A Conceptual Framework for Winter Dormancy in Deciduous Trees




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
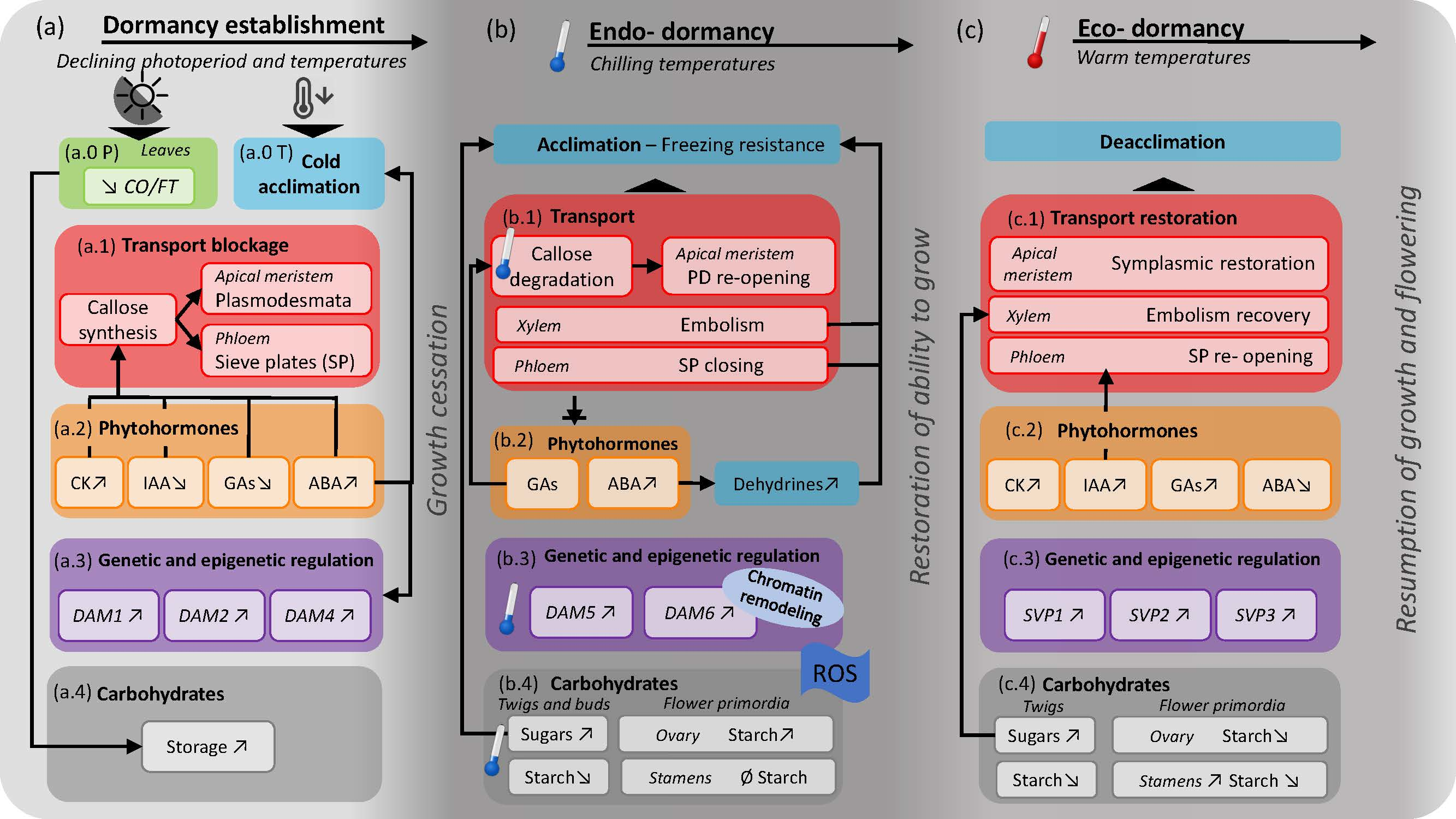
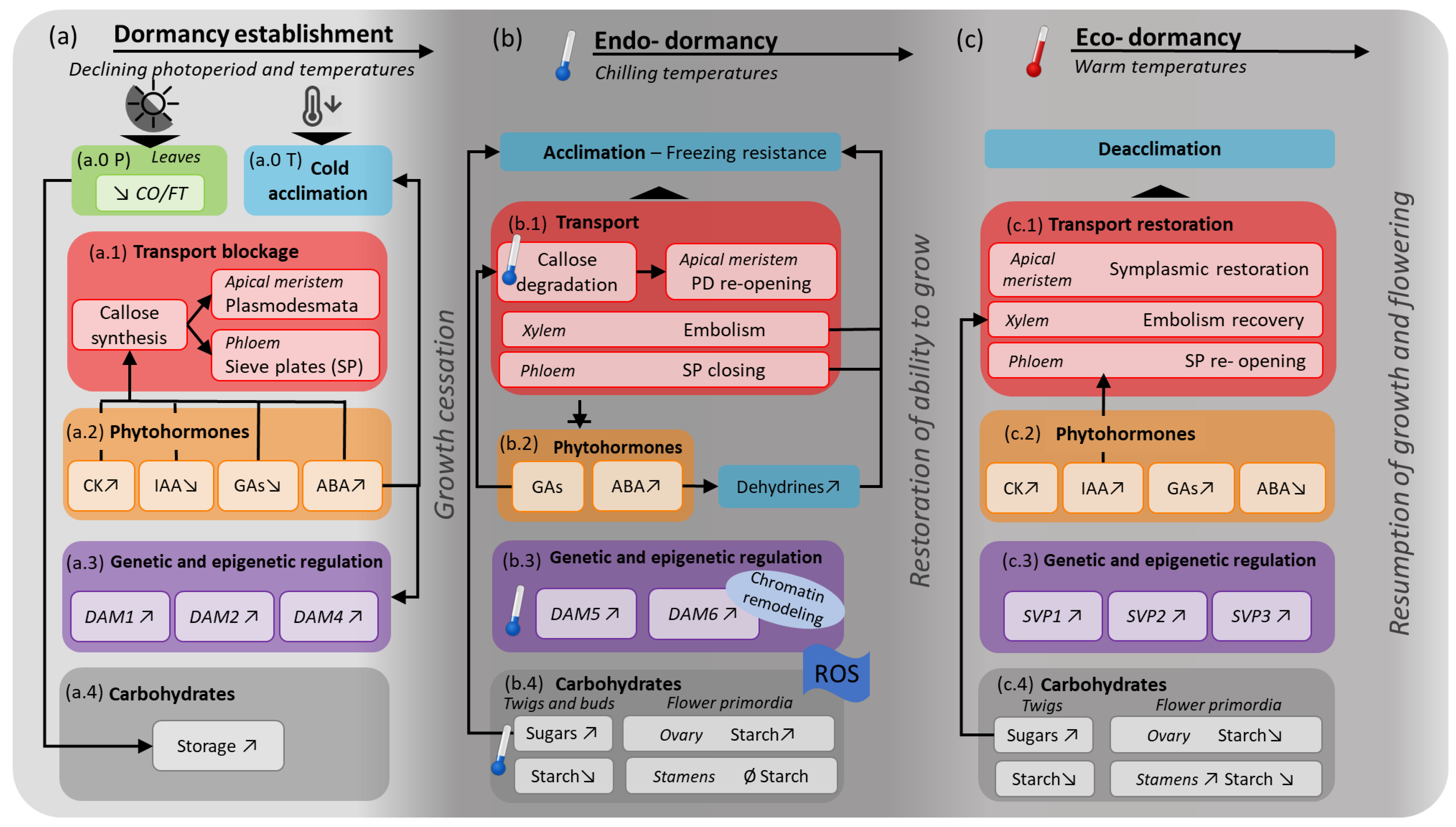
2. A Seasonal Framework of Dormancy
2.1. Dormancy Induction
2.2. Endo-Dormancy
2.3. Eco-Dormancy
3. Physiological Processes that Regulate Dormancy
3.1. Transport
3.2. Phytohormones
3.3. Genetic and Epigenetic Dormancy Regulation
3.4. Carbohydrates
4. Physiological Processes during Dormancy Induction
5. Physiological Processes during Endo-Dormancy
6. Physiological Processes during Eco-Dormancy
7. Conclusions
- –
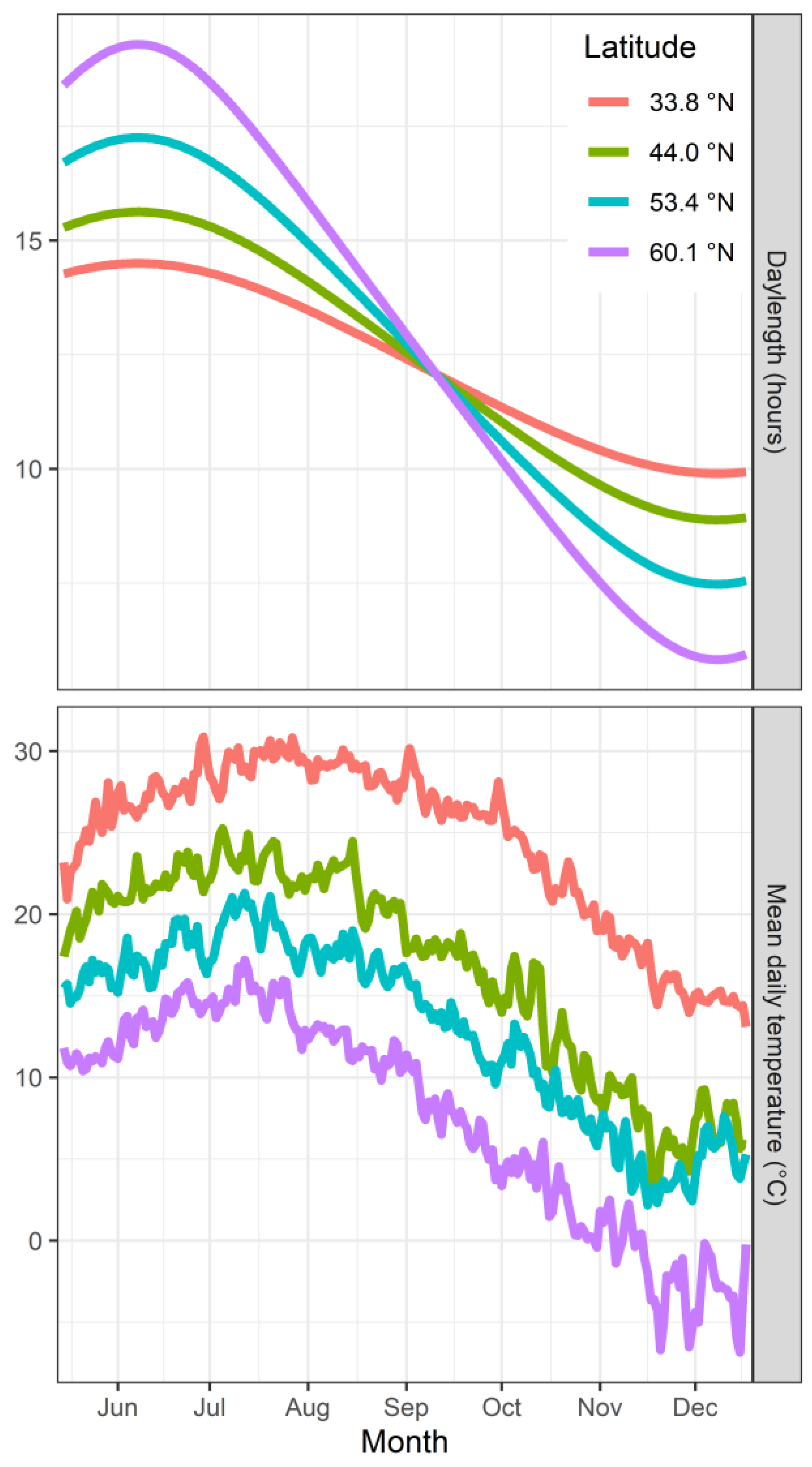
- How dormant trees perceive temperature, translate this perception into a signal and “accumulate” or record it: Chilling temperatures have clearly been established as one of the most important environmental factors that regulate dormancy. However, one of the main knowledge gaps in tree dormancy concerns the mechanisms by which plants perceive temperature. Although this knowledge gap persists, several factors have been related with the accumulation of chilling and dormancy completion.
- –
- How the complex interactions of numerous physiological processes contribute to dormancy dynamics: The significance of each factor appears to vary according to species and location, and there is similar variation in plants’ responsiveness to environmental signals (i.e. temperature and photoperiod). Photoperiod and temperature appear to contribute to dormancy establishment in species-specific ways. The most effective chilling temperatures that contribute to dormancy completion may also differ between species and cultivars.
- –
- Dormancy model improvement: In general, the dormancy models that are currently available are composed of combinations of chill and heat sub-models, both of which are only based on temperature and phenological observations. A modeling approach based on the whole process, rather than on just a single environmental parameter (i.e., temperature), could offer not only a tool to predict flowering dates but also a reliable framework of dormancy progression.
Author Contributions
Funding
Conflicts of Interest
References
- Luedeling, E. Climate change impacts on winter chill for temperate fruit and nut production: A review. Sci. Hortic. 2012, 144, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Luedeling, E.; Zhang, M.; Girvetz, E.H. Climatic changes lead to declining winter chill for fruit and nut trees in California during 1950–2099. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Luedeling, E.; Girvetz, E.H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Knight, T.A. Account of some experiments on the ascent of the sap in trees. Philos. Trans. R. Soc. Lond. 1801, 91, 333–353. [Google Scholar]
- Doorenbos, J. Review of the literature on dormancy in buds of woody plants. Meded. Landbouwhogesch. Wagening. Ned. 1953, 53, 1–24. [Google Scholar]
- Vegis, A. Dormancy in higher plants. Annu. Rev. Plant Physiol. 1964, 15, 185–224. [Google Scholar] [CrossRef]
- Perry, T.O. Dormancy of trees in winter. Science 1971, 171, 29–36. [Google Scholar] [CrossRef]
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Coville, F.V. The influence of cold in stimulating the growth of plants. Proc. Natl. Acad. Sci. USA 1920, 6, 434–735. [Google Scholar] [CrossRef] [Green Version]
- Chuine, I.; Régnière, J. Process-Based Models of Phenology for Plants and Animals. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 159–182. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for “Redhaven” and “Elberta” peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of at two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants—Computer-simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The dynamic model for rest completion in peach buds. Acta Hortic. 1990, 165–174. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy—Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar]
- Considine, M.J.; Considine, J.A. On the language and physiology of dormancy and quiescence in plants. J. Exp. Bot. 2016, 67, 3189–3203. [Google Scholar] [CrossRef] [Green Version]
- Bielenberg, D.G.; Wang, Y.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. A deletion affecting several gene candidates is present in the Evergrowing peach mutant. J. Hered. 2004, 95, 436–444. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Wang, Y.E.; Li, Z.; Zhebentyayeva, T.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genomes 2008, 4, 495–507. [Google Scholar] [CrossRef]
- Zhuang, W.; Gao, Z.; Wang, L.; Zhong, W.; Ni, Z.; Zhang, Z. Comparative proteomic and transcriptomic approaches to address the active role of GA(4) in Japanese apricot flower bud dormancy release. J. Exp. Bot. 2013, 64, 4953–4966. [Google Scholar] [CrossRef]
- Gonzalez, L.M.G.; El Kayal, W.; Ju, C.; Allen, C.C.G.; King-Jones, S.; Cooke, J.E.K.; Gonzalez, L.M.G.; El Kayal, W.; Ju, C.J.-T.; Allen, C.C.G.; et al. Integrated transcriptomic and proteomic profiling of white spruce stems during the transition from active growth to dormancy. Plant Cell Environ. 2012, 35, 682–701. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhuo, X.; Zhao, K.; Zheng, T.; Han, Y.; Yuan, C.; Zhang, Q. Transcriptome profiles reveal the crucial roles of hormone and sugar in the bud dormancy of Prunus mume. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitani, C.; Saito, T.; Ubi, B.E.; Shimizu, T.; Itai, A.; Saito, T.; Yamamoto, T.; Moriguchi, T. Transcriptome analysis of Pyrus pyrifolia leaf buds during transition from endodormancy to ecodormancy. Sci. Hortic. 2012, 147, 49–55. [Google Scholar] [CrossRef]
- Ríos, G.; Leida, C.; Conejero, A.; Badenes, M.L. Epigenetic regulation of bud dormancy events in perennial plants. Front. Plant Sci. 2014, 5, 247. [Google Scholar] [PubMed]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud dormancy in perennial fruit tree species: A pivotal role for oxidative cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- Singh, R.K.; Svystun, T.; AlDahmash, B.; Jönsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef] [Green Version]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef]
- Lloret, A.; Badenes, M.L.; Ríos, G. Modulation of dormancy and growth responses in reproductive buds of temperate trees. Front. Plant Sci. 2018, 9, 1368. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Paul, L.K.; Rinne, P.L.; van der Schoot, C. Shoot meristems of deciduous woody perennials: Self-organization and morphogenetic transitions. Curr. Opin. Plant Biol. 2014, 17, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Falavigna, V.D.S.; Guitton, B.; Costes, E.; Andrés, F. I want to (bud) break free: The potential role of DAM and SVP-Like genes in regulating dormancy cycle in temperate fruit trees. Front. Plant Sci. 2019, 9, 1990. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Spencer, J. Fourier series representation of the position of the Sun. Search 1971, 2, 172. [Google Scholar]
- Almorox, J.; Hontoria, C.; Benito, M. Statistical validation of daylength definitions for estimation of global solar radiation in Toledo, Spain. Energy Convers. Manag. 2005, 46, 1465–1471. [Google Scholar] [CrossRef]
- Luedeling, E. chillR: Statistical Methods for Phenology Analysis in Temperate Fruit Trees; R Package Version 0.70.21; 2019. [Google Scholar]
- Samish, R.M. Dormancy in woody plants. Annu. Rev. Plant Physiol. 1954, 5, 183–204. [Google Scholar] [CrossRef]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Diggle, P.K.; Mulder, C.P.H. Diverse developmental responses to warming temperatures underlie changes in flowering phenologies. Integr. Comp. Biol. 2019, 59, 559–570. [Google Scholar] [CrossRef]
- Rohde, A.; Storme, V.; Jorge, V.; Gaudet, M.; Vitacolonna, N.; Fabbrini, F.; Ruttink, T.; Zaina, G.; Marron, N.; Dillen, S.; et al. Bud set in poplar-genetic dissection of a complex trait in natural and hybrid populations. New Phytol. 2011, 189, 106–121. [Google Scholar] [CrossRef]
- Rinne, P.L.H.; Paul, L.K.; van der Schoot, C. Decoupling photo- and thermoperiod by projected climate change perturbs bud development, dormancy establishment and vernalization in the model tree Populus. BMC Plant Biol. 2018, 18, 220. [Google Scholar] [CrossRef]
- Heide, O.M.; Prestrud, A.K. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heide, O.M. Interaction of photoperiod and temperature in the control of growth and dormancy of Prunus species. Sci. Hortic. 2008, 115, 309–314. [Google Scholar] [CrossRef]
- Wilkie, J.D.; Sedgley, M.; Olesen, T. Regulation of floral initiation in horticultural trees. J. Exp. Bot. 2008, 59, 3215–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanninen, H.; Tanino, K. Tree seasonality in a warming climate. Trends Plant Sci. 2011, 16, 412–416. [Google Scholar] [CrossRef] [PubMed]
- George, A.; Erez, A. Stone fruit species under warm subtropical and tropical climates. In Temperate Fruit Crops in Warm Climates; Springer: Dordrecht, The Netherlands, 2000; pp. 231–265. [Google Scholar]
- Kurokura, T.; Mimida, N.; Battey, N.H.; Hytönen, T. The regulation of seasonal flowering in the Rosaceae. J. Exp. Bot. 2013, 64, 4131–4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julian, C.; Herrero, M.; Rodrigo, J. Flower bud differentiation and development in fruiting and non-fruiting shoots in relation to fruit set in apricot (Prunus armeniaca L.). Trees 2010, 24, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Julian, C.; Rodrigo, J.; Herrero, M. Stamen development and winter dormancy in apricot (Prunus armeniaca). Ann. Bot. 2011, 108, 617–625. [Google Scholar] [CrossRef]
- Felker, F.C.; Robitaille, H.A.; Hess, F.D. Morphological and ultrastructural development and starch accumulation during chilling of sour cherry flower buds. Am. J. Bot. 1983, 70, 376–386. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J.; Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees Struct. Funct. 2018, 32, 1583–1594. [Google Scholar]
- Foster, T.; Johnston, R.; Seleznyova, A. A morphological and quantitative characterization of early floral development in apple (Malus x domestica Borkh.). Ann. Bot. 2003, 92, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Ruelland, E.; Vaultier, M.N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. In Advances in Botanical Research; Kader, J.-C., Delseny, M., Eds.; Elesvier Ltd.: Amsterdam, The Netherlands, 2009; Volume 49, pp. 35–150. ISBN 978-0-12-374735-8. [Google Scholar]
- Basler, D.; Körner, C. Photoperiod and temperature responses of bud swelling and bud burst in four temperate forest tree species. Tree Physiol. 2014, 34, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Basler, D.; Körner, C. Photoperiod sensitivity of bud burst in 14 temperate forest tree species. Agric. For. Meteorol. 2012, 165, 73–81. [Google Scholar] [CrossRef]
- Zohner, C.; Renner, S. Perception of photoperiod in individual buds of mature trees regulates leaf-out. New Phytol. 2015, 1023–1030. [Google Scholar] [CrossRef] [Green Version]
- Caffarra, A.; Donnelly, A.; Chuine, I.; Jones, M. Modelling the timing of Betula pubescens budburst. I. Temperature and photoperiod: A conceptual model. Clim. Res. 2011, 46, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Schaber, J.; Badeck, F.-W. Physiology-based phenology models for forest tree species in Germany. Int. J. Biometeorol. 2003, 47, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Lundell, R.; Hänninen, H.; Saarinen, T.; Åström, H.; Zhang, R. Beyond rest and quiescence (endodormancy and ecodormancy): A novel model for quantifying plant–environment interaction in bud dormancy release. Plant Cell Environ. 2020, 43, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower development in sweet cherry framed in the BBCH scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Charrier, G.; Bonhomme, M.; Lacointe, A.; Améglio, T. Are budburst dates, dormancy and cold acclimation in walnut trees (Juglans regia L.) under mainly genotypic or environmental control? Int. J. Biometeorol. 2011, 55, 763–774. [Google Scholar] [CrossRef]
- Anderson, J.L.; Richardson, E.A.; Kesner, C.D. Validation of Chill Unit and flower bud phenology models for “Montmorency” sour cherry. Acta Hortic. 1986, 184, 71–78. [Google Scholar] [CrossRef]
- Ashcroft, G.L.; Richardson, E.A.; Seeley, S.D. A statistical method of determinig Chill Unit and Growing Degree Hour requirements for deciduous fruit trees. HortScience 1977, 12, 347–348. [Google Scholar]
- Chuine, I.; Bonhomme, M.; Legave, J.M.; de Cortazar-Atauri, I.G.; Charrier, G.; Lacointe, A.; Ameglio, T. Can phenological models predict tree phenology accurately in the future? The unrevealed hurdle of endodormancy break. Glob. Chang. Biol. 2016, 22, 3444–3460. [Google Scholar] [CrossRef]
- Brown, D.S.; Kotob, F.A. Growth of flower buds of apricot, peach, and pear during the rest period. Proc. Am. Soc. Hortic. Sci. 1957, 69, 158–164. [Google Scholar]
- Bennett, J.P. Temperature and bud rest period. Calif. Agric. 1949, 3, 9–12. [Google Scholar]
- Pope, K.S.; Da Silva, D.; Brown, P.H.; DeJong, T.M. A biologically based approach to modeling spring phenology in temperate deciduous trees. Agric. For. Meteorol. 2014, 198–199, 15–23. [Google Scholar] [CrossRef]
- Harrington, C.A.; Gould, P.J.; St.Clair, J.B. Modeling the effects of winter environment on dormancy release of Douglas-fir. For. Ecol. Manag. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Loescher, W.H.; Mccamant, T.; Keller, J.D. Carbohydrate reserves, translocation and storage in woody plant roots. HortScience 1990, 25, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B.K. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- De Schepper, V.; De Swaef, T.; Bauweraerts, I.; Steppe, K. Phloem transport: A review of mechanisms and controls. J. Exp. Bot. 2013, 64, 4839–4850. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-A, J.; Sherman, W.B.; Scorza, R.; Wisniewski, M.; Okie, W.R. “Evergreen” peach, its inheritance and dormant behavior. J. Am. Soc. Hortic. Sci. 1994, 119, 789–792. [Google Scholar] [CrossRef]
- Jiménez, S.; Li, Z.; Reighard, G.L.; Bielenberg, D.G. Identification of genes associated with growth cessation and bud dormancy entrance using a dormancy-incapable tree mutant. BMC Plant Biol. 2010, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Tomes, S.; Karunairetnam, S.; Tustin, S.D.; Hellens, R.P.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. SVP-like MADS Box genes control dormancy and budbreak in apple. Front. Plant Sci. 2017, 8, 477. [Google Scholar] [CrossRef]
- Jiménez, S.; Reighard, G.L.; Bielenberg, D.G. Gene expression of DAM5 and DAM6 is suppressed by chilling temperatures and inversely correlated with bud break rate. Plant Mol. Biol. 2010, 73, 157–167. [Google Scholar] [CrossRef]
- Li, Z.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated MADS genes from the EVG locus of peach [Prunus persica (L.) Batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D. Common mechanisms regulate flowering and dormancy. Plant Sci. 2009, 177, 523–531. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, H.; Abbott, A. Dormancy behaviors and underlying regulatory mechanisms: From perspective of pathways to epigenetic regulation. In Advances in Plant Dormancy; Anderson, J.V., Ed.; Springer: Fargo, ND, USA, 2015; pp. 35–47. ISBN 978-3-319-14450-4. [Google Scholar]
- Karlberg, A.; Englund, M.; Petterle, A.; Molnar, G.; Sjödin, A.; Bako, L.; Bhalerao, R.P. Analysis of global changes in gene expression during activity-dormancy cycle in hybrid aspen apex. Plant Biotechnol. 2010, 27, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhlenius, H.; Huang, T.; Charbonnel-Campaa, L.; Brunner, A.M.; Jansson, S.; Strauss, S.H.; Nilsson, O. Forest CO/FT Regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 2006, 312, 1040–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruelland, Z. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S. Acclimation and adaptive responses of woody plants to environmetal stresses. Bot. Rev. 2002, 279–334. [Google Scholar]
- Rinne, P.L.; Kaikuranta, P.M.; van der Schoot, C. The shoot apical meristem restores its symplasmic organization during chilling-induced release from dormancy. Plant J. 2001, 26, 249–264. [Google Scholar] [CrossRef]
- Rinne, P.L.H.; Welling, A.; Vahala, J.; Ripel, L.; Ruonala, R.; Kangasjarvi, J.; van der Schoot, C. Chilling of dormant buds hyperinduces FLOWERING LOCUS T and recruits GA-inducible 1,3-glucanases to reopen signal conduits and release dormancy in Populus. Plant Cell 2011, 23, 130–146. [Google Scholar] [CrossRef] [Green Version]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M.; et al. Photoperiodic control of seasonal growth is mediated by ABA acting on cell-cell communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-W.; Kumar, R.; Iswanto, A.B.B.; Kim, J.-Y. Callose balancing at plasmodesmata. J. Exp. Bot. 2018, 69, 5325–5339. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.L.; Acheampong, A.K.; Shi, Z.W.; Halaly, T.; Kamiya, Y.; Ophir, R.; Galbraith, D.W.; Or, E. Distinct gibberellin functions during and after grapevine bud dormancy release. J. Exp. Bot. 2018, 69, 1635–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, C.; Ramirez-Parra, E.; Castellano, M.M.; del Pozo, J.C. G1 to S transition: More than a cell cycle engine switch. Curr. Opin. Plant Biol. 2002, 5, 480–486. [Google Scholar] [CrossRef]
- Tuan, P.A.; Bai, S.; Saito, T.; Ito, A.; Moriguchi, T. Dormancy-Associated MADS-Box (DAM) and the abscisic acid pathway regulate pear endodormancy through a feedback mechanism. Plant Cell Physiol. 2017, 58, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Wada, M.; Honda, C.; Matsuura, T.; Ikeda, Y.; Hirayama, T.; Osako, Y.; Gao-Takai, M.; Kojima, M.; Sakakibara, H.; et al. Overexpression of Prunus DAM6 inhibits growth, represses bud break competency of dormant buds and delays bud outgrowth in apple plants. PLoS ONE 2019, 14, e0214788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, C.; Li, X.; Gao, D.; Liu, H.; Li, M. Studies on regulations of endogenous ABA and GA3 in sweet cherry flower buds on dormancy. Acta Hortic. Sin. 2004, 31, 149–154. [Google Scholar]
- Aloni, R.; Baum, S.F.; Peterson, C.A. The role of cytokinin in sieve tube regeneration and callose production in wounded Coleus internodes. Plant Physiol. 1990, 93, 982–989. [Google Scholar] [CrossRef] [Green Version]
- Améglio, T.; Bodet, C.; Lacointe, A.; Cochard, H. Winter embolism, mechanisms of xylem hydraulic conductivity recovery and springtime growth patterns in walnut and peach trees. Tree Physiol. 2002, 22, 1211–1220. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J. Winter xylem embolism and spring recovery in Betula cordifolia, Fagus grandifolia, Abies balsamea and Picea rubens. In Water Transport in Plants under Climatic Stress; Borghetti, M., Grace, J., Raschi, A., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 86–98. ISBN 978-0-511-75330-5. [Google Scholar]
- Mimida, N.; Saito, T.; Moriguchi, T.; Suzuki, A.; Komori, S.; Wada, M. Expression of DORMANCY-ASSOCIATED MADS-BOX (DAM)—Like genes in apple. Biol. Plant. 2015, 59, 237–244. [Google Scholar] [CrossRef]
- Wisniewski, M.; Norelli, J.; Artlip, T. Overexpression of a peach CBF gene in apple: A model for understanding the integration of growth, dormancy, and cold hardiness in woody plants. Front. Plant Sci. 2015, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Yamane, H.; Ooka, T.; Jotatsu, H.; Kitamura, Y.; Akagi, T.; Tao, R. Functional and expressional analyses of PmDAM genes associated with endodormancy in Japanese apricot. Plant Physiol. 2011, 157, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, D.P.; Chao, W.S.; Suttle, J.C.; Thimmapuram, J.; Anderson, J.V. Transcriptome analysis identifies novel responses and potential regulatory genes involved in seasonal dormancy transitions of leafy spurge (Euphorbia esula L.). BMC Genom. 2008, 9, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, D.P.; Sung, S.; Kim, D.; Chao, W.; Anderson, J. Characterization, expression and function of DORMANCY ASSOCIATED MADS-BOX genes from leafy spurge. Plant Mol. Biol. 2010, 73, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.-M.; Yvin, J.-C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 317. [Google Scholar] [CrossRef] [Green Version]
- Dietze, M.C.; Sala, A.; Carbone, M.S.; Czimczik, C.I.; Mantooth, J.A.; Richardson, A.D.; Vargas, R. Nonstructural carbon in woody plants. Annu. Rev. Plant Biol. 2014, 65, 667–687. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, E.; Cuneo, I.F.; Luedeling, E.; Alvarado, L.; Farias, D.; Saa, S. Starch and hexoses concentrations as physiological markers in dormancy progression of sweet cherry twigs. Trees 2019, 33, 1187–1201. [Google Scholar] [CrossRef]
- Fadón, E.; Herrero, M.; Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Front. Plant Sci. 2018, 9, 171. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Herrero, M.; Rodrigo, J. Anther and pollen development in sweet cherry (Prunus avium L.) in relation to winter dormancy. Protoplasma 2019, 256, 733–744. [Google Scholar]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Le Bris, M.; Michaux-Ferrière, N.; Jacob, Y.; Poupet, A.; Barthe, P.; Guigonis, J.-M.; Page-Degivry, M.-T.L. Regulation of bud dormancy by manipulation of ABA in isolated buds of Rosa hybrida cultured in vitro. Funct. Plant Biol. 1999, 26, 273. [Google Scholar] [CrossRef]
- Zheng, C.; Halaly, T.; Acheampong, A.K.; Takebayashi, Y.; Jikumaru, Y.; Kamiya, Y.; Or, E. Abscisic acid (ABA) regulates grape bud dormancy, and dormancy release stimuli may act through modification of ABA metabolism. J. Exp. Bot. 2015, 66, 1527–1542. [Google Scholar] [CrossRef] [PubMed]
- Choubane, D.; Rabot, A.; Mortreau, E.; Legourrierec, J.; Péron, T.; Foucher, F.; Ahcène, Y.; Pelleschi-Travier, S.; Leduc, N.; Hamama, L.; et al. Photocontrol of bud burst involves gibberellin biosynthesis in Rosa sp. J. Plant Physiol. 2012, 169, 1271–1280. [Google Scholar] [CrossRef]
- Wen, L.H.; Zhong, W.J.; Huo, X.M.; Zhuang, W.B.; Ni, Z.J.; Gao, Z.H. Expression analysis of ABA- and GA-related genes during four stages of bud dormancy in Japanese apricot (Prunus mume Sieb. et Zucc). J. Hortic. Sci. Biotechnol. 2016, 91, 362–369. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Y.; Xin, D.; Chen, W.; Shao, X.; Wang, Y.; Guo, W. RNA-Seq-based transcriptome analysis of dormant flower buds of Chinese cherry (Prunus pseudocerasus). Gene 2015, 555, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Sriram, A.; Park, J.; Zhebentyayeva, T.; Main, D.; Abbott, A. Genome wide identification of chilling responsive microRNAs in Prunus persica. BMC Genom. 2012, 13, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leida, C.; Romeu, J.F.; García-Brunton, J.; Ríos, G.; Badenes, M.L. Gene expression analysis of chilling requirements for flower bud break in peach. Plant Breed. 2012, 131, 329–334. [Google Scholar] [CrossRef]
- Kaufmann, H.; Blanke, M. Changes in carbohydrate levels and relative water content (RWC) to distinguish dormancy phases in sweet cherry. J. Plant Physiol. 2017, 218, 1–5. [Google Scholar] [CrossRef]
- Chmielewski, F.-M.; Götz, K.P.; Homman, T.; Huschec, G.; Rawel, H.M. Identification of endodormancy release for cherries (Prunus Avium L.) by Abscisic Acid and Sugars. J. Hortic. 2017, 4. [Google Scholar] [CrossRef]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef]
- Perez, F.J.; Vergara, R.; Or, E. On the mechanism of dormancy release in grapevine buds: A comparative study between hydrogen cyanamide and sodium azide. Plant Growth Regul. 2009, 59, 145–152. [Google Scholar] [CrossRef]
- Hedhly, A.; Vogler, H.; Schmid, M.W.; Pazmino, D.; Gagliardini, V.; Santelia, D.; Grossniklaus, U. Starch turnover and metabolism during flower and early embryo development. Plant Physiol. 2016, 172, 2388–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloni, R.; Peterson, C.A. Auxin promotes dormancy callose removal from the phloem of Magnolia kobus and callose accumulation and earlywood vessel differentiation in Quercus Robur. J. Plant Res. 1997, 110, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Cochard, H.; Lemoine, D.; Ameglio, T.; Granier, A. Mechanisms of xylem recovery from winter embolism in Fagus sylvatica. Tree Physiol. 2001, 21, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Matthews, M.A.; Shackel, K.A. The dynamics of embolism repair in xylem: In vivo visualizations using high-resolution computed tomography. Plant Physiol. 2010, 154, 1088–1095. [Google Scholar] [CrossRef] [Green Version]
- Brodersen, C.R.; McElrone, A.J. Maintenance of xylem network transport capacity: A review of embolism repair in vascular plants. Front. Plant Sci. 2013, 4, 108. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Wang, T.; Warren, B.A.W.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. Kiwifruit SVP2 gene prevents premature budbreak during dormancy. J. Exp. Bot. 2017, 68, 1071–1082. [Google Scholar] [CrossRef]
- Wu, R.M.; Walton, E.F.; Richardson, A.C.; Wood, M.; Hellens, R.P.; Varkonyi-Gasic, E. Conservation and divergence of four kiwifruit SVP-like MADS-box genes suggest distinct roles in kiwifruit bud dormancy and flowering. J. Exp. Bot. 2012, 63, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, J.; Herrero, M.; Hormaza, J.I. Pistil traits and flower fate in apricot (Prunus armeniaca). Ann. Appl. Biol. 2009, 154, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo, J.; Hormaza, J.I.; Herrero, M. Ovary starch reserves and flower development in apricot (Prunus armeniaca). Physiol. Plant. 2000, 108, 35–41. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Hormaza, J.I.; Rodrigo, J. Ovary starch reserves and pistil development in avocado (Persea americana). Physiol. Plant. 2010, 140, 395–404. [Google Scholar] [CrossRef]
- Alcaraz, M.L.; Hormaza, J.I.; Rodrigo, J. Pistil starch reserves at anthesis correlate with final flower fate in avocado (Persea americana). PLoS ONE 2013, 8, e78467. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy 2020, 10, 241. https://doi.org/10.3390/agronomy10020241
Fadón E, Fernandez E, Behn H, Luedeling E. A Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy. 2020; 10(2):241. https://doi.org/10.3390/agronomy10020241
Chicago/Turabian StyleFadón, Erica, Eduardo Fernandez, Helen Behn, and Eike Luedeling. 2020. "A Conceptual Framework for Winter Dormancy in Deciduous Trees" Agronomy 10, no. 2: 241. https://doi.org/10.3390/agronomy10020241
APA StyleFadón, E., Fernandez, E., Behn, H., & Luedeling, E. (2020). A Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy, 10(2), 241. https://doi.org/10.3390/agronomy10020241