Environmental Factors Affecting the Mineralization of Crop Residues



{kind=link}
{kind=link}
Abstract
:1. Introduction
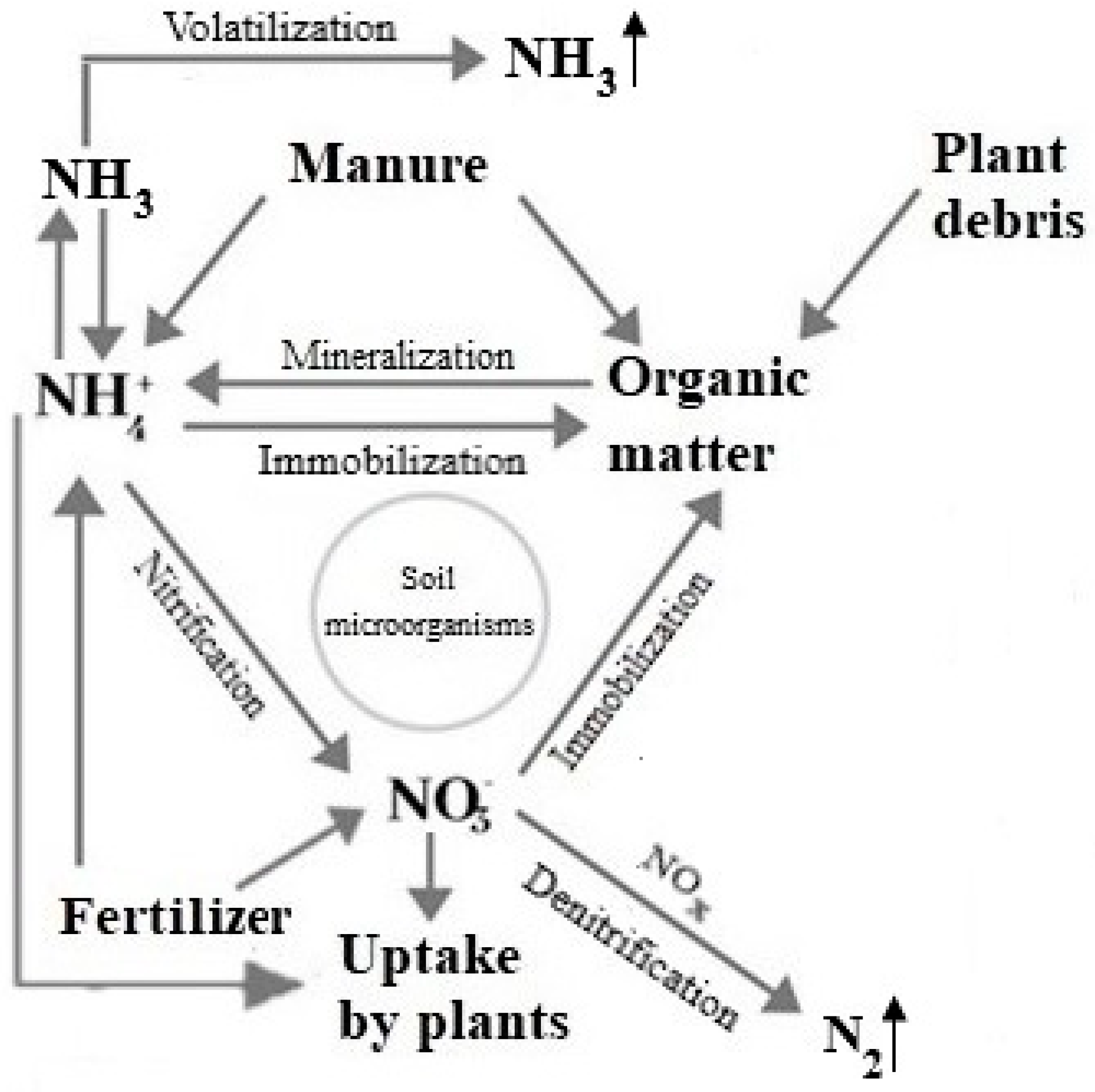
2. Microbiological Mineralization of Crop Residues
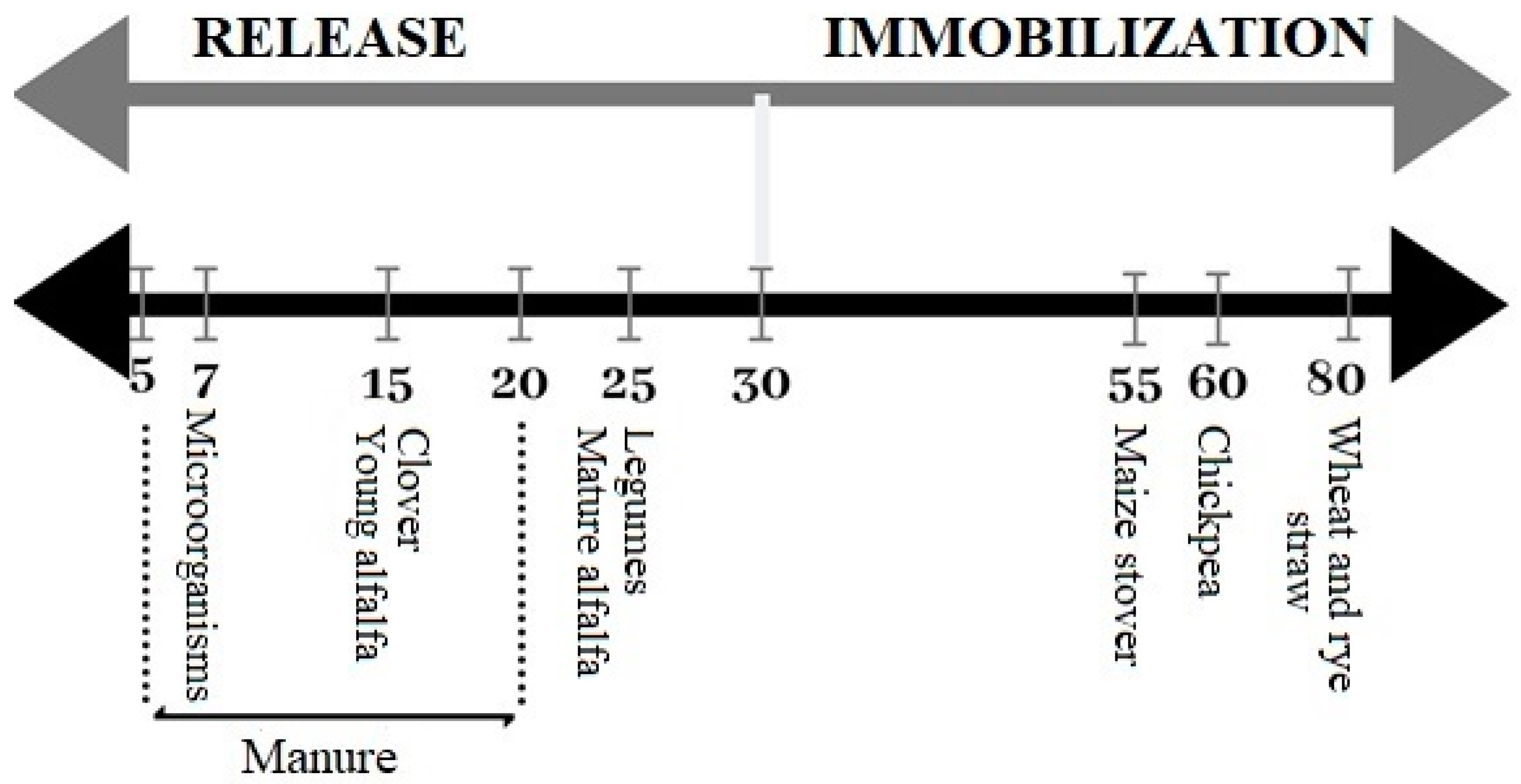
3. The Influence of Environmental Factors on the Rate of Mineralization of Crop Residues
4. The Benefits of Using Crop Residues in Agricultural Production
5. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Franzluebbers, A.J. Ecology: Cycling of carbon and nitrogen. In Encyclopedia of Soil Science; Lal, R., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 711–715. [Google Scholar]
- Lal, R. World crop residues production and implications of its use as a biofuel. Environ. Int. 2005, 31, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.K.; Tahir, M.M.; Sabir, N.; Khurshid, M. Impact of the addition of different plant residues on nitrogen mineralization–immobilization turnover and carbon content of a soil incubated under laboratory conditions. Solid Earth 2015, 6, 197–205. [Google Scholar] [CrossRef] [Green Version]
- Khalil, M.I.; Hossain, M.B.; Schmidhalter, U. Carbon and nitrogen mineralization in different upland soils of the subtropics treated with organic materials. Soil Biol. Biochem. 2005, 37, 1507–1518. [Google Scholar] [CrossRef]
- Martens, D. Plant residue biochemistry regulates soil carbon cycling and carbon sequestration. Soil Biol. Biochem. 2000, 32, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Ajwa, H.A.; Tabatabai, M.A. Decomposition of different organic materials in soils. Biol. Fertil. Soils 1994, 18, 175–182. [Google Scholar] [CrossRef]
- Clayden, J.; Greeves, N.; Warren, S.; Wathers, P. Chemia Organiczna Cz. IV; WNT: Warszawa, Polska, 2016; p. 384. [Google Scholar]
- Hood, R.C. The Effect of soil temperature and moisture on organic matter decomposition and plant growth. Isotopes Environ. Health Stud. 2001, 37, 25–41. [Google Scholar] [CrossRef]
- Gough, C.M.; Seiler, J.R. The influence of environmental, soil carbon, root, and stand characteristics on soil CO2 efflux in loblolly pine (Pinus taeda L.) plantations located on the South Carolina Coastal Plain. For. Ecol. Manag. 2004, 191, 353–363. [Google Scholar] [CrossRef]
- Wallenstein, M.; Allison, S.D.; Ernakovich, J.; Steinweg, J.M.; Sinsabaugh, R. Controls on the temperature sensitivity of soil enzymes: A Key Driver of In Situ Enzyme Activity Rates. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 22, pp. 245–258. [Google Scholar]
- Butterly, C.R.; Baldock, J.A.; Tang, C. The contribution of crop residues to changes in soil pH under field conditions. Plant Soil 2013, 366, 185–198. [Google Scholar] [CrossRef]
- Hassan, W.; David, J.; Abbas, F. Effect of type and quality of two contrasting plant residues on CO2 emission potential of Ultisol soil: Implications for indirect influence of temperature and moisture. Cantena 2014, 114, 90–96. [Google Scholar] [CrossRef]
- Conant, R.T.; Ryan, M.G.; Ågren, G.I.; Birge, H.E.; Davidson, E.A.; Eliasson, P.E.; Evans, S.E.; Frey, S.D.; Giardina, C.P.; Hopkins, F.M.; et al. Temperature and soil organic matter decomposition rates-synthesis of current knowledge and a way forward. Glob. Chang. Biol. 2011, 17, 3392–3404. [Google Scholar] [CrossRef]
- McMurtrie, R.E.; Iversen, C.M.; Dewar, R.C.; Medlyn, B.E.; Näsholm, T.; Pepper, D.A.; Norby, R.J. Plant root distributions and nitrogen uptake predicted by a hypothesis of optimal root foraging: Optimal Root Foraging. Ecol. Evol. 2012, 2, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Whalen, J.K. Managing Soil biota-mediated decomposition and nutrient mineralization in sustainable agroecosystems. Adv. Agric. 2014, 1–13. [Google Scholar] [CrossRef]
- Bridgham, S.D.; Ye, R. Organic matter mineralization and decomposition. In SSSA Book Series; DeLaune, R.D., Reddy, K.R., Richardson, C.J., Megonigal, J.P., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 2015; pp. 385–406. [Google Scholar]
- Rusnak, J. How to Improve Soil Fertility? Agricultural Advisory Center: Karniowice, Poland, 2017. [Google Scholar]
- Kumar, K.; Goh, K.M. Crop residues and management practices: Effects on soil quality, Soil nitrogen dynamics, crop yield, and nitrogen recovery. Adv. Agron. 1999, 68, 197–319. [Google Scholar]
- Singh, B.; Rengel, Z. The role of crop residues in improving soil fertility. In Nutrient Cycling in Terrestrial Ecosystems; Marschner, P., Rengel, Z., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2007; Volume 10, pp. 183–214. [Google Scholar]
- Henriksen, T.; Breland, T. Carbon mineralization, fungal and bacterial growth, and enzyme activities as affected by contact between crop residues and soil. Biol. Fertil. Soils 2002, 35, 41–48. [Google Scholar] [CrossRef]
- Chen, B.; Liu, E.; Tian, Q.; Yan, C.; Zhang, Y. Soil nitrogen dynamics and crop residues. A review. Agron. Sustain. Dev. 2014, 34, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Yadvinder-Singh, B.S.; Timsina, J. Research Publications Repository-Crop residue management for nutrient cycling and improving soil productivity in rice-based cropping systems in the tropics. Adv. Agron. 2005, 85, 269–407. [Google Scholar]
- Sapek, B. Nitrogen and phosphorus release from soil organic matter. Water Environ. Rural Areas 2010, 10, 229–256. [Google Scholar]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Jamir, E.; Kangabam, R.D.; Borah, K.; Tamuly, A.; Deka Boruah, H.P.; Silla, Y. Role of Soil microbiome and enzyme activities in plant growth nutrition and ecological restoration of soil health. In Microbes and Enzymes in Soil Health and Bioremediation; Kumar, A., Sharma, S., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2019; pp. 99–132. [Google Scholar]
- Nieder, R.; Benbi, D.K. Carbon and nitrogen transformations in soils. In Carbon and Nitrogen in the Terrestrial Environment; Nieder, R., Benbi, D.K., Eds.; Springer Science & Business Media: Dordrecht, The Netherlands, 2008; pp. 137–159. [Google Scholar]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef]
- Arcand, M.M.; Helgason, B.L.; Lemke, R.L. Microbial crop residue decomposition dynamics in organic and conventionally managed soils. Appl. Soil Ecol. 2016, 107, 347–359. [Google Scholar] [CrossRef]
- Gupta, V.V. Biological factors influence N mineralization from soil organic matter and crop residues in Australian cropping systems. Red 2016, 500, 35–55. [Google Scholar]
- Strickland, M.S.; Lauber, C.; Fierer, N.; Bradford, M.A. Testing the functional significance of microbial community composition. Ecology 2009, 90, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevins, C.J.; Nakatsu, C.; Armstrong, S. Characterization of microbial community response to cover crop residue decomposition. Soil Biol. Biochem. 2018, 127, 39–49. [Google Scholar] [CrossRef]
- Kerdraon, L.; Balesdent, M.-H.; Barret, M.; Laval, V.; Suffert, F. Crop residues in wheat-oilseed rape rotation system: A Pivotal, Shifting Platform for Microbial Meetings. Microb. Ecol. 2019, 77, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Ruess, L.; Ferris, H. Decomposition pathways and successional changes. Nematol. Monogr. Perspect. 2004, 2, 547–556. [Google Scholar]
- Bastian, F.; Bouziri, L.; Nicolardot, B.; Ranjard, L. Impact of wheat straw decomposition on successional patterns of soil microbial community structure. Soil Biol. Biochem. 2009, 41, 262–275. [Google Scholar] [CrossRef]
- Galus-Barchan, A.; Chmiel, M.J. The role of microorganisms in acqisition of nutrients by plants. Kosmos 2019, 68, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Koeck, D.E.; Pechtl, A.; Zverlov, V.V.; Schwarz, W.H. Genomics of cellulolytic bacteria. Curr. Opin. Biotechnol. 2014, 29, 171–183. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Zühlke, D.; Becher, D.; Riedel, K.; Baldrian, P. Cellulose and hemicellulose decomposition by forest soil bacteria proceeds by the action of structurally variable enzymatic systems. Sci. Rep. 2016, 6, 25279. [Google Scholar] [CrossRef]
- Pérez, J.; Muñoz-Dorado, J.; de la Rubia, T.; Martínez, J. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: An overview. Int. Microbiol. 2002, 5, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Sharma, V.; Saxena, J.; Chandra, R.; Alam, A.; Prakash, A. Isolation and screening of amylolytic bacteria from soil. Int. J. Sci. Res. Agric. Sci. 2015, 2, 159–165. [Google Scholar] [CrossRef]
- Breza-Boruta, B. Occurrence of pectinolytic microorganisms in soil cultivated under organic and conventional cropping systems. Pol. J. Agron. 2013, 15, 32–37. [Google Scholar]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial pectinolytic enzymes: A review. Process Biochem. 2005, 40, 2931–2944. [Google Scholar] [CrossRef]
- Johnson, C.; Albrecht, G. Nitrogen basics–the nitrogen cycle. Agron. Fact Sheet Ser. Fact Sheet 2005, 2, 1–2. [Google Scholar]
- Albornoz, F. Crop responses to nitrogen overfertilization: A review. Sci. Hortic. 2016, 205, 79–83. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef] [Green Version]
- El-Jaoual, T.; Cox, D. Effects of plant age on nitrogen uptake and distribution by greenhouse plants. J. Plant Nutr. 1998, 21, 1055–1066. [Google Scholar] [CrossRef]
- Jones, D.L.; Healey, J.R.; Willett, V.B.; Farrar, J.F.; Hodge, A. Dissolved organic nitrogen uptake by plants—An important N uptake pathway? Soil Biol. Biochem. 2005, 37, 413–423. [Google Scholar] [CrossRef]
- Wyczółkowski, A.; Dąbek-Szreniawska, M. Enzymes taking part in organic nitrogen mineralization. Acta Agroph. 2005, 3, 37–61. [Google Scholar]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment: Activity, Ecology, and Applications; Burns, R.G., Dick, R.P., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 1–33. [Google Scholar]
- Piotrowska, M.; Kusewicz, D. Soil microflora. In Technical Microbiology; Microorganisms and their environment; Libudzisz, Z., Kowal, K., Eds.; Scientific Publisher PWN: Warszawa, Poland, 2007; pp. 195–200. [Google Scholar]
- Jisha, V.; Smitha, R.B.; Pradeep, S.; Sreedevi, S.; Unni, K.N.; Sajith, S.; Priji, P.; Sarath Josh, M.; Benjamin, S. Versatility of microbial proteases. Adv. Enzym. Res. 2013, 1, 39–51. [Google Scholar] [CrossRef]
- Dąbek-Szreniawska, M.; Zimon, A.; Wyczółkowski, A.I. Soil enzymes activity of amonification process in the soil with the addition of organic nitrogen. Acta Agroph. 2006, 8, 23–33. [Google Scholar]
- Strock, J.S. Ammonification. In Encyclopedia of Ecology, Five-Volume Set; Joegensen, S.E., Fath, B.D., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2008; pp. 162–165. [Google Scholar]
- Romillac, N. Ammonification. In Encyclopedia of Ecology; Fath, B.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 256–263. [Google Scholar]
- Deacon, L.J.; Pryce-Miller, E.J.; Frankland, J.C.; Bainbridge, B.W.; Moore, P.D.; Robinson, C.H. Diversity and function of decomposer fungi from a grassland soil. Soil Biol. 2006, 38, 7–20. [Google Scholar] [CrossRef]
- Deubel, A.; Merbach, W. Influence of Microorganisms on phosphorus bioavailability in soils. In Microorganisms in Soils: Roles in Genesis and Functions; Varma, A., Buscot, F., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2005; Volume 3, pp. 177–191. [Google Scholar]
- Potarzycki, J. Phosphorus in soil. J. Elem. 2003, 8, 19–32. [Google Scholar]
- Kennelly, P.J. Protein phosphatases−a phylogenetic perspective. Chem. Rev. 2001, 101, 2291–2312. [Google Scholar] [CrossRef]
- Quiquampoix, H.; Mousain, D. Enzymatic hydrolysis of organic phosphorus. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D.S., Eds.; CABI Pub: Wallingford, UK; Cambridge, MA, USA, 2005; pp. 89–112. [Google Scholar]
- Martínez-Espinosa, R.M.; Cole, J.A.; Richardson, D.J.; Watmough, N.J. Enzymology and ecology of the nitrogen cycle. Biochem. Soc. Trans. 2011, 39, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of phosphatase enzymes in soil. In Phosphorus in Action; Bünemann, E., Oberson, A., Frossard, E., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 215–243. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soils. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Bielińska, E. Methods of determination of photosphatase activity. Acta Agroph. 2005, 3, 63–74. [Google Scholar]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Harasim, A. Straw Management; Publisher IUNG-PIB: Puławy, Poland, 2011; p. 77. [Google Scholar]
- Smernik, R.J.; Baldock, J.A. Solid-state 15N NMR analysis of highly 15N-enriched plant materials. Plant Soil 2005, 275, 271–283. [Google Scholar] [CrossRef]
- Zhao, F.J.; McGrath, S.P.; Blake-Kalff, M.M.A.; Link, A.; Tucker, M. Crop Responses to Sulphur Fertilisation in Europe; The International Fertiliser Society: York, UK, 2002; pp. 1–28. [Google Scholar]
- Niknahad-Gharmakher, H.; Piutti, S.; Machet, J.-M.; Benizri, E.; Recous, S. Mineralization-immobilization of sulphur in a soil during decomposition of plant residues of varied chemical composition and S content. Plant Soil 2012, 360, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, J. Gross sulphur mineralisation–immobilisation turnover in soil amended with plant residues. Soil Biol. Biochem. 2005, 37, 2216–2224. [Google Scholar] [CrossRef]
- Nicolardot, B.; Recous, S.; Mary, B. Simulation of C and N mineralisation during crop residue decomposition: A simple dynamic model based on the C:N ratio of the residues. Plant Soil 2001, 228, 83–103. [Google Scholar] [CrossRef]
- Hadas, A.; Kautsky, L.; Goek, M.; Kara, E.E. Rates of decomposition of plant residues and available nitrogen in soil, related to residue composition through simulation of carbon and nitrogen turnover. Soil Biol. Biochem. 2004, 36, 255–266. [Google Scholar] [CrossRef]
- Dannehl, T.; Leithold, G.; Brock, C. The effect of C:N ratios on the fate of carbon from straw and green manure in soil: The effect of C:N ratios on the fate of carbon from straw and green manure. Eur. J. Soil Sci. 2017, 68, 988–998. [Google Scholar] [CrossRef]
- Hessen, D.O.; Ågren, G.I.; Anderson, T.R.; Elser, J.J.; De Ruiter, P.C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology 2004, 85, 1179–1192. [Google Scholar] [CrossRef]
- Lynch, M.J.; Mulvaney, M.J.; Hodges, S.C.; Thompson, T.L.; Thomason, W.E. Decomposition, nitrogen and carbon mineralization from food and cover crop residues in the central plateau of Haiti. Springer Plus 2016, 5, 973. [Google Scholar] [CrossRef] [Green Version]
- Thuriès, L. Biochemical composition and mineralization kinetics of organic inputs in a sandy soil. Soil Biol. Biochem. 2002, 34, 239–250. [Google Scholar] [CrossRef]
- Magdoff, F.; Weil, R. Soil organic matter management strategies. In Soil Organic Matter in Sustainable Agriculture; Magdoff, F., Weil, R., Eds.; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Machinet, G.E.; Bertrand, I.; Chabbert, B.; Recous, S. Decomposition in soil and chemical changes of maize roots with genetic variations affecting cell wall quality. Eur. J. Soil Sci. 2009, 60, 176–185. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Decomposition of sun and shade leaves from three deciduous tree species, as affected by their chemical composition. Biol. Fertil. Soils 2003, 37, 137–146. [Google Scholar] [CrossRef]
- Annoussamy, M.; Richard, G.; Recous, S.; Guérif, J. Change in mechanical properties of wheat straw due to decomposition and moisture. Appl. Eng. Agric. 2000, 16, 657–664. [Google Scholar] [CrossRef]
- Magid, J.; Luxhøi, J.; Lyshede, O.B. Decomposition of plant residues at low temperatures separates turnover of nitrogen and energy rich tissue components in time. Plant Soil 2004, 258, 351–365. [Google Scholar] [CrossRef]
- Abiven, S.; Recous, S.; Reyes, V.; Oliver, R. Mineralisation of C and N from root, stem and leaf residues in soil and role of their biochemical quality. Biol. Fertil. Soils 2005, 42, 119–128. [Google Scholar] [CrossRef]
- Prieto, I.; Stokes, A.; Roumet, C. Root functional parameters predict fine root decomposability at the community level. J. Ecol. 2016, 104, 725–733. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W. The decomposition of fine and coarse roots: Their global patterns and controlling factors. Sci. Rep. 2015, 5, 9940. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef]
- Johnson, J.M.-F.; Barbour, N.W.; Weyers, S.L. Chemical composition of crop biomass impacts its decomposition. Soil Sci. Soc. Am. J. 2007, 71, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Kriaučiūnienė, Z.; Velička, R.; Raudonius, S. The influence of crop residues type on their decomposition rate in the soil: A litterbag study. Zemdirb. Agric. 2012, 99, 227–236. [Google Scholar]
- Cabrera, M.L.; Kissel, D.E.; Vigil, M.F. Nitrogen mineralization from organic residues: Research opportunities. J Environ. Qual. 2005, 34, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Paśmionka, I. Microbiological transformations of soil nitrogen. Kosmos 2017, 66, 185–192. [Google Scholar]
- Sapek, B.; Kalinowska, D. Mineralization of soil organic nitrogen compounds in the light of long-term grassland experiments in IMUZ. Water Environ. Rural Areas 2004, 4, 183–200. [Google Scholar]
- Pangnakorn, U.; George, D.L.; Tullberg, J.N.; Gupta, M.L. Effect of tillage and traffic on earthworm populations in a vertosol in south-east Queensland. In Proceedings of the Soil Management for Sustainability, Brisbane, Australia, 13–18 July 2003; International Soil Tillage Research Organisation: Queensland, Australia, 2003. [Google Scholar]
- Hu, R.; Wang, X.; Pan, Y.; Zhang, Y.; Zhang, H. The response mechanisms of soil N mineralization under biological soil crusts to temperature and moisture in temperate desert regions. Eur. J. Soil Biol. 2014, 62, 66–73. [Google Scholar] [CrossRef]
- Chow, A.T.; Tanji, K.K.; Gao, S.; Dahlgren, R.A. Temperature, water content and wet–dry cycle effects on DOC production and carbon mineralization in agricultural peat soils. Soil Biol. Biochem. 2006, 38, 477–488. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology and Biochemistry, 4th ed.; Academic Press: Waltham, MA, USA, 2014. [Google Scholar]
- Paul, K.I.; Polglase, P.J.; O’Connell, A.M.; Carlyle, J.C.; Smethurst, P.J.; Khanna, P.K. Defining the relation between soil water content and net nitrogen mineralization. Eur. J. Soil Sci. 2003, 54, 39–48. [Google Scholar] [CrossRef]
- Devêvre, O.C.; Horwáth, W.R. Decomposition of rice straw and microbial carbon use efficiency under different soil temperatures and moistures. Soil Biol. Biochem. 2000, 32, 1773–1785. [Google Scholar] [CrossRef]
- Kruse, J.S.; Kissel, D.E.; Cabrera, M.L. Effects of drying and rewetting on carbon and nitrogen mineralization in soils and incorporated residues. Nutr. Cycl. Agroecosyst. 2004, 69, 247–256. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Klocke, N.L.; Currie, R.S.; Aiken, R.M. Soil water evaporation and crop residues. Trans. ASABE 2009, 52, 103–110. [Google Scholar] [CrossRef]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef]
- Frøseth, R.B.; Bleken, M.A. Effect of low temperature and soil type on the decomposition rate of soil organic carbon and clover leaves, and related priming effect. Soil Biol. Biochem. 2015, 80, 156–166. [Google Scholar] [CrossRef]
- Schmatz, R.; Recous, S.; Aita, C.; Tahir, M.M.; Schu, A.L.; Chaves, B.; Giacomini, S.J. Crop residue quality and soil type influence the priming effect but not the fate of crop residue C. Plant Soil 2017, 414, 229–245. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M.A. Effects of trace elements on b-glucosaminidase activity in soils. Soil Biol. 2002, 34, 1829–1832. [Google Scholar] [CrossRef]
- Kemmitt, S.; Wright, D.; Goulding, K.; Jones, D. pH regulation of carbon and nitrogen dynamics in two agricultural soils. Soil Biol. Biochem. 2006, 38, 898–911. [Google Scholar] [CrossRef]
- Aciego Pietri, J.C.; Brookes, P.C. Nitrogen mineralisation along a pH gradient of a silty loam UK soil. Soil Biol. Biochem. 2008, 40, 797–802. [Google Scholar] [CrossRef]
- Mandal, K.G.; Misra, A.K.; Hati, K.M.; Bandyopadhyay, K.K.; Ghosh, P.K.; Mohanty, M. Rice residue- management options and effects on soil properties and crop productivity. J. Food Agric. Environ. 2004, 2, 224–231. [Google Scholar]
- Kumar, K.; Goh, K.M. Nitrogen Release from crop residues and organic amendments as affected by biochemical composition. Commun. Soil Sci. Plant Anal. 2003, 34, 2441–2460. [Google Scholar] [CrossRef]
- Bergman, I.; Lundberg, P.; Nilsson, M. Microbial carbon mineralisation in an acid surface peat: Effects of environmental factors in laboratory incubations. Soil Biol. Biochem. 1999, 31, 1867–1877. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Müller, C.; Cai, Z. Effects of pH and mineralisation on nitrification in a subtropical acid forest soil. Soil Res. 2018, 56, 275–283. [Google Scholar] [CrossRef]
- Hu, H.-W.; Xu, Z.-H.; He, J.-Z. Chapter Six-Ammonia-oxidizing archaea play a predominant role in acid soil nitrification. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2014; Volume 125, pp. 261–302. [Google Scholar]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Brankatschk, G.; Finkbeiner, M. Crop rotations and crop residues are relevant parameters for agricultural carbon footprints. Agron. Sustain. Dev. 2017, 37, 58. [Google Scholar] [CrossRef] [Green Version]
- Pandiaraj, T.; Selvaraj, S.; Ramu, N. Effects of crop residue management and nitrogen fertilizer on soil nitrogen and carbon content and productivity of wheat (Triticum aestivum l.) in two cropping systems. J. Agric. Sci. Technol. 2015, 17, 249–260. [Google Scholar]
- Surekha, K.; Kumari, A.P.P.; Reddy, M.N.; Satyanarayana, K.; Cruz, S. Crop residue management to sustain soil fertility and irrigated rice yields. Nutr. Cycl. Agroecosyst. 2003, 67, 145–154. [Google Scholar] [CrossRef]
- Shafi, M.; Bakht, J.; Khan, M.A. Effects of crop sequence and crop residues on soil C, soil N and yield of maize. Pak. J. Bot. 2010, 42, 1651–1664. [Google Scholar]
- Arnault, I.; Fleurance, C.; Vey, F.; Fretay, G.D.; Auger, J. Use of Alliaceae residues to control soil-borne pathogens. Ind. Crops Prod. 2013, 49, 265–272. [Google Scholar] [CrossRef]
- Arnault, I.; Huchette, O.; Auger, J. Characterisation of Aroma “Type” in Allium species according to their s-alk(en)yl cysteine sulfoxides and γ-glutamyl dipeptides contents. In Proceedings of the International Symposium on Medicinal and Aromatic Plants, Djerba, Tunisia, 26–28 March 2009; ISHS, International Society for Horticultural Science: Hague, The Natherlands, 2010; pp. 171–182. [Google Scholar]
- Wang, D.; Rosen, C.; Kinkel, L.; Cao, A.; Tharayil, N.; Gerik, J. Production of methyl sulfide and dimethyl disulfide from soil-incorporated plant materials and implications for controlling soilborne pathogens. Plant Soil 2009, 324, 185–197. [Google Scholar] [CrossRef]
- Kerdraon, L.; Laval, V.; Suffert, F. Microbiomes and pathogen survival in crop residues, an ecotone between plant and soil. Phytobiomes J. 2019, 3, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Suffert, F.; Sache, I. Relative importance of different types of inoculum to the establishment of Mycosphaerella graminicola in wheat crops in north-west Europe. Plant Pathol. 2011, 60, 878–889. [Google Scholar] [CrossRef]
- Vera, D.I.; Murray, T.D. Occurrence and survival of spothecia of the syespot pathogens oculimacula acuformis and O. yallundae on wheat stubble in the U.S. Pacific Northwest. Plant Dis. 2016, 100, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.; Jessop, R.S.; Sindel, B.M.; Hoult, A. Utilising crop residues to control weeds. In Proceedings of the 12th Australian Weeds Conference, Papers and Proceedings, Weed Management into the 21st Century: Do We Know Where We’re Going? Hobart, Tasmania, Australia, 12–16 September 1999; University of Tasmania: Hobart, Australia, 1999. [Google Scholar]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides, Text with EEA Relevance. Official Journal of the European Union 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex%3A32009L0128 (accessed on 1 September 2020).
- Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Official Journal of the European Union 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32009R1107 (accessed on 1 September 2020).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzyb, A.; Wolna-Maruwka, A.; Niewiadomska, A. Environmental Factors Affecting the Mineralization of Crop Residues. Agronomy 2020, 10, 1951. https://doi.org/10.3390/agronomy10121951
Grzyb A, Wolna-Maruwka A, Niewiadomska A. Environmental Factors Affecting the Mineralization of Crop Residues. Agronomy. 2020; 10(12):1951. https://doi.org/10.3390/agronomy10121951
Chicago/Turabian StyleGrzyb, Aleksandra, Agnieszka Wolna-Maruwka, and Alicja Niewiadomska. 2020. "Environmental Factors Affecting the Mineralization of Crop Residues" Agronomy 10, no. 12: 1951. https://doi.org/10.3390/agronomy10121951
APA StyleGrzyb, A., Wolna-Maruwka, A., & Niewiadomska, A. (2020). Environmental Factors Affecting the Mineralization of Crop Residues. Agronomy, 10(12), 1951. https://doi.org/10.3390/agronomy10121951