The Effects of Alkaline Pretreatment on Agricultural Biomasses (Corn Cob and Sweet Sorghum Bagasse) and Their Hydrolysis by a Termite-Derived Enzyme Cocktail



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lignocellulosic Biomass Pretreatment
2.3. Biomass Composition and Scanning Electron Microscope (SEM) Analysis
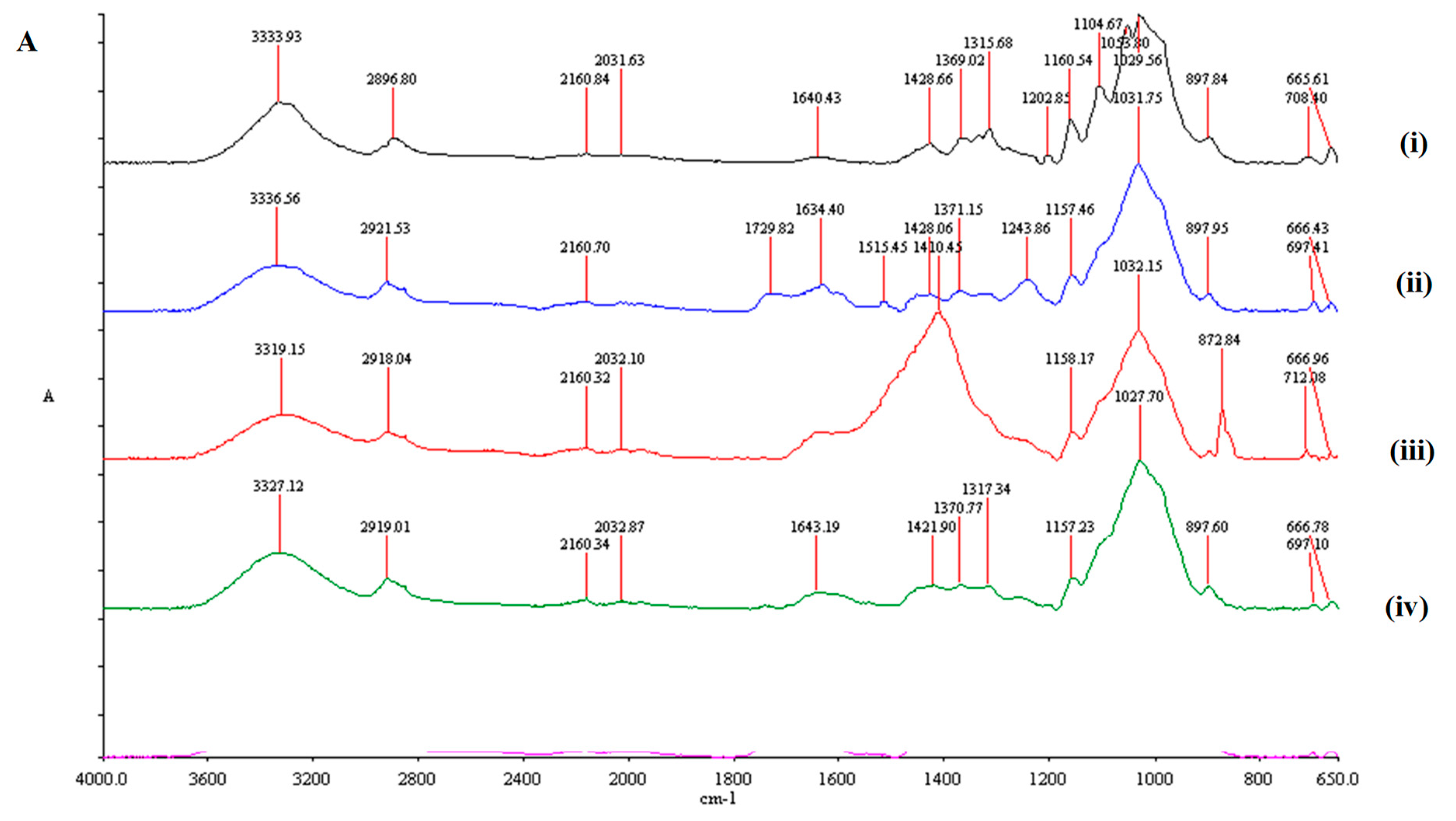
2.4. Fourier-Transform Infrared Spectroscopy Analysis
2.5. Formulation of the Holoenzyme Cocktail (HEC)
2.6. Data Analysis
3. Results
3.1. Biomass Composition and SEM Analysis of Pre-Treated Feedstocks
3.2. FTIR Analysis of Pretreated Feedstock
3.3. Substrate Specificity
3.4. Holocellulolytic Enzyme Cocktail Formulation and Its Application on Pretreated Biomass
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Qi, W.; He, C.; Wang, Q.; Liu, S.; Yu, Q.; Wang, W.; Leksawasdi, N.; Wang, C.; Yuan, Z. Carbon-based solid acid pretreatment in corncob saccharification: Specific xylose production and efficient enzymatic hydrolysis. ACS Sustain. Chem. Eng. 2018, 6, 3640–3648. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K. Sweet sorghum bagasse pretreatment by Coriolus versicolor in mesh tray bioreactor for selective delignification and improved saccharification. Waste Biomass Valorization 2019, 10, 2689–2702. [Google Scholar] [CrossRef]
- Takada, M.; Niu, R.; Minami, E.; Saka, S. Characterization of three tissue fractions in corn (Zea mays) cob. Biomass Bioenergy 2018, 115, 130–135. [Google Scholar] [CrossRef]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jørgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic cross-links: Building and de-constructing the plant cell wall. Nat. Prod. Rep. 2019. [Google Scholar] [CrossRef] [PubMed]
- Malgas, S.; Mafa, M.S.; Mkabayi, L.; Pletschke, B. A mini review of xylanolytic enzymes with regards to their synergistic interactions during hetero-xylan degradation. World J. Microbiol. Biotechnol. 2019, 35. [Google Scholar] [CrossRef] [PubMed]
- Van Dyk, J.S.; Pletschke, B.I. A review of lignocellulose bioconversion using enzymatic hydrolysis and synergistic cooperation between enzymes-factors affecting enzymes, conversion and synergy. Biotechnol. Adv. 2012, 30, 1458–1480. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Deraz, S.; Khalil, A. Bioethanol production from lignocellulosic feedstocks based on enzymatic hydrolysis: Current status and recent developments. Biotechnology 2014, 13, 1–21. [Google Scholar]
- Sharma, H.K.; Xu, C.; Qin, W. Biological Pretreatment of Lignocellulosic Biomass for Biofuels and Bioproducts: An Overview. Waste Biomass Valor 2019, 10, 235–251. [Google Scholar] [CrossRef]
- Dietrich, J.F.R.; Goring, D.A.I. Effect of mercerization on the crystallite size and crystallinity index in cellulose from different sources. Can. J. Chem. 1986, 65, 1724–1725. [Google Scholar]
- Beukes, N.; Pletschke, B.I. Effect of lime pre-treatment on the synergistic hydrolysis of sugarcane bagasse by hemicellulases. Bioresour. Technol. 2010, 101, 4472–4478. [Google Scholar] [CrossRef]
- Oka, D.; Kobayashi, K.; Isobe, N.; Ogawa, Y.; Yokoyama, T.; Kimura, S.; Kim, U.; Tokuyasu, K.; Wada, M. Enzymatic hydrolysis of wood with alkaline treatment. J. Wood Sci. 2013, 59, 484–488. [Google Scholar] [CrossRef]
- Grimaldi, M.P.; Marques, M.P.; Laluce, C.; Cilli, E.M.; Pombeiro Sponchiado, S.R.P. Evaluation of lime and hydrothermal pretreatments for efficient enzymatic hydrolysis of raw sugarcane bagasse. Biotechnol. Biofuels 2015, 8, 1–14. [Google Scholar]
- Kim, J.S.; Lee, Y.Y.; Kim, T.H. A review on alkaline pretreatment technology for bioconversion of lignocellulosic biomass. Bioresour. Technol. 2016, 199, 42–48. [Google Scholar] [CrossRef] [PubMed]
- da Silva Neto, J.M.; Oliveira, C.; da Silva, L.S.H.; Tabosa, F.L.; Pacheco, J.N.; da Silva, M.J.V. Use of sweet sorghum bagasse (Sorghum bicolor (L.) Moench) for cellulose acetate synthesis. BioRes 2019, 14, 3534–3553. [Google Scholar]
- Carrillo, F.; Colom, X.; Sunol, J.J.; Saurina, J. Structural FTIR analysis and thermal characterization of lyocell and viscose-type fibres. Eur. Polym. J. 2004, 40, 2229–2234. [Google Scholar] [CrossRef]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef]
- Ni., J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef]
- Bastien, G.; Arnal, G.; Bozonnet, S.; Laguerre, S.; Ferreira, F.; Fauré, R.; Henrissat, B.; Lefèvre, F.; Robe, P.; Bouchez, O.; et al. Mining for hemicellulases in the fungus-growing termite Pseudacanthotermes militaris using functional metagenomics. Biotechnol. Biofuels 2013, 6, 1–15. [Google Scholar]
- Rashamuse, K.; Ronneburg, T.; Sanyika, W.; Mathiba, K.; Mmutlane, E.; Brady, D. Metagenomic mining of feruloyl esterases from termite enteric flora. Appl. Microbiol. Biotechnol. 2013, 98, 727–737. [Google Scholar] [CrossRef]
- Rashamuse, K.; Sanyika, T.W.; Mathiba, K.; Ngcobo, T.; Mtimka, S.; Brady, D. Metagenomic mining of glycoside hydrolases from the hindgut bacterial symbionts of a termite., Trinervitermise trinervoides and the characterisation of a multimodular β-1, 4-Xylanase (GH11). Biotechnol. Appl. Biochem. 2016, 64. [Google Scholar] [CrossRef]
- Panagiotopoulos, I.A.; Bakker, R.R.; De Vrije, T.; Koukios, E.G.; Claassen, P.A.M. Pretreatment of sweet sorghum bagasse for hydrogen production by Caldicellulosiruptor saccharolyticus. Int. J. Hydrogen. Energy 2010, 35, 7738–7747. [Google Scholar] [CrossRef]
- Sluiter, J.B.; Ruiz, R.O.; Scarlata, C.J.; Sluiter, A.D.; Templeton, D.W. Compositional analysis of lignocellulosic feedstocks. 1. Review and description of methods. J. Agric. Food Chem. 2010, 58, 9043–9053. [Google Scholar] [CrossRef] [PubMed]
- Mafa, M.S.; Dirr, H.W.; Malgas, S.; Krause, R.W.M.; Rashamuse, K.; Pletschke, B.I. A novel dimeric exoglucanase (GH5_38): Biochemical and structural characterisation towards its application in alkyl cellobioside synthesis. Molecules 2020, 25, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, L.G. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Álvarez, C.; Reyes-Sosa, F.M.; Díez, B. Enzymatic hydrolysis of biomass from wood. Microb. Biotechnol. 2016, 9, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Liu, J.; Chang, X.; Xue, Y.; Liu, P.; Lin, H.; Han, S. A review on the pretreatment of lignocellulose for high-value chemicals. Fuel Proc. Technol. 2017, 160, 196–206. [Google Scholar] [CrossRef]
- Kumar, A.; Negi, Y.S.; Choudhary, V.; Bhardwaj, N.K. Characterization of cellulose nanocrystals produced by acid-hydrolysis from sugarcane bagasse as agro-waste. J. Mater. Phys. Chem. 2014, 2, 1–8. [Google Scholar]
- Fry, S.C.; York, W.S.; Albersheim, P.; Darvill, A.; Hayashi, T.; Joseleau, J.-P.; Kato, Y.; Lorences, P.E.; Maclachlan, G.A.; McNeil, M.; et al. An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol. Plant. 1993, 89, 1–3. [Google Scholar] [CrossRef]
- Rashmi, R.; Siddalingamurthy, K.R. Microbial xyloglucanases: A comprehensive review. Biocatal. Biotransform. 2018, 36, 280–295. [Google Scholar] [CrossRef]
- Cheng, X.-Y.; Huang, W.-J.; Hu, S.-C.; Zhang, H.-L.; Wang, H.; Zhang, J.-X.; Lin, H.-H.; Zou, Y.-Z.Q.; Ji, Z.-L. Global Characterization and Identification of Multifunctional Enzymes. PLoS ONE 2012, 7, e38979. [Google Scholar] [CrossRef] [Green Version]
- Huberts, D.H.E.W.; van der Klei, I.J. Moonlighting proteins: An intriguing mode of multitasking. Biochim. Biophys. Acta 2010, 1803, 520–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, J.C. Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 2003, 19, 415–417. [Google Scholar] [CrossRef]
- Aspeborg, H.; Coutinho, P.M.; Wang, Y.; Brumer, H.; Henrissat, B. Evolution, substrate specificity and subfamily classification of glycoside hydrolase family 5 (GH5). BMC Evol. Biol. 2012, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Liu, H.; Xu, Q.; Sun, J.; Shi, H. Identification and characterization of two endogenous β-glucosidases from the termite Coptotermes formosanus. Appl. Biochem. Biotechnol. 2015, 176, 2039–2052. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.; Kovaleva, E.S.; Slack, J.M.; Brown, S.; Buchman, G.W.; Scharf, M.E. A GHF7 cellulase from the protist symbiont community of Reticulitermes flavipes enables more efficient lignocellulose processing. Arch. Insect Biochem. Physiol. 2013, 84, 175–193. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass Pre-Treatment | Biomass Composition (%) | ||
|---|---|---|---|
| Corncob (CC) | Lignin | Cellulose | Hemicellulose |
| Untreated | 22.51 ± 0.19 | 23.58 ± 0.47 | 33.34 ± 0.56 |
| Ca(OH)2 | 14.81 ± 0.10 # | 27.34 ± 0.33 # | 33.49 ± 0.11 |
| NaOH | 9.50 ± 0.04 # | 33.32 ± 0.41 # | 35.03 ± 0.63 * |
| Sweet sorghum bagasse (SSB) | |||
| Untreated | 29.34 ± 0.042 | 17.75 ± 0.36 | 16.28 ± 0.46 |
| Ca(OH)2 | 29.08 ± 0.092 | 19.60 ± 0.1 # | 11.54 ± 0.47 * |
| NaOH | 10.88 ± 0.031 # | 23.81 ± 0.22 # | 13.05 ± 0.25 # |
| Substrates | Enzyme Specific Activity (U/mg Protein) | ||
|---|---|---|---|
| MFE-5E | MFE-5H | MFE-45 | |
| CMC | 17.189 ± 0.049 | 28.584 ± 0.025 | 4.75 ± 0.053 |
| Avicel | N/A | N/A | N/A |
| Beechwood xylan | 7.483 ± 0.013 | 52.015 ± 0.071 | 4.55 ± 0.013 |
| Wheat flour xylan | 13.471 ± 0.161 | 67.365 ± 0.116 | # |
| Xyloglucan | 6.831 ± 0.008 | 84.975 ± 0.012 | 12.86 ± 0.19 |
| Locus bean gum | N/A | N/A | N/A |
| pNP-A | N/A | N/A | NA |
| pNP-G | N/A | N/A | N/A |
| pNP-X | N/A | N/A | N/A |
| pNP-C | N/A | N/A | N/A |
| pNP-M | N/A | N/A | N/A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mafa, M.S.; Malgas, S.; Bhattacharya, A.; Rashamuse, K.; Pletschke, B.I. The Effects of Alkaline Pretreatment on Agricultural Biomasses (Corn Cob and Sweet Sorghum Bagasse) and Their Hydrolysis by a Termite-Derived Enzyme Cocktail. Agronomy 2020, 10, 1211. https://doi.org/10.3390/agronomy10081211
Mafa MS, Malgas S, Bhattacharya A, Rashamuse K, Pletschke BI. The Effects of Alkaline Pretreatment on Agricultural Biomasses (Corn Cob and Sweet Sorghum Bagasse) and Their Hydrolysis by a Termite-Derived Enzyme Cocktail. Agronomy. 2020; 10(8):1211. https://doi.org/10.3390/agronomy10081211
Chicago/Turabian StyleMafa, Mpho. S., Samkelo Malgas, Abhishek Bhattacharya, Konanani Rashamuse, and Brett I. Pletschke. 2020. "The Effects of Alkaline Pretreatment on Agricultural Biomasses (Corn Cob and Sweet Sorghum Bagasse) and Their Hydrolysis by a Termite-Derived Enzyme Cocktail" Agronomy 10, no. 8: 1211. https://doi.org/10.3390/agronomy10081211
APA StyleMafa, M. S., Malgas, S., Bhattacharya, A., Rashamuse, K., & Pletschke, B. I. (2020). The Effects of Alkaline Pretreatment on Agricultural Biomasses (Corn Cob and Sweet Sorghum Bagasse) and Their Hydrolysis by a Termite-Derived Enzyme Cocktail. Agronomy, 10(8), 1211. https://doi.org/10.3390/agronomy10081211