CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- -
- Enzymes are produced from raw materials that are inexpensive, readily available and renewable, which means that prices are not subject to large fluctuations.
- -
- Enzymes are essentially non-hazardous, non-toxic, biocompatible and biodegradable. In contrast, serious environmental costs are associated with mining precious metal catalysts and large price fluctuations are the norm. Moreover, significant costs are associated with the removal of traces of noble metals from end-products as is required by regulatory agencies in, for example, the pharmaceutical industry.
- -
- Enzymatic reactions can be performed in conventional reactors, without any need for specialized equipment, under mild conditions (ambient temperature and pressure, physiological pH) in an environmentally acceptable solvent (water).
- -
- Enzymatic reactions involving multifunctional molecules can proceed with high regio- and stereoselectivity and generally without the need for functional group activation and protection and deprotection steps. This affords synthetic routes that, compared with conventional organic syntheses, are more step economic, more energy efficient, generate less waste and provide products in exquisite stereochemical purities that are difficult to compete with.
- -
- Many more enzymes have been identified through (meta)genome mining, that is the in silico analysis of publicly accessible genome sequence data bases that have been generated as a result of next generation genome sequencing [16].
- -
- Advances in gene synthesis have enabled the synthesis of identified genes, ready for cloning into a host production organism, in a few weeks at relatively low cost. This has significantly reduced the cost of development and subsequent production of enzymes at industrial scale.
- -
- Advances in protein engineering, using directed evolution techniques [17,18], have enabled optimization of enzyme performance under the challenging conditions encountered in industrial-scale processes, namely, high (stereo)selectivities, activities and space-time yields with non-natural substrates at high substrate concentrations, in the presence of organic solvents.
2. Enzyme Immobilization
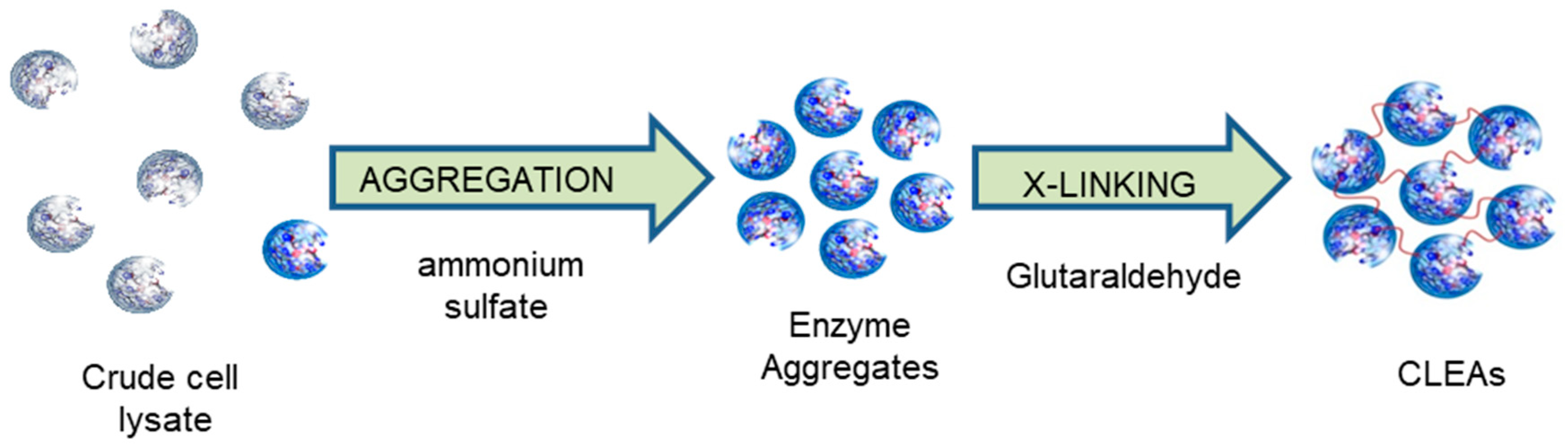
3. Cross-Linked Enzyme Aggregates (CLEAs)
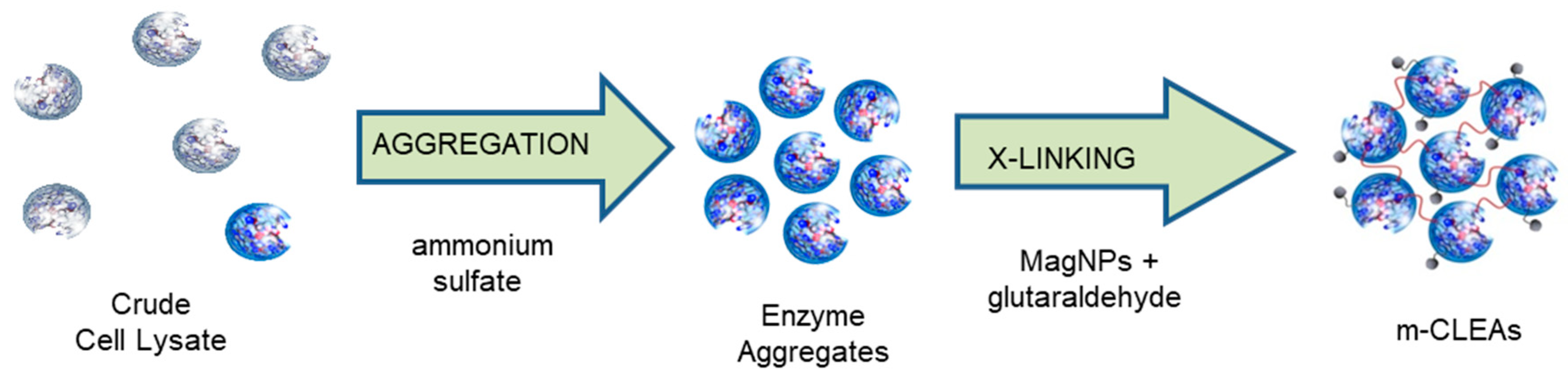
3.1. Cross-Linked Enzyme Aggregate (CLEA) Preparation
3.2. Effect of Additives
4. Scope of CLEA Technology
4.1. Hydrolase CLEAs
4.2. Oxidoreductase and Lyase CLEAs
5. Advantages, Limitations and Optimization of CLEAs
5.1. Bioimprinting
5.2. Reactor Configurations
5.3. Multi- and Combi-CLEAs
6. Magnetic CLEAs: The New Frontier
6.1. Preparation of m-CLEAs
6.2. Applications of m-CLEAs and m-Combi-CLEAs
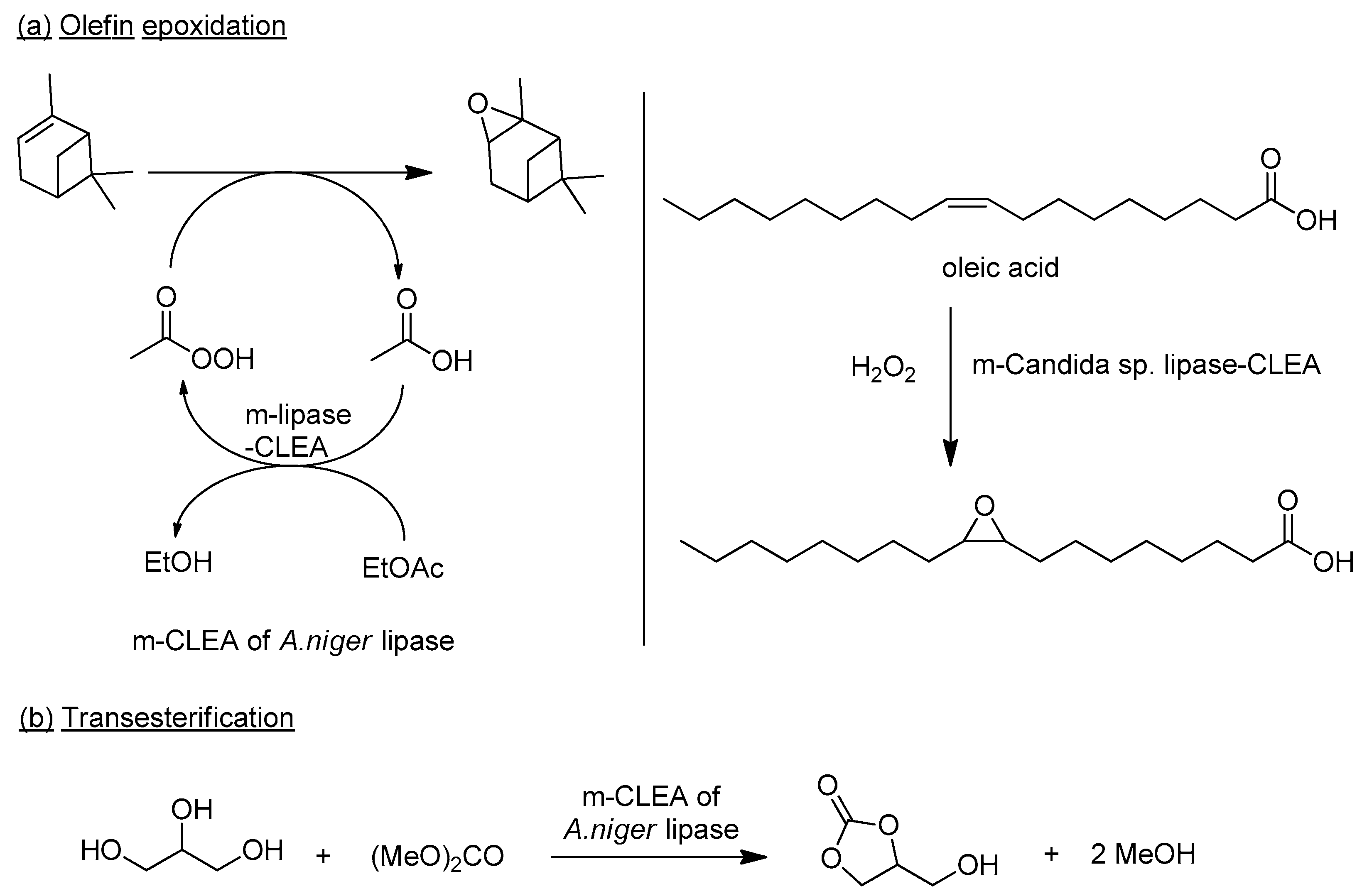
6.3. Lipase-m-CLEAs
6.4. Ligninolytic Enzymes and Waste Water Treatment
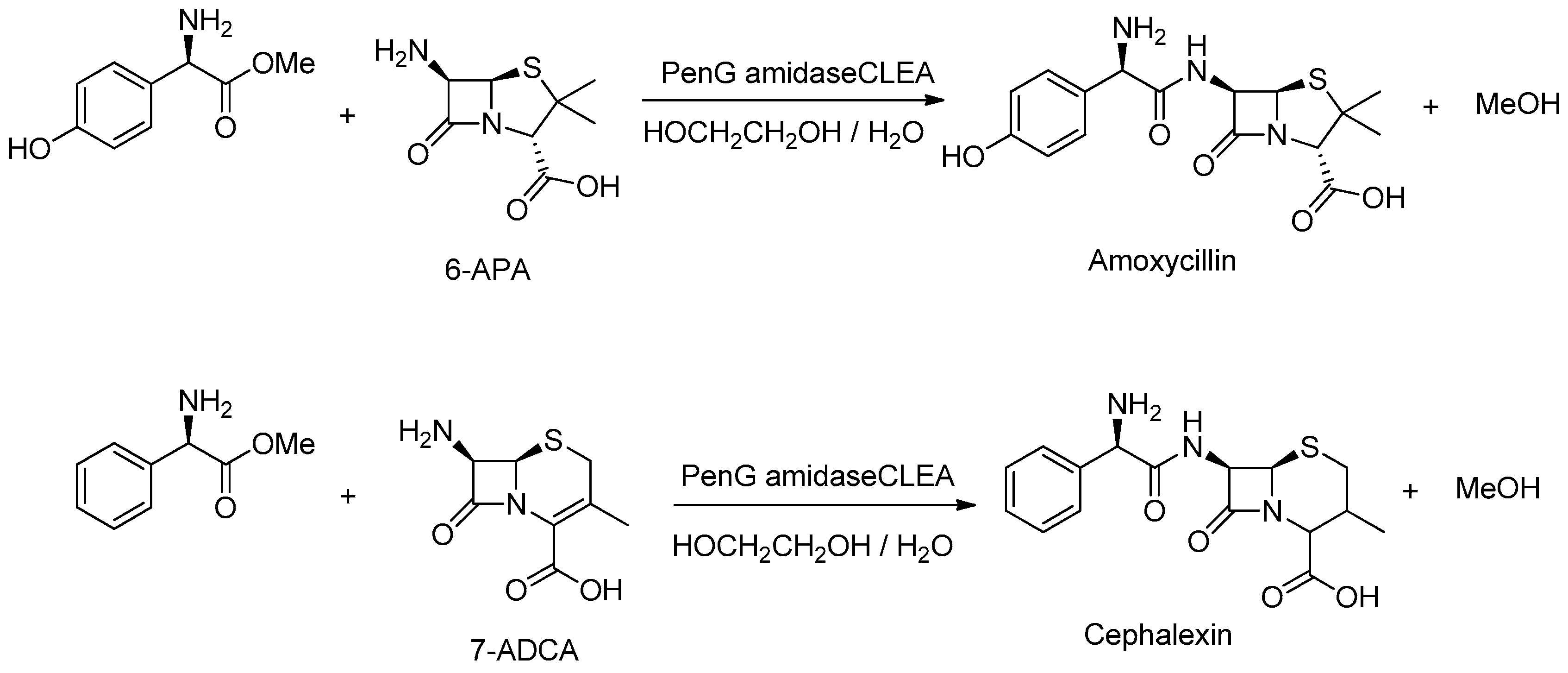
6.5. Synthesis of Semi-Synthetic Penicillin and Cephalosporin Antibiotics
6.6. Other Potential Applications
7. Conclusions and Outlook
Funding
Acknowledgments
Conflicts of Interest
References
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Kazlaukas, R.J. Biocatalysis for Green Chemistry and Chemical Process Development; Wiley: Hoboken, NJ, USA, 2011. [Google Scholar]
- Sheldon, R.A.; Brady, D. The limits to biocatalysis: Pushing the envelope. Chem. Commun. 2018, 54, 6088–6104. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis-key to sustainable chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Hu, J.-H. Biocatalysis in developments of green pharmaceutical processes. Curr. Opin. Chem. Biol. 2009, 13, 43–50. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis for the synthesis of pharmaceuticals. Bioorg. Med. Chem. 2018, 26, 1252–1274. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, J.-B.; Reetz, M.T. Biocatalysts for the pharmaceutical industry created by structure-guided directed evolution of stereoselective enzymes. Bioorg. Med. Chem. 2018, 26, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the synthesis of pharmaceuticals and pharmaceutical intermediates. Bioorg. Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, K.; Lütz, S. Recent developments and challenges of biocatalytic processes in the pharmaceutical industry. Curr. Opin. Green Sustain. Chem. 2018, 11, 58–64. [Google Scholar] [CrossRef]
- Huisman, G.W.; Collier, S.J. On the development of new biocatalytic processes for practical pharmaceutical synthesis. Curr. Opin. Chem. Biol. 2013, 17, 284–292. [Google Scholar] [CrossRef]
- Lalonde, J. Highly engineered biocatalysts for efficient small molecule pharmaceutical synthesis. Curr. Opin. Biotechnol. 2016, 42, 152–158. [Google Scholar] [CrossRef]
- Truppo, M.D. Biocatalysis in the Pharmaceutical Industry: The Need for Speed. ACS Med. Chem. Lett. 2017, 8, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Heux, S.; Meynial-Salles, I.; O’Donohue, M.J.; Dumon, C. White biotechnology: State of the art strategies for the development of biocatalysts for biorefining. Biotechnol. Adv. 2015, 33, 1653–1670. [Google Scholar] [CrossRef]
- Sheldon, R.A. Biocatalysis and Biomass Conversion in Alternative Reaction Media. Chem. Eur. J. 2016, 22, 1–17. [Google Scholar] [CrossRef]
- Sheldon, R.A. Chemicals from Renewable Biomass: A Renaissance in Carbohydrate Chemistry. Curr. Opin. Green Sustain. Chem. 2018, 14, 89–95. [Google Scholar] [CrossRef]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef]
- Reetz, M.T. Biocatalysis in Organic Chemistry and Biotechnology: Past, Present, and Future. J. Am. Chem. Soc. 2013, 135, 12480–12496. [Google Scholar] [CrossRef]
- Sun, Z.; Witmark, Y.; Bäckvall, J.-E.; Reetz, M.T. New Concepts for Increasing the Efficiency in Directed Evolution of Stereoselective Enzymes. Chem. Eur. J. 2016, 22, 5046–5054. [Google Scholar] [CrossRef]
- Madhavan, A.; Sindhu, R.; Binod, P.; Sukumaran, R.K.; Pandey, A. Strategies for design of improved biocatalysts for industrial applications. Bioresour. Technol. 2017, 245, 1304–1313. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- Bommarius, A.S.; Paye, M.F. Stabilizing Biocatalysts. Chem. Soc. Rev. 2013, 42, 6534–6565. [Google Scholar] [CrossRef]
- Thompson, M.P.; Peñafiel, I.; Cosgrove, S.C.; Turner, N.J. Biocatalysis Using Immobilized Enzymes in Continuous Flow for the Synthesis of Fine Chemicals. Org. Process Res. Dev. 2019, 23, 9–18. [Google Scholar] [CrossRef]
- Tamborini, L.; Fernandes, P.; Paradisi, F.; Molinari, F. Flow Bioreactors as Complementary Tools for Biocatalytic Process Intensification. Trends Biotechnol. 2018, 36, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Enzyme Immobilization: The Quest for Optimum Performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme Immobilization: Why, What and How? Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- DiCosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Immobilization Strategies to Improve the Performance of Enzymes. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Prasad, S.; Roy, I. Converting Enzymes into Tools of Industrial Importance. Recent Pat. Biotechnol. 2018, 12, 33–56. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J. Advances in enzyme immobilization. Biotechnol. Lett. 2009, 31, 1639–1650. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked Enzyme Aggregates(CLEAs) as Industrial Biocatalysts. In Biocatalysis: An Industrial Perspective; de Gonzalo, G., de Maria, P.D., Eds.; Royal Society of Chemistry: London, UK, 2018; Chapter 14. [Google Scholar]
- Sheldon, R.A. Multi-Step Enzyme Catalysis: Biotransformations and Chemoenzymatic Synthesi; Garcia-Junceda, E., Ed.; Wiley-VCH: Weinheim, Germany, 2008; pp. 109–135. [Google Scholar]
- Mateo, C.; Chmura, A.; Rustler, S.; van Rantwijk, F.; Stolz, A.; Sheldon, R.A. Synthesis of enantiomerically pure (S)-mandelic acid using an oxynitrilase-nitrilase bienzymatic cascade: A nitrilase surprisingly shows nitrile hydratase activity. Tetrahedron Asymmetry 2006, 17, 320–323. [Google Scholar] [CrossRef]
- Cao, L. Carrier-Bound Immobilized Enzymes, Principles, Applications, and Design; Wiley-VCH: Weinheim, Germany, 2005. [Google Scholar]
- Truppo, M.D.; Strotman, H.; Hughes, G. Development of an Immobilized Transaminase Capable of Operating in Organic Solvent. ChemCatChem 2012, 4, 1071–1074. [Google Scholar] [CrossRef]
- Cao, L.; van Langen, L.; Sheldon, R.A. Immobilized enzymes: Carrier-bound or carrier-free? Curr. Opin. Biotechnol. 2003, 14, 387–394. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Rantwijk, F.; van Langen, L.M.; Wegman, M.A.; Cao, L.; Janssen, M.H.A. Biocatalysts and Biocatalysis in the Synthesis of ß-Lactam Antibiotics. In Synthesis of β-Lactam Antibiotics: Chemistry, Biocatalysis and Process Integration; Bruggink, A., Ed.; Kluwer Academic: Dordrecht, The Netherlands, 2001; pp. 102–148. ISBN 0-7923-7060-0. [Google Scholar]
- Kallenberg, A.I.; van Rantwijk, F.; Sheldon, R.A. Immobilization of Penicillin G acylase: The Key to Optimum Performance. Adv. Synth. Catal. 2005, 347, 905–926. [Google Scholar] [CrossRef]
- Kosseva, M.R.; Panesar, P.S.; Kaur, G.; Kennedy, J.F. Use of immobilised biocatalysts in the processing of cheese whey. Int. J. Biol. Macromol. 2009, 45, 437–447. [Google Scholar] [CrossRef]
- Thompson, M.P.; Derrington, S.R.; Heath, R.S.; Porter, J.L.; Mangas-Sanchez, J.; Devine, P.N.; Truppo, M.D.; Turner, N.J. A generic platform for the immobilisation of engineered biocatalysts. Tetrahedron 2019, 75, 327–334. [Google Scholar] [CrossRef]
- Margolin, A.; Navia, M.A. Protein crystals as novel catalytic materials. Angew. Chem. Int. Ed. 2001, 40, 2204–2222. [Google Scholar] [CrossRef]
- Cao, L.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Enzyme Aggregates: A Simple and Effective Method for the Immobilization of Penicillin Acylase. Org. Lett. 2000, 2, 1361–1364. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEAs): Stable and recyclable biocatalysts. Biochem. Soc. Trans. 2007, 35, 1583. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Cross-Linked Enzyme Aggregates as Industrial Biocatalysts. Org. Proc. Res. Dev. 2011, 15, 213–223. [Google Scholar] [CrossRef]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Kiyota, Y.; Miyazaki, M. Techniques for Preparation of Cross-Linked Enzyme Aggregates and Their Applications in Bioconversions. Catalysts 2018, 8, 174. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Talekar, S.; Nadar, S.; Joshi, A.; Joshi, G. Pectin cross-linked enzyme aggregates (pectin-CLEAs) of glucoamylase. RSC Adv. 2014, 4, 59444–59453. [Google Scholar] [CrossRef]
- Weiser, D.; Varga, A.; Kovaca, K.; Nagy, F.; Szilagy, A.; Vertessy, B.G.; Paizs, C.; Poppe, L. Bisepoxide Cross-Linked Enzyme Aggregates—New Immobilized Biocatalysts for Selective Biotransformations. ChemCatChem 2014, 6, 1463–1469. [Google Scholar] [CrossRef]
- Pchelintsev, N.A.; Youshko, M.I.; Svedas, V.K. Quantitative characteristic of the catalytic properties and microstructure of cross-linked enzyme aggregates of penicillin acylase. J. Mol. Catal. B Enzym. 2009, 56, 202–207. [Google Scholar] [CrossRef]
- López-Gallego, F.; Betancor, L.; Hidalgo, A.; Alonso, N.; Fernández-Lafuente, R.; Guisán, J.M. Co-aggregation of Enzymes and Polyethyleneimine: A Simple Method to Prepare Stable and Immobilized Derivatives of Glutaryl Acylase. Biomacromolecules 2005, 6, 1839–1842. [Google Scholar] [CrossRef]
- Wilson, L.; Illanes, A.; Abian, O.; Pessela, B.C.C.; Fernandez-Lafuenta, R.; Guisan, J.M. Co-Aggregation of Penicillin G Acylase and Polyionic Polymers: An Easy Methodology To Prepare Enzyme Biocatalysts Stable in Organic Media. Biomacromolecules 2005, 5, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, B.K.; Kumar, S.S.; Golegaonkar, S.B.; Nene, S.N. Preparation of cross-linked enzyme aggregates of L-aminoacylase via co-aggregation with polyethyleneimine. J. Mol. Catal. B Enzym. 2012, 74, 184–191. [Google Scholar] [CrossRef]
- Yan, J.; Gui, X.; Wang, G.; Yan, Y. Improving and Activity of Cross-linked Enzyme Aggregates Based on Polyethylenimine in Hydrolysis of Fish Oil for Enrichment of Polyunsaturated Fatty Acids. Appl. Biochem. Biotechnol. 2012, 166, 925–932. [Google Scholar] [CrossRef]
- Perez, D.I.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Enzyme Aggregates of Chloroperoxidase: Synthesis, Optimization and Characterization. Adv. Synth. Catal. 2009, 351, 2133–2139. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Miyazaki, M.; Asanomi, Y.; Maeda, H. Poly-lysine supported cross-linked enzyme aggregates with efficient enzymatic activity and high operational stability. Catal. Sci. Technol. 2011, 1, 1256–1261. [Google Scholar] [CrossRef]
- Shah, S.; Sharma, A.; Gupta, M.N. Preparation of cross-linked enzyme aggregates by using bovine serum albumin as a proteic feeder. Anal. Biochem. 2006, 351, 207–213. [Google Scholar] [CrossRef]
- Dong, T.; Zhao, L.; Huang, Y.; Tan, X. Preparation of cross-linked aggregates of aminoacylase fron Aspergillus melleus by using bovine serum albumin as an inert additive. Bioresour. Technol. 2010, 101, 6569–6571. [Google Scholar] [CrossRef] [PubMed]
- Karimpil, J.J.; Melo, J.S.; D‘Souza, S.F. Hen egg white as a feeder protein for lipase immobilization. J. Mol. Catal. B Enzym. 2011, 71, 113–118. [Google Scholar] [CrossRef]
- Goetze, D.; Foletto, E.F.; da Silva, H.B.; Vitória, C.C.; Silveira, V.C.C.; Dal Magro, L.; Rodrigues, R.C. Effect of feather meal as a proteic feeder on combi-CLEA preparation for grape fruit clarification. Proc. Biochem. 2017, 62, 122–127. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S.; Kanbak-Aksu, S.; Rasmussen, J.; Janssen, M.H.A. Cross-Linked Enzyme Aggregates in Organic Synthesis. Aldrichim. Acta 2013, 46, 81–93. [Google Scholar]
- Sheldon, R.A. Industrial Applications of Asymmetric Synthesis using Cross-Linked Enzyme Aggregates. In Comprehensive Chirality; Carreira, E.M., Yamamoto, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 9, pp. 353–366. [Google Scholar]
- GuauqueTorres, M.P.; Foresti, M.L.; Ferreira, M.L. Cross-linked enzyme aggregates (CLEAs) of selected lipases: A procedure for the proper calculation of their recovered activity. AMB Express 2013, 3, 25. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, S.; Khanahmadi, S. Optimizing the preparation conditions and characterization of a stable and recyclable cross-linked enzyme aggregate (CLEA)-protease. Bioresour. Bioprocess. 2016, 3, 3. [Google Scholar] [CrossRef]
- Montoro-Garcia, S.; Gil-Ortiz, F.; Navarro-Fernandez, J.; Rubio, V.; Garcia-Carmona, F.; Sanchez-Ferrer, A. Improved cross-linked enzyme aggregates for the production of desacetyl beta-lactam antibiotics intermediates. Bioresour. Technol. 2009, 101, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzalez, L.; Garcia-Carmona, F. Characterization of cross-linked immobilized arylesterase from Gluconobacter oxydans 621H with activity toward cephalosporin C and 7-aminocephalosporanic acid. Biotechnol. Prog. 2016, 32, 36–42. [Google Scholar] [CrossRef]
- Park, H.J.; Uhm, K.N.; Kim, H.K. Biotransformation of amides to acids using a co-cross-linked enzyme aggregate of Rhodococcus erythropolis amidase. J. Microbiol. Biotechnol. 2010, 20, 325–331. [Google Scholar]
- Malandra, A.; Cantarella, M.; Kaplan, O.; Vejvoda, V.; Uhnakova, B.; Stepankova, B.; Kubac, D.; Martinkova, L. Continuous hydrolysis of 4-cyanopyridine by nitrilases from Fusarium solani O1 and Aspergillus niger K10. Appl. Microbiol. Biotechnol. 2009, 85, 277–284. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Li, X.-F.; Lou, W.-Y.; Zong, M.H. Cross-linked enzyme aggregates of Mung bean epoxide hydrolases: A highly active, stable and recyclable biocatalyst for asymmetric hydrolysis of epoxides. J. Biotechnol. 2013, 166, 12–19. [Google Scholar] [CrossRef]
- Cruz, J.; Barbosa, O.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Rodrigo Torres, R.; Claudia Ortiz, C. Optimized preparation of CALB-CLEAs by response surface methodology: The necessity to employ a feeder to have an effective crosslinking. J. Mol. Catal. B Enzym. 2012, 80, 7–14. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. CLEAs of Candida antarctica lipase B (CaLB) with a bovine serum albumin (BSA) cofeeder core: Study of their catalytic activity. Biochem. Eng. J. 2014, 90, 36–43. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. Effect of different parameters on the hydrolytic activity of cross-linked enzyme aggregates (CLEAs) of lipase from Thermomyces lanuginose. Biochem. Eng. J. 2013, 72, 18–23. [Google Scholar] [CrossRef]
- Lai, J.-Q.; Hu, Z.-L.; Sheldon, R.A.; Yang, Z. Catalytic performance of cross-linked enzyme aggregates of Penicillium expansum lipase and their use as catalyst for biodiesel production. Proc. Biochem. 2012, 47, 2058–2063. [Google Scholar] [CrossRef]
- Rehman, S.; Bhatti, H.N.; Bilal, M.; Asgher, M. Cross-linked enzyme aggregates (CLEAs) of Pencilluim notatum lipaseenzyme with improved activity, stability and reusabilitycharacteristics. Int. J. Biol. Macromol. 2016, 91, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Lozano, S.; López-Gallego, F.; Rocha-Martin, J.; Guisán, J.M.; Favela-Torres, E. Improving enantioselectivity of lipase from Candida rugosa bycarrier-bound and carrier-free immobilization. J. Mol. Catal. B Enzym. 2016, 130, 32–39. [Google Scholar] [CrossRef]
- Chavez, G.; Rasmussen, J.-A.; Janssen, M.H.A.; Mamo, G.; Hatti-Kaul, R.; Sheldon, R.A. Baeyer-Villiger Oxidation of Cyclohexanone in Aqueous Medium with In Situ Generation of Peracid Catalyzed by Perhydrolase CLEA. Top. Catal. 2014, 57, 349–355. [Google Scholar] [CrossRef]
- Schroeder, H.; Strohmeier, G.A.; Leypold, M.; Nuijens, T.; Quaedflieg, P.J.L.M.; Breinbauer, R. Racemization-Free Chemoenzymatic Peptide Synthesis Enabled by the Ruthenium-Catalyzed Synthesis of Peptide Enol Esters via Alkyne-Addition and Subsequent Conversion Using Alcalase-Cross-Linked Enzyme Aggregates. Adv. Synth. Catal. 2013, 355, 1799–1807. [Google Scholar] [CrossRef]
- Illanes, A.; Wilson, L.; Caballero, E.; Fernandez-Lafuenta, R.; Guisan, J.M. Crosslinked penicillin acylase aggregates for synthesis of β-lactam antibiotics in organic medium. Appl. Biochem. Biotechnol. 2006, 133, 189–202. [Google Scholar] [CrossRef]
- Illanes, A.; Wilson, L.; Altamirano, C.; Cabrera, Z.; Alvarez, L.; Aguirre, C. Production of cephalexin in organic medium at high substrate concentrations with CLEA of penicillin acylase and PGA-450. Enzym. Microb. Technol. 2007, 40, 195–203. [Google Scholar] [CrossRef]
- Contesini, F.; de Alencar Figueira, J.; Kawaguti, H.; de Barros Fernandes, P.; de Oliveira Carvalho, P.; da Graça Nascimento, M.; Sato, H. Potential Applications of Carbohydrases Immobilization in the Food Industry. Int. J. Mol. Sci. 2013, 14, 1335–1369. [Google Scholar] [CrossRef] [Green Version]
- Gauer, R.; Paul, H.; Jain, R.; Khare, S.K. Galacto-oligosaccharide synthesis by immobilized Aspergillus oryzae β-galactosidase. Food Chem. 2006, 97, 426–430. [Google Scholar] [CrossRef]
- Li, L.; Li, G.; Cao, L.; Ren, G.; Kong, W.; Wang, S.; Guo, G.; Liu, Y. Characterization of the cross-linked enzyme aggregates of a novel β-galactosidase, a potential catalyst for the synthesis of galacto-oligosaccharides. J. Agric. Food Chem. 2015, 63, 894–901. [Google Scholar] [CrossRef]
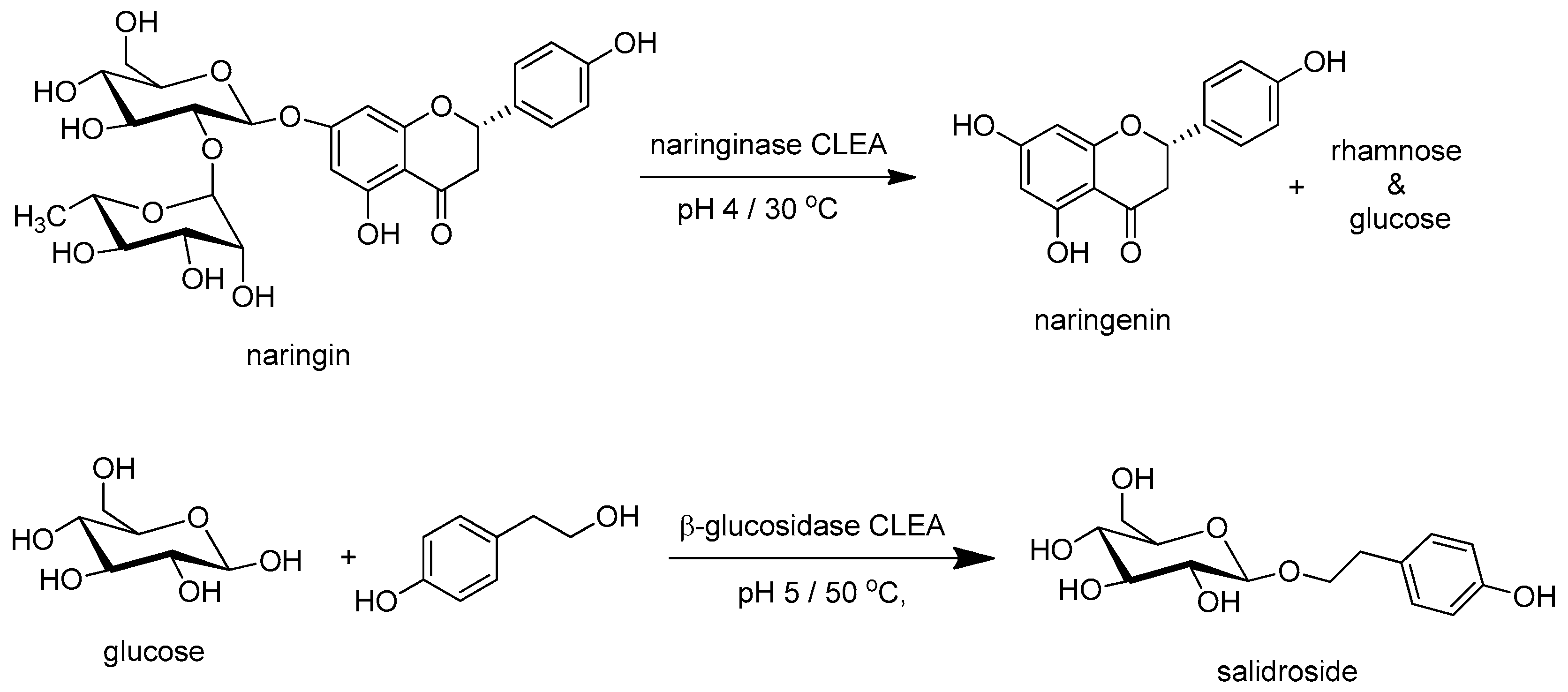
- Ribeiro, M.H.L.; Rabaça, M. Cross-Linked Enzyme Aggregates of Naringinase: Novel Biocatalysts for Naringin Hydrolysis. Enzym. Res. 2011, 2011, 851272. [Google Scholar] [CrossRef]
- Chen, L.; Hu, Y.-D.; Li, N.; Zong, M.-H. Cross-linked enzyme aggregates of β-glucosidase from Prunus domestica seeds. Biotechnol. Lett. 2012, 34, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
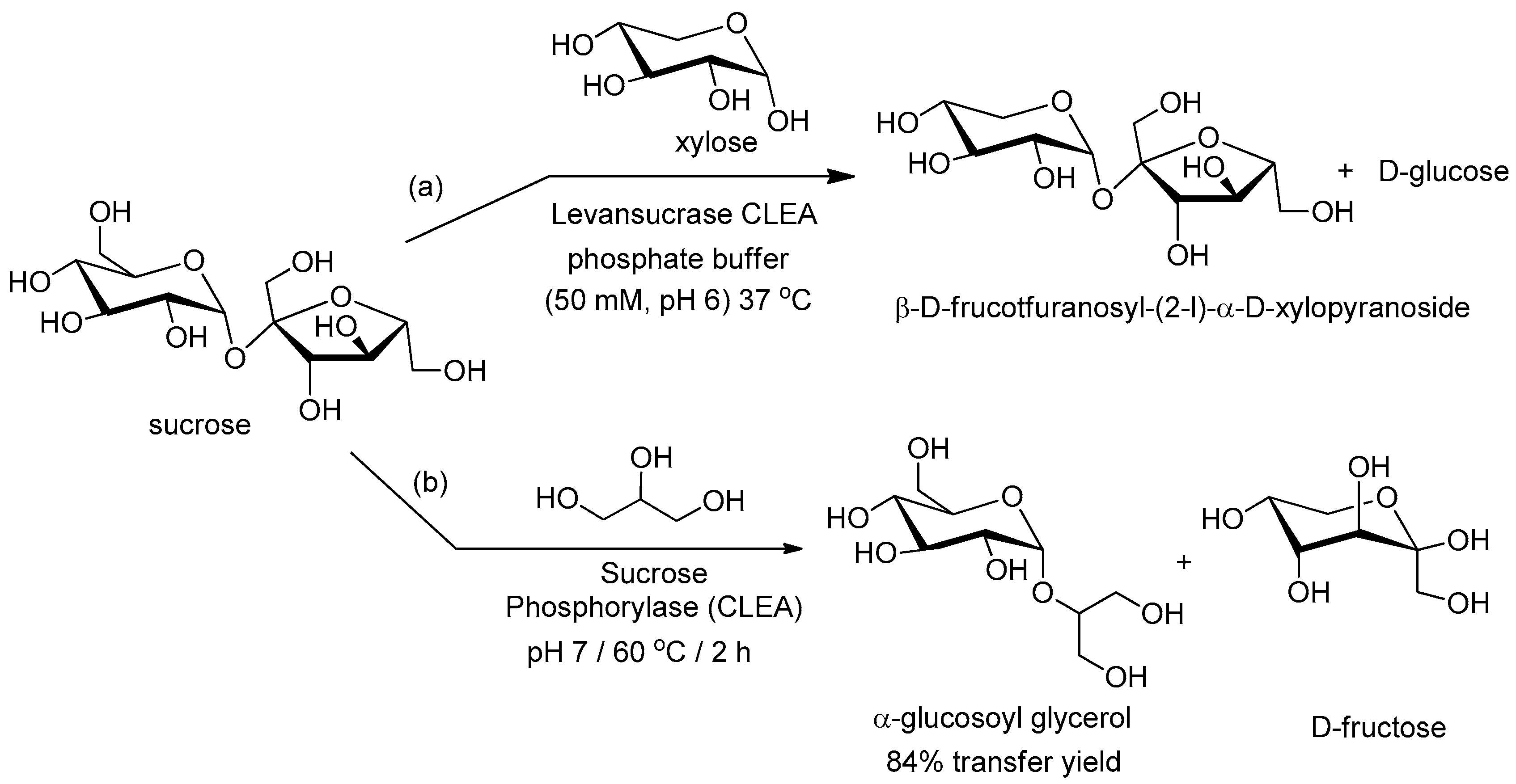
- Ortiz-Soto, M.E.; Rudiño-Piñera, E.; Rodriguez-Alegria, M.E.; Manguia, A.L. Evaluation of cross-linked aggregates from purified Bacillus subtilis levansucrase mutants for transfructosylation reactions. BMC Biotechnol. 2009, 9, 68. [Google Scholar] [CrossRef]
- Cerdobbel, A.; De Winter, K.; Desmet, T.; Soetaert, W. Sucrose phosphorylase as cross-linked enzyme aggregate: Improved thermal stability for industrial applications. Biotechnol. J. 2010, 5, 1192–1197. [Google Scholar] [CrossRef] [Green Version]
- De Winter, K.; Soetaert, W.; Desmet, T. An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity. Int. J. Mol. Sci. 2012, 13, 11333–11342. [Google Scholar] [CrossRef] [Green Version]
- Talekar, S.; Ghodake, V.; Kate, A.; Samant, N.; Kumar, C.; Gadagkar, S. Preparation and Characterization of Cross-linked Enzyme Aggregates of Saccharomyces Cerevisiae Invertase. Aust. J. Basic Appl. Sci. 2010, 4, 4760–4765. [Google Scholar]
- Diaz-Rodriguez, A.; Lavandera, I.; Kanbak-Aksu, S.; Sheldon, R.A.; Gotor, V.; Gotor-Fernandez, V. From Diols to Lactones under Aerobic Conditions Using a Laccase/TEMPO Catalytic System in Aqueous Medium. Adv. Synth. Catal. 2012, 354, 3405–3408. [Google Scholar] [CrossRef]
- Matijosyte, I.; Arends, I.W.C.E.; de Vries, S.; Sheldon, R.A. Preparation and use of cross-linked enzyme aggregates (CLEAs) of laccase. J. Mol. Catal. B Enzym. 2010, 62, 142–148. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, Y.-W. Optimizing the preparation conditions and characterization of cross-linked enzyme aggregates of a monoamine oxidase. Food Sci. Biotechnol. 2016, 25, 1421–1425. [Google Scholar] [CrossRef]
- Roberge, C.; Amos, D.; Pollard, D.; Devine, P. Preparation and application of cross-linked aggregates of chloroperoxidase with enhanced hydrogen peroxide tolerance. J. Mol. Catal. B Enzym. 2009, 56, 41–45. [Google Scholar] [CrossRef]
- Sulek, F.; Perez Fernandez, D.; Kneza, Z.; Habulina, M.; Sheldon, R.A. Immobilization of horseradish peroxidase as crosslinked enzyme aggregates (CLEAs). Process Biochem. 2011, 46, 765–769. [Google Scholar] [CrossRef]
- Morales, A.; Barbosa, O.; Rueda, N.; Fonseca, Z.; Torres, R.; Rodriguez, R.C.; Ortiz, C.; Fernandez-Lafuenta, R. Optimization and characterization of CLEAs of the very thermostable dimeric peroxidase from Roystonea regia. RSC Adv. 2015, 5, 53047–53053. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, S.; Kim, Y.H.; Won, K.; Lee, S.H. Immobilization of formate dehydrogenase from Candida boidinii through cross-linked enzyme aggregates. J. Mol. Catal. B Enzym. 2013, 97, 209–214. [Google Scholar] [CrossRef]
- Sinirlioglu, Z.A.; Sinirlioglu, D.; Akbas, F. Preparation and characterization of stable cross-linked enzyme aggregates of novel laccase enzyme from Shewanella putrefaciens and using malachite green decolorization. Bioresour. Technol. 2013, 146, 807–811. [Google Scholar] [CrossRef]
- Ba, S.; Arsenault, A.; Hassani, T.; Jones, J.P.; Cabana, H. Laccase immobilization and insolubilization: From fundamentals to applications for the elimination of emerging contaminants in wastewater treatment. Crit. Rev. Biotechnol. 2013, 33, 404–418. [Google Scholar] [CrossRef]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Preparation and characterization of cross-linked laccase aggregates and their application to the elimination of endocrine disrupting chemicals. J. Biotechnol. 2007, 132, 23–31. [Google Scholar] [CrossRef] [PubMed]
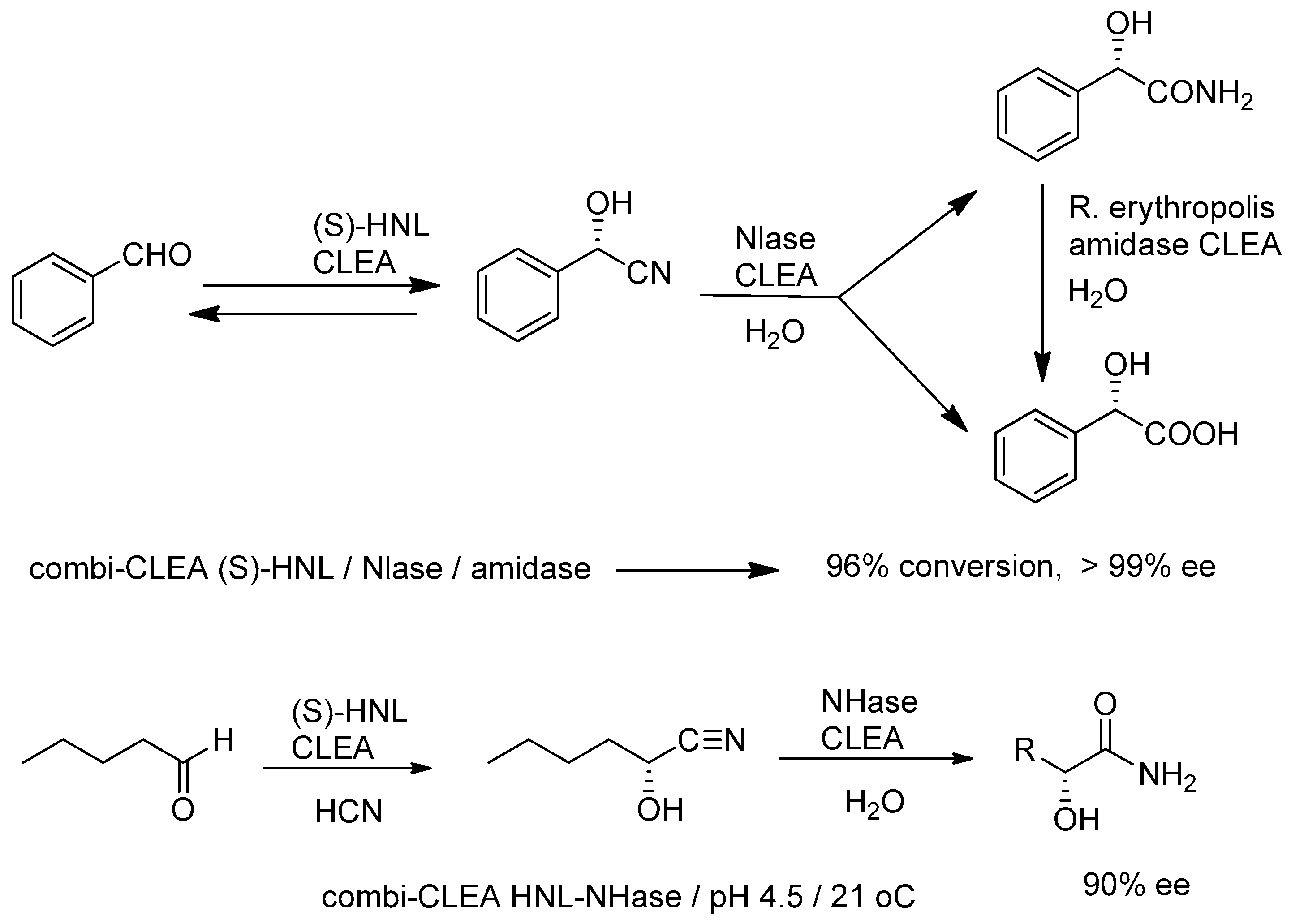
- Cabirol, F.L.; Lim, A.E.C.; Hanefeld, U.; Sheldon, R.A. Straightforward Enzymatic Process Based on HNL CLEA-Catalysis towards Cyanohydrin Derivatives. Org. Proc. Res. Dev. 2010, 14, 114–118. [Google Scholar] [CrossRef]
- Alagoz, D.; Tukel, S.S.; Yildirim, D. Enantioselective Synthesis of Various Cyanohydrins Using Covalently Immobilized Preparations of Hydroxynitrile Lyase from Prunus dulcis. Appl. Biochem. Biotechnol. 2015, 177, 1348–1363. [Google Scholar] [CrossRef]
- Yildirim, D.; Tuekel, S.S.; Alagoez, D. Crosslinked enzyme aggregates of hydroxynitrile lyase partially purified from Prunus dulcis seeds and its application for the synthesis of enantiopure cyanohydrins. Biotechnol. Prog. 2014, 30, 818–826. [Google Scholar] [CrossRef]
- Cabirol, F.L.; Hanefeld, U.; Sheldon, R.A. Immobilized hydroxynitrile lyases for enantioselective ynthesis of cyanohydrins: Sol-gels and cross-linked enzyme aggregates. Adv. Synth. Catal. 2006, 348, 1645–1654. [Google Scholar] [CrossRef]
- Chmura, A.; van der Kraan, G.M.; Kielar, F.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. Cross-linked aggregates of the hydroxynitrile lyase from Manihot esculenta: Highly active and robust Biocatalysts. Adv. Synth. Catal. 2006, 348, 1655–1661. [Google Scholar] [CrossRef]
- Cabirol, F.L.; Tan, P.L.; Tay, B.; Cheng, S.; Hanefeld, U.; Sheldon, R.A. Linum usitatissimum hydroxynitrile lyase cross-linked enzyme aggregates: A recyclable enantioselective catalysis. Adv. Synth. Catal. 2008, 350, 2329–2338. [Google Scholar] [CrossRef]
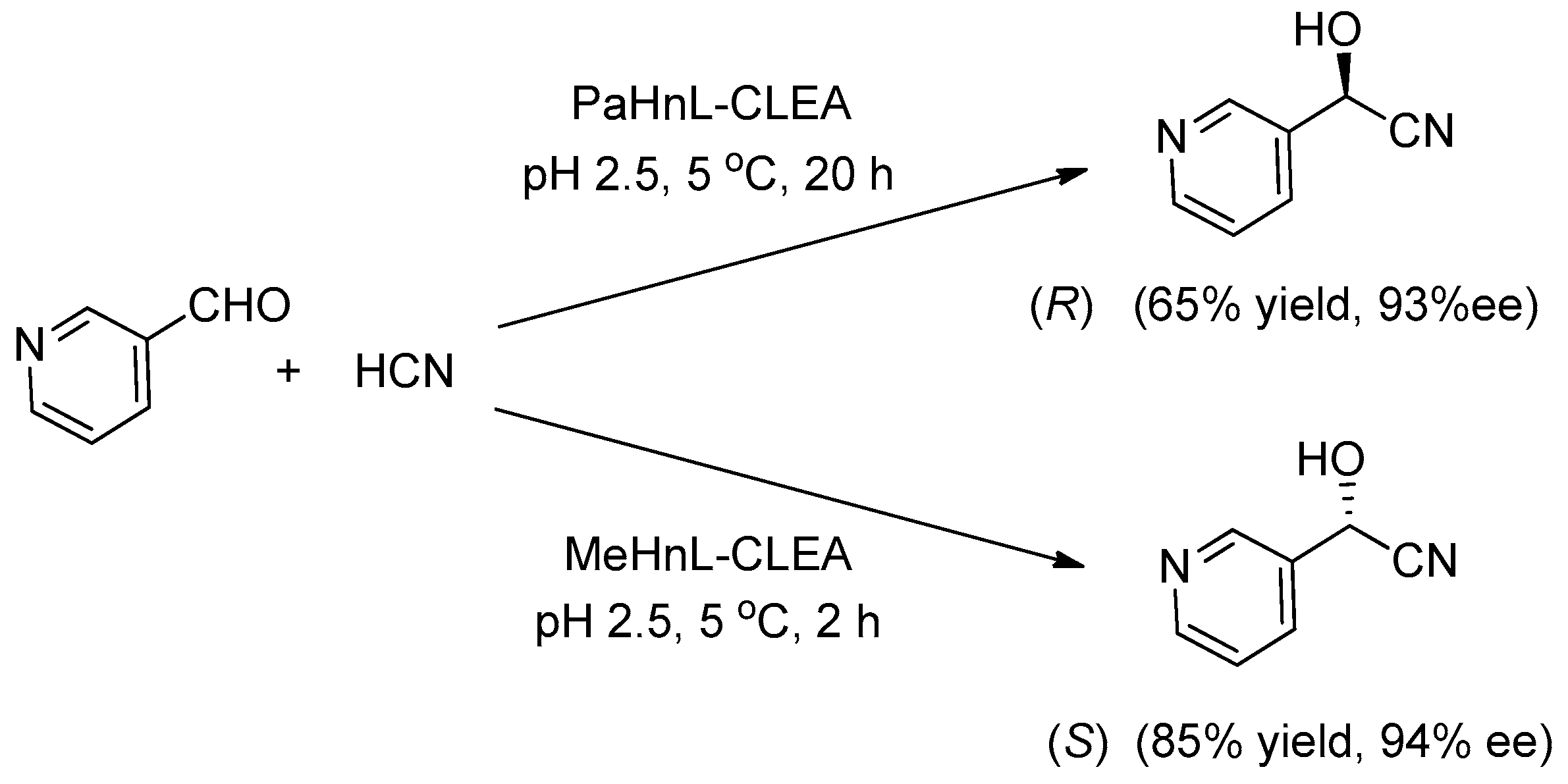
- Roberge, C.; Fleitz, F.; Pollard, D.; Devine, P. Asymmetric synthesis of cyanohydrin derived from pyridine aldehyde with cross-linked aggregates of hydroxynitrile lyases. Tetrahedron Lett. 2007, 48, 1473–1477. [Google Scholar] [CrossRef]
- Lanfranchi, E.; Grill, B.; Raghoeber, Z.; Van Pelt, S.; Sheldon, R.A.; Steiner, K.; Glieder, A.; Winkler, M. Production of hydroxynitrile lyase from davallia tyermannii (DtHNL) in Komagataella phaffii and its immobilization as a CLEA to generate a robust biocatalyst. ChemBioChem 2018, 19, 312–316. [Google Scholar] [CrossRef]
- Jangir, N.; Padhi, S.K. Immobilized Baliospermum montanum hydroxynitrile lyase catalyzed suynthesis of chiral cyanohydrins. Bioorg. Chem. 2019, 84, 32–40. [Google Scholar] [CrossRef]
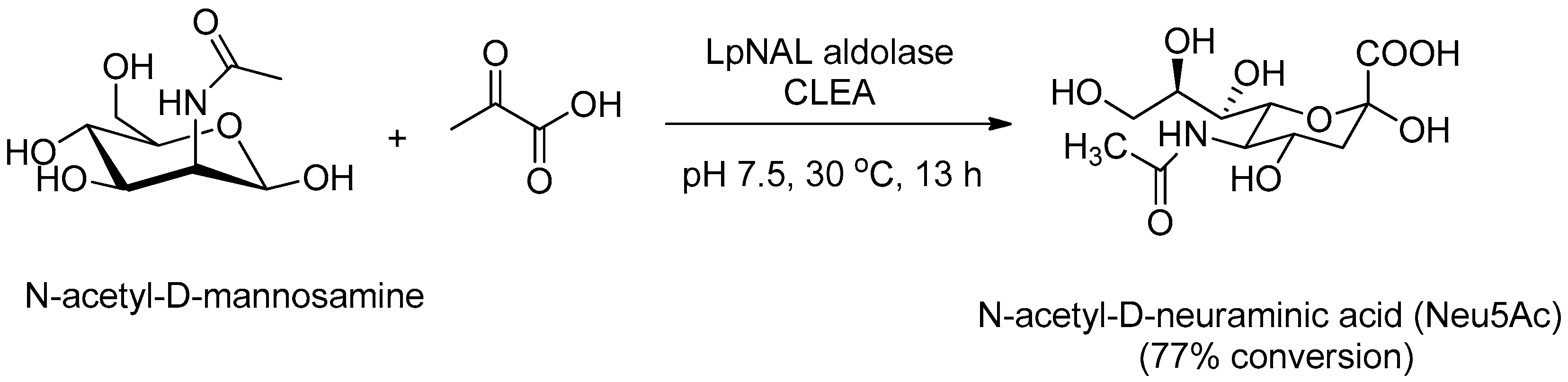
- Garcia-Garcia, M.I.; Sola-Carvajal, A.; Garcia Carmona, F.; Sanchez Ferrer, A. Improved production of 2-keto-3-deoxy-d-glycero-galactononulosonic acid (KDN) using FastPrep-CLEAs. Process Biochem. 2014, 49, 90–94. [Google Scholar] [CrossRef]
- Garcia-Garcia, M.I.; Sola-Carvajal, A.; Sanchez-Carron, G.; Garcia-Carmona, F.; Sanchez-Ferrer, A. New stabilized FastPrep-CLEAs for sialic acid synthesis. Bioresour. Technol. 2011, 102, 6186–6191. [Google Scholar] [CrossRef] [PubMed]
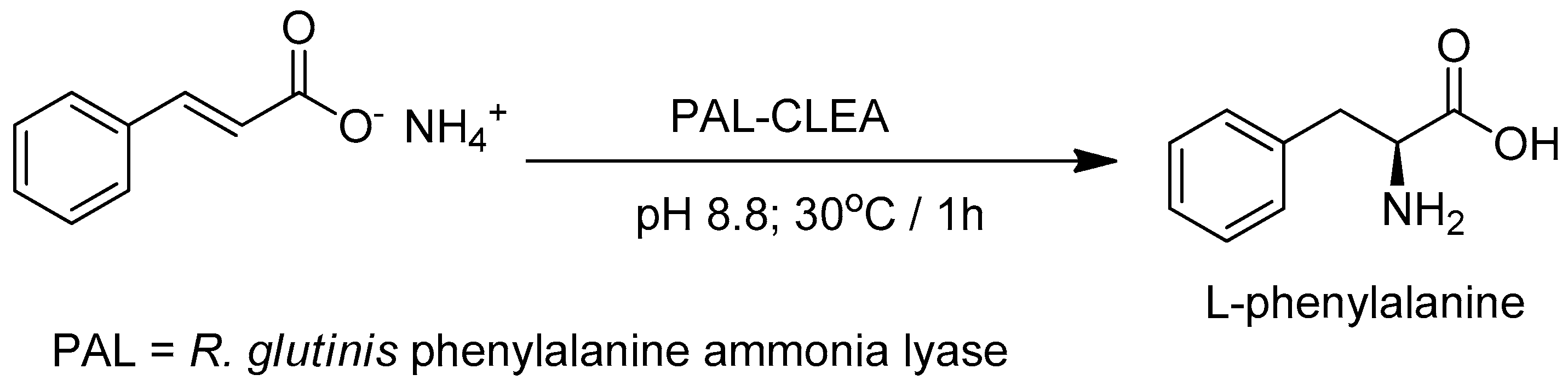
- Cui, J.D.; Zhang, S.; Sun, L.M. Cross-Linked Enzyme Aggregates of Phenylalanine Ammonia Lyase: Novel Biocatalysts for Synthesis of L-Phenylalanine. Appl. Biochem. Biotechnol. 2012, 167, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D.; Sun, L.M.; Li, L.L. A Simple Technique of Preparing Stable CLEAs of Phenylalanine Ammonia Lyase Using Co-aggregation with Starch and Bovine Serum Album. Appl. Biochem. Biotechnol. 2013, 170, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, S.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratases in synthesis. Chim. Oggi 2008, 26, 2. [Google Scholar]
- Van Pelt, S.; Quignard, S.; Kubac, D.; Sorokin, D.Y.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratase CLEAs: The immobilization and stabilization of an industrially important enzyme. Green Chem. 2008, 10, 395–400. [Google Scholar] [CrossRef]
- Van Langen, L.M.; Selassa, R.P.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Aggregates of (R)-Oxynitrilase: A Stable, Recyclable Biocatalyst for Enantioselective Hydrocyanation. Org. Lett. 2005, 7, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Betancor, L.; Fernandez-Lorente, G.; Fuentes, M.; Hidalgo, A.; Guisan, J.M.; Pessela, B.C.C.; Fernandez-Lafuente, R. Cross-Linked Aggregates of Multimeric Enzymes: A Simple and Efficient Methodology To Stabilize Their Quaternary Structure. Biomacromolecules 2004, 5, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Du, X.; Jiang, W.; Tong, Y.; Zhao, Z.; Fang, R.; Feng, J. Cross-Linked Enzyme Aggregates (CLEAs) of halohydrin dehalogenase from Agrobacterium radiobacter AD1: Preparation, Characterization and application as a biocatalyst. J. Biotechnol. 2018, 272–273, 48–55. [Google Scholar] [CrossRef]
- Schoevaart, R.; Wolbers, M.W.; Golubovic, M.; Ottens, M.; Kieboom, P.G.; van Rantwijk, F.; Van der Wielen, L.A.M.; Sheldon, R.A. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol. Bioeng. 2004, 87, 754–762. [Google Scholar] [CrossRef]
- Khanahmadi, S.; Yusof, F.; Amid, A.; Mahmod, S.S.; Mahat, M.K. Optimized preparation and characterization of CLEA-lipase from cocoa pod husk. J. Biotechnol. 2015, 202, 153–161. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Cui, J.D.; Ja, S.R. Optimization protocols and improved strategies of cross-linked enzyme aggregates technology: Current development and future challenges. Crit. Rev. Biotechnol. 2015, 35, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Valesco-Lozano, S.; Lopez-Gallego, F.; Mateoes-Diaz, J.C.; Favela-Torres, E. Cross-linked enzyme aggregates (CLEA) in enzyme improvement—A review. Biocatalysis 2015, 1, 166–177. [Google Scholar] [CrossRef]
- Talekar, S.; Shah, V.; Patil, S.; Nimbalkar, M. Porous cross linked enzyme aggregates (p-CLEAs) of Saccharomyces cerevisiae invertase. Catal. Sci. Technol. 2012, 2, 1575–1579. [Google Scholar] [CrossRef]
- Wang, M.; Jia, C.; Qi, W.; Yu, Q.; Peng, X.; Su, R.; He, Z. Porous-CLEAs of papain: Application to enzymatic hydrolysis of macromolecules. Bioresour. Technol. 2010, 102, 3541–3545. [Google Scholar] [CrossRef] [PubMed]
- Vinoth Kumar, V.; Prem Kumar, M.P.; Thiruvenkadaravi, K.V.; Baskaralingram, P.; Senthil Kumar, P.; Sivanesan, S. Preparation and characterization of porous cross linked laccase aggregates for the decolorization of triphenyl methane and reactive dyes. Bioresour. Technol. 2012, 119, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Q.; Wang, M.; Qi, W.; Su, R.; He, Z. Preparation of β-mannanase CLEAs using macromolecular cross-linkers. Catal. Sci. Technol. 2013, 3, 1937–1941. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J.; Simpson, C.; Chetty, A.; Arumugam, C.; Moolman, F. Spherezymes: A novel structured self-immobilisation enzyme technology. BMC Biotechnol. 2008, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Mbanjwa, M.B.; Land, K.J.; Windvoel, T.; Papala, P.M.; Korvink, J.G.; Visser, D.; Brady, D. Production of self-immobilised enzyme microspheres using microfluidics. Proc. Biochem. 2018, 69, 75–81. [Google Scholar] [CrossRef]
- Lopez-Serrano, P.; Cao, L.; van Rantwijk, F.; Sheldon, R.A. Cross-linked enzyme aggregates with enhanced activity: Application to lipases. Biotechnol. Lett. 2002, 24, 1379–1383. [Google Scholar] [CrossRef]
- Sorgedrager, M.J.; Verdoes, D.; van der Meer, H.; Sheldon, R.A. Cross-linked enzyme aggregates in a membrane slurry reactor. Continuous production of 6-APA by enzymatic hydrolysis of penicillin. Chim. Oggi 2008, 26, 23–25. [Google Scholar]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Utilization of cross-linked laccase aggregates in a perfusion basket reactor for the continuous elimination of endocrine-disrupting chemicals. Biotechnol. Bioeng. 2009, 102, 1582–1592. [Google Scholar] [CrossRef]
- Hilterhaus, L.; Thum, O.; Liese, A. Reactor Concept for Lipase-Catalyzed Solvent-Free Conversion of Highly Viscous Reactants Forming Two-Phase Systems. Org. Process Res. Dev. 2008, 12, 618–625. [Google Scholar] [CrossRef]
- Hickey, A.M.; Marle, L.; McCreedy, T.; Watts, P.; Greenway, G.M.; Littlechild, J.A. Immobilization of thermophilic enzymes in miniaturized flow reactors. Biochem. Soc. Trans. 2007, 35, 1621–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.-Q.; Wang, S.S.; Li, L.-N.; Gao, J.; Zhang, Y.-W. Combined Cross-linked Enzyme Aggregates as Biocatalysts. Catalysts 2018, 8, 460. [Google Scholar] [CrossRef]
- Dalal, S.; Kapoor, M.; Gupta, M.N. Preparation and characterization of combi-CLEAs catalyzing multiple non-cascade reactions. J. Mol. Catal. B Enzym. 2007, 44, 128–132. [Google Scholar] [CrossRef]
- Dalal, S.; Sharma, A.; Gupta, M.N. A multipurpose immobilized biocatalyst with pectinase, xylanase and cellulase activities. Chem. Cent. J. 2007, 1, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Magro, L.; Hertz, P.F.; Fernandez-Lafuenta, R.; Klein, M.P.; Rodrigues, R.C. Preparation and characterization of a Combi-CLEA from pectinases and cellulases: Potential biocatalyst for grape juice clarification. RSC Adv. 2016, 6, 27242–27251. [Google Scholar] [CrossRef]
- Touahar, I.E.; Haroune, L.; Ba, S.; Belenger, J.-P.; Cabana, H. Characterization of combined cross-linked enzyme aggregates from laccase, versatile peroxidase and glucose oxidase and their utilization for the elimination of pharmaceuticals. Sci. Total Environ. 2014, 481, 90–99. [Google Scholar] [CrossRef]
- Ba, S.; Mialon, A.; Jones, J.P.; Cabana, H. Laccase insolubilized as combination of crosslinked enzyme aggregates: Characterization and potential application in wastewater treatment. J. Enzym. Res. 2012, 3, 23–28. [Google Scholar]
- Vafiadi, C.; Topakas, E.; Christakopoulos, P. Preparation of multipurpose cross-linked enzyme aggregates and their application to production of alkyl ferulates. J. Mol. Catal. B Enzym. 2008, 54, 35–41. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, M.S.; Khanahmadi, S.; Shah, H. Development of an immobilized biocatalyst with lipase and protease activities as a multipurpose cross-linked enzyme aggregate. Process. Biochem. 2015, 50, 2144–2157. [Google Scholar] [CrossRef]
- Muschiol, J.; Peters, C.; Oberleitner, N.; Mihovilovic, M.D.; Bornscheuer, U.T.; Rudroff, F. Cascade catalysis--strategies and challenges en route to preparative synthetic biology. Chem. Commun. 2015, 51, 5798–5811. [Google Scholar] [CrossRef]
- Schrittweiser, J.H.; Sattler, J.; Resch, V.; Mutti, F.G.; Kroutil, W. Recent biocatalytic oxidation-reduction cascades. Curr. Opin. Chem. Biol. 2011, 15, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Woodley, J.M. Process technology for multi-enzymatic reaction systems. Bioresour. Technol. 2012, 115, 183–195. [Google Scholar] [CrossRef]
- Santacoloma, P.A.D.; Sin, G.; Gernacy, K.V.; Woodley, J.M. Multienzyme-Catalyzed Processes: Next-Generation Biocatalysis. Org. Proc. Res. Dev. 2011, 15, 203–212. [Google Scholar] [CrossRef]
- Land, H.; Hendit-Forssell, P.; Martinelle, M.; Berglund, P. One-pot biocatalytic amine transaminase/acyl transferase cascade for aqueous formation of amides from aldehydes or ketones. Catal. Sci. Technol. 2016, 6, 2897–2900. [Google Scholar] [CrossRef] [Green Version]
- Quin, M.B.; Wallin, K.K.; Zhang, G.; Schmidt-Dannert, C. Spatial organization of multi-enzyme biocatalytic cascades. Org. Biomol. Chem. 2017, 15, 4260–4271. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Yang, K.-L. Combined cross-linked enzyme aggregates of horseradish peroxidase and glucose oxidase for catalyzing cascade chemical reactions. Enzym. Microb. Technol. 2017, 100, 52–59. [Google Scholar] [CrossRef]
- Taboada-Puig, R.; Junghanns, C.; Demarche, P.; Moreira, M.T.; Feijoo, G.; Lema, J.M.; Agathos, S.N. Combined cross-linked enzyme aggregates from versatile peroxidase and glucose oxidase: Production, partial characterization and application for the elimination of endocrine disruptors. Bioresour. Technol. 2011, 102, 6593–6599. [Google Scholar] [CrossRef]
- Ning, C.; Su, E.; Tian, Y.; Wei, D. Combined cross-linked enzyme aggregates (combi-CLEAs) for efficient integration of a ketoreductase and a cofactor regeneration system. J. Biotechnol. 2014, 184, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, L.; Chen, D.; Wang, Y.; Zhang, J.; Shao, L. Co-expression of the recombined alcohol dehydrogenase and glucose dehydrogenase and cross-linked enzyme aggregates stabilization. Bioresour. Technol. 2017, 224, 531–535. [Google Scholar] [CrossRef]
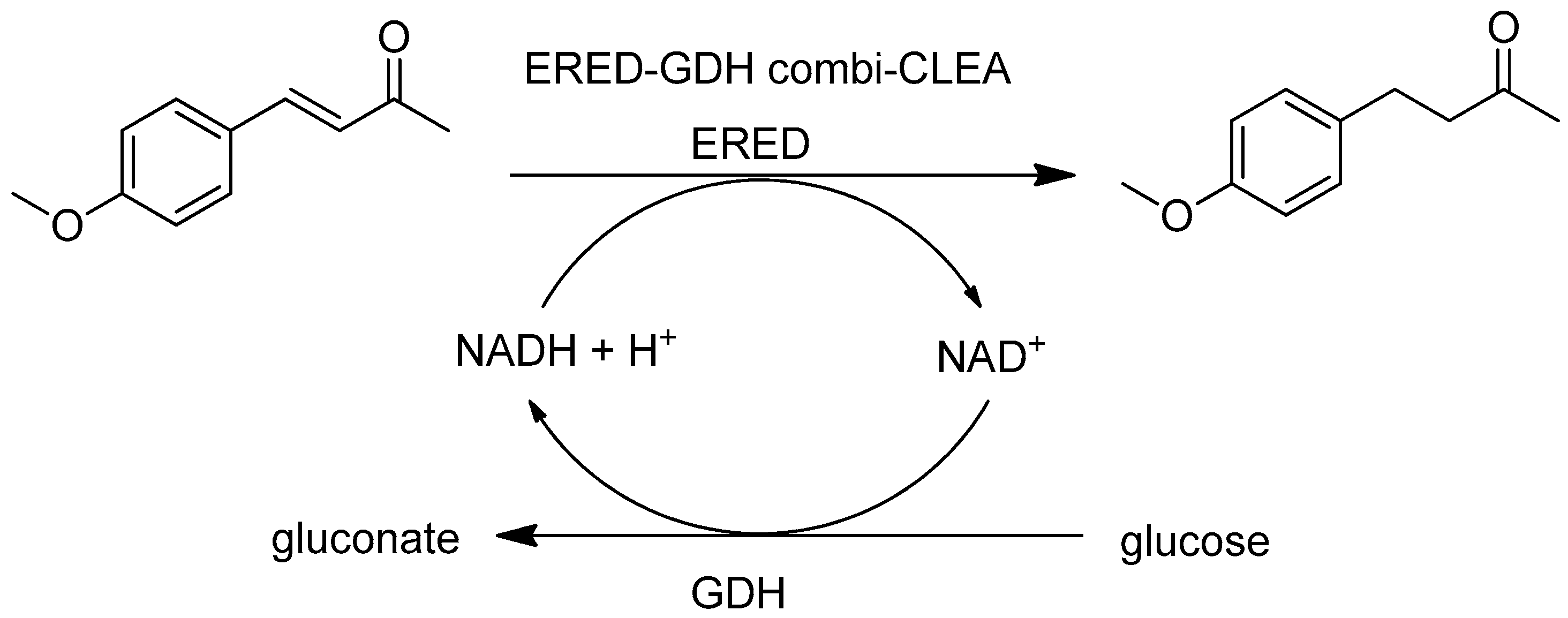
- Li, H.; Xiao, W.; Xie, P.; Zheng, L. Co-immobilization of enoate reductase with a cofactor-recycling partner enzyme. Enzym. Microb. Technol. 2018, 109, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Chmura, A.; Rustler, S.; Paravidino, M.; van Rantwijk, F.; Stolz, A.; Sheldon, R.A. The combi-CLEA approach: Enzymatic cascade synthesis of enantiomerically pure (S)-mandelic acid. Tetrahedron Asymmetry 2013, 24, 1225–1232. [Google Scholar] [CrossRef]
- Van Rantwijk, F.; Stolz, A. Enzymatic cascade synthesis of (S)-2-hydroxycarboxylic amides and acids: Cascade reactions employing a hydroxynitrile lyase, nitrile-converting enzymes and an amidase. J. Mol. Catal. B Enzym. 2015, 114, 25–30. [Google Scholar] [CrossRef]
- Van Pelt, S.; van Rantwijk, F.; Sheldon, R.A. Synthesis of Aliphatic (S)-α-Hydroxycarboxylic Amides using a One-Pot Bienzymatic Cascade of Immobilised Oxynitrilase and Nitrile Hydratase. Adv. Synth. Catal. 2009, 351, 397–404. [Google Scholar] [CrossRef]
- Stressler, T.; Ewert, J.; Eisele, T.; Fischer, L. Cross-linked enzyme aggregates (CLEAs) of PepX and PepN-production, partial characterization and application of combi-CLEAs for milk protein hydrolysis. Biocatal. Agric. Biotechnol. 2015, 50, 752–760. [Google Scholar] [CrossRef]
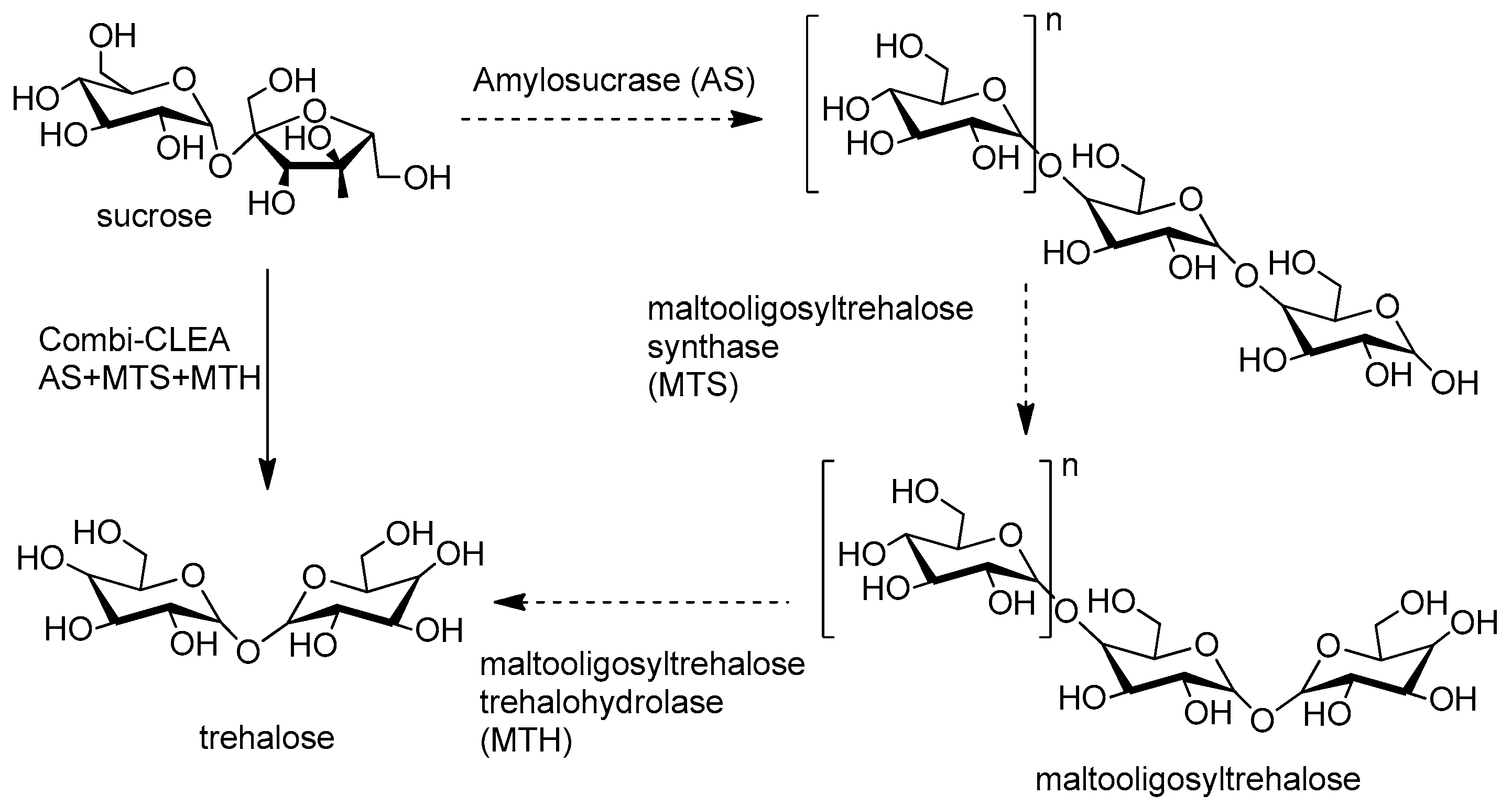
- Jung, D.-H.; Jung, J.-H.; Seo, D.-H.; Ha, S.-J.; Kweon, D.-K.; Park, C.-S. One-pot conversion of sucrose to trehalose using enzymatic sequential reactions in combined cross-linked enzyme aggregates. Bioresour. Technol. 2013, 130, 801–804. [Google Scholar] [CrossRef] [PubMed]
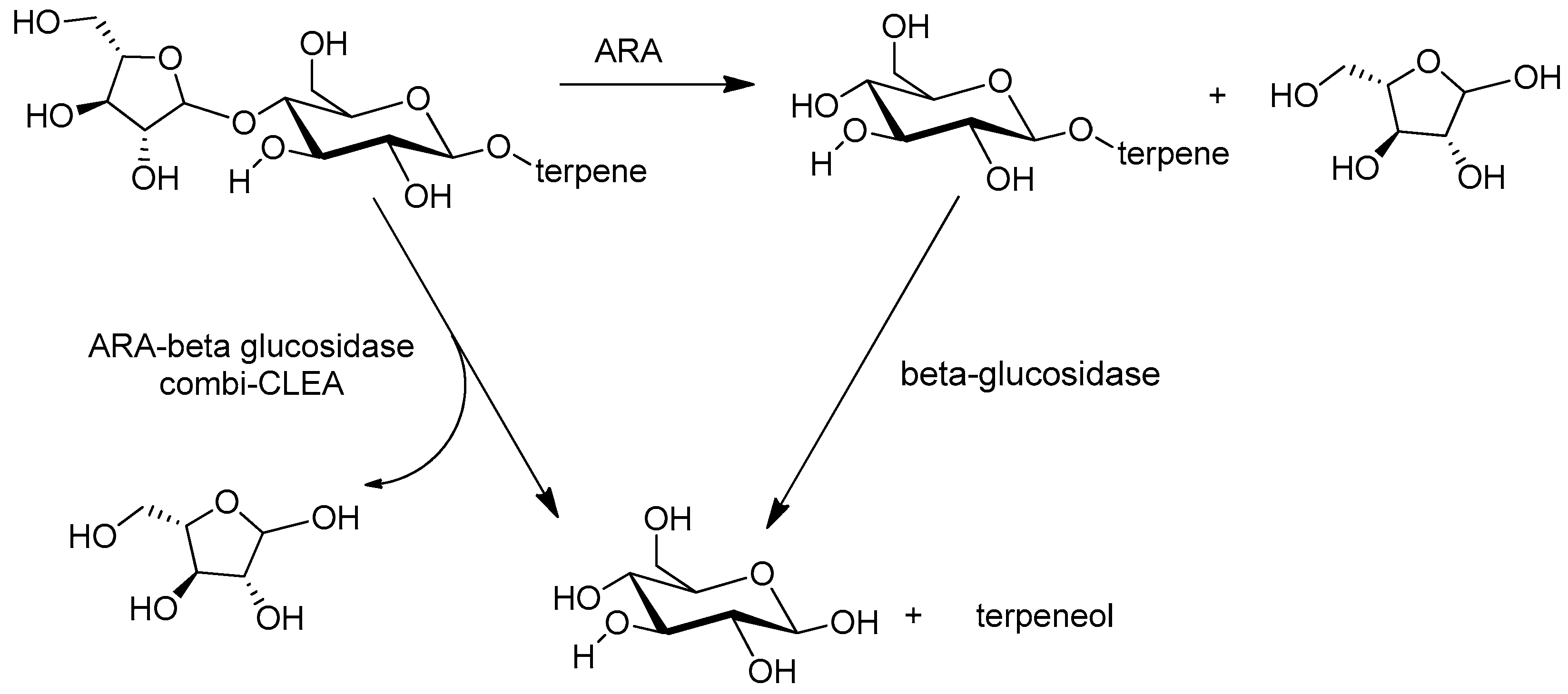
- Ahumada, K.; Urrutia, P.; Illanes, A.; Wilson, L. Production of combi-CLEAs of glycosidases utilized for aroma enhancement in wine. Food Bioprod. Process. 2015, 94, 555–560. [Google Scholar] [CrossRef]
- Talekar, S.; Waingade, S.; Gaikwad, V.; Patil, S.; Nagavekar, N. Preparation and characterization of crosslinked enzyme aggregates (CLEAs) of Bacillus amyloliquefaciens alpha amylase. J. Biochem. Technol. 2012, 3, 349–353. [Google Scholar]
- Sahutoglu, A.S.; Akgul, C. Immobilisation of Aspergillus oryzae α-amylase and Aspergillus niger glucoamylase enzymes as cross-linked enzyme aggregates. Chem. Pap. 2015, 69, 433–439. [Google Scholar] [CrossRef]
- Nadar, S.S.; Muley, A.B.; Ladole, M.R.; Joshi, P.U. Macromolecular cross-linked enzyme aggregates (m-CLEAs) of α-amylase. Int. J. Biol. Macromol. 2016, 84, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-D.; Wu, J.; Jia, D.-C.; Wan, Y.H.; Yang, N.; Qiao, M. Preparation of Cross-Linked Glucoamylase Aggregates Immobilization by Using Dextrin and Xanthan Gum as Protecting Agents. Catalysts 2016, 6, 77. [Google Scholar] [CrossRef]
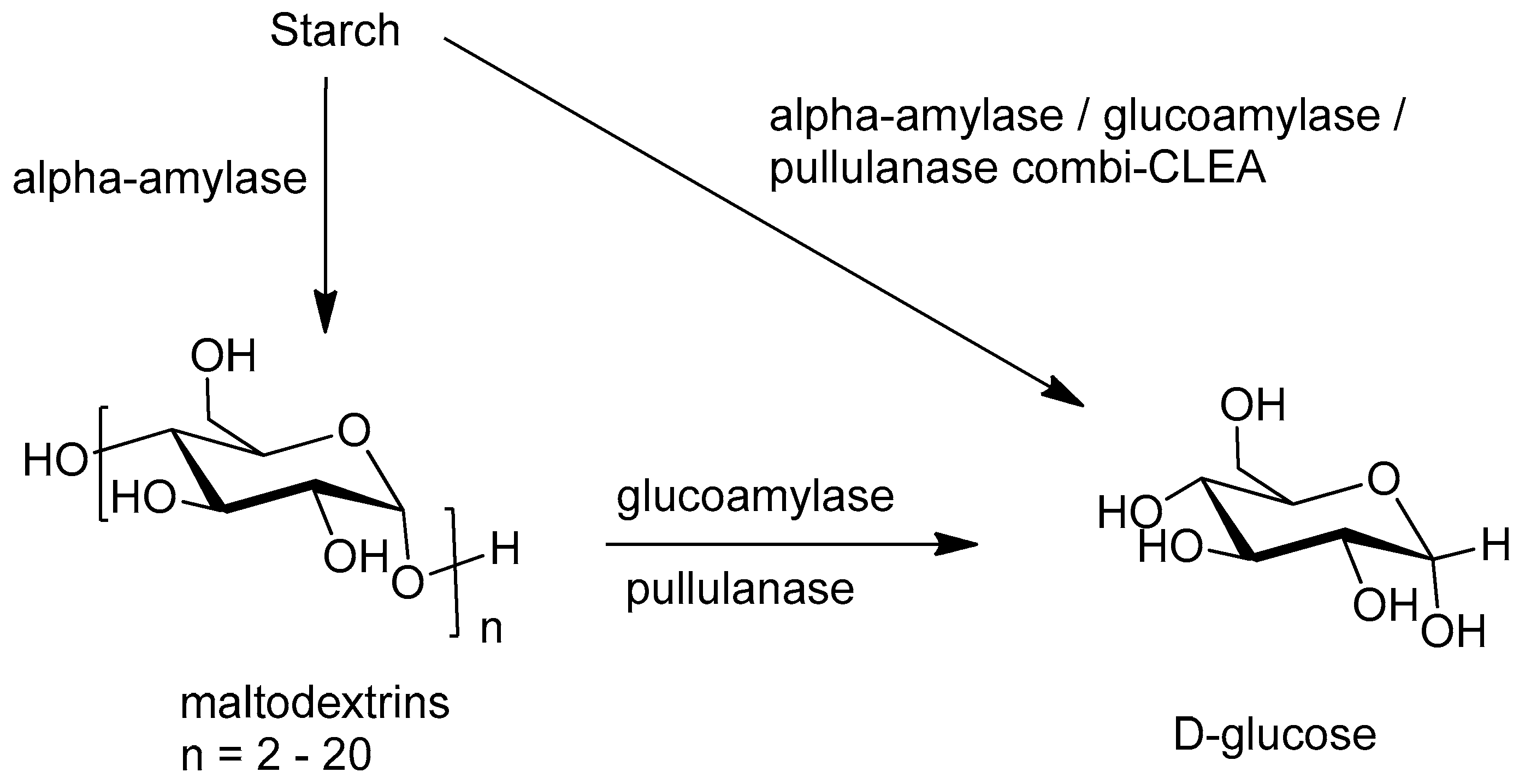
- Talekar, S.; Desai, S.; Pillai, M.; Nagavekar, N.; Ambarkar, S.; Surnis, S.; Ladole, M.; Nadar, S.; Mulla, N. Carrier free co-immobilization of glucoamylase and pullulanase as combi-cross linked enzyme aggregates (combi-CLEAs). RSC Adv. 2013, 3, 2265–2271. [Google Scholar] [CrossRef]
- Talekar, S.; Pandharbale, A.; Ladole, M.; Nadar, S.; Mulla, M.; Japhalekar, K.; Pattankude, K.; Arage, D. Carrier free co-immobilization of alpha amylase, glucoamylase and pullulanase as combined cross-linked enzyme aggregates (combi-CLEAs). Bioresour. Technol. 2013, 147, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Bhattachariya, A.S.; Bhattachariya, A.; Pletschke, B.I. Synergism of fungal and bacterial cellulases and hemicellulases: A novel perspective for enhanced bio-ethanol production. Biotechnol. Lett. 2015, 37, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Bayer, E.A.; Belaich, J.-P.; Shoham, Y.; Lamed, R. The Cellulosomes: Multienzyme Machines for Degradation of Plant Cell Wall Polysaccharides. Ann. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Bhattachariya, A.; Pletschke, B.I. Strategic optimization of xylanase-mannanase combi-CLEAs for synergistic and efficient hydrolysis of complex lignocellulosic substrates. J. Mol. Catal. B Enzym. 2015, 115, 140–150. [Google Scholar] [CrossRef]
- Periyasamy, K.; Santhalembi, L.; Mortha, G.; Aurousseau, M.; Subramanian, S. Carrier-free co-immobilization of xylanase, cellulase and β-1,3-glucanase as combined cross-linked enzyme aggregates (combi-CLEAs) for one-pot saccharification of sugarcane bagasse. RSC Adv. 2016, 6, 32849–32857. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Sorgedrager, M.J.; Kondor, B. Non-Leachable Magnetic Cross-Linked Enzyme Aggregate. WO Patent 2012/023847 A2, 23 February 2012. [Google Scholar]
- Whitesides, G.M.; Hill, C.L.; Brunie, J.-C. Magnetic Filtration of Small Heterogeneous Catalyst Particles. Preparation of Ferrimagnetic Catalyst Supports. Ind. Eng. Chem. Process Des. Dev. 1976, 15, 226–227. [Google Scholar] [CrossRef]
- Cui, J.D.; Cui, L.L.; Zhang, S.P.; Zhang, Y.F.; Su, Z.G.; Ma, G.H. Hybrid Magnetic Cross-Linked Enzyme Aggregates of Phenylalanine Ammonia Lyase from Rhodotorula glutinis. PLoS ONE 2014, 9, e97221. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, S.; Janssen, M.H.A.; Rasmussen, J.-A.M.; Sorgedrager, M.J.; Sheldon, R.A.; Koning, P. Cross-Linked Enzyme Aggregates Comprising Magnetizable Particles. WO Patent 2018/004341 A1, 4 January 2018. [Google Scholar]
- Talekar, S.; Ghodake, V.; Ghotage, T.; Rathod, P.; Deshmukh, P.; Nadar, S.; Mulla, M.; Ladole, M. Novel magnetic cross-linked enzyme aggregates (magnetic CLEAs) of alpha amylase. Bioresour. Technol. 2012, 123, 542–547. [Google Scholar] [CrossRef]
- Nadar, S.S.; Rathod, V.K. Magnetic macromolecular cross linked enzyme aggregates (CLEAs) of glucoamylase. Enzym. Microb. Technol. 2016, 83, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Kumar Jana, A.; Kumar, S.; Maiti, M. Immobilization of amyloglucosidase from SSF of Aspergillus niger by crosslinked enzyme aggregate onto magnetic nanoparticles using minimum amount of carrier and characterizations. J. Mol. Catal. B Enzym. 2013, 98, 30–36. [Google Scholar] [CrossRef]
- Amaral-Fonseca, M.; Kopp, W.; de Lima Camargo Giordano, R.; Fernandez-Lafuenta, R.; Tardioli, P.W. Preparation of Magnetic Cross-Linked Amyloglucosidase Aggregates: Solving Some Activity Problems. Catalysts 2018, 8, 496. [Google Scholar] [CrossRef]
- Asgher, M.; Shahid, M.; Kamal, S.; Iqbal, H.M.N. Recent trends and valorization of immobilization strategies and ligninolytic enzymes by industrial biotechnology. J. Mol. Catal. B Enzym. 2014, 101, 56–66. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Pletschke, B.I. Magnetic cross-linked enzyme aggregates (CLEAs): A novel concept towards carrier free immobilization of lignocellulolytic enzymes. Enzym. Microb. Technol. 2014, 61–62, 17–27. [Google Scholar] [CrossRef]
- Khorshadi, K.J.; Lenjannezhadian, H.; Jamalan, M.; Zeinali, M. Preparation and characterization of nanomagnetic cross-linked cellulase aggregates for cellulose bioconversion. J. Chem. Technol. Biotechnol. 2016, 91, 539–546. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, W.; Yang, Z.; Yang, X.; Wang, N.; Yu, X. Novel Magnetic Cross-Linked Cellulase Aggregates with a Potential Application in Lignocellulosic Biomass Conversion. Molecules 2017, 22, 269. [Google Scholar] [CrossRef]
- Hwangbo, M.; Tran, J.L.; Chu, K.-H. Effective one-step saccharification of lignocellulosic biomass using magnetite-biocatalysts containing saccharifying enzymes. Sci. Total Environ. 2019, 647, 806–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaarani, S.M.; Jahim, J.M.; Rahman, R.A.; Idris, A.; Murad, A.M.A.; Illias, R.M. Silanized maghemite for cross-linked enzyme aggregates of recombinant xylanase from Trichoderma reesei. J. Mol. Catal. B Enzym. 2016, 133, 65–76. [Google Scholar] [CrossRef]
- Olofsson, K.; Bertilsson, M.; Liden, G. A short review on SSF—An interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol. Biofuels 2008, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Sojitra, U.V.; Nadar, S.S.; Rathod, V.K. A magnetic tri-enzyme nanobiocatalyst for fruit juice clarification. Food Chem. 2016, 213, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Pfaltzgraff, L.A.; De Bruyn, M.; Cooper, E.C.; Budarin, V.; Clark, J.H. Food waste biomass: A resource for high-value chemicals. Green Chem. 2013, 15, 307–314. [Google Scholar] [CrossRef]
- Mehde, A.A.; Mehdi, W.A.; Severgün, O.; Çakar, S.; Özacar, M. Lipase-based on starch material as a development matrix with magnetite cross-linked enzyme aggregates and its application. Int. J. Biol. Macromol. 2018, 120, 1533–1543. [Google Scholar] [CrossRef]
- Cruz-Izquierdo, A.; Pico, E.A.; Lopez, C.; Serra, J.L.; Lama, M.J. Magnetic Cross-Linked Enzyme Aggregates (mCLEAs) of Candida antarctica Lipase: An Efficient and Stable Biocatalyst for Biodiesel Synthesis. PLoS ONE 2014, 9, e115202. [Google Scholar] [CrossRef]
- Picó, E.A.; López, C.; Cruz-Izquierdo, A.; Munarriz, M.; Iruretagoyena, F.J.; Serra, J.L.; Llama, M.J. Easy reuse of magnetic cross-linked enzyme aggregates of lipase B from Candida antarctica to obtain biodiesel from Chlorella vulgaris lipids. J. Biosci. Bioeng. 2018, 126, 451–457. [Google Scholar] [CrossRef]
- Zhang, W.W.; Yang, X.L.; Jia, J.Q.; Wang, N.; Hu, C.J.; Yu, X.Q. Surfactant-activated magnetic cross-linked enzyme aggregates (magnetic CLEAs) of Thermomyces lanuginosus lipase for biodiesel production. J. Mol. Catal. B Enzym. 2015, 115, 83–89. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, H.; Liu, W.; Wang, N.; Yu, X. Improved Performance of Magnetic Cross-Linked Lipase Aggregates by Interfacial Activation: A Robust and Magnetically Recyclable Biocatalyst for Transesterification of Jatropha Oil. Molecules 2017, 22, 2157. [Google Scholar] [CrossRef]
- Cui, J.; Cui, L.; Jia, S.; Su, Z.; Zhang, S. Hybrid Cross-Linked Lipase Aggregates with Magnetic Nanoparticles: A Robust and Recyclable Biocatalyst for the Epoxidation of Oleic Acid. J. Agric. Food Chem. 2016, 64, 7179–7187. [Google Scholar] [CrossRef]
- Tudorache, M.; Gheorghe, A.; Viana, A.S.; Parvulescu, V.I. Biocatalytic epoxidation of α-pinene to oxy-derivatives over cross-linked lipase aggregates. J. Mol. Catal. B Enzym. 2016, 134, 9–15. [Google Scholar] [CrossRef]
- Tudorache, M.; Nae, A.; Coman, S.; Parvulescu, V.I. Strategy of cross-linked enzyme aggregates onto magnetic particles adapted to the green design of biocatalytic synthesis of glycerol carbonate. RSC Adv. 2013, 3, 4052–4058. [Google Scholar] [CrossRef]
- Zheng, D.; Wang, S.; Qiu, S.; Lin, J.; Diao, X. Synthesis of butyl oleate catalyzed by cross-linked enzyme aggregates with magnetic nanoparticles in rotating magneto-micro-reactor. J. Biotechnol. 2018, 281, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Loos, K. Enzymatic Synthesis of Biobased Polyesters and Polyamides. Polymers 2016, 8, 243. [Google Scholar] [CrossRef]
- Stavila, E.; Arsyi, R.Z.; Petrovic, D.M.; Loos, K. Fusarium solani pisi cutinase-catalyzed synthesis of polyamides. Eur. Polym. J. 2013, 49, 834–842. [Google Scholar] [CrossRef]
- Steunenberg, P.; Uiterweerd, M.; Sijm, M.; Scott, E.L.; Zuilhof, H.; Sanders, J.P.M.; Franssen, M.C.R. Enzyme-Catalyzed Polymerization of β-alanine Esters, A Sustainable Route Towards the Formation of Poly-β-alanine. Curr. Org. Chem. 2013, 17, 682–690. [Google Scholar] [CrossRef]
- Kumar, V.V.; Sivaesan, S.; Cabana, H. Magnetic cross-linked laccase aggregates—Bioremediation tool for decolorization of distinct classes of recalcitrant dyes. Sci. Total Environ. 2014, 487, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, Z.; Lin, Y.; Ng, T.B.; Ye, X.; Lin, J. Immobilized Cerrena sp. Laccase: Preparation, thermal inactivation and operational stability in malachite green decolorization. Sci. Rep. 2017, 7, 16429. [Google Scholar] [CrossRef]
- Zhou, L.; Tang, W.; Jiang, Y.; Ma, L.; He, Y.; Gao, J. Magnetic combined cross-linked enzyme aggregates of horseradish peroxidase and glucose oxidase: An efficient biocatalyst for dye decolourization. RSC Adv. 2016, 6, 90061–90068. [Google Scholar] [CrossRef]
- Kumar, V.V.; Cabana, H. Towards high potential magnetic biocatalysts for on-demand elimination of pharmaceuticals. Bioresour. Technol. 2016, 200, 81–89. [Google Scholar] [CrossRef]
- Bedade, D.K.; Muley, A.B.; Singhal, R.S. Magnetic cross-linked aggregates of acrylamidase from Cupriavidus oxalaticus ICTDB921 for biodegradation of acrylamide from industrial waste water. Bioresour. Technol. 2019, 272, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Grooten, H.M.J.; Rijkers, M.P.W.M.; Moody, H.M.; Webbers, J.J.P.; De Vroom, E. Method for the Separation of a Solid Substance. WO Patent 98/56486, 17 December 1998. [Google Scholar]
- Kopp, W.; Da Costa, T.P.; Pereira, S.C.; Jafelicci, M.; Giordano, R.C.; Marques, R.F.C.; Araujo-Moreira, F.M.; Giordano, R.L.C. Easily handling penicillin G acylase magnetic cross-linked enzymes aggregates: Catalytic and morphological studies. Process Biochem. 2014, 49, 38–46. [Google Scholar] [CrossRef]
- Ling, X.-M.; Wang, X.-Y.; Ma, P.; Yang, Y.; Qin, J.-M.; Zhang, X.-J.; Zhang, Y.-W. Covalent Immobilization of Penicillin G Acylase onto Fe3O4@Chitosan Magnetic Nanoparticles. J. Microbiol. Biotechnol. 2016, 26, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Illanes, A.; Soler, L.; Henriquez, M.J. Effect of the degree of cross-linking on the properties of different CLEAs of penicillin acylase. J. Process Biochem. 2009, 44, 322–326. [Google Scholar] [CrossRef]
- Su, E.; Meng, Y.; Ning, C.; Ma, X.; Deng, S. Magnetic combined cross-linked enzyme aggregates (Combi-CLEAs) for cofactor regeneration in the synthesis of chiral alcohol. J. Biotechnol. 2018, 271, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, Q.; Sun, L.; Li, Q.; Zhou, L.; Chen, Q.; Li, S.; Yu, M.; Li, W. Magnetic Combined Cross-Linked Enzyme Aggregates of Ketoreductase and Alcohol dehydrogenase: An Efficient and Stable Biocatalyst for Asymmetric Synthesis of (R)-3-Quinuclidinol with Regeneration of Coenzymes In Situ. Catalysts 2018, 8, 334. [Google Scholar] [CrossRef]
- Peirce, S.; Russo, M.E.; De Luca, V.; Capasso, C.; Rossi, M.; Olivieri, G.; Salatino, P.; Marzocchella, A. Immobilization of carbonic anhydrase for biomimetic CO2 capture in a slurry absorber as cross-linked enzyme aggregates (CLEA). Chem. Eng. Trans. 2015, 43, 259–264. [Google Scholar] [CrossRef]
- Gupta, A.; Srivastava, S.K. Study of cross linked enzyme aggregate of glucose isomerase of Streptomyces thermonitrificans immobilised on magnetic particle. J. Biochem. Technol. 2016, 7, 1102–1106. [Google Scholar]














© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheldon, R.A. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts 2019, 9, 261. https://doi.org/10.3390/catal9030261
Sheldon RA. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts. 2019; 9(3):261. https://doi.org/10.3390/catal9030261
Chicago/Turabian StyleSheldon, Roger A. 2019. "CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy" Catalysts 9, no. 3: 261. https://doi.org/10.3390/catal9030261
APA StyleSheldon, R. A. (2019). CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts, 9(3), 261. https://doi.org/10.3390/catal9030261