
The Influence of Silicateins on the Shape and Crystalline Habit of Silica Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr)

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Silicatein Activity Assay
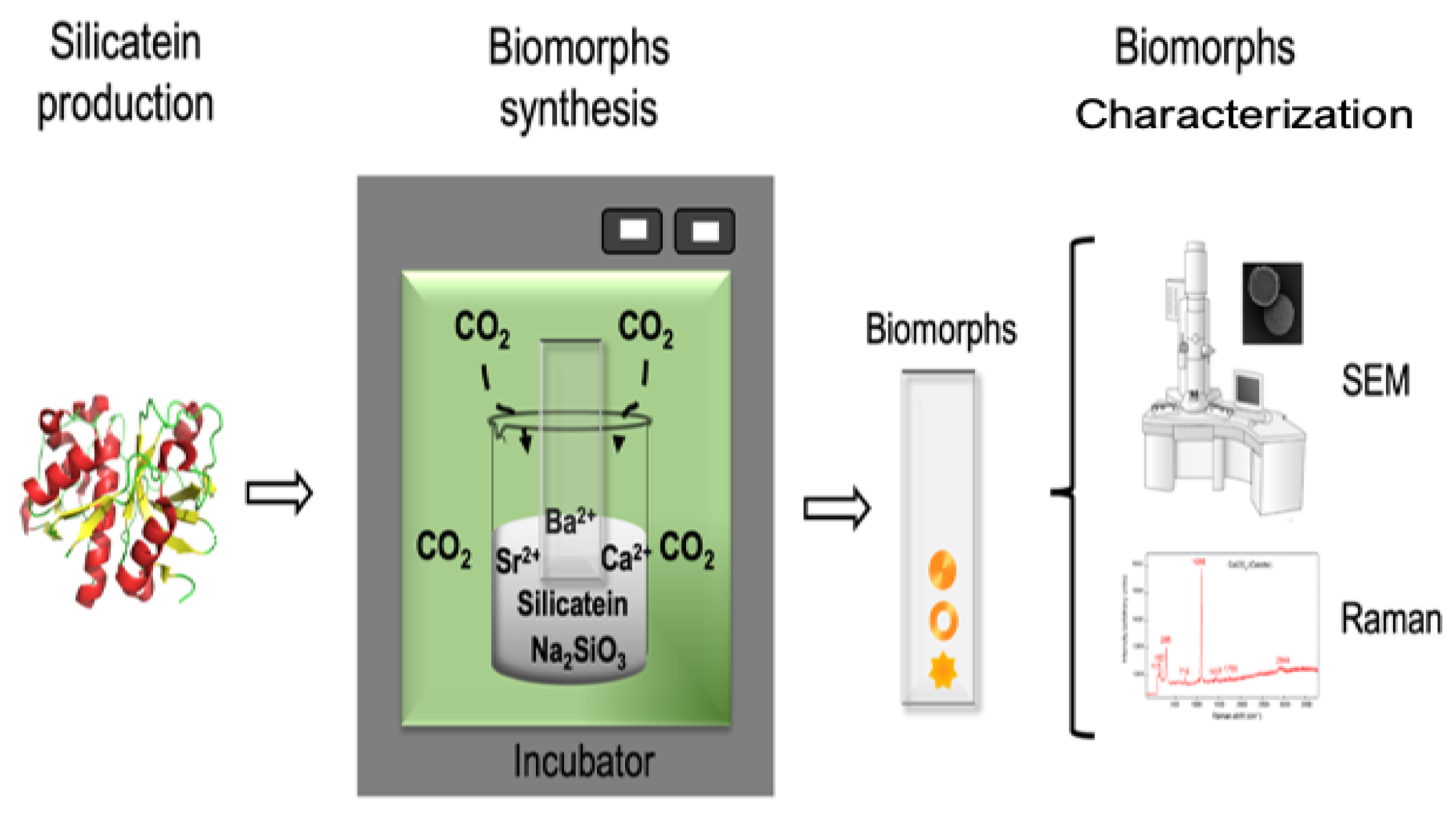
2.3. Biomorph Synthesis
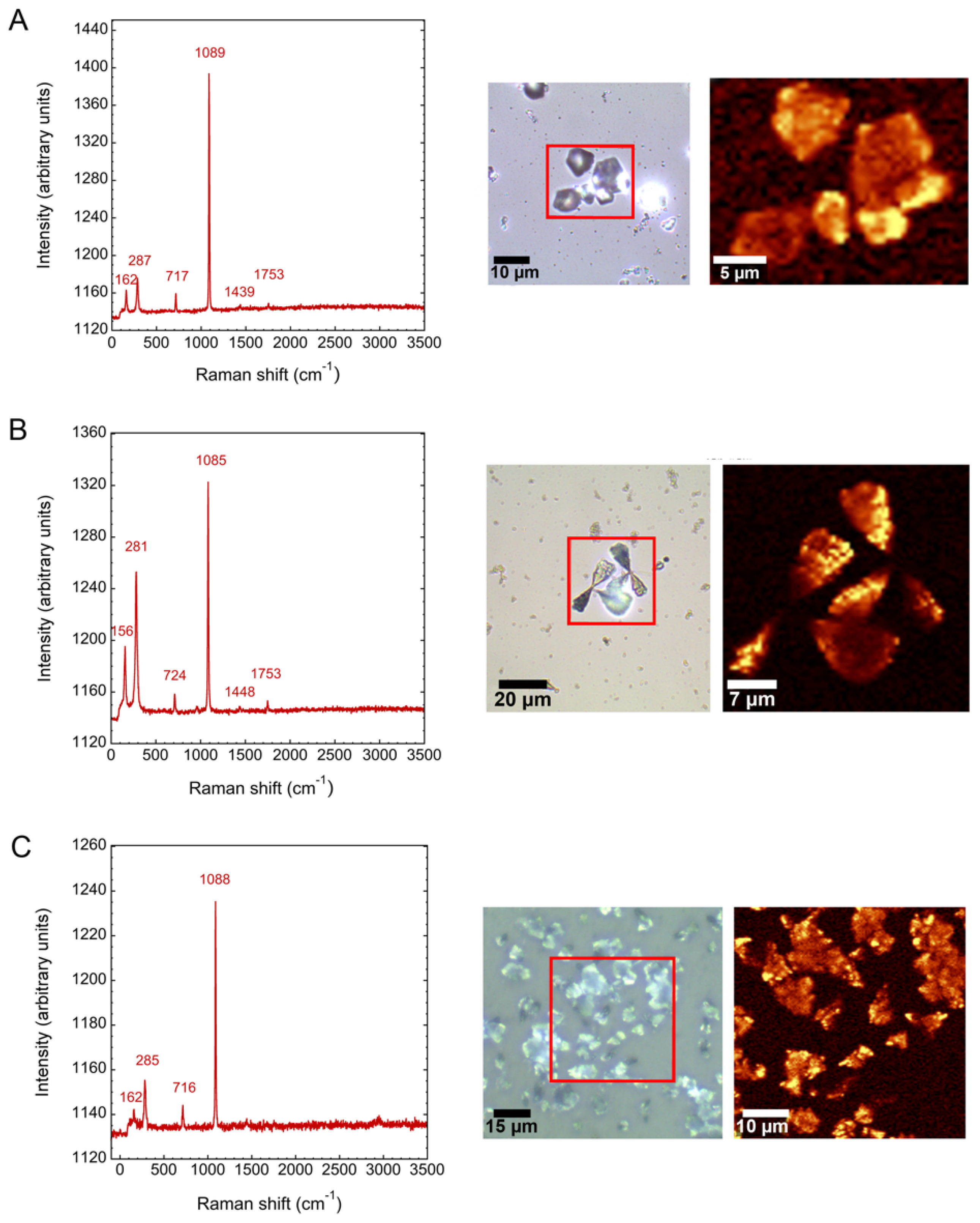
2.4. Characterization of Biomorphs
3. Results and Discussion
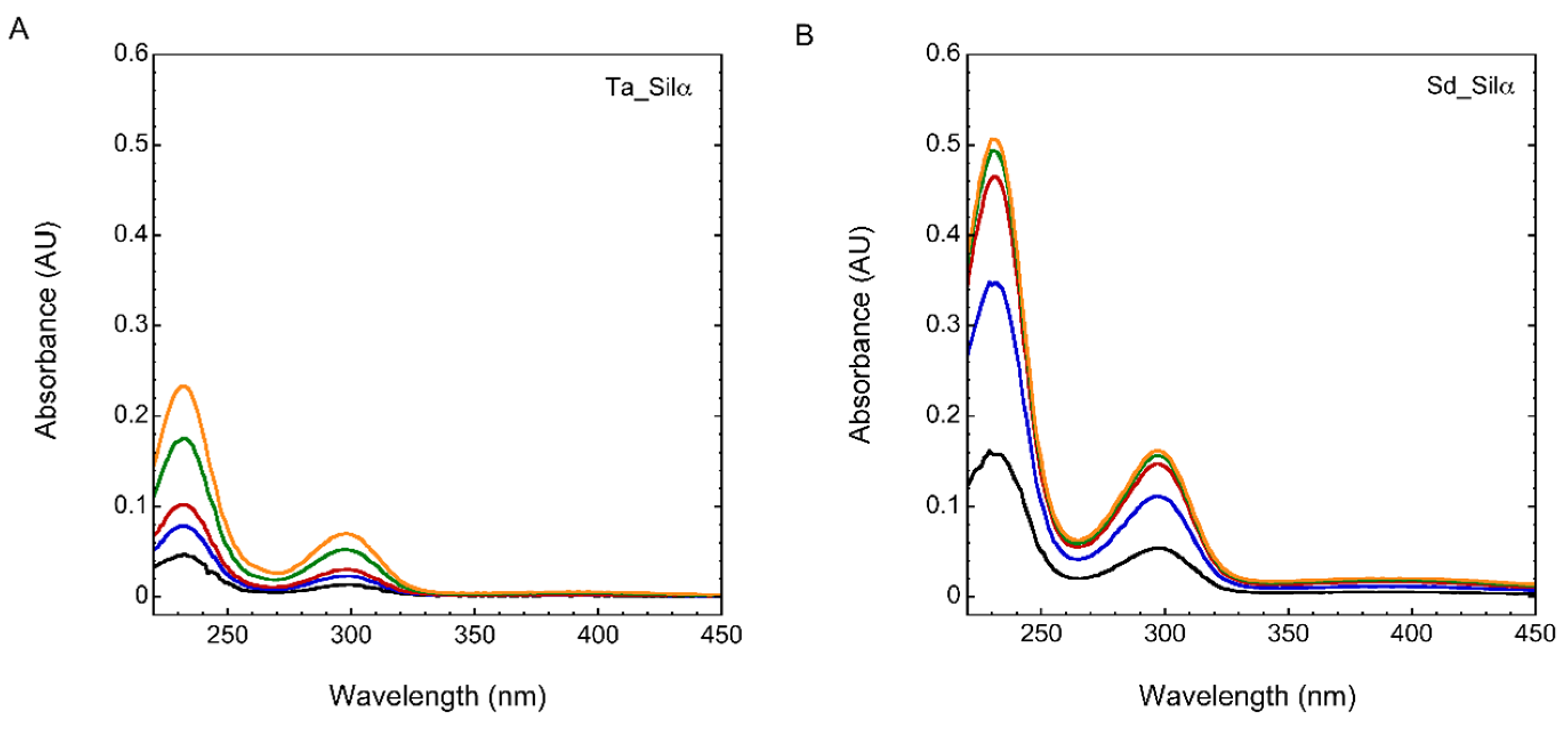
3.1. Silicatein-α Production
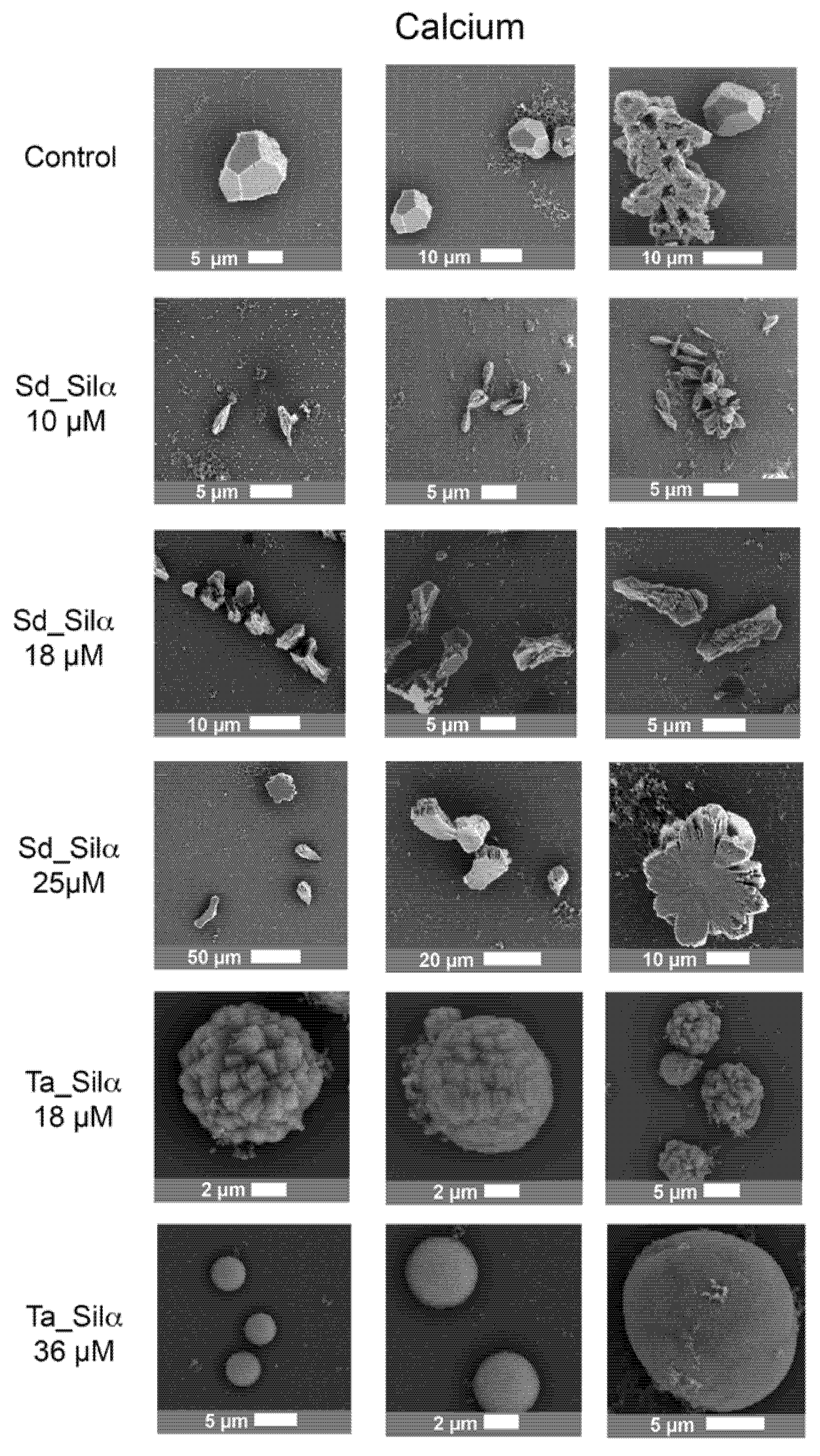
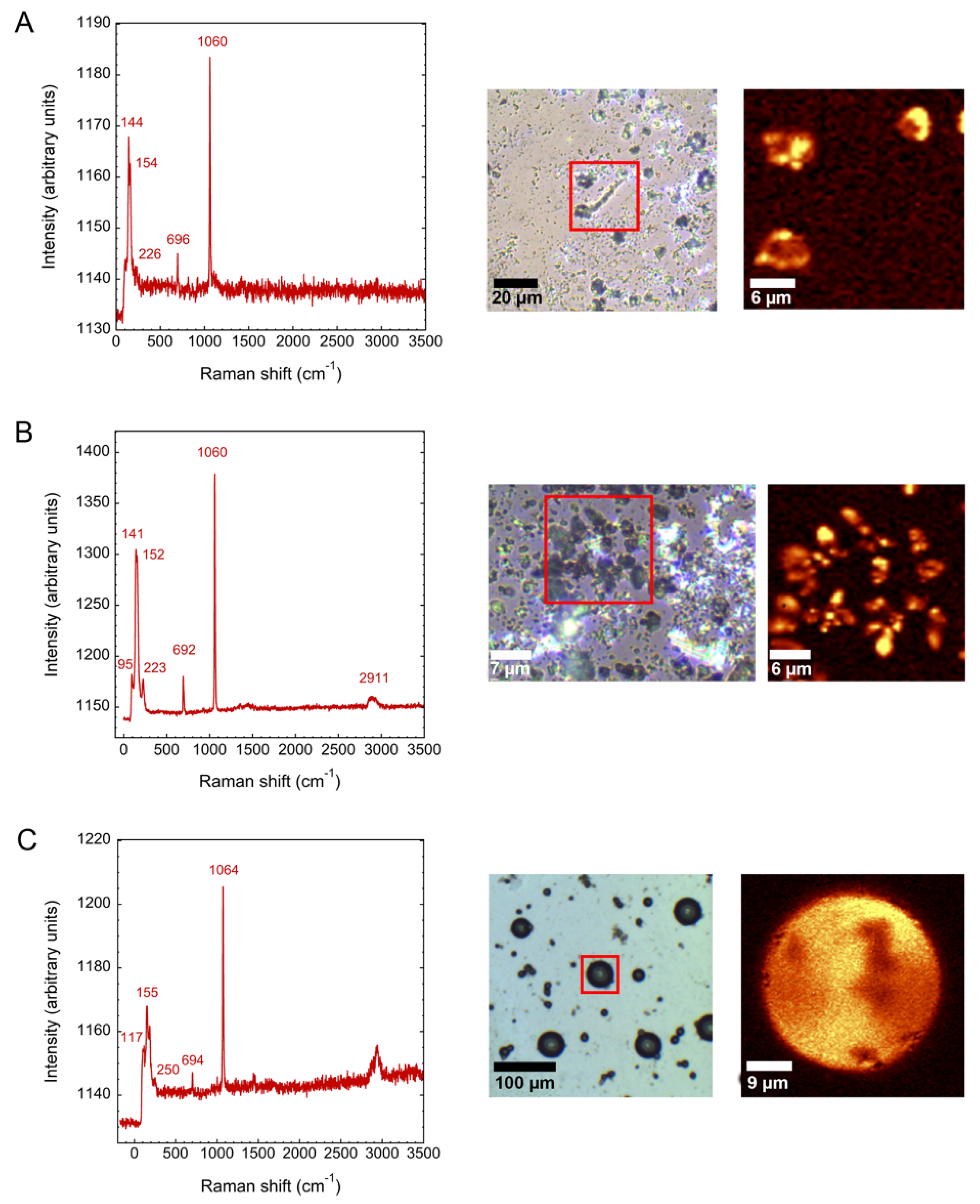
3.2. Calcium Silica Carbonate Biomorphs in the Presence of Silicatein-α
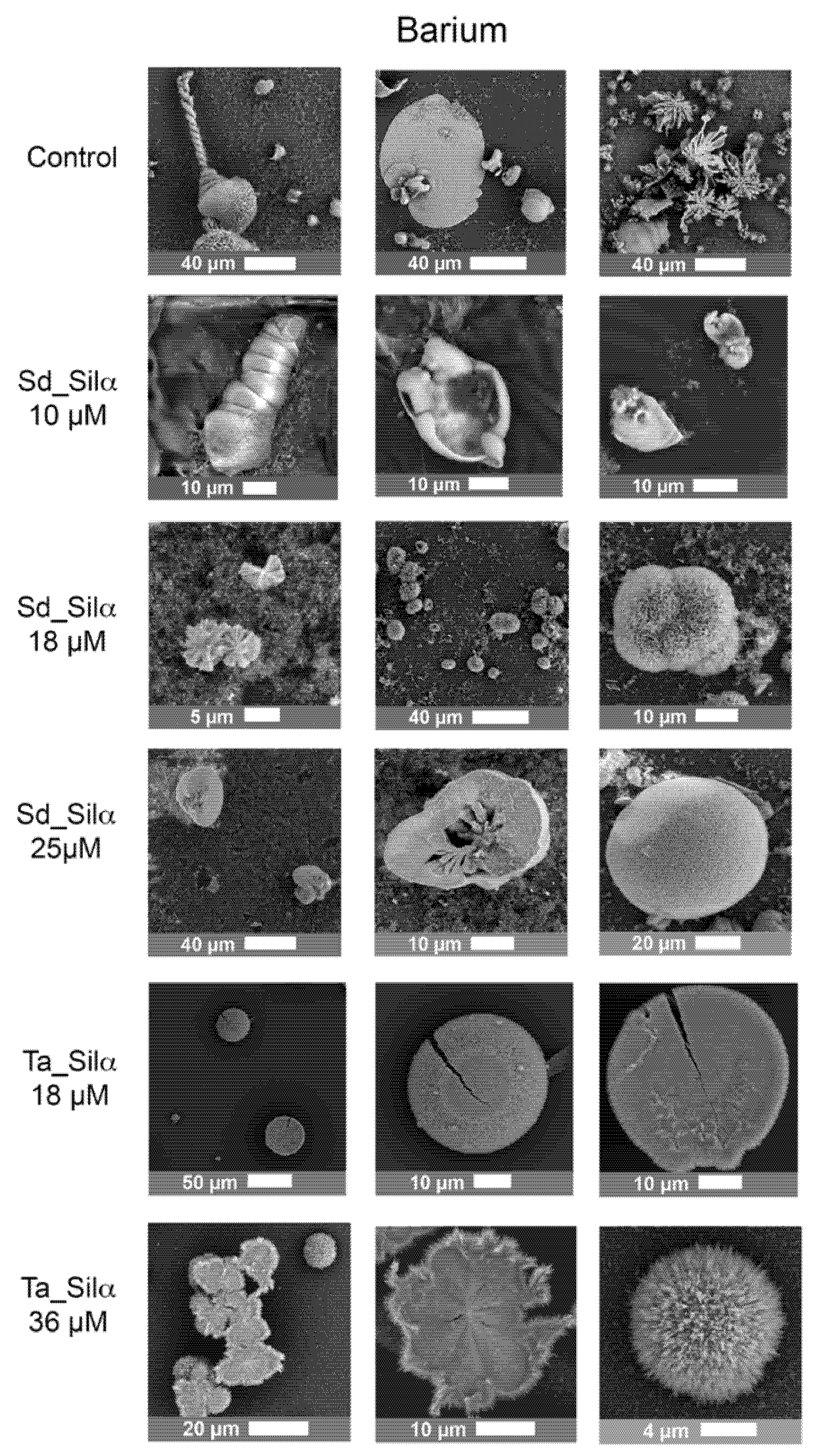
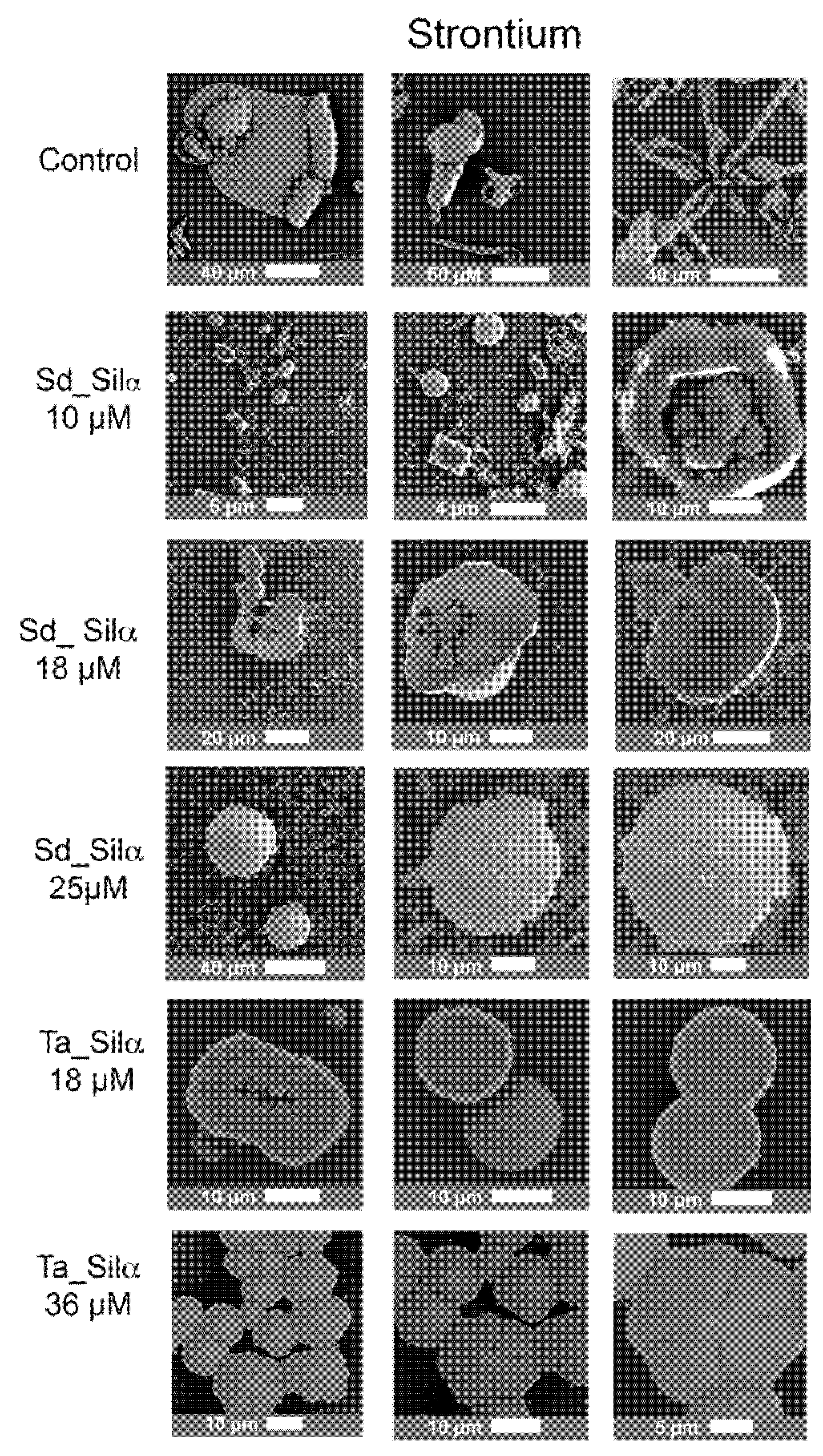
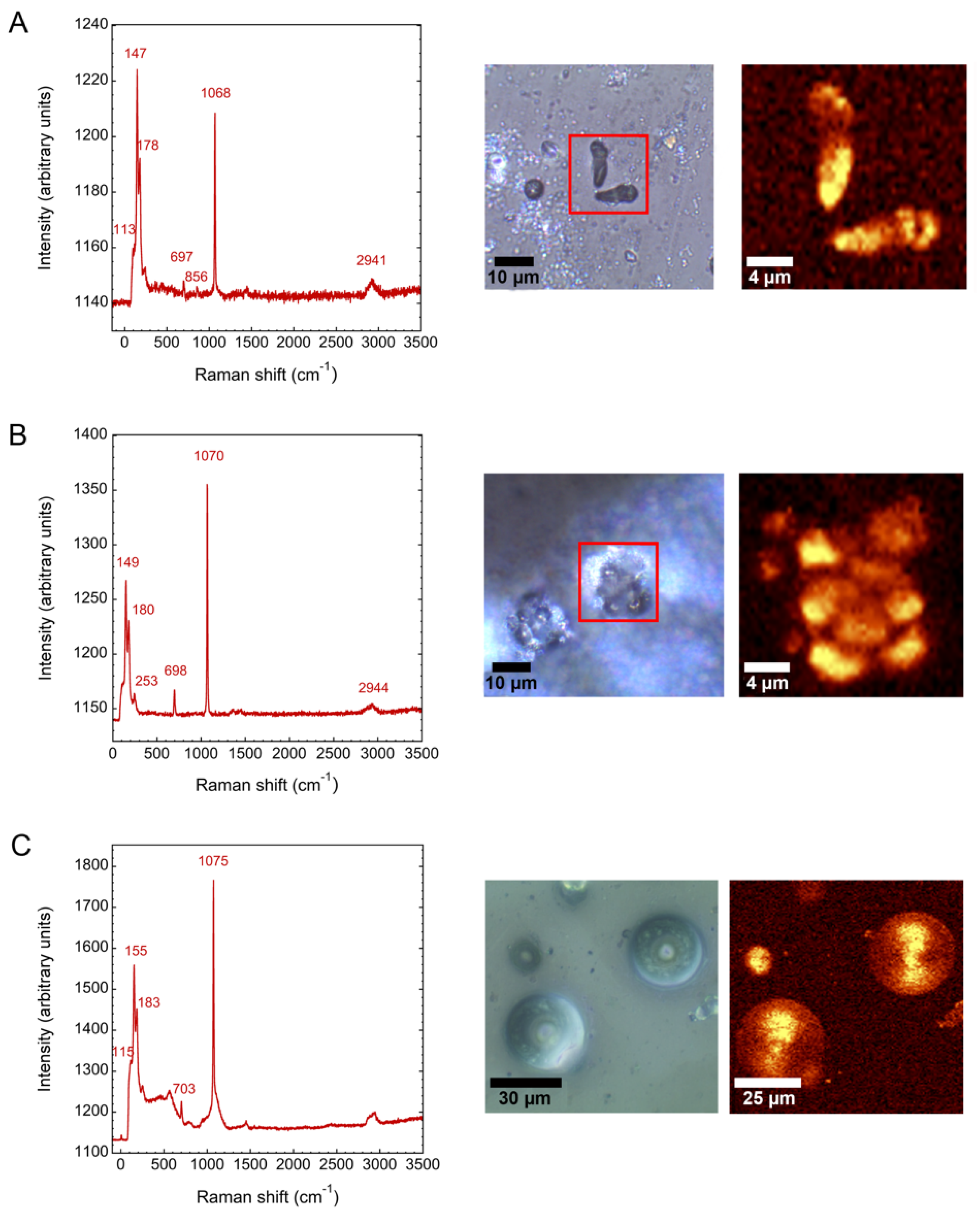
3.3. Barium and Strontiun Silica Carbonate Biomorphs in the Presence of Silicatein-α
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, R.Z.; Addadi, L.; Weiner, S. Design strategies of sea urchin teeth: Structure, composition and micromechanical relations to function. Philos. Trans. R. Soc. B Biol. Sci. 1997, 352, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, A.L. A New Theory of the Origin and Nature of Life. Science 1942, 96, 14. [Google Scholar] [CrossRef]
- Lillie, R.S.; Johnston, E.N. Precipitation-Structures Simulating Organic Growth. II. A Contribution to the Physico-Chemical Analysis of Growth and Heredity. Biol. Bull. 1919, 36, 225–272. [Google Scholar] [CrossRef]
- Ruiz, J.M.G.; Kubatko, K.-A.H.; Helean, K.B.; Navrotsky, A.; Burns, P.C. Self-Assembled Silica-Carbonate Structures and Detection of Ancient Microfossils. Science 2003, 302, 1194–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, S.T.; Carnerup, A.M.; Larsson, A.-K.; Christy, A.G.; García-Ruiz, J.M. Self-assembly of carbonate-silica colloids: Between living and non-living form. Phys. A: Stat. Mech. Its Appl. 2004, 339, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Nakouzi, E.; Knoll, P.; Steinbock, O. Biomorph growth in single-phase systems: Expanding the structure spectrum and pH range. Chem. Commun. 2015, 52, 2107–2110. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, J.M.; Melero-García, E.; Hyde, S.T. Morphogenesis of Self-Assembled Nanocrystalline Materials of Barium Carbonate and Silica. Science 2009, 323, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Noorduin, W.L.; Grinthal, A.; Mahadevan, L.; Aizenberg, J. Rationally Designed Complex, Hierarchical Microarchitectures. Science 2013, 340, 832–837. [Google Scholar] [CrossRef] [Green Version]
- Cuéllar-Cruz, M.; Moreno, A. Synthesis of Crystalline Silica-Carbonate Biomorphs of Ba(II) under the Presence of RNA and Positively and Negatively Charged ITO Electrodes: Obtainment of Graphite via Bioreduction of CO2 and Its Implications to the Chemical Origin of Life on Primitive Earth. ACS Omega 2020, 5, 5460–5469. [Google Scholar] [CrossRef]
- Pérez, K.S.; Moreno, A. Influence of Pyruvic Acid and UV Radiation on the Morphology of Silica-carbonate Crystalline Biomorphs. Crystals 2019, 9, 67. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, J.M.; Nakouzi, E.; Kotopoulou, E.; Tamborrino, L.; Steinbock, O. Biomimetic mineral self-organization from silica-rich spring waters. Sci. Adv. 2017, 3, e1602285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opel, J.; Kellermeier, M.; Sickinger, A.; Morales, J.; Cölfen, H.; Garcia-Ruiz, J.-M. Structural Transition of Inorganic Silica–Carbonate Composites Towards Curved Lifelike Morphologies. Minerals 2018, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Opel, J.; Wimmer, F.P.; Kellermeier, M.; Cölfen, H. Functionalisation of silica–carbonate biomorphs. Nanoscale Horiz. 2016, 1, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Morales, J.; García-Ruiz, J.M. Growth behaviour of silica/carbonate nanocrystalline composites of calcite and aragonite. J. Mater. Chem. B 2017, 5, 1658–1663. [Google Scholar] [CrossRef] [Green Version]
- Kellermeier, M.; Glaab, F.; Carnerup, A.M.; Drechsler, M.; Gossler, B.; Hyde, S.T.; Kunz, W. Additive-induced morphological tuning of self-assembled silica–barium carbonate crystal aggregates. J. Cryst. Growth 2009, 311, 2530–2541. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Islas, S.R.; Gonzalez, G.; Moreno, A. Influence of Nucleic Acids on the Synthesis of Crystalline Ca(II), Ba(II), and Sr(II) Silica–Carbonate Biomorphs: Implications for the Chemical Origin of Life on Primitive Earth. Cryst. Growth Des. 2019, 19, 4667–4682. [Google Scholar] [CrossRef]
- Elejalde-Cadena, N.R.; Cuéllar-Cruz, M.; Moreno, A. The role of silica and alkaline earth metals with biomolecules in the biomineralization processes: The eggshell’s formation and the crystallization in vivo for x-ray crystallography. Prog. Cryst. Growth Charact. Mater. 2020, 66, 100473. [Google Scholar] [CrossRef]
- Kröger, N.; Deutzmann, R.; Sumper, M. Polycationic Peptides from Diatom Biosilica That Direct Silica Nanosphere Formation. Science 1999, 286, 1129–1132. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Cha, J.; Stucky, G.D.; Morse, D.E. Silicatein: Cathepsin L-like protein in sponge biosilica. Proc. Natl. Acad. Sci. USA 1998, 95, 6234–6238. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Amano, T.; Bari, R.; Weaver, J.C.; Arima, J.; Mori, N. Glassin, a histidine-rich protein from the siliceous skeletal system of the marine sponge Euplectella, directs silica polycondensation. Proc. Natl. Acad. Sci. USA 2015, 112, 11449–11454. [Google Scholar] [CrossRef] [Green Version]
- Gehling, J.G.; Rigby, J.K. Long expected sponges from the Neoproterozoic Ediacara fauna of South Australia. J. Paleéontol. 1996, 70, 185–195. [Google Scholar] [CrossRef]
- Yin, Z.; Zhu, M.; Davidson, E.H.; Bottjer, D.J.; Zhao, F.; Tafforeau, P. Sponge grade body fossil with cellular resolution dating 60 Myr before the Cambrian. Proc. Natl. Acad. Sci. USA 2015, 112, E1453–E1460. [Google Scholar] [CrossRef] [Green Version]
- Nosenko, T.; Schreiber, F.; Adamska, M.; Adamski, M.; Eitel, M.; Hammel, J.; Maldonado, M.; Müller, W.E.; Nickel, M.; Schierwater, B.; et al. Deep metazoan phylogeny: When different genes tell different stories. Mol. Phylogenet. Evol. 2013, 67, 223–233. [Google Scholar] [CrossRef]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World Porifera Database. Available online: http://www.marinespecies.org/porifera (accessed on 14 April 2021).
- Cha, J.N.; Shimizu, K.; Zhou, Y.; Christiansen, S.C.; Chmelka, B.F.; Stucky, G.D.; Morse, D.E. Silicatein filaments and subunits from a marine sponge direct the polymerization of silica and silicones in vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 361–365. [Google Scholar] [CrossRef] [Green Version]
- Müller, W.E.G.; Rothenberger, M.; Boreiko, A.; Tremel, W.; Reiber, A.; Schröder, H.C. Formation of siliceous spicules in the marine demosponge Suberites domuncula. Cell Tissue Res. 2005, 321, 285–297. [Google Scholar] [CrossRef]
- Murr, M.M.; Morse, D.E. Fractal intermediates in the self-assembly of silicatein filaments. Proc. Natl. Acad. Sci. USA 2005, 102, 11657–11662. [Google Scholar] [CrossRef] [Green Version]
- Schloßmacher, U.; Wiens, M.; Schröder, H.C.; Jochum, K.P.; Wang, X.; Müller, W.E.G. Silintaphin-1—Interaction with silicatein during structure-guiding bio-silica formation. FEBS J. 2011, 278, 1145–1155. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Schloßmacher, U.; Wang, X.; Boreiko, A.; Brandt, D.; Wolf, S.E.; Tremel, W.; Schröder, H.C. Poly(silicate)-metabolizing silicatein in siliceous spicules and silicasomes of demosponges comprises dual enzymatic activities (silica polymerase and silica esterase). FEBS J. 2007, 275, 362–370. [Google Scholar] [CrossRef]
- Dakhili, S.Y.T.; Caslin, S.A.; Faponle, A.S.; Quayle, P.; De Visser, S.P.; Wong, L.S. Recombinant silicateins as model biocatalysts in organosiloxane chemistry. Proc. Natl. Acad. Sci. USA 2017, 114, E5285–E5291. [Google Scholar] [CrossRef] [Green Version]
- Kisailus, D.; Choi, J.H.; Weaver, J.C.; Yang, W.; Morse, D.E. Enzymatic Synthesis and Nanostructural Control of Gallium Oxide at Low Temperature. Adv. Mater. 2005, 17, 314–318. [Google Scholar] [CrossRef]
- Kisailus, D.; Truong, Q.; Amemiya, Y.; Weaver, J.C.; Morse, D.E. Self-assembled bifunctional surface mimics an enzymatic and templating protein for the synthesis of a metal oxide semiconductor. Proc. Natl. Acad. Sci. USA 2006, 103, 5652–5657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahir, M.N.; Théato, P.; Müller, W.E.G.; Schröder, H.C.; Borejko, A.; Faiss, S.; Janshoff, A.; Huth, J.; Tremel, W. Formation of layered titania and zirconia catalysed by surface-bound silicatein. Chem. Commun. 2005, 5533–5535. [Google Scholar] [CrossRef]
- Sánchez-Puig, N.; Guerra-Flores, E.; López-Sánchez, F.; Juárez-Espinoza, P.A.; Ruiz-Arellano, R.; González-Muñoz, R.; Arreguin-Espinosa, R.; Moreno, A. Controlling the morphology of silica–carbonate biomorphs using proteins involved in biomineralization. J. Mater. Sci. 2011, 47, 2943–2950. [Google Scholar] [CrossRef]
- Schröder, H.C.; Wang, X.; Manfrin, A.; Yu, S.-H.; Grebenjuk, V.A.; Korzhev, M.; Wiens, M.; Schlossmacher, U.; Müller, W.E. Acquisition of Structure-guiding and Structure-forming Properties during Maturation from the Pro-silicatein to the Silicatein Form. J. Biol. Chem. 2012, 287, 22196–22205. [Google Scholar] [CrossRef] [Green Version]
- Simpson, T.L.; Volcani, B.E. Silicon and Siliceous Structures in Biological Systems; Springer: Berlin/Heidelberg, Germany, 1981; p. 587. [Google Scholar]
- Voronkov, M.G.; Zelchan, G.I.; Lukevits, E.J. Silicon and Life; Zinatne Publishing: Vilnius, Lithuania, 1977. [Google Scholar]
- Ruiz-Arellano, R.R.; Moreno, A. Obtainment of Spherical-Shaped Calcite Crystals Induced by Intramineral Proteins Isolated from Eggshells of Ostrich and Emu. Cryst. Growth Des. 2014, 14, 5137–5143. [Google Scholar] [CrossRef]
- De La Pierre, M.; Carteret, C.; Maschio, L.; André, E.; Orlando, R.; Dovesi, R. The Raman spectrum of CaCO3polymorphs calcite and aragonite: A combined experimental and computational study. J. Chem. Phys. 2014, 140, 164509. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Grajeda, J.P.; Moreno, A.; Romero, A. Crystal Structure of Ovocleidin-17, a Major Protein of the Calcified Gallus gallus Eggshell. J. Biol. Chem. 2004, 279, 40876–40881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Li, X. Uncovering Aragonite Nanoparticle Self-assembly in Nacre—A Natural Armor. Cryst. Growth Des. 2012, 12, 4306–4310. [Google Scholar] [CrossRef]
- García-Ruiz, J.; Amorós, J. Morphological aspects of some symmetrical crystal aggregates grown by silica gel technique. J. Cryst. Growth 1981, 55, 379–383. [Google Scholar] [CrossRef]
- Schopf, J.W. Solution to Darwin’s dilemma: Discovery of the missing Precambrian record of life. Proc. Natl. Acad. Sci. USA 2000, 97, 6947–6953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Stefano, M.; Marino, D. Morphology and taxonomy of Amphicocconeisgen. nov. (Achnanthales, Bacillariophyceae, Bacillariophyta) with considerations on its relationship to other monoraphid diatom genera. Eur. J. Phycol. 2003, 38, 361–370. [Google Scholar] [CrossRef]
- Pamirsky, I.E.; Golokhvast, K.S. Silaffins of Diatoms: From Applied Biotechnology to Biomedicine. Mar. Drugs 2013, 11, 3155–3167. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurti, D. The Raman spectra of aragonite, strontianite and witherite. Proc. Indian Acad. Sci.—Sect. A 1960, 51, 285–295. [Google Scholar] [CrossRef]
- Görlich, S.; Samuel, A.J.; Best, R.J.; Seidel, R.; Vacelet, J.; Leonarski, F.K.; Tomizaki, T.; Rellinghaus, B.; Pohl, D.; Zlotnikov, I. Natural hybrid silica/protein superstructure at atomic resolution. Proc. Natl. Acad. Sci. USA 2020, 117, 31088–31093. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Puig, N.; Cuéllar-Cruz, M.; Islas, S.R.; Tapia-Vieyra, J.V.; Arreguín-Espinosa, R.A.; Moreno, A. The Influence of Silicateins on the Shape and Crystalline Habit of Silica Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr). Crystals 2021, 11, 438. https://doi.org/10.3390/cryst11040438
Sánchez-Puig N, Cuéllar-Cruz M, Islas SR, Tapia-Vieyra JV, Arreguín-Espinosa RA, Moreno A. The Influence of Silicateins on the Shape and Crystalline Habit of Silica Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr). Crystals. 2021; 11(4):438. https://doi.org/10.3390/cryst11040438
Chicago/Turabian StyleSánchez-Puig, Nuria, Mayra Cuéllar-Cruz, Selene R. Islas, Juana V. Tapia-Vieyra, Roberto A. Arreguín-Espinosa, and Abel Moreno. 2021. "The Influence of Silicateins on the Shape and Crystalline Habit of Silica Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr)" Crystals 11, no. 4: 438. https://doi.org/10.3390/cryst11040438
APA StyleSánchez-Puig, N., Cuéllar-Cruz, M., Islas, S. R., Tapia-Vieyra, J. V., Arreguín-Espinosa, R. A., & Moreno, A. (2021). The Influence of Silicateins on the Shape and Crystalline Habit of Silica Carbonate Biomorphs of Alkaline Earth Metals (Ca, Ba, Sr). Crystals, 11(4), 438. https://doi.org/10.3390/cryst11040438