
Characterization of Dental Pulp Stem Cells Response to Bone Substitutes Biomaterials in Dentistry


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human DPSCs Isolation and Immune Selection
2.2. Culture of hDPSCs in Presence of Bone Substitutes Biomaterials
2.3. PBMCs Isolation and Co-Culture with hDPSCs
2.4. Evaluation of Cell Morphology and Stemness Markers in hDPSCs
2.5. Cell Adhesion, Proliferation and Viability Bioluminescent Assays
2.6. Biological Effects of BSBs Treatment on hDPSCs
2.7. Evaluation of Inflammatory Cytokines in Cell Culture Media
2.8. Evaluation of Osteogenic Differentiation
2.9. Statistical Analysis
3. Results
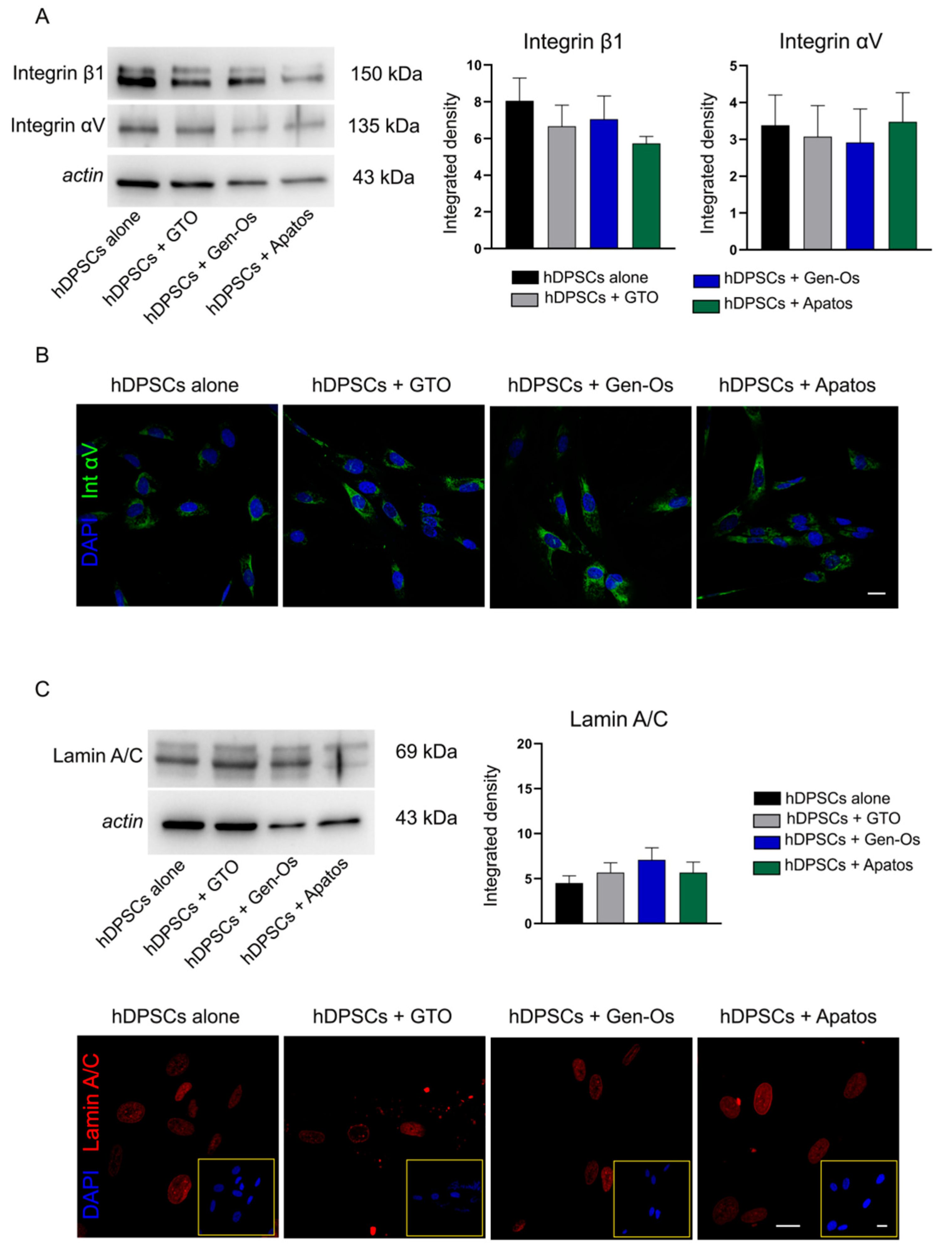
3.1. Analysis of the Effects Induced by BSBs on hDPSCs Morphology and Stemness Properties
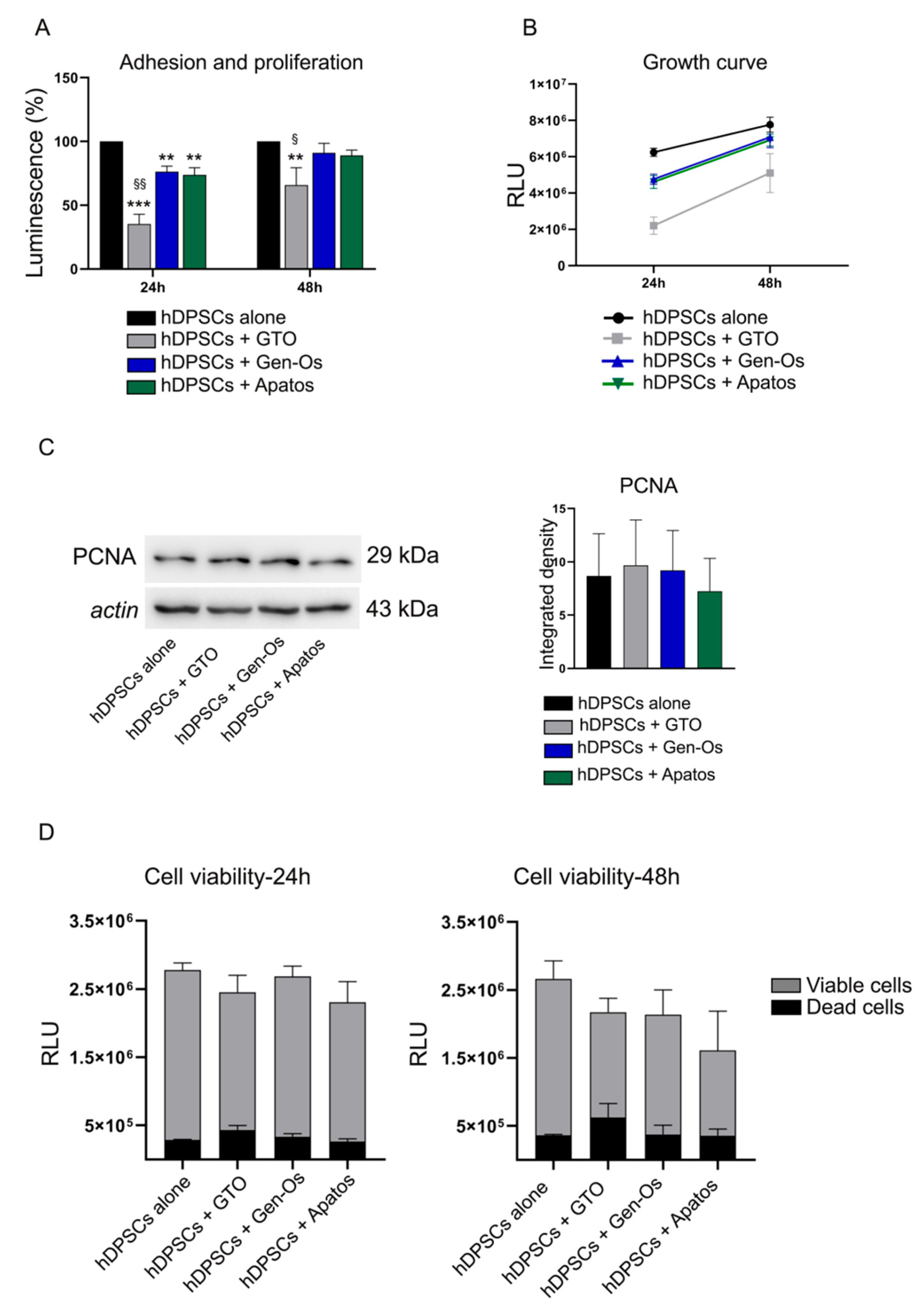
3.2. Investigation of Stem Cell Adhesion, Proliferation and Viability after BSBs Treatments
3.3. Evaluation of Biological Effects of BSBs Treatment on hDPSCs
3.4. Osteogenic Differentiation of hDPSCs Treated with BSBs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barone, A.; Toti, P.; Quaranta, A.; Alfonsi, F.; Cucchi, A.; Calvo-Guirado, J.L.; Negri, B.; di Felice, R.; Covani, U. Volumetric Analysis of Remodelling Pattern after Ridge Preservation Comparing Use of Two Types of Xenografts. A Multicentre Randomized Clinical Trial. Clin. Oral Implant. Res. 2016, 27, e105–e115. [Google Scholar] [CrossRef] [PubMed]
- Wallace, S.S.; Froum, S.J. Effect of Maxillary Sinus Augmentation on the Survival of Endosseous Dental Implants. A Systematic Review. Ann. Periodontol. 2003, 8, 328–343. [Google Scholar] [CrossRef] [Green Version]
- Del Fabbro, M.; Rosano, G.; Taschieri, S. Implant Survival Rates after Maxillary Sinus Augmentation. Eur. J. Oral Sci. 2008, 116, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.K.; Yun, P.Y.; Lee, H.J.; Ahn, J.Y.; Kim, S.G. Ridge Preservation of the Molar Extraction Socket Using Collagen Sponge and Xenogeneic Bone Grafts. Implant. Dent. 2011, 20, 267–272. [Google Scholar] [CrossRef]
- Mardas, N.; D’Aiuto, F.; Mezzomo, L.; Arzoumanidi, M.; Donos, N. Radiographic Alveolar Bone Changes Following Ridge Preservation with Two Different Biomaterials. Clin. Oral Implant. Res. 2011, 22, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Gholami, G.A.; Najafi, B.; Mashhadiabbas, F.; Goetz, W.; Najafi, S. Clinical, Histologic and Histomorphometric Evaluation of Socket Preservation Using a Synthetic Nanocrystalline Hydroxyapatite in Comparison with a Bovine Xenograft: A Randomized Clinical Trial. Clin. Oral Implant. Res. 2012, 23, 1198–1204. [Google Scholar] [CrossRef] [PubMed]
- Festa, V.M.; Addabbo, F.; Laino, L.; Femiano, F.; Rullo, R. Porcine-Derived Xenograft Combined with a Soft Cortical Membrane versus Extraction Alone for Implant Site Development: A Clinical Study in Humans. Clin. Implant. Dent. Relat. Res. 2013, 15, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Pradel, W.; Eckelt, U.; Lauer, G. Bone Regeneration after Enucleation of Mandibular Cysts: Comparing Autogenous Grafts from Tissue-Engineered Bone and Iliac Bone. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2006, 101, 285–290. [Google Scholar] [CrossRef]
- Velasco-Ortega, E.; Valente, N.A.; Iezzi, G.; Petrini, M.; Derchi, G.; Barone, A. Maxillary Sinus Augmentation with Three Different Biomaterials: Histological, Histomorphometric, Clinical, and Patient-Reported Outcomes from a Randomized Controlled Trial. Clin. Implant. Dent. Relat. Res. 2021, 23, 86–95. [Google Scholar] [CrossRef]
- Hartlev, J.; Erik Nørholt, S.; Spin-Neto, R.; Kraft, D.; Schou, S.; Isidor, F. Histology of Augmented Autogenous Bone Covered by a Platelet-Rich Fibrin Membrane or Deproteinized Bovine Bone Mineral and a Collagen Membrane: A Pilot Randomized Controlled Trial. Clin. Oral Implant. Res. 2020, 31, 694–704. [Google Scholar] [CrossRef]
- Sakkas, A.; Wilde, F.; Heufelder, M.; Winter, K.; Schramm, A. Autogenous Bone Grafts in Oral Implantology—Is It Still a “Gold Standard”? A Consecutive Review of 279 Patients with 456 Clinical Procedures. Int. J. Implant. Dent. 2017, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, J.P.A.; ten Bruggenkate, C.M.; Krekeler, G.; Bram Tuinzing, D. Sinusfloor Elevation and Grafting with Autogenous Iliac Crest Bone. Clin. Oral Implant. Res. 1998, 9, 429–435. [Google Scholar] [CrossRef]
- Olson, J.W.; Dent, C.D.; Morris, H.F.; Ochi, S. Long-Term Assessment (5 to 71 Months) of Endosseous Dental Implants Placed in the Augmented Maxillary Sinus. Ann. Periodontol./Am. Acad. Periodontol. 2000, 5, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Pikdöken, L.; Gürbüzer, B.; Küçükodaci, Z.; Urhan, M.; Bariş, E.; Tezulaş, E. Scintigraphic, Histologic, and Histomorphometric Analyses of Bovine Bone Mineral and Autogenous Bone Mixture in Sinus Floor Augmentation: A Randomized Controlled Trial--Results after 4 Months of Healing. J. Oral Maxillofac. Surg. 2011, 69, 160–169. [Google Scholar] [CrossRef]
- Tomas, M.; Čandrlić, M.; Juzbašić, M.; Ivanišević, Z.; Matijević, N.; Včev, A.; Peloza, O.C.; Matijević, M.; Kačarević, Ž.P. Synthetic Injectable Biomaterials for Alveolar Bone Regeneration in Animal and Human Studies. Materials 2021, 14, 2858. [Google Scholar] [CrossRef]
- Yamada, M.; Egusa, H. Current Bone Substitutes for Implant Dentistry. J. Prosthodont. Res. 2018, 62, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.; Henriques, J.; Martins, G.; Guerra, F.; Judas, F.; Figueiredo, H. Physicochemical Characterization of Biomaterials Commonly Used in Dentistry as Bone Substitutes--Comparison with Human Bone. J. Biomed. Mater. Res. B Appl. Biomater. 2010, 92, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Barone, A.; Toti, P.; Quaranta, A.; Alfonsi, F.; Cucchi, A.; Negri, B.; di Felice, R.; Marchionni, S.; Calvo-Guirado, J.L.; Covani, U.; et al. Clinical and Histological Changes after Ridge Preservation with Two Xenografts: Preliminary Results from a Multicentre Randomized Controlled Clinical Trial. J. Clin. Periodontol. 2017, 44, 204–214. [Google Scholar] [CrossRef]
- Orsini, G.; Scarano, A.; Piattelli, M.; Piccirilli, M.; Caputi, S.; Piattelli, A. Histologic and Ultrastructural Analysis of Regenerated Bone in Maxillary Sinus Augmentation Using a Porcine Bone-Derived Biomaterial. J. Periodontol. 2006, 77, 1984–1990. [Google Scholar] [CrossRef]
- Marconcini, S.; Giammarinaro, E.; Derchi, G.; Alfonsi, F.; Covani, U.; Barone, A. Clinical Outcomes of Implants Placed in Ridge-Preserved versus Nonpreserved Sites: A 4-Year Randomized Clinical Trial. Clin. Implant. Dent. Relat. Res. 2018, 20, 906–914. [Google Scholar] [CrossRef]
- Angelova Volponi, A.; Zaugg, L.K.; Neves, V.; Liu, Y.; Sharpe, P.T. Tooth Repair and Regeneration. Curr. Oral Health Rep. 2018, 5, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, L.; Decker, A.M.; Nibali, L.; Pilipchuk, S.P.; Berglundh, T.; Giannobile, W.V. Regenerative Medicine for Periodontal and Peri-Implant Diseases. J. Dent. Res. 2016, 95, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.; Pisciotta, A.; Bertoni, L.; Berni, M.; Gambardella, A.; Visani, A.; Russo, A.; de Pol, A.; Carnevale, G. Osteogenic Differentiation of HDPSCs on Biogenic Bone Apatite Thin Films. Stem Cells Int. 2017, 2017, 3579283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Tinco, R.; Bertani, G.; Pisciotta, A.; Bertoni, L.; Pignatti, E.; Maccaferri, M.; Bertacchini, J.; Sena, P.; Vallarola, A.; Tupler, R.; et al. Role of PD-L1 in Licensing Immunoregulatory Function of Dental Pulp Mesenchymal Stem Cells. Stem Cell Res. Ther. 2021, 12, 598. [Google Scholar] [CrossRef] [PubMed]
- Machado-Pineda, Y.; Cardeñes, B.; Reyes, R.; López-Martín, S.; Toribio, V.; Sánchez-Organero, P.; Suarez, H.; Grötzinger, J.; Lorenzen, I.; Yáñez-Mó, M.; et al. CD9 Controls Integrin A5β1-Mediated Cell Adhesion by Modulating Its Association with the Metalloproteinase ADAM17. Front. Immunol. 2018, 9, 2474. [Google Scholar] [CrossRef] [Green Version]
- Dubik, N.; Mai, S. Lamin A/C: Function in Normal and Tumor Cells. Cancers 2020, 12, 3688. [Google Scholar] [CrossRef] [PubMed]
- Malashicheva, A.; Perepelina, K. Diversity of Nuclear Lamin A/C Action as a Key to Tissue-Specific Regulation of Cellular Identity in Health and Disease. Front. Cell Dev. Biol. 2021, 9, 2834. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zhang, L.; Xu, C.; Zheng, C.; Han, Y.; Li, W.; Huang, Y.; Zhang, X.; Shao, C.; et al. An Osteopontin-Integrin Interaction Plays a Critical Role in Directing Adipogenesis and Osteogenesis by Mesenchymal Stem Cells. Stem Cells 2014, 32, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.X.; Denhardt, D.T. Osteopontin: Role in Immune Regulation and Stress Responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Mortada, I.; Mortada, R. Dental Pulp Stem Cells and Osteogenesis: An Update. Cytotechnology 2018, 70, 1479. [Google Scholar] [CrossRef]
- Chaikiawkeaw, D.; Khorattanakulchai, N.; Nammultriputtar, K.; Rattanapisit, K.; Everts, V.; Kubera, A.; Phoolchareon, W.; Pavasant, P. Osteopontin Induces Osteogenic Differentiation by Human Periodontal Ligament Cells via Calcium Binding Domain-Activin Receptor-like Kinase (ALK-1) Interaction. J. Periodontol. 2021, 93, e11–e23. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Afif, I.Y.; Maula, M.I.; Winarni, T.I.; Tauviqirrahman, M.; Akbar, I.; Basri, H.; van der Heide, E.; Jamari, J. Tresca Stress Simulation of Metal-on-Metal Total Hip Arthroplasty during Normal Walking Activity. Materials 2021, 14, 7554. [Google Scholar] [CrossRef] [PubMed]
- Jamari, J.; Ammarullah, M.I.; Saad, A.P.M.; Syahrom, A.; Uddin, M.; van der Heide, E.; Basri, H. The Effect of Bottom Profile Dimples on the Femoral Head on Wear in Metal-on-Metal Total Hip Arthroplasty. J. Funct. Biomater. 2021, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Yang, R.; Cooper, P.R.; Khurshid, Z.; Shavandi, A.; Ratnayake, J. Bone Grafts and Substitutes in Dentistry: A Review of Current Trends and Developments. Molecules 2021, 26, 3007. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M. Bone Tissue Regeneration: Biology, Strategies and Interface Studies. Prog. Biomater. 2019, 8, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Pagni, G.; Pellegrini, G.; Giannobile, W.V.; Rasperini, G. Postextraction Alveolar Ridge Preservation: Biological Basis and Treatments. Int. J. Dent. 2012, 2012, 151030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sculean, A.; Gruber, R.; Bosshardt, D.D. Soft Tissue Wound Healing around Teeth and Dental Implants. J. Clin. Periodontol. 2014, 41, S6–S22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinheinz, J.; Stratmann, U.; Joos, U.; Wiesmann, H.P. VEGF-Activated Angiogenesis During Bone Regeneration. J. Oral Maxillofac. Surg. 2005, 63, 1310–1316. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Tan, Y.Y.; Wong, R.; Wenden, A.; Zhang, L.K.; Rabie, A.B.M. The Role of Vascular Endothelial Growth Factor in Ossification. Int. J. Oral Sci. 2012, 4, 64–68. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Cui, Z.; Wang, L.; Xia, Z.; Hu, Y.; Xian, L.; Li, C.; Xie, L.; Crane, J.; Wan, M.; et al. PDGF-BB Secreted by Preosteoclasts Induces Angiogenesis during Coupling with Osteogenesis. Nat. Med. 2014, 20, 1270–1278. [Google Scholar] [CrossRef] [Green Version]
- Dirckx, N.; Moorer, M.C.; Clemens, T.L.; Riddle, R.C. The Role of Osteoblasts in Energy Homeostasis. Nat. Rev. Endocrinol. 2019, 15, 651–665. [Google Scholar] [CrossRef]
- Lee, M.N.; Hwang, H.S.; Oh, S.H.; Roshanzadeh, A.; Kim, J.W.; Song, J.H.; Kim, E.S.; Koh, J.T. Elevated Extracellular Calcium Ions Promote Proliferation and Migration of Mesenchymal Stem Cells via Increasing Osteopontin Expression. Exp. Mol. Med. 2018, 50, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.S.; Cabral, J.M.S.; da Silva, C.L.; Vashishth, D. Synergistic Effect of Extracellularly Supplemented Osteopontin and Osteocalcin on Stem Cell Proliferation, Osteogenic Differentiation, and Angiogenic Properties. J. Cell Biochem. 2019, 120, 6555–6569. [Google Scholar] [CrossRef] [PubMed]
- Giachelli, C.M.; Steitz, S. Osteopontin: A Versatile Regulator of Inflammation and Biomineralization. Matrix Biol. 2000, 19, 615–622. [Google Scholar] [CrossRef]
- Hadjidakis, D.J.; Androulakis, I.I. Bone Remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Tinco, R.; Consolo, U.; Pisciotta, A.; Orlandi, G.; Bertani, G.; Nasi, M.; Bertacchini, J.; Carnevale, G. Characterization of Dental Pulp Stem Cells Response to Bone Substitutes Biomaterials in Dentistry. Polymers 2022, 14, 2223. https://doi.org/10.3390/polym14112223
Di Tinco R, Consolo U, Pisciotta A, Orlandi G, Bertani G, Nasi M, Bertacchini J, Carnevale G. Characterization of Dental Pulp Stem Cells Response to Bone Substitutes Biomaterials in Dentistry. Polymers. 2022; 14(11):2223. https://doi.org/10.3390/polym14112223
Chicago/Turabian StyleDi Tinco, Rosanna, Ugo Consolo, Alessandra Pisciotta, Giulia Orlandi, Giulia Bertani, Milena Nasi, Jessika Bertacchini, and Gianluca Carnevale. 2022. "Characterization of Dental Pulp Stem Cells Response to Bone Substitutes Biomaterials in Dentistry" Polymers 14, no. 11: 2223. https://doi.org/10.3390/polym14112223
APA StyleDi Tinco, R., Consolo, U., Pisciotta, A., Orlandi, G., Bertani, G., Nasi, M., Bertacchini, J., & Carnevale, G. (2022). Characterization of Dental Pulp Stem Cells Response to Bone Substitutes Biomaterials in Dentistry. Polymers, 14(11), 2223. https://doi.org/10.3390/polym14112223