Institute (MARDI): Serdang, Malaysia, 2014. [Google Scholar]
- Loh, T.C. Livestock Production and the Feed Industry in Malaysia. Food and Agriculture Organization. 2002. Available online: http://www.fao.org/docrep/007/y5019e/y5019e0l.htm#bm21 (accessed on 31 August 2020).
- Sukri, S.M.; Rahman, R.A.; Illias, R.M.; Yaakob, H. Optimization of alkaline pretreatment conditions of oil palm fronds in improving the lignocelluloses contents for reducing sugar production. Biotechnol. Lett. 2014, 19, 9006–9018. [Google Scholar]
- Rahman, M.M.; Lourenço, M.; Hassim, H.A.; Baars, J.J.; Sonnenberg, A.S.; Cone, J.W.; De Boever, J.; Fievez, V. Improving ruminal degradability of oil palm fronds using white rot fungi. Anim. Feed Sci. Technol. 2011, 169, 157–166. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, Z.K. Microwave-assisted conversion of lignocellulosic biomass into furans in ionic liquid. Bioresour. Technol. Rep. 2010, 101, 1111–1114. [Google Scholar] [CrossRef]
- Binod, P.; Satyanagalakshmi, K.; Sindhu, R.; Janu, K.U.; Sukumaran, R.K.; Pandey, A. Short duration microwave assisted pretreatment enhances the enzymatic saccharification and fermentable sugar yield from sugarcane bagasse. Renew. Energy 2012, 37, 109–116. [Google Scholar] [CrossRef]
- Salami, S.A.; Luciano, G.; O’Grady, M.N.; Biondi, L.; Newbold, C.J.; Kerry, J.P.; Priolo, A. Sustainability of feeding plant by-products: A review of the implications for ruminant meat production. Anim. Feed Sci. Technol. 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Laser, M.; Schulman, D.; Allen, S.G.; Lichwa, J.; Antal, M.J.; Lynd, L.R. A comparison of liquid hot water and steam pretreatments of sugar cane bagasse for bioconversion to ethanol. Bioresour. Technol. Rep. 2009, 81, 33–44. [Google Scholar] [CrossRef]
- Rabelo, S.C.; Maciel, R.; Costa, A.C. Lime pretreatment of sugarcane bagasse for bioethanol production. Int. J. Appl. Biotechnol. Biochem. 2009, 153, 139–150. [Google Scholar] [CrossRef]
- Hassim, H.A.; Lourenço, M.; Goh, Y.M.; Baars, J.J.P.; Fievez, V. Rumen degradation of oil palm fronds is improved through pre-digestion with white rot fungi but not through supplementation with yeast or enzymes. Can. J. Anim. Sci. 2012, 92, 79–87. [Google Scholar] [CrossRef]
- Jang, S.; Kwon, S.L.; Lee, H.; Jang, Y.; Park, M.S.; Lim, Y.W.; Changmu, K.; Kim, J.J. New Report of Three Unrecorded Species in Trichoderma harzianum Species Complex in Korea. Mycobiology 2018, 46, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Kudur, M.H.; Prakash, P.Y.; Savitha, M. Fusarium solani causing quasi-invasive infection of the foot in an immunocompetent middle-aged man from South India. Indian J. Dermatol. 2013, 58, 241. [Google Scholar] [CrossRef]
- Levin, L.; Herrmann, C.; Papinutti, V.L. Optimization of lignocellulolytic enzyme production by the white-rot fungus Trametes trogii in solid-state fermentation using response surface methodology. Biochem. Eng. J. 2008, 39, 207–214. [Google Scholar] [CrossRef]
- Revankar, M.S.; Lele, S.S. Enhanced production of laccase using a new isolate of white rot fungus WR-1. Process Biochem. 2006, 41, 581–588. [Google Scholar] [CrossRef]
- Bonnarme, P.; Jeffries, T.W. Mn (II) regulation of lignin peroxidases and manganese-dependent peroxidases from lignin-degrading white rot fungi. J. Appl. Environ. Microbiol. 1990, 56, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Sulaiman, O.; Hashim, R.; Peng, L.C.; Singh, R.P. Evaluating biopulping as an alternative application on oil palm trunk using the white-rot fungus Trametes versicolor. Int. Biodeterior. Biodegrad. 2013, 82, 96–103. [Google Scholar] [CrossRef]
- Li, P.; Wang, H.; Liu, G.; Li, X.; Yao, J. The effect of carbon source succession on laccase activity in the co-culture process of Ganoderma lucidum and a yeast. Enzym. Microb. Technol. 2011, 48, 1–6. [Google Scholar] [CrossRef]
- Arora, D.S.; Chander, M.; Gill, P.K. Involvement of lignin peroxidase, manganese peroxidase and laccase in degradation and selective ligninolysis of wheat straw. Int. Biodeterior. Biodegrad. 2002, 50, 115–120. [Google Scholar] [CrossRef]
- Chang, A.J.; Fan, J.; Wen, X. Screening of fungi capable of highly selective degradation of lignin in rice straw. Int. Biodeterior. Biodegrad. 2012, 72, 26–30. [Google Scholar] [CrossRef]
- Alam, M.Z.; Muhammad, N.; Mahmat, M.E. Solid State Bioconversion. Am. J. Appl. Sci. 2005, 2, 569–572. [Google Scholar]
- Khan, I.A.; Butt, W.A.; Ali, S.; Qadeer, M.A. Effect of carbon and nitrogen sources on Xylanase production by mutant strain of Aspergillus niger GCBMX-45. J. Biol. Sci. 2002, 2, 143–144. [Google Scholar]
- Lynd, L.R.; Zhang, Y. Quantitative determination of cellulase concentration as distinct from cell concentration in studies of microbial cellulose utilization: Analytical framework and methodological approach. Biotechnol. Bioeng. 2002, 77, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Aslam, N.; Latif, F.; Rajoka, M.I.; Jamil, A. Molecular cloning of cellulase genes from Trichoderma harzianum. Front. Nat. Prod. Chem. 2005, 1, 73–75. [Google Scholar] [CrossRef]
- Malik, N.N.; Akhtar, M.W.; Naz, B.A. Production of cellulase enzymes by Trichoderma harzianum. In Proceedings of the PAEC-KFK. Symposium Workshop on Biotechnology in Agriculture and Energy, Faisalabad, Pakistan, 3–7 March 1986; p. 10. [Google Scholar]
- Old, K.M.; Schippers, B. Electron microscopical studies of chlamydospores of Fusarium solani f. cucurbitae formed in natural soil. Soil Biol. Biochem. 1973, 5, 613–620. [Google Scholar] [CrossRef]
- Astuti, W.D.; Wiryawan, K.G.; Wina, E.; Widyastuti, Y.; Suharti, S.; Ridwan, R. Effects of Selected Lactobacillus plantarum as Probiotic on in vitro Ruminal Fermentation and Microbial Population. Pak. J. Nutr. 2018, 17, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef] [Green Version]
- Morand-Fehr, P. Recent developments in goat nutrition and application: A review. Small Rumin. Res. 2002, 60, 25–43. [Google Scholar] [CrossRef]
- Ali, R.E.; Mansfield, S.D.; Saddler, S.D. Cellulases: Agents for Fiber Modification or Bioconversion? The effect of substrate accessibility on cellulose enzymatic hydrolyzability. Prog. Biotechnol. 2002, 21, 21–36. [Google Scholar]
- Khezri, A.; Rezayazdi, K.; Mesgaran, M.D.; Moradi-Sharbabk, M. Effect of different rumen-degradable carbohydrates on rumen fermentation, nitrogen metabolism and lactation performance of Holstein dairy cows. Asian Australas. J. Anim. Sci. 2009, 22, 651–658. [Google Scholar] [CrossRef]
- Borneman, W.S.; Hartley, R.D.; Morrison, W.H.; Akin, D.E.; Ljungdahl, L.G. Feruloyl and p-coumaroyl esterase from anaerobic fungi in relation to plant cell wall degradation. Appl. Microbiol. Biotechnol. 1990, 33, 345–351. [Google Scholar] [CrossRef]
- Wong, D.W.S. Feruloyl esterase, a key enzyme in biomass degradation. Appl. Biochem. Biotechnol. 2005, 133, 87–112. [Google Scholar] [CrossRef]
- Namoolnoy, P.; Phoolphundh, S.; Wongwicharn, A. Biodegradation of lignin in oil palm fronds by white rot fungi. Agric. Nat. Resour. 2011, 45, 254–259. [Google Scholar]
- Dijkstra, J. Production and absorption of volatile fatty acids in the rumen. Livest. Prod. Sci. 1994, 39, 61–69. [Google Scholar] [CrossRef]
- Pandey, A.; Selvakumar, P.; Soccol, C.R.; Nigam, P. Solid state fermentation for the production of industrial enzymes. Curr. Sci. 1999, 10, 149–162. [Google Scholar]
- Pandey, A. Solid-state fermentation. Biochem. Eng. J. 2003, 13, 81–84. [Google Scholar] [CrossRef]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.D.; Castel, V.; Rosales, M.; Rosales, M.; de Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food & Agriculture Org.: Rome, Italy, 2006. [Google Scholar]
- Dinis, M.J.; Bezerra, R.M.F.; Nunes, F.; Dias, A.A.; Guedes, C.V.; Ferreira, L.M.M.; Rodrigues, M.A.M. Modification of wheat straw lignin by solid state fermentation with white-rot fungi. Bioresour. Technol. Rep. 2009, 100, 4829–4835. [Google Scholar] [CrossRef] [Green Version]
- Kusai, N.A.; Ayob, Z.; Maidin, M.S.T.; Safari, S.; Ali, S.R.A. Characterization of fungi from different ecosystems of tropical peat in Sarawak, Malaysia. Rend. Lincei Sci. Fis. Nat. 2018, 29, 469–482. [Google Scholar] [CrossRef]
- Dereeper, A.; Audic, S.; Claverie, J.M.; Blanc, G. BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 2010, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, G.; Lama, A.; Rodríguez, R.; Jiménez, A.; Guillén, R.; Fernández-Bolaños, J. Olive stone an attractive source of bioactive and valuable compounds. Bioresour. Technol. Rep. 2008, 99, 5261–5269. [Google Scholar] [CrossRef]
- Demeyer, D.I. Quantitative Aspects of Microbial Metabolism in the Rumen and Hindgut. In Rumen Microbial Metabolism and Ruminant Digestion; Jouany., J.P., Ed.; INRA Editions: Paris, France, 1991; pp. 217–237. [Google Scholar]
- Kaik, Y. The bread and butter of statistical analysis “t-test”: Uses and misuses. Pak. J. Med. Sci. 2015, 31, 1558–1559. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azmi, M.A.; Nicholas, A.F.; Azmi, A.F.M.; Zainudin, M.H.M.; Yusof, M.T.; Zakaria, Z.; Mustapha, N.M.; Zamri Saad, M.; Rusli, N.D.; Hassim, H.A. Enzymatic Pretreatment Improved the In Vitro Ruminal Degradability of Oil Palm Fronds. Catalysts 2022, 12, 461. https://doi.org/10.3390/catal12050461
Azmi MA, Nicholas AF, Azmi AFM, Zainudin MHM, Yusof MT, Zakaria Z, Mustapha NM, Zamri Saad M, Rusli ND, Hassim HA. Enzymatic Pretreatment Improved the In Vitro Ruminal Degradability of Oil Palm Fronds. Catalysts. 2022; 12(5):461. https://doi.org/10.3390/catal12050461
Chicago/Turabian StyleAzmi, Mohammad Azri, Ahmad Fariz Nicholas, Amirul Faiz Mohd Azmi, Mohd Huzairi Mohd Zainudin, Mohd Termizi Yusof, Zunita Zakaria, Noordin Mohamed Mustapha, Mohd Zamri Saad, Nor Dini Rusli, and Hasliza Abu Hassim. 2022. "Enzymatic Pretreatment Improved the In Vitro Ruminal Degradability of Oil Palm Fronds" Catalysts 12, no. 5: 461. https://doi.org/10.3390/catal12050461
APA StyleAzmi, M. A., Nicholas, A. F., Azmi, A. F. M., Zainudin, M. H. M., Yusof, M. T., Zakaria, Z., Mustapha, N. M., Zamri Saad, M., Rusli, N. D., & Hassim, H. A. (2022). Enzymatic Pretreatment Improved the In Vitro Ruminal Degradability of Oil Palm Fronds. Catalysts, 12(5), 461. https://doi.org/10.3390/catal12050461