Wild Blueberry Fruit Drop: A Consequence of Seed Set?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Time Sequence of Fruit Drop (2008 and 2011)
2.2. Seed Set and Fruit Drop (2011)
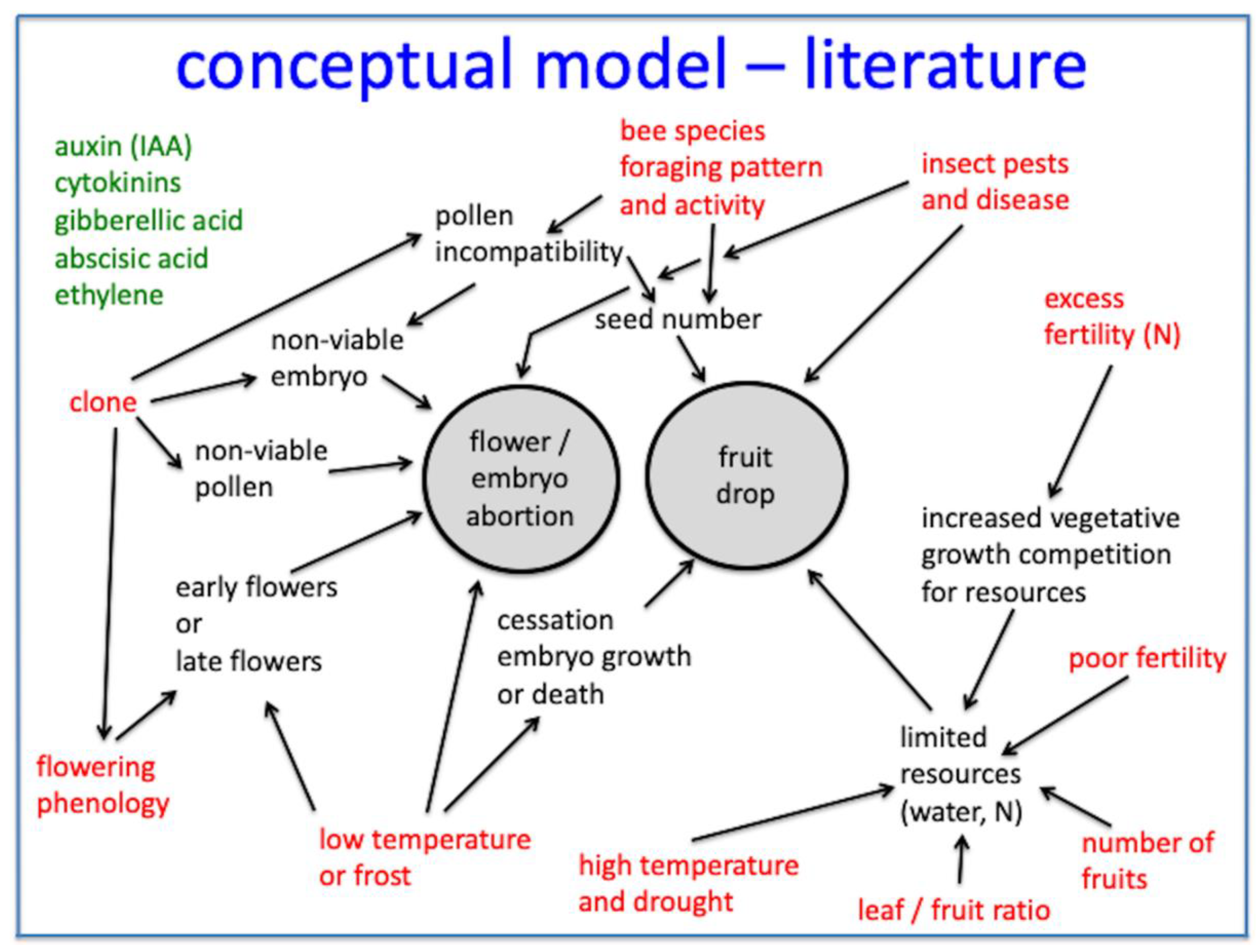
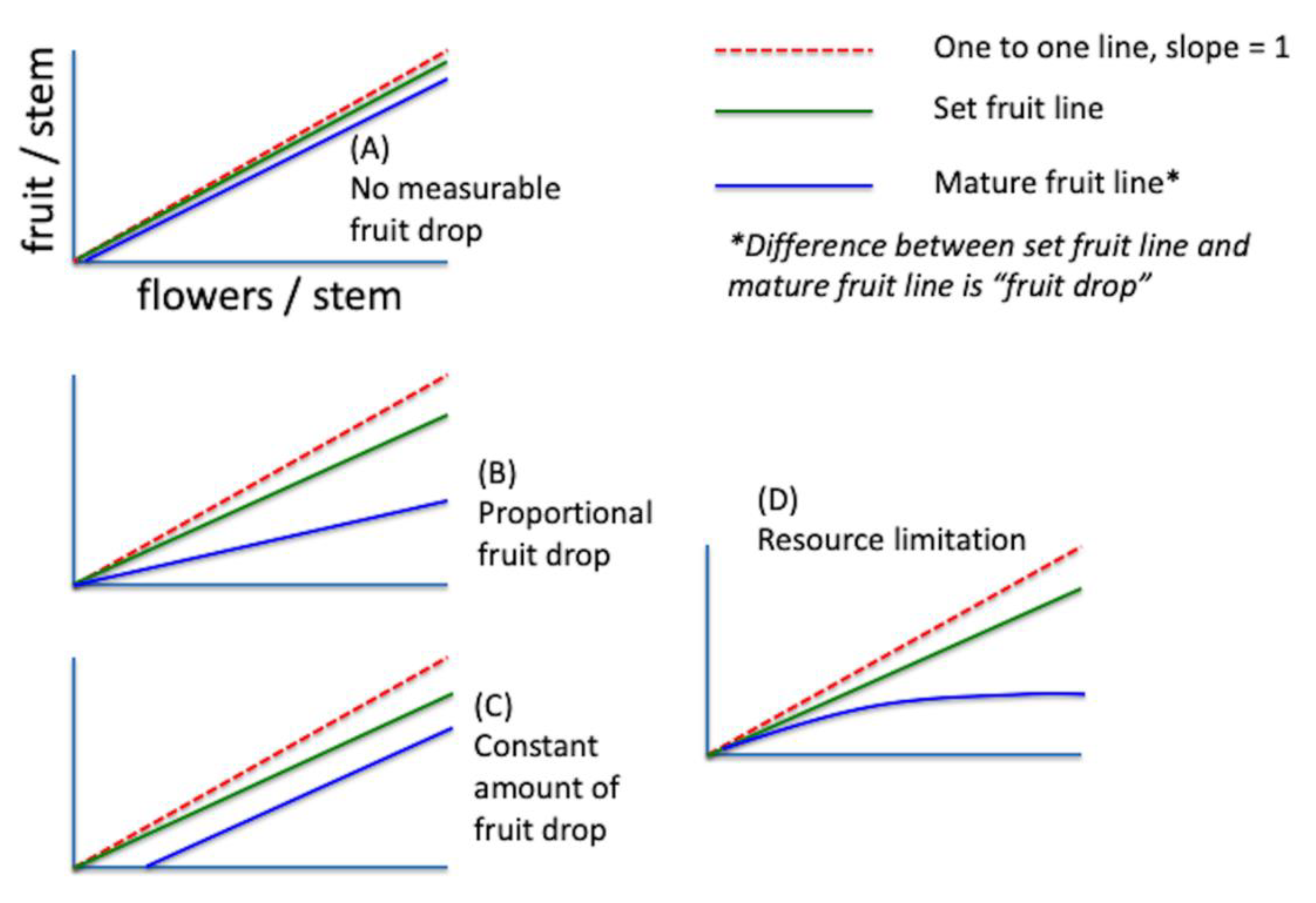
2.3. Fruit Drop Hypotheses (Models—1997)
2.4. Within and between Fields and Years (1996–1999)
3. Results
3.1. Time Sequence of Fruit Drop (2008 and 2011)
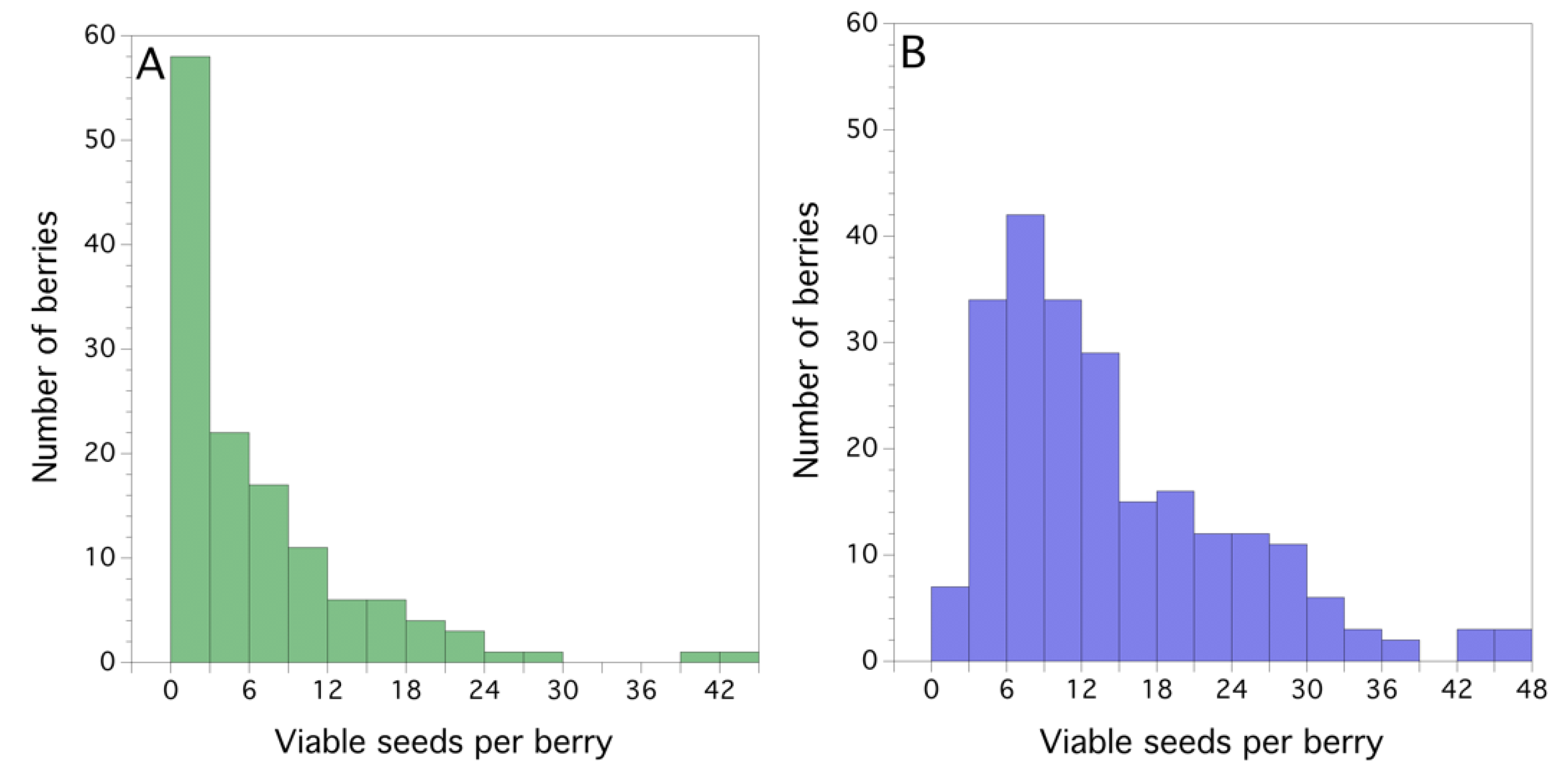
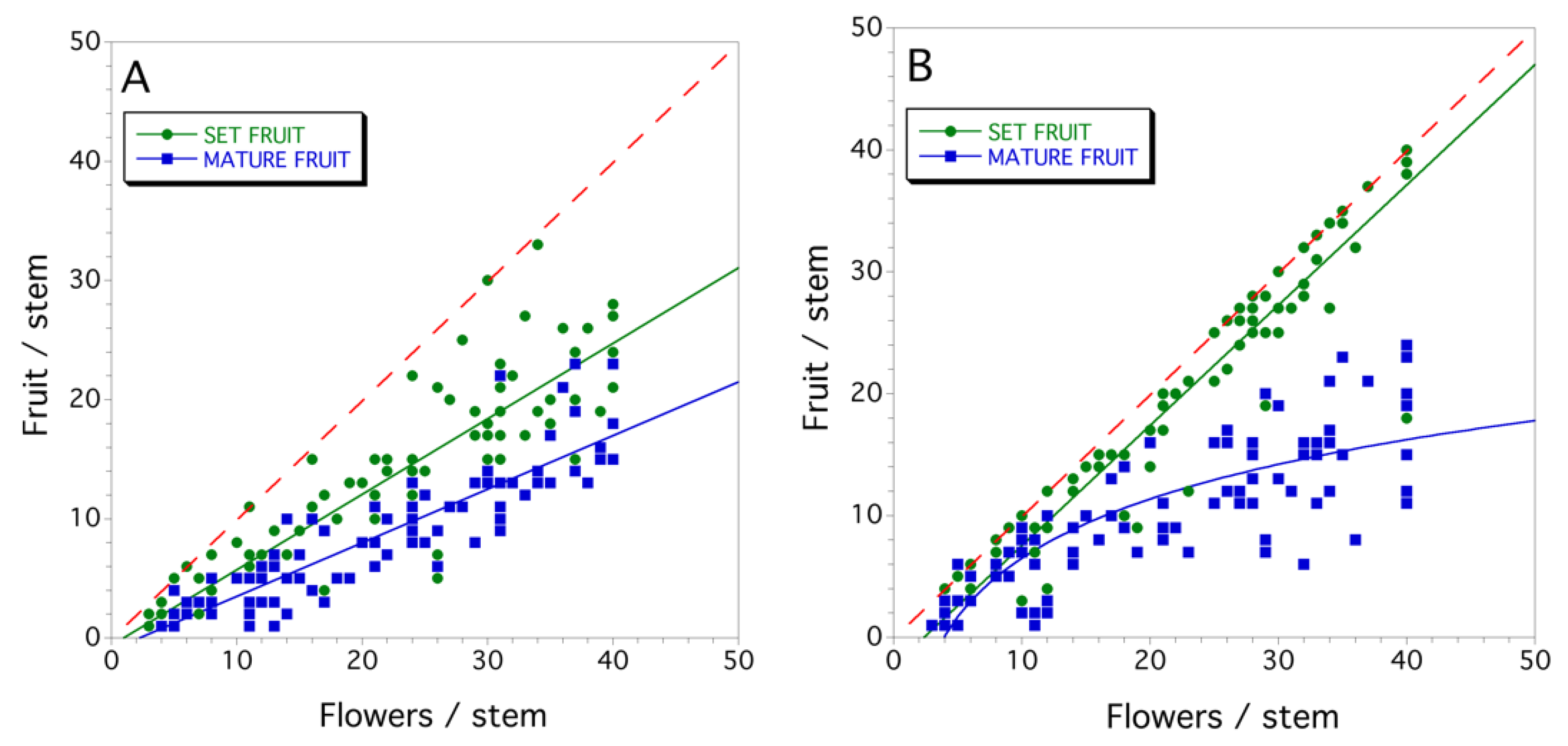
3.2. Seed Set and Fruit Drop (2011)
3.3. Fruit Drop Hypotheses (Models—1997)
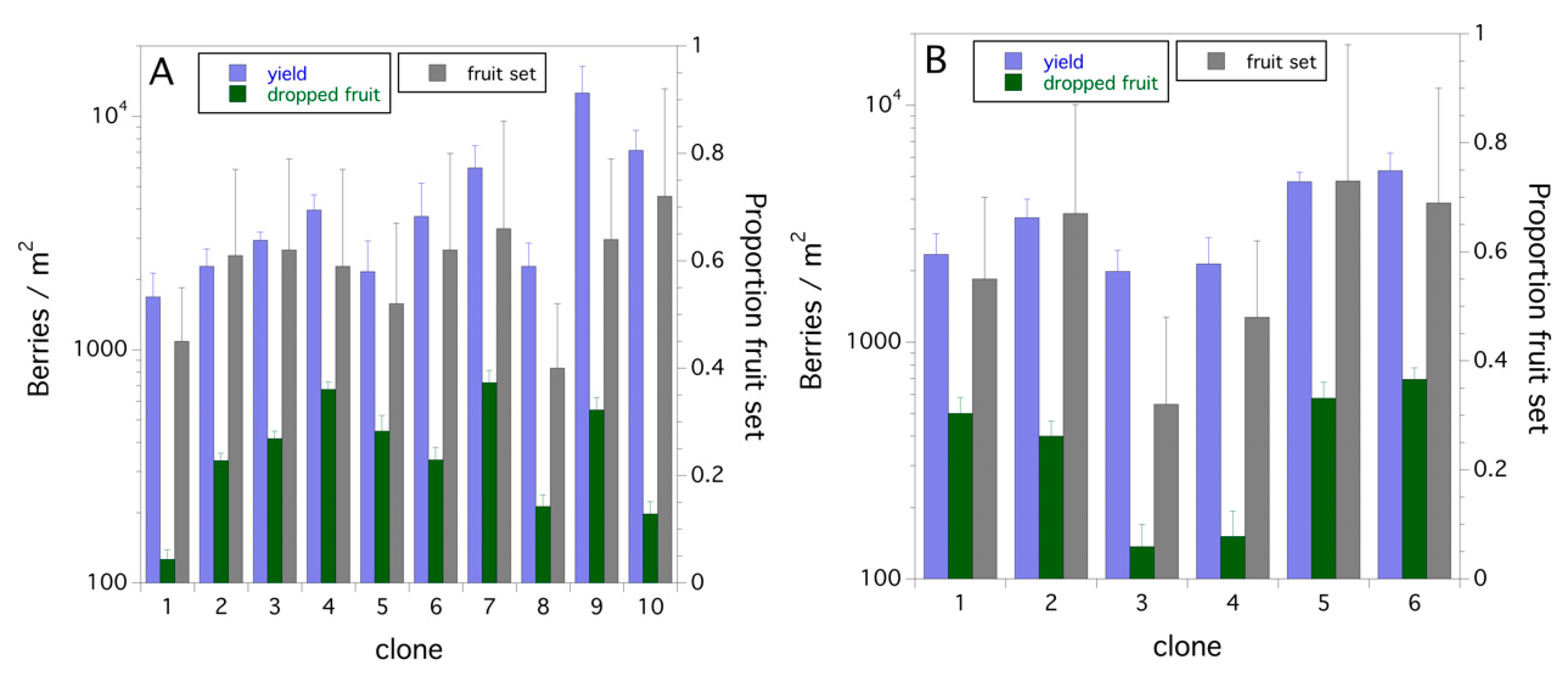
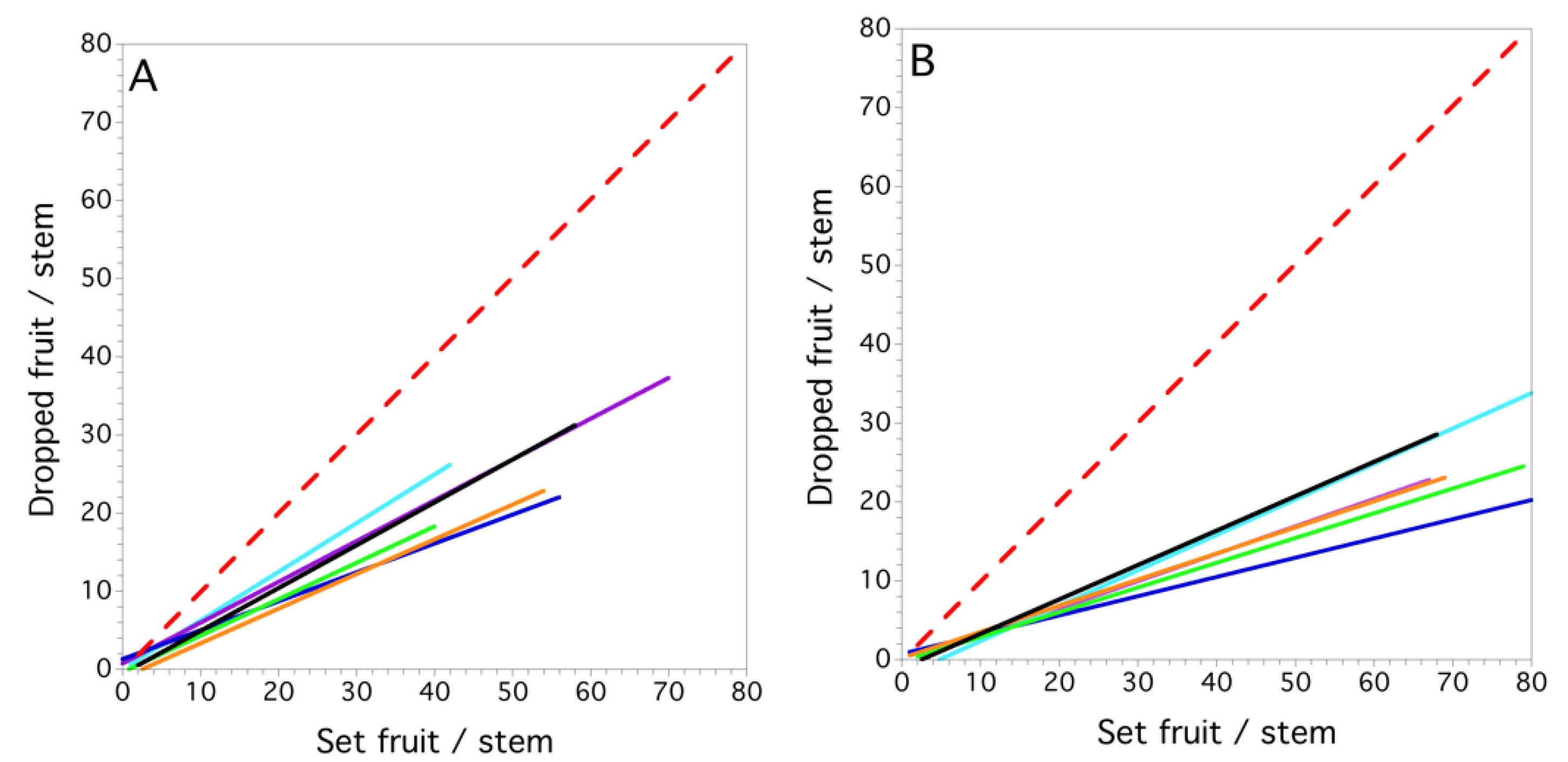
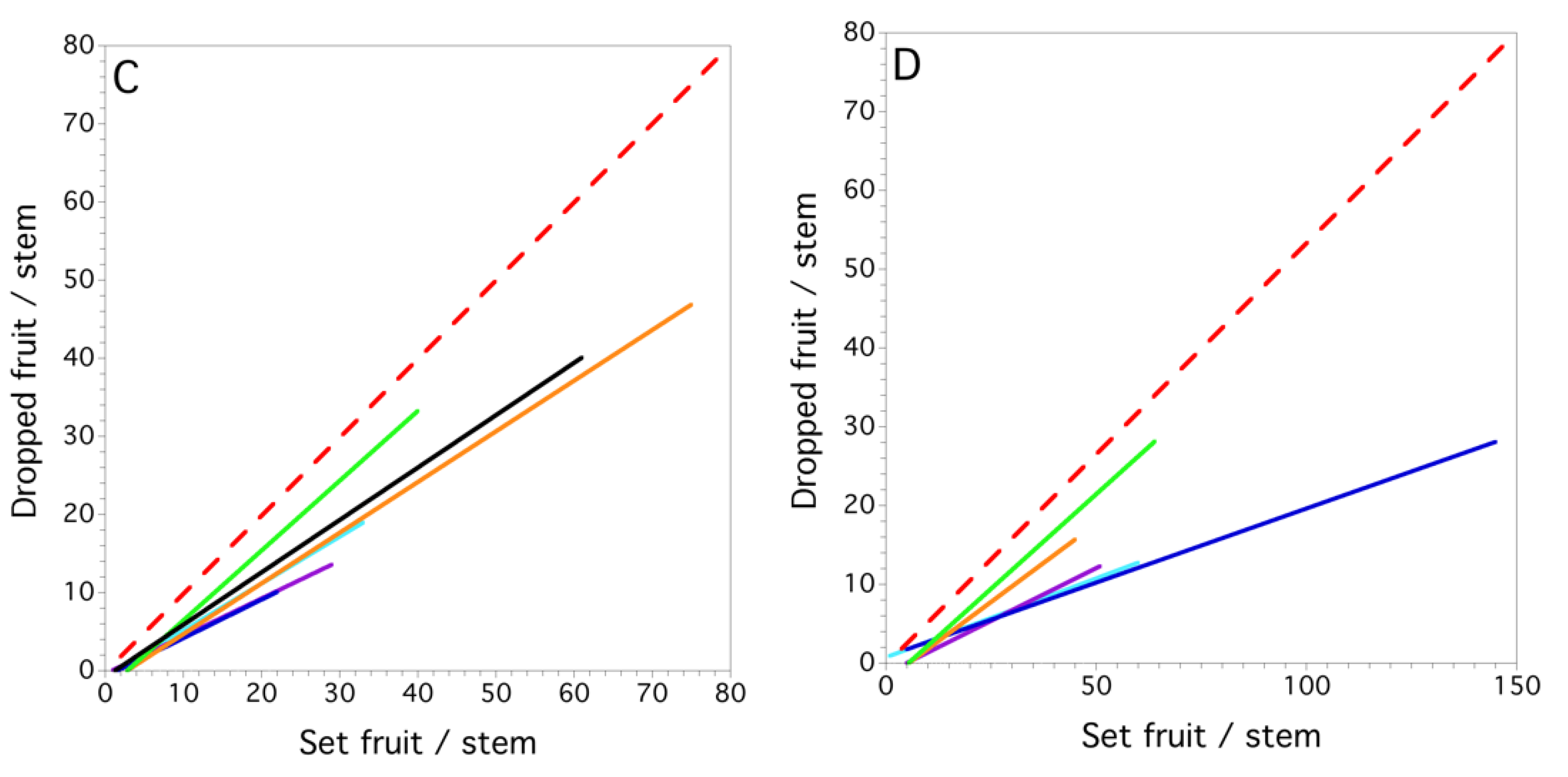
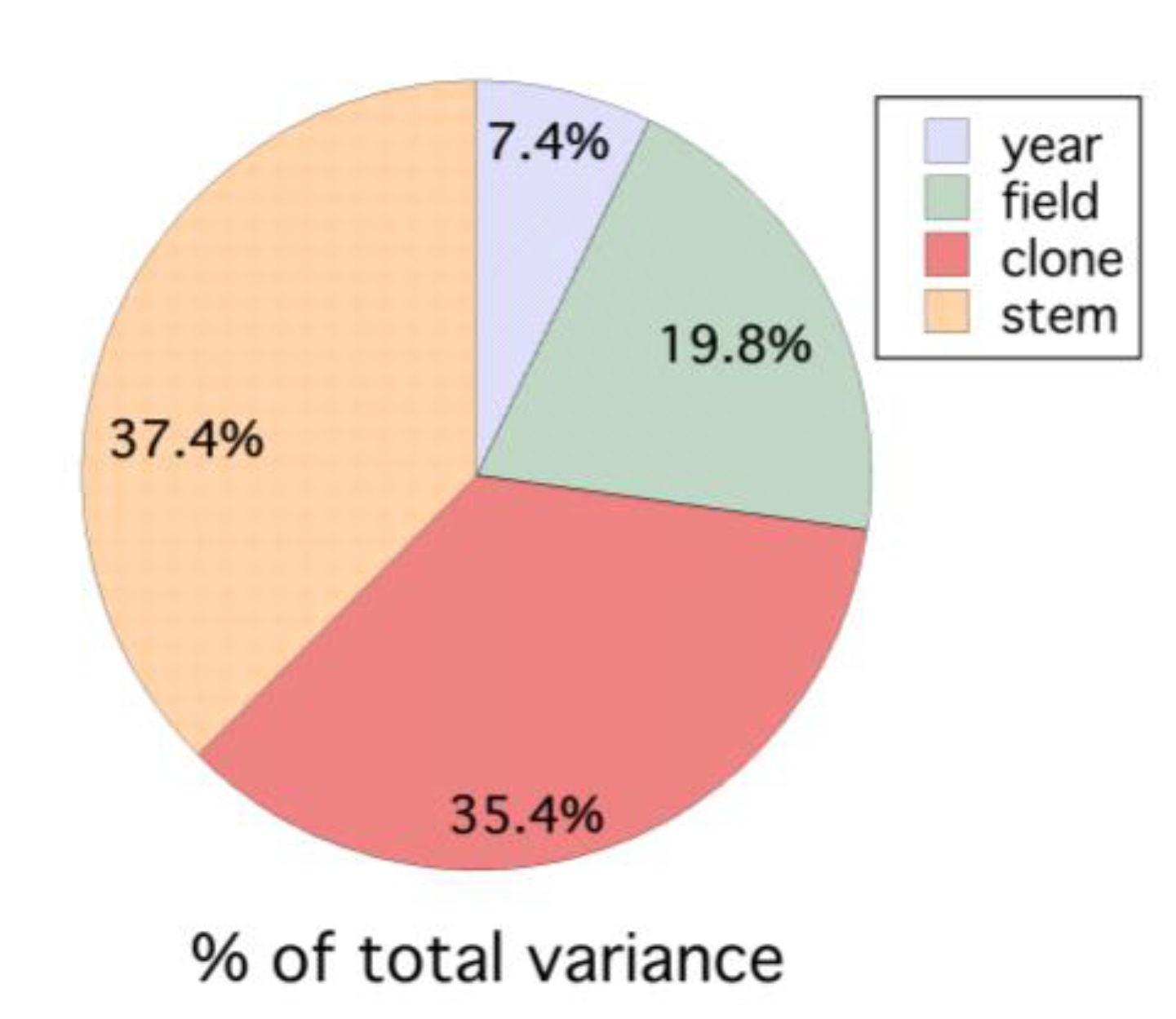
3.4. Within and between Fields and Years (1996–1999)
4. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- Calderwood, L.; Yarborough, D.E.; Tooley, B. Maine Wild Blueberry Production Statistics. Wild Blueberry Fact Sheet. 2019. Available online: https://extension.umaine.edu/blueberries/factsheets/statistics-2/crop-production-statistics-2019/ (accessed on 30 April 2020).
- Jones, M.S.; Vanhanen, H.; Peltola, R.; Drummond, F.A. A global review of arthropod-mediated ecosystem-services in Vaccinium berry agroecosystems. Terr. Arthropod Rev. 2014, 7, 41–78. [Google Scholar] [CrossRef]
- Yarborough, D.E. Wild Blueberry Culture in Maine. Wild Blueberry Fact Sheet. No. 220, Bulletin No. 2088. 2009. Available online: https://extension.umaine.edu/blueberries/factsheets/production/wildblueberryculture-in-maine/ (accessed on 6 May 2020).
- Bell, D.J.; Rowland, L.J.; Smagula, J.; Drummond, F.A. Recent advances in the biology and genetics of lowbush blueberry. Maine Agric. For. Exp. Stn. Tech. Bull. 2009, 203, 1–28. [Google Scholar]
- Eaton, L.J.; Murray, J.E. Relationships of pollinator numbers in blueberry fields to fruit development and yields. Acta Hort. 1996, 446, 181–188. [Google Scholar] [CrossRef]
- Bushmann, S.; Drummond, F.A. A literature review and analysis of pollination services provided by wild and managed bees (Apoidea) in wild blueberry (Vaccinium angustifolium Aiton) production. In Prep.
- Drummond, F.A. Commercial bumblebee pollination of lowbush blueberry. Int. J. Fruit Sci. 2012, 12, 54–64. [Google Scholar] [CrossRef]
- Drummond, F.A. Reproductive biology of wild blueberry (Vaccinium angustifolium Aiton). Agriculture 2019, 9, 69. [Google Scholar] [CrossRef] [Green Version]
- Drummond, F.A. Factors that affect yield in wild blueberry, Vaccinium angustifolium Aiton. Agirc. Res. Technol. Open Access J. 2019, 22. [Google Scholar] [CrossRef]
- Yarborough, D.; Drummond, F.A.; Annis, S.; D’Appollonio, J. Maine Wild blueberry systems analysis. Acta Hortic. 2017, 1180, 151–160. [Google Scholar] [CrossRef]
- Racsko, J.; Nagy, J.; Soltész, M.; Nyeki, J.; Szabo, Z. Fruit drop: I. Specific characteristics and varietal properties of fruit drop. Int. J. Hortic. Sci. 2006, 12, 59–67. [Google Scholar] [CrossRef]
- Bookman, S.S. Costs and benefits of flower abscission and fruit abortion in Asclepias speciosa. Ecology 1983, 64, 264–273. [Google Scholar] [CrossRef]
- Allen, J.C. The effect of citrus rust mite damage on citrus fruit drop. J. Econ. Entomol. 1978, 71, 746–750. [Google Scholar] [CrossRef]
- Smagula, J.M. Effect of boron on lowbush blueberry fruit set and yield. Acta Hortic. 1993, 346, 183–192. [Google Scholar] [CrossRef]
- Milić, B.; Tarlanović, J.; Keserović, Z.; Magazin, N.; Miodragović, M.; Popara, G. Bioregulators can improve fruit size, yield and plant growth of northern highbush blueberry (Vaccinium corymbosum L.). Sci. Hortic. 2018, 235, 214–220. [Google Scholar] [CrossRef]
- Gerbrandt, E.M.; Mouritzen, C.; Sweeney, M. Foliar calcium corrects a deficiency causing green fruit drop in ‘Draper’ highbush blueberry (Vaccinium corymbosum L.). Agriculture 2019, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Koura, S.; Minowa, Y.; Tachikawa, S.; Kato, T.; Taniguchi, M. Study on Control of Fruit Drop of Rabbiteye Blueberry (Vaccinium ashei), 1: The Effect of GA3 and Aminoethoxyvinylglycine on Fruit Setting under Severe Condition. 2000. Available online: https://agris.fao.org/agris-search/search.do?recordID=JP2000003686 (accessed on 6 May 2020).
- Diaz-Lara, A.; Martin, R.R. Blueberry fruit drop-associated virus: A new member of the family Caulimoviridae isolated from blueberry exhibiting fruit-drop symptoms. Plant Dis. 2016, 100, 2211–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racskó, J.; Leite, G.B.; Petri, J.L.; Zhongfu, S.; Wang, Y.; Szabó, Z.; Soltész, M.; Nyéki, J. Fruit drop: The role of inner agents and environmental factors in the drop of flowers and fruits. Int. J. Hortic. Sci. 2007, 13, 13–23. [Google Scholar] [CrossRef]
- Drummond, F.A.; Collins, J. Dispersal from Overwintering Sites, Action Thresholds for Rhagoletis mendax (Diptera: Tephritidae), and Factors that can Influence Variation in Predicted Fruit Infestation Levels in Maine Wild Blueberry. Part I. J. Econ. Entomol. 2020, 113, 851–859. [Google Scholar] [CrossRef]
- Drummond, F.A.; Collins, J.; Ballman, E. Population dynamics of spotted wing drosophila (Drosophila suzukii (Matsumura)) in Maine wild blueberry. Insects 2019, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Penman, L.N.; Annis, S.L. Leaf and flower blight caused by Monilinia vaccinii-corymbosi on lowbush blueberry: Effects on yield and relationship to bud phenology. Phytopathology 2005, 95, 1174–1182. [Google Scholar] [CrossRef] [Green Version]
- Bajcz, A.W.; Drummond, F.A. Flower power: Floral and resource manipulations reveal the consequences of reproductive effort in lowbush blueberry (Vaccinium angustifolium). Ecol. Evol. 2017, 7, 5645–5659. [Google Scholar] [CrossRef] [Green Version]
- Tasnim, R.; Calderwood, L.; Annis, S.; Drummond, F.A.; Zhang, Y.-J. The future of wild blueberries: Testing warming impacts using open-top chambers. Spire 2020. Available online: https://umaine.edu/spire/2020/02/10/ wildblueberries/ (accessed on 17 June 2020).
- Bajcz, A.W.; Drummond, F.A. Bearing fruit: Flower removal reveals the trade-offs associated with high reproductive effort for lowbush blueberry. Oecologia 2017, 185, 13–26. [Google Scholar] [CrossRef]
- Bell, D.J.; Rowland, L.J.; Stommel, J.; Drummond, F.A. Yield variation among clones of lowbush blueberry as a function of kinship and self-compatibility. J. Am. Soc. Hortic. Sci. 2010, 135, 1–12. [Google Scholar]
- Bell, D.J.; Drummond, F.A.; Rowland, J.L. Evidence of functional gender polymorphisms in a population of the hermaphroditic lowbush blueberry (Vaccinium angustifolium Ait.). Botany 2012, 90, 393–399. [Google Scholar] [CrossRef]
- Yarborough, D. Blueberry Enterprise Budget. Wild Blueberry Fact Sheet. No. 260. Bulletin No. 2016. 2011. Available online: https://extension.umaine.edu/blueberries/factsheets/marketing-and-business-manage-ment/260-blueberry-enterprise-budget/ (accessed on 30 April 2020).
- Asare, E.; Hoshide, A.K.; Drummond, F.A.; Chen, X.; Criner, G.K. Economic risk of bee pollination in Maine wild blueberry, Vaccinium angustifolium Aiton. J. Econ. Entomol. 2017, 110, 1980–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS Institute. JMP® Version 14; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.V.; Christensen, R.H.B. Lmertest Package: Tests in linear mixed effects models. J. Stat. Software 2017, 88, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall/Pearson: Upper Saddle River, NJ, USA, 2010; 944p. [Google Scholar]
- Drummond, F.; Collum, K.; Hanes, S.; Wilson, M.; Skinner, J.; Collins, J. A Pollination Toolbox for Wild Blueberry Growers. J. Res. Exten. Workers Conf. 2015. [Google Scholar] [CrossRef]
- Drummond, F.A. Estimating fruit set and pollinator force in wild blueberry. Wild Blueberry Fact Sheet. 2020. Univ. Maine Coop. Ext. Fact Sheet No. 2019. Available online: https://extension.umaine.edu/blueberries/estimating-the-strength-of-your-pollinator-force-in-wild-blueberry/ (accessed on 9 May 2020).
- Du Clos, B.; Loftin, C.; Drummond, F.A. BeeMapper Tool Quick Reference Guide. 2017. Available online: https://umaine.edu/beemapper/wp-content/uploads/sites/353/2017/07/BeeMapper-Quick-Guide-for-Web.pdf (accessed on 5 May 2020).
- Ehrlen, J. Why do plants produce surplus flowers? A reserve-ovary model. Am. Nat. 1991, 138, 918–933. [Google Scholar] [CrossRef]
- Stephenson, A.G. Flower and fruit abortion: Proximate causes and ultimate functions. Ann. Rev. Ecol. Syst. 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Sadras, V.O. Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 2007, 100, 125–138. [Google Scholar] [CrossRef]
- Qu, H.; Drummond, F.A. Simulation-based modeling of wild blueberry pollination. Electron. Comp. Agric. 2018, 144, 94–101. [Google Scholar] [CrossRef]
- Bell, D.J.; Rowland, L.J.; Drummond, F.A. Fine-scale spatial genetic structure associated with Vaccinium angustifolium Aiton (Ericaceae). Int. J. Bot. 2012, 2, 72–82. [Google Scholar]
- Rowland, L.J.; Ogden, E.L.; Bell, D.J.; Drummond, F.A. Pollen-mediated gene flow in managed fields of lowbush blueberry. Can. J. Plant Sci. 2019, 100, 95–102. [Google Scholar] [CrossRef]
- Drummond, F.A. Behavior of bees associated with the wild blueberry agro-ecosystem in the USA. Int. J. Entomol. Nematol. 2016, 2, 27–41. [Google Scholar]
- Quinlan, J.D.; Preston, A.P. Effects of thinning blossom and fruitlets on growth and cropping of Sunset apple. J. Hortic. Sci. 1968, 43, 373–381. [Google Scholar] [CrossRef]
- Willson, M.F.; Price, P.W. The evolution of inflorescence size in Asclepias (Asclepiadaceae). Evolution 1977, 31, 495–511. [Google Scholar] [CrossRef]










© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drummond, F.A. Wild Blueberry Fruit Drop: A Consequence of Seed Set? Agronomy 2020, 10, 939. https://doi.org/10.3390/agronomy10070939
Drummond FA. Wild Blueberry Fruit Drop: A Consequence of Seed Set? Agronomy. 2020; 10(7):939. https://doi.org/10.3390/agronomy10070939
Chicago/Turabian StyleDrummond, Francis A. 2020. "Wild Blueberry Fruit Drop: A Consequence of Seed Set?" Agronomy 10, no. 7: 939. https://doi.org/10.3390/agronomy10070939
APA StyleDrummond, F. A. (2020). Wild Blueberry Fruit Drop: A Consequence of Seed Set? Agronomy, 10(7), 939. https://doi.org/10.3390/agronomy10070939