
Structural and Biochemical Studies of Bacillus subtilis MobB

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression, and Purification
2.2. Crystallization, Data Collection, and Structure Determination
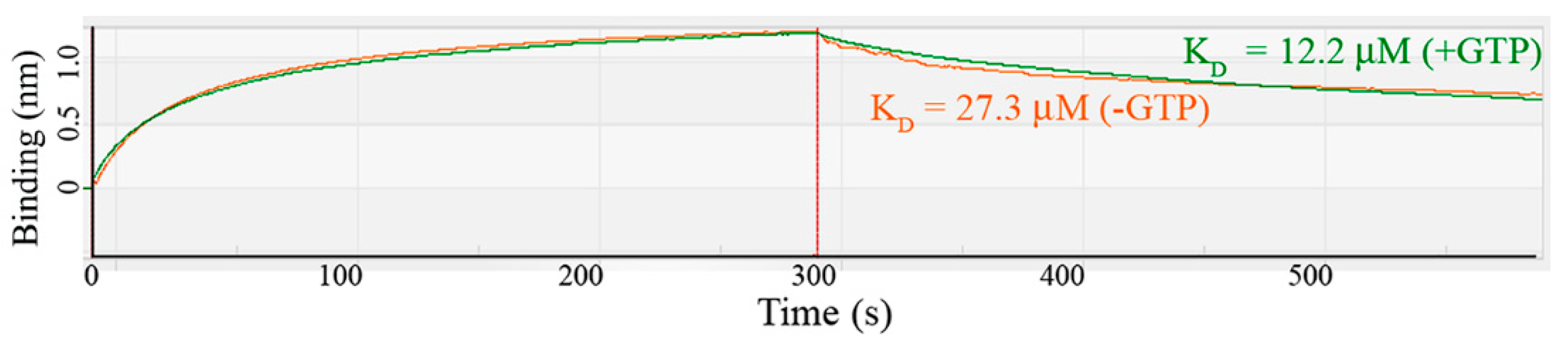
2.3. MobA Preparation and MobA:MobB Binding Assay
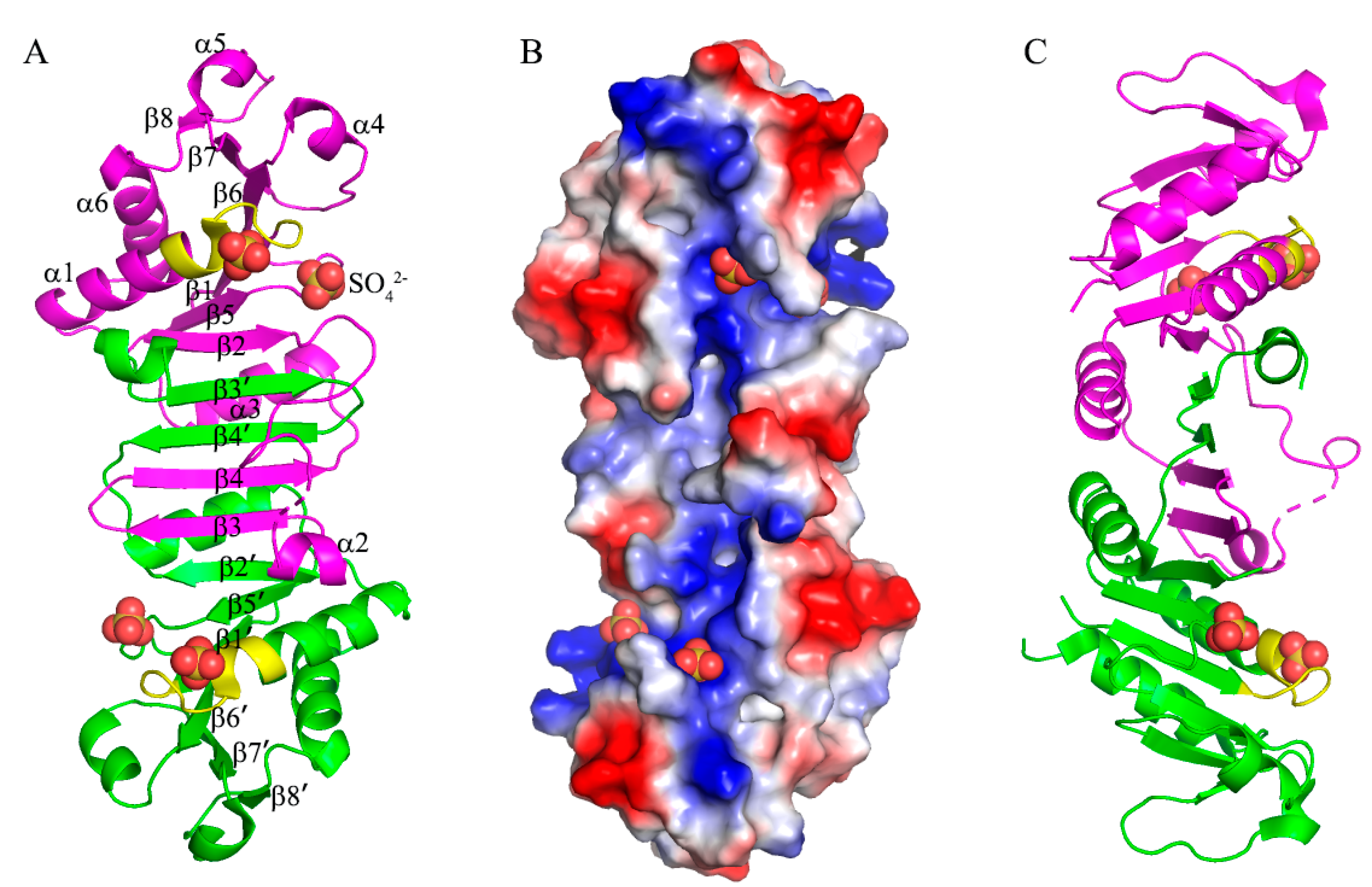
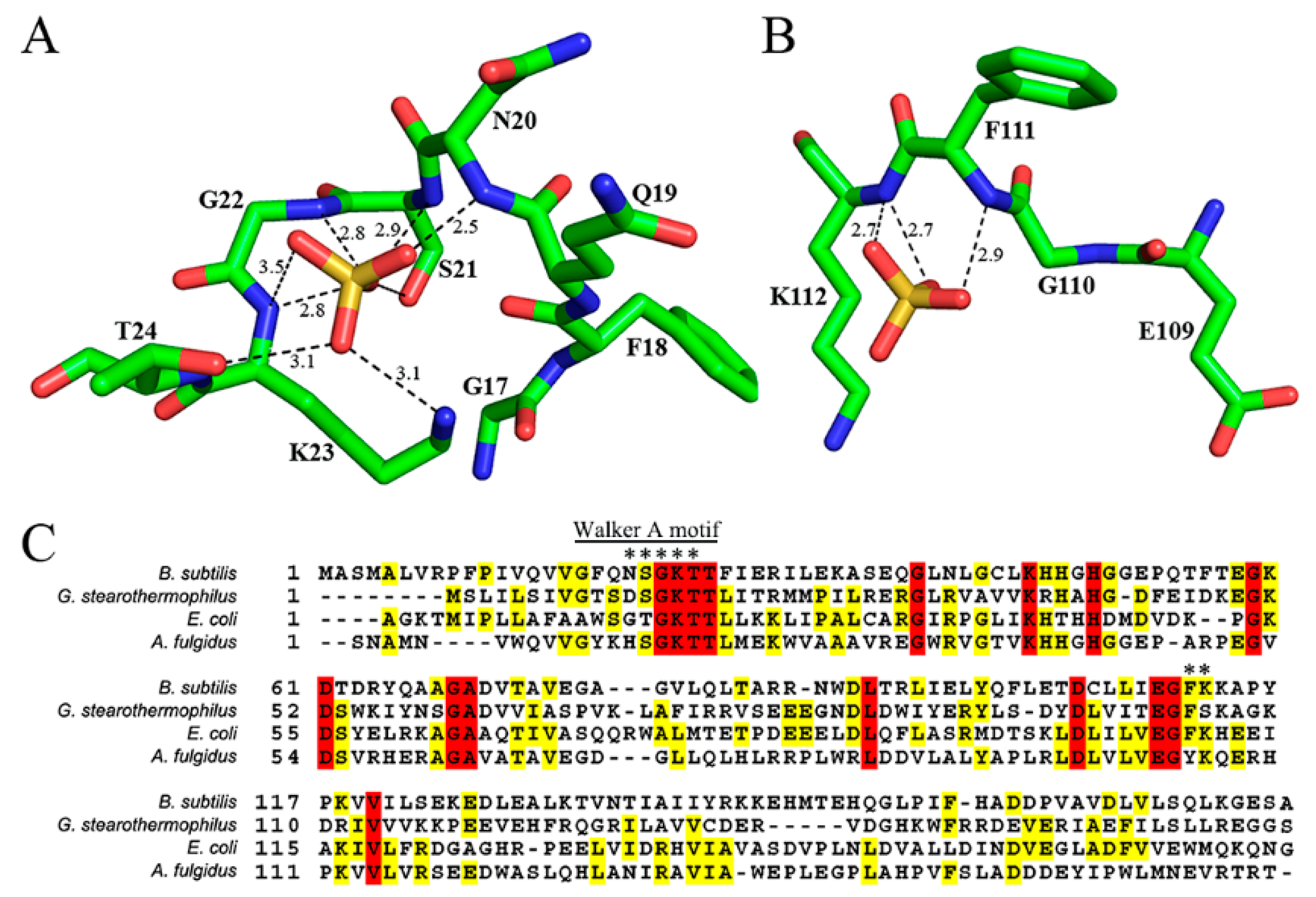
3. Results and Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hille, R. The mononuclear molybdenum enzymes. Chem. Rev. 1996, 96, 2757–2816. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, K.V.; Johnson, J.L. The pterin molybdenum cofactors. J. Biol. Chem. 1992, 267, 10199–10202. [Google Scholar] [CrossRef]
- Hover, B.M.; Loksztejn, A.; Ribeiro, A.A.; Yokoyama, K. Identification of a Cyclic Nucleotide as a Cryptic Intermediate in Molybdenum Cofactor Biosynthesis. J. Am. Chem. Soc. 2013, 135, 7019–7032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutzke, G.; Fischer, B.; Mendel, R.R.; Schwarz, G. Thiocarboxylation of molybdopterin synthase provides evidence for the mechanism of dithiolene formation in metal-binding pterins. J. Biol. Chem. 2001, 276, 36268–36274. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, M.J.; Wuebbens, M.M.; Turque, O.; Rajagopalan, K.V.; Schindelin, H. Structural studies of molybdopterin synthase provide insights into its catalytic mechanism. J. Biol. Chem. 2003, 278, 14514–14522. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.S.; Johnson, J.L.; Rajagopalan, K.V. Molybdenum cofactor biosynthesis in Escherichia coli mod and mog mutants. J. Bacteriol. 1996, 178, 4310–4312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuper, J.; Winking, J.; Hecht, H.J.; Mendel, R.R.; Schwarz, G. The active site of the molybdenum cofactor biosynthetic protein domain Cnx1G. Arch. Biochem. Biophys. 2003, 411, 36–46. [Google Scholar] [CrossRef]
- Llamas, A.; Mendel, R.R.; Schwarz, N. Synthesis of adenylated molybdopterin—An essential step for molybdenum insertion. J. Biol. Chem. 2004, 279, 55241–55246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krausze, J.; Hercher, T.W.; Zwerschke, D.; Kirk, M.L.; Blankenfeldt, W.; Mendel, R.R.; Kruse, T. The functional principle of eukaryotic molybdenum insertases. Biochem. J. 2018, 475, 1739–1753. [Google Scholar] [CrossRef]
- Probst, C.; Yang, J.; Krausze, J.; Hercher, T.W.; Richers, C.P.; Spatzal, T.; Kc, K.; Giles, L.J.; Rees, D.C.; Mendel, R.R.; et al. Mechanism of molybdate insertion into pterin-based molybdenum cofactors. Nat. Chem. 2021, 13, 758–765. [Google Scholar] [CrossRef]
- Palmer, T.; Vasishta, A.; Whitty, P.W.; Boxer, D.H. Isolation of Protein Fa, a Product of the Mob Locus Required for Molybdenum Cofactor Biosynthesis in Escherichia-Coli. Eur. J. Biochem. 1994, 222, 687–692. [Google Scholar] [CrossRef]
- Neumann, M.; Mittelstadt, G.; Seduk, F.; Iobbi-Nivol, C.; Leimkuhler, S. MocA Is a Specific Cytidylyltransferase Involved in Molybdopterin Cytosine Dinucleotide Biosynthesis in Escherichia coli. J. Biol. Chem. 2009, 284, 21891–21898. [Google Scholar] [CrossRef] [Green Version]
- Reschke, S.; Sigfridsson, K.G.; Kaufmann, P.; Leidel, N.; Horn, S.; Gast, K.; Schulzke, C.; Haumann, M.; Leimkuhler, S. Identification of a bis-molybdopterin intermediate in molybdenum cofactor biosynthesis in Escherichia coli. J. Biol. Chem. 2013, 288, 29736–29745. [Google Scholar] [CrossRef] [Green Version]
- Eaves, D.J.; Palmer, T.; Boxer, D.H. The product of the molybdenum cofactor gene mobB of Escherichia coli is a GTP-binding protein. Eur. J. Biochem. 1997, 246, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Magalon, A.; Frixon, C.; Pommier, J.; Giordano, G.; Blasco, F. In vivo interactions between gene products involved in the final stages of molybdenum cofactor biosynthesis in Escherichia coli. J Biol. Chem. 2002, 277, 48199–48204. [Google Scholar] [CrossRef] [Green Version]
- McLuskey, K.; Harrison, J.A.; Schuttelkopf, A.W.; Boxer, D.H.; Hunter, W.N. Insight into the role of Escherichia coli MobB in molybdenum cofactor biosynthesis based on the high resolution crystal structure. J. Biol. Chem. 2003, 278, 23706–23713. [Google Scholar] [CrossRef] [Green Version]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Method Enzym. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Cryst. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Cryst. D 2011, 67, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Cryst. D 1997, 53, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S. The Ccp4 Suite—Programs for Protein Crystallography. Acta Cryst. D 1994, 50, 760–763. [Google Scholar]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D Biol. Cryst. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Cryst. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Concepcion, J.; Witte, K.; Wartchow, C.; Choo, S.; Yao, D.F.; Persson, H.; Wei, J.; Li, P.; Heidecker, B.; Ma, W.L.; et al. Label-Free Detection of Biomolecular Interactions Using BioLayer Interferometry for Kinetic Characterization. Comb. Chem. High. T Scr. 2009, 12, 791–800. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly Related Sequences in the Alpha-Subunits and Beta-Subunits of Atp Synthase, Myosin, Kinases and Other Atp-Requiring Enzymes and a Common Nucleotide Binding Fold. EMBO J. 1982, 1, 945–951. [Google Scholar] [CrossRef]
- DeLano, W.L.; Lam, J.W. PyMOL: A communications tool for computational models. Abstr. Pap. Am. Chem. S 2005, 230, U1371–U1372. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | |
|---|---|
| Space Group | C2 |
| Unit Cell parameters (Å, °) | a = 225.53, b = 42.11, c = 93.62, β = 100.99 |
| Wavelength (Å) | 0.97921 |
| Temperature (K) | 100 |
| Resolution (Å) a | 50.0–2.40 (2.44–2.40) |
| No. of observed reflections | 117,931 |
| No. of unique reflections | 33,530 |
| Completeness (%) | 98.8 (99.7) |
| Rsym b | 0.109 (0.666) |
| <I/σ (I)> c | 9.0 (3.1) |
| Refinement statistics | |
| No. of Residues | 746 |
| No. of water molecules | 218 |
| No. of sulfate ions | 13 |
| Rcryst (%), Rfree (%) d | 21.0, 30.3 |
| rmsd bonds (Å) | 0.014 |
| rmsd angles (°) | 1.559 |
| Ramachandran plot | |
| Most favored region (%) | 96.8 |
| Additionally allowed region (%) | 3.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Choi, S.; Kim, H.; Choe, J. Structural and Biochemical Studies of Bacillus subtilis MobB. Crystals 2021, 11, 1262. https://doi.org/10.3390/cryst11101262
Kim D, Choi S, Kim H, Choe J. Structural and Biochemical Studies of Bacillus subtilis MobB. Crystals. 2021; 11(10):1262. https://doi.org/10.3390/cryst11101262
Chicago/Turabian StyleKim, Dajeong, Sarah Choi, Hyunjin Kim, and Jungwoo Choe. 2021. "Structural and Biochemical Studies of Bacillus subtilis MobB" Crystals 11, no. 10: 1262. https://doi.org/10.3390/cryst11101262
APA StyleKim, D., Choi, S., Kim, H., & Choe, J. (2021). Structural and Biochemical Studies of Bacillus subtilis MobB. Crystals, 11(10), 1262. https://doi.org/10.3390/cryst11101262