1. Introduction
Cowpea (
Vigna unguiculata L. Walp.) is an important food legume throughout West and Central Africa. It occupies a smaller proportion of the crop area compared to cereals and contributes significantly to household food security in West and Central Africa [
1]. Globally, the total estimated area of cowpea production is about 14 million ha. Out of this, West Africa accounts for almost 9.3 million ha, deriving an annual production of 2.9 million Mg. Cowpea makes a valuable contribution towards human food and livestock fodder and its dual-purpose character makes it a very attractive crop where land is becoming scarce [
2]. It has a unique ability to fix atmospheric nitrogen through its nodules and grows well even in poor soils with more than 85% sand and less than 0.2% organic matter and low levels of phosphorus [
3]. Additionally, it is shade tolerant and, therefore, compatible as an inter-crop with maize, millet, sorghum, sugarcane and cotton, as well as with several plantation crops [
4]. Coupled with these characteristics, its quick growth and rapid ground cover make it a suitable plant to prevent soil erosion. As relatively inexpensive sources of high-quality protein [
5], cowpeas are important in the nutrition of the poor. The economic and nutritional importance of cowpea in West Africa makes it a key subject for research.
Cowpea is a drought-tolerant and tropical crop, which is well adapted to the semi-arid regions of the tropics where other food legumes do not perform well [
2]. Despite its ability to thrive under various drought conditions which would otherwise render other field crops unproductive [
6], there have been various reports demonstrating significant drought tolerance existing among cowpea genotypes [
7,
8]. Tolerance to drought in cowpea has been attributed to increased photosynthetic response to drought stress and is highly complex. It involves the interplay of processes taking place at different sites of the cell or leaf and at different time scales concerning plant development [
9]. The reduction in photosynthetic activity is due to several coordinated events, such as stomatal closure and the reduced activity of photosynthetic enzyme depth [
10], while greater nutrient acquisition has been associated with topsoil foraging, especially in the case of immobile resources such as phosphorus [
11]. Mid- and late-season drought stresses have received considerable attention, given their negative effects on yield [
12,
13,
14]. Low levels of soil available N and P and large crop responses to N and P fertilizer applications are common for both cereals and legumes in the moist savanna of West Africa [
15]. The P fertilizer use efficiency (grain yield per unit of nutrient supplied) [
16] has been established to a certain extent in cowpea [
17]. Root symbiosis with Arbuscular Mycorrhizal fungi (AMF) has also been shown to enhance P absorption by increasing the effective root area [
18].
Water and phosphorus can represent extremely contrasting resource availability in time and space, where soil water is ephemeral, mobile and in dry periods only in the subsoil, whereas soil phosphorus is stable, immobile and usually concentrated in the topsoil. Studies have indicated that moisture content in soil has a strong effect on P uptake by plants [
19]. While the single effects of irrigation or P application on the grain yield, grain yield components and P-related parameters of cowpea have been reported in numerous studies, not much has been published about the interactive effects of these factors, especially on highly weathered, acidic and low-P soils in Africa. The objective of this study is to identify cowpea genotypes that are tolerant to both P and drought stress on a highly weathered tropical soil with strong P fixation capacity. We hypothesize that (1) genotypes that have the highest grain yield under optimum conditions do not perform best under water or P stress conditions and (2) genotypes that have the highest grain yield under P stress conditions (2.1) perform well under water stress conditions and (2.2) also perform well under combined water and P stress.
2. Materials and Methods
Two field experiments were conducted at the Crops Research Institute (CRI), Fumesua, in the Ashanti Region of Ghana in the dry season of 2017 and 2018, respectively. The mean daily temperature during the experimental period in the two years of this study was 30.4 °C and 32.0 °C with a relative humidity of 79.9% and 83.3%, respectively. Total rainfall during the growing cycle was 143 and 119 mm in 2017 and 2018, respectively. Physical and chemical soil analyses were conducted on soil samples from four different soil depths before the beginning of the field experiments. The soil texture of both experimental sites was sandy loam in the topsoil and clay in the subsoil (>60 cm;
Table 1). Water-holding capacity of the soil layers is expressed as water content at field capacity (0.33 bar). Ammonium acetate, aluminum (titration method) and Bray 1 methods were used to extract exchangeable bases, exchangeable acids and available P, respectively (
Table 2). The results of the physical analyses of the soils for experiments 1 and 2 are presented in
Table 1 and
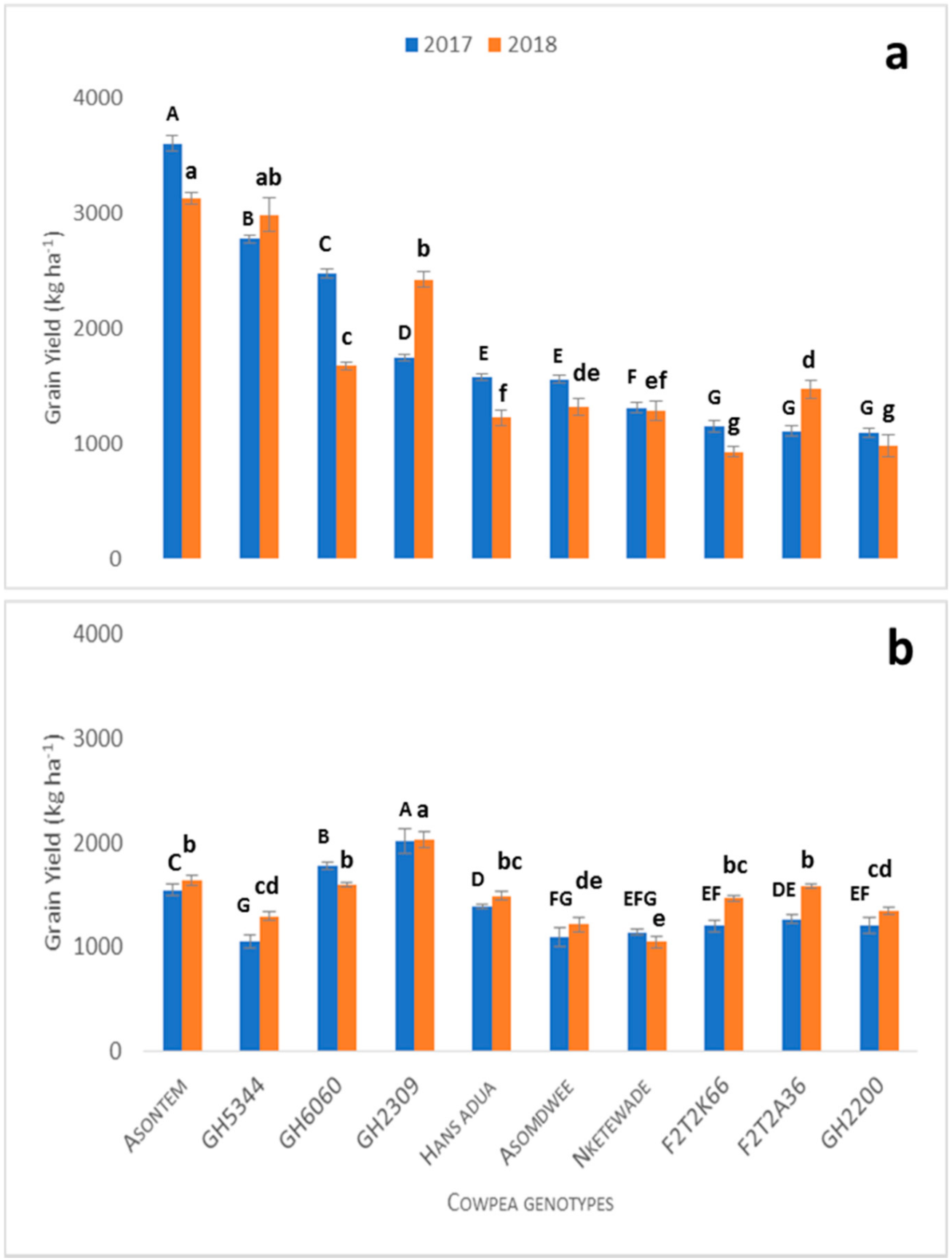
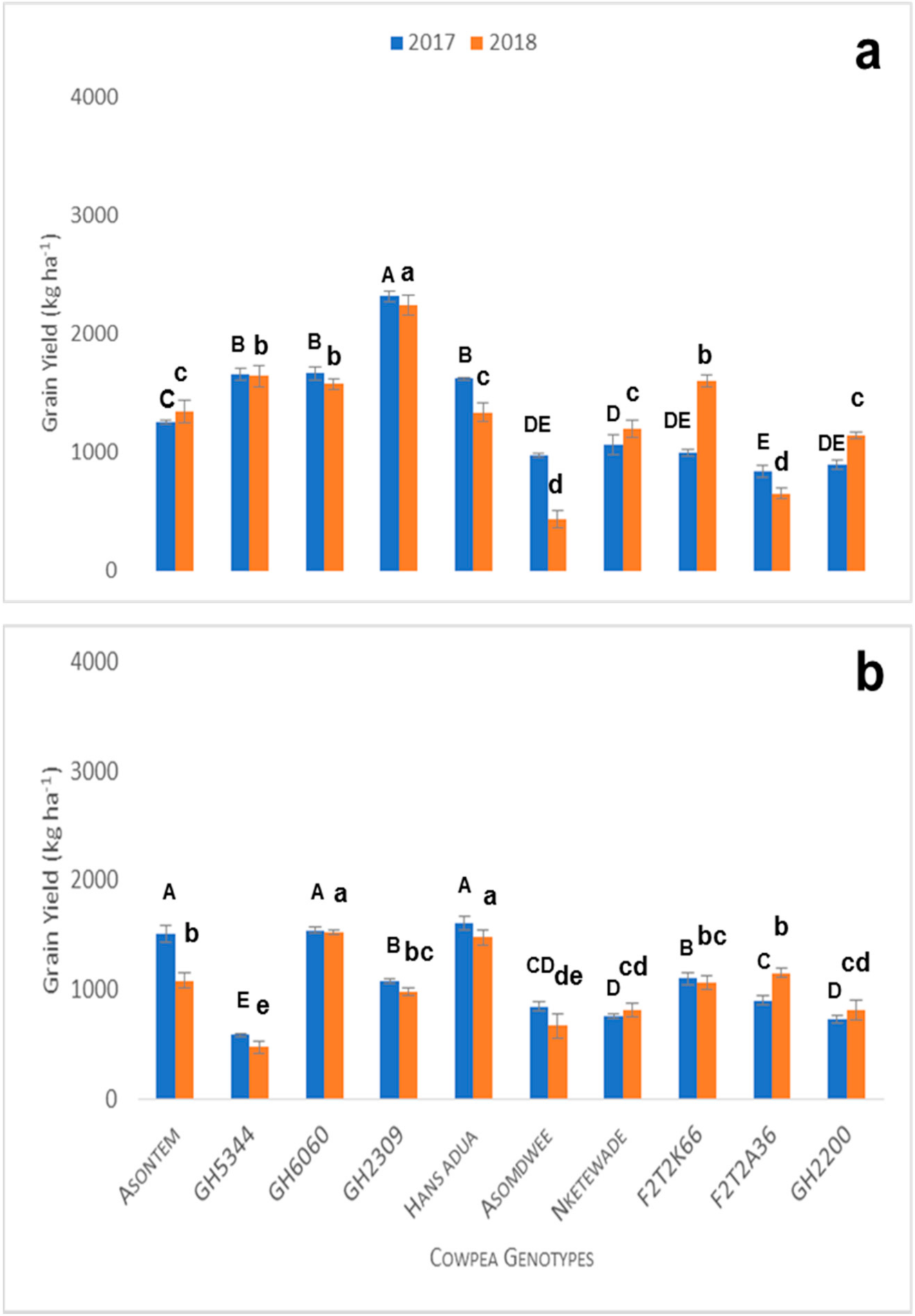
Table 2. Ten (10) cowpea lines were selected based on screening trials in pots in the greenhouse, including the best five genotypes (Asontem, GH-2200, GH-2309, F2T2K66 and GH-6060), that were tolerant to P and water stress and the worst five (Asomdwee, GH-5344, Hans adua, Nketewade and F2T2A36) genotypes that were susceptible to P and water stress. The selection criterion was aboveground biomass productivity.
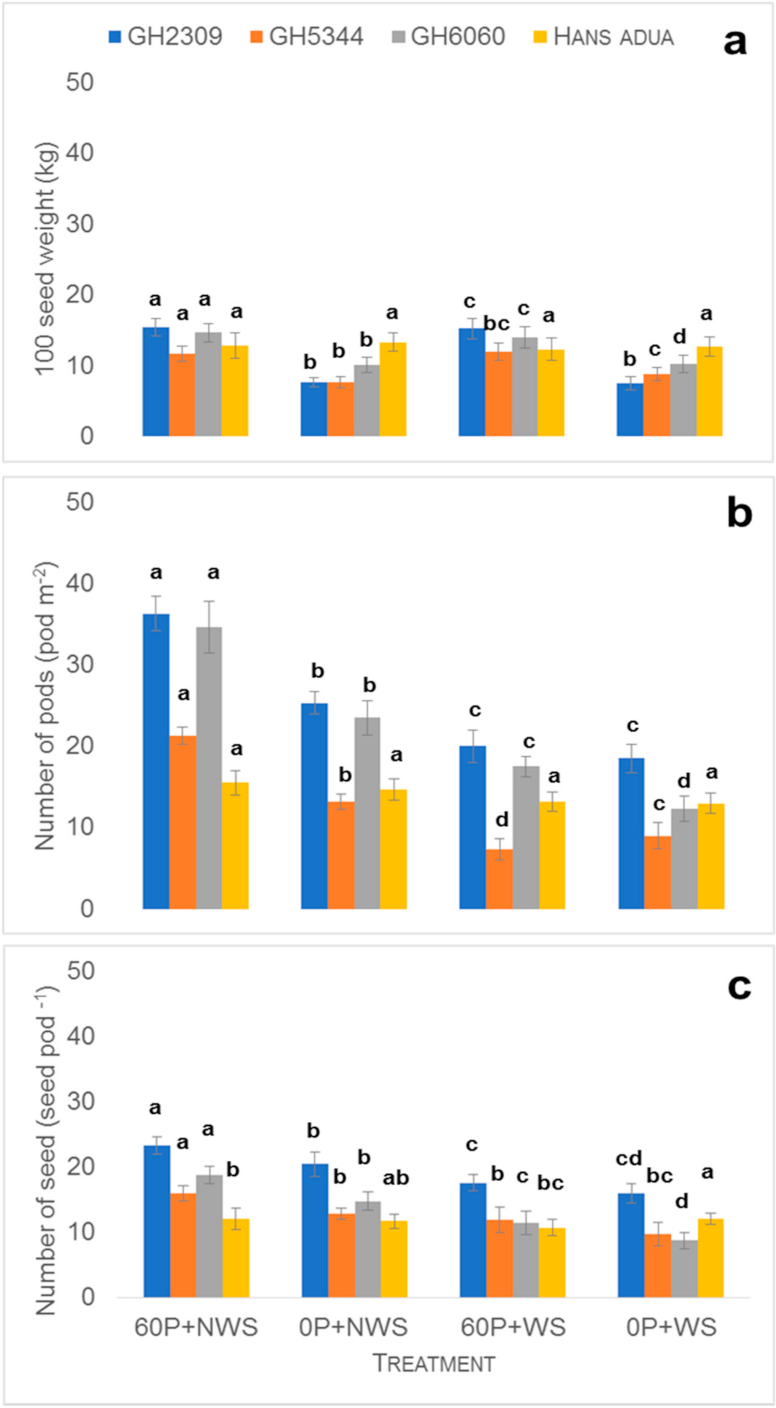
Both field experiments were conducted in a split plot design with 3 factors (P fertilizer application levels, water regimes and cowpea genotypes) with three replicates. The main plot comprised the 10 selected cowpea genotypes. The subplot was P fertilizer application (triple superphosphate) with two levels (0 kg Pha−1 and 60 kg Pha−1) with water treatment as a sub-subplot level (with water stress and without water stress). A total of four combinations of P fertilizer and water treatment were established in the field experiment (sub- and sub-subplots): 0 kg Pha−1 + water stress (0P + WS, control treatment); 0 kg Pha−1 + no water stress (0P + NWS); 60 kg Pha−1 + water stress (60P + WS); 60 kg Pha−1 + no water stress (60P + NWS, optimum condition) for the field experiments in both years.
All cowpea seeds were surface sterilized with moncerene at 5 mL of chemical to 15 liters of water. The seeds were dipped in the chemical solution for 30 s and allowed to dry for 15−20 min. Seeds for the first and second experiments were sown on 23rd December 2017 and 29th December 2018, respectively. Four seeds of each variety were sown per hill and thinned to one plant per hill 10 days after emergence. Both experiments were conducted within a land area of 1354.4 m2 with a 0.60 m distance between rows and 0.40 m between hills. The area occupied by each plant was 0.24 m2. Blocks and plots in both experiments were separated by a spacing of 1 m. In both experiments, there were 7 rows in each plot and each row was 1.8 m.
In all treatments, plants were sufficiently irrigated twice daily (morning and evening) to avoid any occurrence of water stress until 21 days after planting to ensure uniform germination and good seedling establishment. Then, in the water stress treatment (WS), irrigation was interrupted at vegetative and flowering stages of growth, i.e., no water was applied for 2 weeks. At the vegetative stage, stress was imposed at 21 days after planting for 2 weeks and at the flowering stage after the emergence of flower buds for 2 weeks (
Table 3). A line source irrigation system with impact sprinklers was used to irrigate the amount of water stated in
Table 3. All field observations and plant samples were obtained from the central five rows of each plot. At maturity, the five rows were harvested for grain yield. Outer rows were not included during the sampling processes in order to avoid border effects on the measurements.
A mixture of Karate
® 2.5 E.C. (a.i. 25 g lambda-cyhalothrin (Hangzhou Tianlong Biotechnology Co., Ltd., Hangzhou, China) per liter; 4 mL L
−1 Vertimec (Syngenta Agro Maintal, Am Technologiepark 1-5 ·63477- Maintal, Germany), 1.8%w v
−1 Abamectin (King Tech Corporation, Shenzhen, chiana) (18 g L
−1; 1.5 mL in 1 L of water) [
3] was sprayed twice (third and fifth weeks) to control insects in the vegetative phase. Then Sunpyrifos
® (5.3 mL in 1 L of water) was sprayed twice (before and after flowering) to control insect pests and diseases, especially white flies (
Trialeurodes vaporariorum) and bacterial blight disease (
Pseudomonas savastanoi), which were very prominent during the experimental period. The bacterial blight disease infestation in 2018 was more severe than in 2017, as well as white flies. Roundup (Monsanto Agrar Deutschland GmbH, Düsseldorf, Germany) and Gramoquat were used to control weeds.
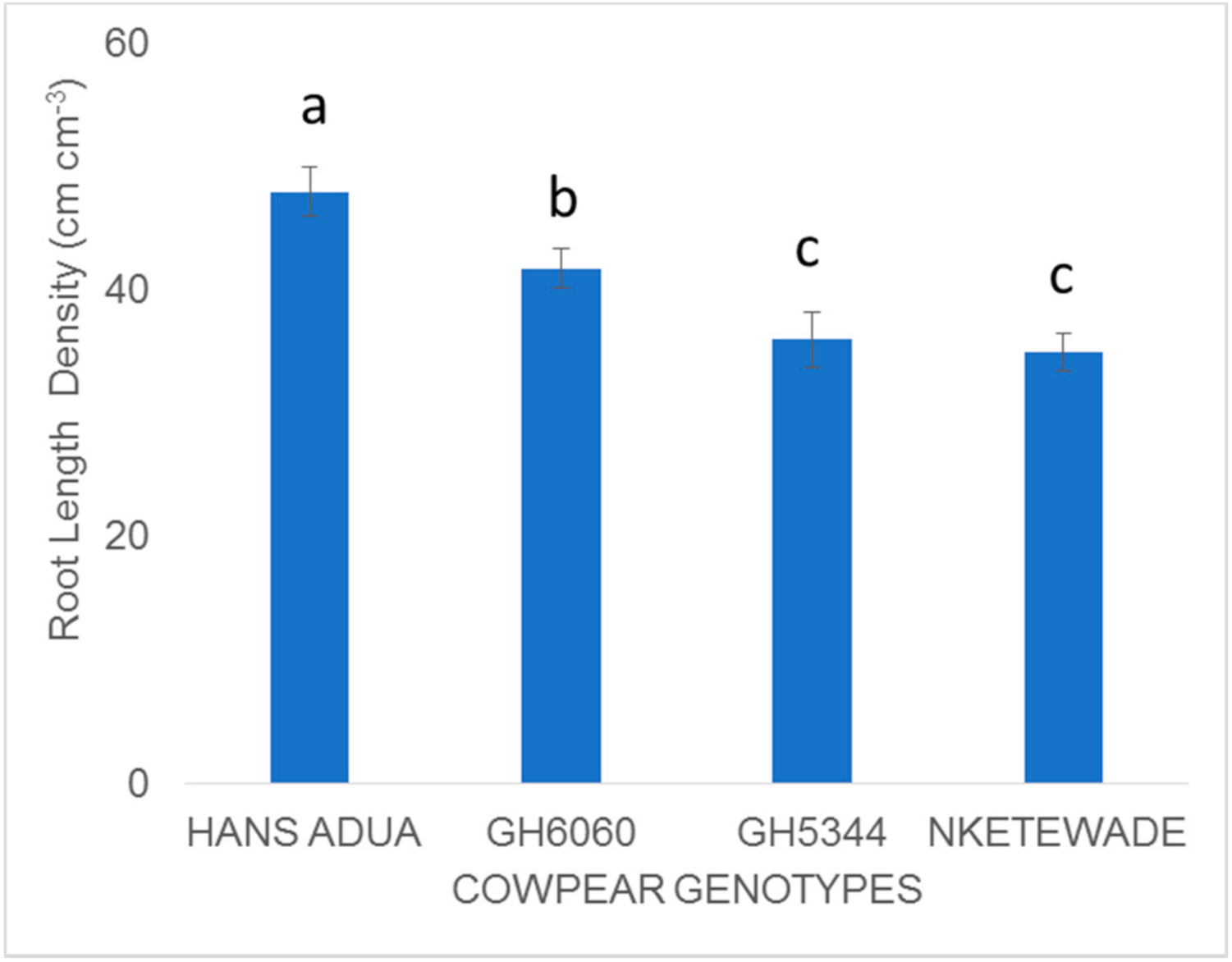
Harvesting was carried out manually in both experiments. Harvesting of plant samples was carried out 60 days after planting onwards depending on the physiological maturity of the different genotypes. Plants were sampled in the five central rows in an area of 1.2 m2 to determine the final biomass and grain yield. After each treatment (water stress application) during the vegetative stage, 3 plants were harvested from each plot and oven-dried at 65 °C for 72 h. The samples were weighed to determine dry shoot biomass. The greenness of leaves was a proxy for chlorophyll content and was measured using a Soil Plant Analysis Development (SPAD ) 502 Plus Chlorophyll Meter (Spectrum Technologies, 3600 Thayer Ct, IL, USA). Three plants in each plot of each replicate from genotypes Hans adua, GH6060, GH5344 and Nketewade roots were excavated to a soil depth of 90 cm together with the soil at the stage of pod initiation around 40 days after planting and gently washed to determine root length density using WINRHIZO PRO 2012 Software (Regent Instruments Inc, Ville de Québec, QC, Canada) to understand how these genotypes responded to the four treatments (0 kg Pha−1 + NWS; 0 kg Pha−1 + WS; 60 kg Pha−1 + NWS; 60 kg Pha−1 + WS). Yield data and root length density obtained from both experiments were analyzed using GenStat (17th edition, VSN INTERNATIONAL LIMITED, 5 The Waterhouse Street, Hemel Hempstead, United Kingdom). A linear model procedure using the restricted maximum likelihood method was performed for ANOVA. Treatment means were separated by the least significant differences (LSD) test at p < 0.05.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}